行政院國家科學委員會專題研究計畫 成果報告
HO-1/hDAF/HLA-DR 多基因轉殖豬細胞體外之功能測試(2/2)
計畫類別: 整合型計畫 計畫編號: NSC92-2314-B-002-226- 執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日 執行單位: 國立臺灣大學醫學院外科 計畫主持人: 李章銘 計畫參與人員: 李玉汶 報告類型: 完整報告 處理方式: 本計畫可公開查詢中 華 民 國 93 年 11 月 9 日
行政院國家科學委員會補助專題研究計畫
異種器官供人體移植之可行性研究-使用多基因轉殖小鼠與豬器官
- HO-1/hDAF/HLA-DR 多基因轉殖豬細胞體外之功能測試
計畫類別: 個別型計畫 □整合型計畫
計畫編號:NSC
92-2314-B-002-226
執行期間: 九十一 年 八 月 一 日至 九十三 年 七 月 三十一 日
計畫主持人:李章銘醫師
共同主持人:
計畫參與人員:李玉汶小姐
成果報告類型(依經費核定清單規定繳交):□精簡報告 完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列
管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查詢
執行單位:臺大醫院外科
中 華 民 國 九十三 年 十一 月 八 日
成果報告
□期中進度報告
附件一中文摘要: 關鍵詞:基因轉殖豬,人類白血球抗原,人類延遲加速因子,第一型 Heme 氧化酵素 以豬器官來源的異種器官,為目前臨床移植所面臨的器官短缺的問題,提供了解決之道。 但是其可行性,目前仍受限於人類對豬隻的異種抗體所誘發的超急性,及亞急性異種器官排 斥,及物種間組織抗原歧異所引起的 T 細胞排斥。先前在我們所產製的 HLA DPW0401 基因轉殖 豬中,發現人類 HLA DP 的抗原性,可完整表達在豬細胞表面,而相較於其他非基因轉殖同胞 豬,HLA DPW0401 基因轉殖豬所引起人類周邊單核球(具 HLA DPW0401 基因型者)的增生反應, 有減少的趨勢。 HO-1 是代謝 Heme 的主要酵素,而能保護細胞抵抗異種抗體所引發的細胞裂解及凋亡,及 使異種移植器官在異種抗體存在之下,達到”適存化”的狀態。而人類延遲加速因子(hDAF, CD55)為一補體活化鏈的調節者。目前發現 hDAF 的基因轉殖豬器官,包括心臟,腎臟及肝臟 可在靈長類體內, 存活 7 天至 3 個月。
在過去一年中,我們已經成功地產製了 HLA-DR 及 HO-1 之基因轉殖豬,其 mRNA 及其蛋白 的表達都已得到確定,針對 HO-1 保護作用的測試,我們已經成功地完成自然殺手細胞毒性測 試的檢驗模式,我們發現殺手細胞與標的細胞於 5:1 的情形有最明顯的毒殺現象,之後我們 將運用於 HO-1 轉殖豬細胞的測試,檢測此 HO-1 轉殖基因是否有保護作用。我們也完成了 HLA-DR 基因轉殖豬細胞對人類週邊單核球的增生測試,我們發覺這種基因轉殖豬細胞,與非 基因轉殖同胞豬細胞相較,其引起的異種細胞免疫反應,並沒有明顯的差別,至於這些轉殖豬 細胞,是否對於其他的異種免疫反應有保護作用,則有待進一步確定。 Abstract:
Keywords: Transgenic Pig , Xenotransplantation, HLA, HO-1, hDAF
Pig to human xenotransplatation is a promising strategy to overcome the organ shortage in clinical transplantation. However, the application is hampered by the xenoreactive antibody mediated hyperacute rejection, delayed xenograft rejection and MHC disparity associated T cell rejection. Previous, we have produced the HLA DPW0401 transgenic pig, which was shown equipped with the human HLA antigenecity and, as compared to the non-transgenic litermate pig, induced a minor cellular response to the human PBMCs that came from the HLA DPW 0401(+) human. HO-1 (heme oxygenase-1) is the main enzyme metabolizing the heme, which has been demonstrated to offer a protective effect for the xenograft to achieve accommodation under the presence of xenoreactive antibodies, and help to attenuate xenoantibody mediated celll lysis and apoptosis. To test the protective effect of HO-1 exogene from the NK related cytotoxicity, we have establish the optimal condition for this cytotoxic assay using the NK cell line, K562. The cell killing efficacy will be maximal when the effector/target ratio being 5: 1, 47% of cell death of the total cell population. We have also successfully cultured the aortic endothelial cells of pig (PAEC) from the non-transgenic pig. The culture of the PAEC of the transgenic pig is ongoing now. Because the HLA-DR 1501 has been successfully produced, we also test the xenogenic MLC for the HLA DR transgenic pig. We found that the tendency about xenogenic cellular response was not evidently different between the HLADR transgenic and non-transgenic littermate control pis PBMC. Whether this transgenic pig cell can obtain protective effect in another process of xenogenic immune response required to be examined further.
Background
Xenotransplantation with graft organ from pig is a promising strategy to overcome the shortage of organ, which has been the main restriction for clinical transplantation (1,2). The xenoreactive antibody-mediated hyperacute rejection (HAR) and delayed xenograft rejection (DXR) (or acute vascular rejection) are the major obstacles to impede the survival of xenofgraft organ in human (3-7). Various strategies have been developed to suppress the HAR or DXR successfully, including using organs from transgenic pigs of human Decay Accelerating Factor (8,9), depletion, or suppression of production of xenoreactive antibodies (10-14). The cellular response induced by differences of MHC between disconcordant species can provide a continuous detrimental effect to graft organ, even though the xenograft has gone through the stages of HAR or acute vascular rejection. It has been demonstrated that the porcine MHC molecules can effectively induce a strong human T cell response, through direct or indirect antigen recognition (15-17). The human NK cell associated anti-porcine cytotoxicity was also maintained, which was T cell and antibody-independent (18).
Heme oxygenase (HO) is responsible for the metabolism of hemin and can be detected in all mammalian tissues. At least three isoenzymes, i.e., HO-1, HO-2, and HO-3, were included in the HO family (20-2). HO-1 is a stress-inducible protein which can be up-regulated by stimulation by cytokines, heavy metals, hypoxia, or oxygen-free radical (23-5). HO-2 is a constitutively expressed protein, particularly aboundant in brain, testis, or liver of mammals (26). The role of HO-3 is relatively obscured, which is believed to be a regulator rather than a direct catalyst in the heme processing (22). HO-1 can catalyze the conversion of heme to biliverdin, Iron, and carbon monooxide (27). Under usual circumstance, which is rich in biliverdine reductase, the biliverdin can be rapidly transformed into bilirubin, the more familiar form. The CO can help to maintain the vascular tone in small and median sized arteries, or aorta (28-9). Bilirubin is an antioxidant (30) and may be important in maintaining the viability of cells under stress environment. Although the iron is toxic to the cells by creating detrimental reactive oxygen species, the iron can be neutralized by ferritin right away after its production (27). Ferritin is a strong antioxidant that protects the cells from oxidative stress. Its expression can in turn be up-regulated by iron through binding to its mRNA iron-responsive elements (31-2). Blocking the activity of HO-1 by tin mesoporphyrin can deteriorate renal function of rat kidney after ischemia and induce an extensive injury in the renal tubule (33). Up-regulation of HO-1 by hemin can also ameliorate the halothane-induced hepatotoxicity (34). Recently it was found that the expression of HO-1 in the endothelium of genograft could essential for the xenograft accommodation. The mouse heart xenotransplant can survived long-term in the rats receiving CVF and CyA. However, the heart from a HO-1 deficient mouse was eventually rejected three to seven days after transplantation in rats under the similar treatment (35). Based on these observations, we expect that through expression of human HO-1 on the porcine cells might also contribute to the survival porcine xenograft.
Materials and Methods
Establishing the NK cell cytotoxicity assay .
This assay was based on a dual fluorescent staining of target cells. The dye, DIOC18 (3,3’-dioctadecyloxacarbocyanine perchlorate) is a carbocyanine membrane dye that label
cellmembrane by inserting two lond (C18 carbon) hydrocarbonchains into the lipid bilayers, is used to stain the membrane of the target cells. Propidium Iodide (PI) is used to label dead cells. The K562 cell line was used as the effector cell to test the protective effect of HO-1 transgene for the NK associated cytotoxicity (Gong et al., 1994). The procedure was listed as below
1. Wash the tatget cells in PBS and re-suspend in PBS at a concentration of 1X 106 cell/ml. 2. Stain the target cell with DIOC 18 10 ul per 1X 106 cells
3. Incubation of the cells for 20 min at 37oC, 5% CO2
4. Wash the cells twice with buffer solution.and resuspend in complete culture media at a concentration of 1X 106 cells/ml
5. Prepare the target and effector cells and made a co-culture of them with the desire ratio, i.e. E:T: 1:5; 1:2.5; 1:1; 1:0.5; 1:0.2. Incubate the co-culture for 4 hours.
6. Centrifuge the cell mixture at 250 g for 5 min after wash, discard suernatant. 7. Washed and re-suspend the cultured cells
8. Label the cells with Propidium iodide (5 ul/per test) incubate at room temperature in dark. 9. Analysis with flowcytometry.
Culture of porcine endothelial cells(PAEC):
1) Treat the porcine aorta with collagenase (2.5 mg/ml; Sigma).
2) Seeding of PAECs in 25cm2 gelatin-coated culture flasks in RPMI. 1640 medium (Gibco,
Paisleg UK), supplemented with 10% heat-
inactivated FCS(Life Technology), 25mM sodium bicarbonate(Flow, Les Ulis, France), 2mM glutamine (Flow), 1 mM sodium pyruvate (Flow), 60 µg/ml tylocin(Life Technologies), 50 IU/ml penicillin (Flow), and 50 µg/ml streptomycin (Flow) (referred
as culture medium)
3) Subculture the PAEC with treatment of trypsin-EDTA(Life Technologies)-with the use of 2nd to 8th subculture
4) Incubate the PAEC with recombinant human TNF-α (Genzyme , Boston, MA) (100U/ml fo36 hours), the optimal condition for inducing MHC expression in previous report.(24)
Mixed lymphocyte culture:
1. Dilution the heparinized human blood with double volume of HBSS 2. Overlayer the above mentioned mixture with 10ml Ficoll-paque 3. Centrifuge with 1500 rpm for 30 minutes
4. Harvest the mononuclear cell layer in the interface 5. Wash with HBSS for three times (PBMC)
6. Incubate the PBMC for 4 hours at 37o C in 75-cm2 plastic flask (Falcon #3023, Becton Dickinson Labware, Lincoln Park, NJ) to deplete the adherent cells.
7. Recover the adherent cells with rubber policemen and analysis with flowcytometry
8. Incubate non-adherent cells for 60 min at 37o C on nylon wool (Fenwal Laboratories, Deerfield, IL) and then gently elute enrich the T cells
9. Treat the T cells with 1:100 dilution of the ascitic fluid of a murine anti-porcine macrophage- and granulocyte-specific antibody for at 4oC for 30 min.
10. Add rabbit complement 9HLA-ABC, Pel Freeze, Roger, AR) at 1:8 dilution and incubate for 45 min at 37oC
11. Wash the cells with culture media
12. Prepare the responder and stimulator cells a) responder cells adjust the
concentration to 1x106 cells/ml
b) stimulator cell:(PBMC) adjust the concentration to 2x106
cells/ml
c) Irradiate the stimulator cells with 4500 cGy and incubate in 37
o
C, 5% CO2 for 20 minutes.
d)Wash the stimulator cells with HBSS for three times e)Adjust the concentration of stimulator cells to 1x106 cells/ml
13. Add into each well with 100 µl of stimulator and responder cells respectively and done in triplicate
14. Incubate in 37o C and 5% CO2 for 6 days 15. Add 20 µl H3 thymidine incubating for 6 hours. 16. Harvest with glass fiber and dry in air
17. Add scintillating cocktail 18. Count the cpm with β-counter.
Cellular Proliferative Assay (Xenogenic MLC)
Responders: PBMCs from the persons with the same genotype of the HLAII exogene expressed on
the transgenic pig cells.
Stimulators ( porcine PBMCs, porcine aortic endothelial cells (PAEC), and porcine Islet cells):
a. Cells from non-transgenic littermate pig b. Cells from DP transgenic pig
c. Cells from DR transgenic pig d. Cells from DP + DR transgenic pig
Results and discussion:
The establishment of NK cytotoxicity assay
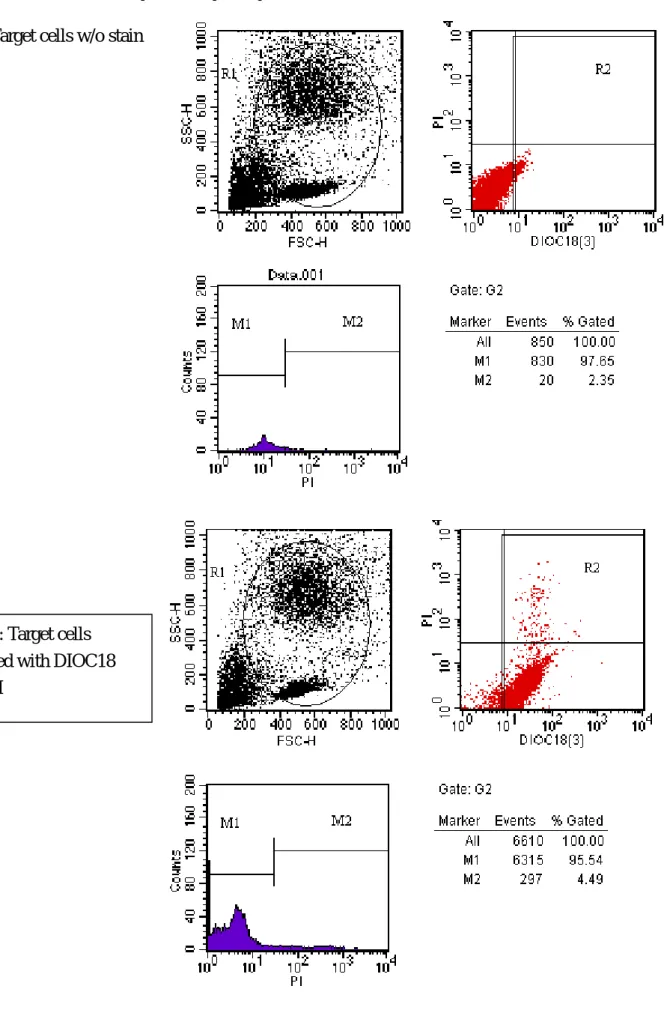
Using the NK cell line, K 526, we will be able to evalaue the protective effect of the HO-1 exogene from the NK related cytotoxicity. We have already established the optimal condition to assay the
cytoxicity. The effector/ target ratio of 5/1 will produce a maximal efficacy of NK cytotoxicity with 46% of the cells stained with propidium iodide. About 27% of cell killing was found when the effector / target ratio being 1 / 1 or 2 / 1. The efficacy was not satisfactory when the target cell was more than the effector cells. This will provide a useful tool to evaluate the HO-1 exogene effect. The detail of the results were demonstrated in Fig 1-7. Next step we will test the system using the swine PBMC, PAEC and islet cell as the target cells.
Culture of the PAEC
At the present time, we have susceccfully produced the HLA DR 1501 transgenic pig. The PAEC of the non-transgenic pig has been harvested and cultured successfully. The culture PAEC of the HLA DR transgenic pig was ongoing at the present time.
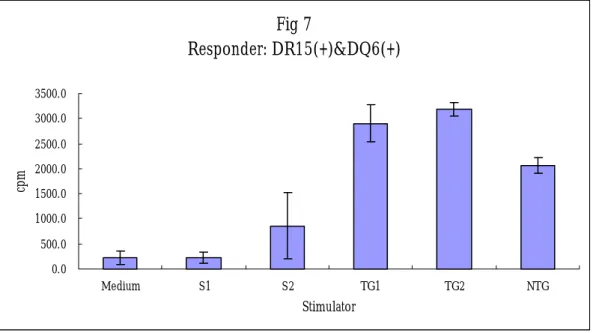
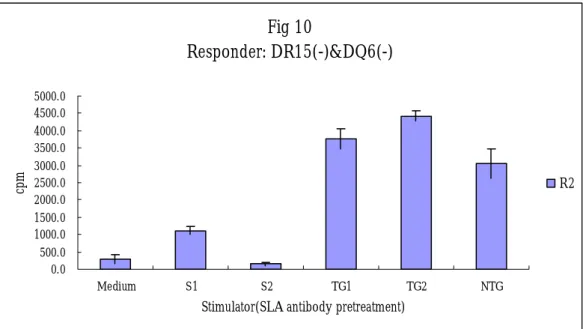
Xenogenic MLC for the HLA DR transgenic pig.
Xenogenic MLC with coculture of the human PBMC and the transgenic and non-transgenic swine PBMC was performed. After seven days of culture, the CPM of the human responder cells did not produced differences of cellular proliferation to the stimulator cells from the non-transgenic or transgenic pig. The results are demonstrated in the figures of 8 and 9.
References
1. Sachs DH, Leight G, Cone J, Schwarz S, Stuart L, Rosenberg S. Transplantation in miniature swine. I. Fixation of the major histocompatibility complex. Transplantation 1976; 22: 559.
2. Sachs DH. The pig as a potential xenograft donor. Vet Immunol Immunopathol 1994; 43: 185. 3. Alwayn IP, Basker M, Buhler L, Cooper DK The problem of anti-pig antibodies in pig-to-primate xenografting: current and novel methods of depletion and/or suppression of production of anti-pig antibodies. Xenotransplantation 6:157-68, 1999.
4.Cozzi E, Masroor S, Soin B, Vial C, White DJ. Progress in xenotransplantation. Clin Nephrol 2000; 53: 13.
5. Galili U, Rachmilewitz EA, Peleg A, Flechner I. A unique natural human IgG antibody with anti-alpha-galactosyl specificity. J Exp Med 1984; 160:1519.
6. Cooper DK, Good AH, Koren E, Oriol R, Malcolm AJ, Ippolito RM, Neethling FA, Ye Y, Romano E, Zuhdi N. Identification of alpha-galactosyl and other carbohydrate epitopes that are bound by human anti-pig antibodies: relevance to discordant xenografting in man. Transpl Immunol 1993; 1: 198.
7. Bach FH, Soares M, Lin Y, Ferran C. Barriers to xenotransplantation. Transplant Proc 1999; 31: 1819.
8. Cozzi E, Bhatti F, Schmoeckel M, Chavez G, Smith KG, Zaidi A,
Bradley JR, Thiru S, Goddard M, Vial C, Ostlie D, Wallwork J, White DJ, Friend PJ. Long-term survival of nonhuman primates receiving life- supporting transgenic porcine kidney xenografts. Transplantation 2000; 70: 15.
9. Vial CM, Ostlie DJ, Bhatti FN, Cozzi E, Goddard M, Chavez GP, Wallwork J, White DJ, Dunning JJ. Life supporting function for over one month of a transgenic porcine heart in a baboon. J Heart Lung Transplant 2000; 19:224.
10. Alexandre GPJ, Gianello P, Latinne D, et al. Plasmapheresis and splenectomy in experimental renal xenotransplantation. In : Hardy MA, ed. Xenograft New York:, Elsevier, 1989: 25.
11. Kozlowski T, Ierino FL, Lambrigts D, Foley A, Andrews D, Awwad M, Monroy R, Cosimi AB, Cooper DK, Sachs DH. Depletion of anti-Gal(alpha)1-3Gal antibody in baboons by specific alpha-Gal immunoaffinity columns. Xenotransplantation 1998; 5: 122.
Monoclonal antiidiotypic antibodies neutralize cytotoxic effects of anti-alphaGal antibodies. Transplantation 1996; 62: 837.
13. Magee JC, Collins BH, Harland RC, Lindman BJ. Immunoglobulin prevents complement-mediated hyperacute rejection in swine-to-primate xenotransplantation. J Clin Invest 1995; 96: 2404.
14. Kozlowski T, Shimizu A, Lambrigts D, Yamada K, Fuchimoto Y, Glaser R, Monroy R, Xu Y, Awwad itzer TR, Cooper DK, Sachs DH. Porcine kidney and heart transplantation in baboons undergoing a tolerance induction regimen and antibody adsorption. Transplantation 1999; 67:18. 15. Yamada K, Sachs DH, DerSimonian H. Human anti-porcine xenogeneic T cell response. Evidence for allelic specificity of mixed leukocyte reaction and for both direct and indirect pathways of recognition. J Immunol 1995; 155: 5249.
16. Dorling A, Lombardi G, Binns R, Lechler RI. Detection of primary direct and indirect human anti-porcine T cell responses using a porcine dendritic cell population. Eur J Immunol 1996; 26:1378.
17. Rollins SA, Kennedy SP, Chodera AJ, Elliott EA, Zavoico GB, Matis LA. Evidence that activation of human T cells by porcine endothelium involves direct recognition of porcine SLA and costimulation by porcine ligands for LFA-1 and CD2. Transplantation 1994; 57: 1709.
18. Kirk AD, Li RA, Kinch MS, Abernethy KA, Doyle C, Bollinger RR. The human antiporcine cellular repertoire. In vitro studies of acquired and innate cellular responsiveness. Transplantation 1993; 55: 924.
20. Shibahara S, Muller R, Taguchi H, Yoshida T. 1985. Cloning and expression of cDNA for rat heme oxygenase. Proc Natl Acad Sci U S A 82:7865-7869
21. Maines MD, Trakshel GM, Kutty RK. 1986. Characterization of two constitutive forms of rat liver microsomal heme oxygenase. Only one molecular species of the enzyme is inducible.
J Biol Chem 261:411-419.
22. McCoubrey WK Jr, Huang TJ, Maines MD. 1997. Isolation and characterization of a cDNA from the rat brain that encodes hemoprotein heme oxygenase-3. Eur J Biochem 247:725-732
23. Tacchini L, Schiaffonati L, Pappalardo C, Gatti S, Bernelli-Zazzera A. 1993. Expression of HSP 70, immediate-early response and heme oxygenase genes in ischemic-reperfused rat liver. Lab
24. Choi AM, Alam J. 1996. Heme oxygenase-1: function, regulation, and implication of a novel stress-inducible protein in oxidant-induced lung injury. Am J Respir Cell Mol Biol 15:9-19
25. Motterlini R, Foresti R, Bassi R, Calabrese V, Clark JE, Green CJ. 2000. Endothelial heme oxygenase-1 induction by hypoxia. Modulation by inducible nitric-oxide synthase and S-nitrosothiols. J Biol Chem 275:13613-13620
26. Maines MD. 1988. Heme oxygenase: function, multiplicity, regulatory mechanisms, and clinical applications. : FASEB J 2):2557-2568
27. Galbraith R. 1999. Heme oxygenase: who needs it? Proc Soc Exp Biol Med 222:299-305
28. Kozma F, Johnson RA, Zhang F, Yu C, Tong X, Nasjletti A. 1999. Contribution of endogenous carbon monoxide to regulation of diameter in resistance vessels. Am J Physiol 276:R1087-94
29. Sammut IA, Foresti R, Clark JE, Exon DJ, Vesely MJ, Sarathchandra P, Green CJ, Motterlini R. 1998. Carbon monoxide is a major contributor to the regulation of vascular tone in aortas expressing high levels of haeme oxygenase-1. Br J Pharmacol 125:1437-1444
30. Stocker R, Yamamoto Y, McDonagh AF, Glazer AN, Ames BN. 1987. Bilirubin is an antioxidant of possible physiological importance. : Science 235:1043-1046
31. Thomson AM, Rogers JT, Leedman PJ. 1999. Iron-regulatory proteins, iron-responsive elements and ferritin mRNA translation. Int J Biochem Cell Biol 31:1139-1152
32. Pantopoulos K, Hentze MW. 1998. Activation of iron regulatory protein-1 by oxidative stress in vitro. Proc Natl Acad Sci U S A 95:10559-10563
33. Shimizu H, Takahashi T, Suzuki T, Yamasaki A, Fujiwara T, Odaka Y, Hirakawa M, Fujita H, Akagi R. 2000. Protective effect of heme oxygenase induction in ischemic acute renal failure. Crit
Care Med 28:809-817
34. Odaka Y, Takahashi T, Yamasaki A, Suzuki T, Fujiwara T, Yamada T, Hirakawa M, Fujita H, Ohmori E, Akagi R. 2000. Prevention of halothane-induced hepatotoxicity by hemin pretreatment: protective role of heme oxygenase-1 induction. : Biochem Pharmacol 59:871-880
35. Soares MP, Lin Y, Anrather J, Csizmadia E, Takigami K, Sato K, Grey ST, Colvin RB, Choi AM, Poss KD, Bach FH. 1998. Expression of heme oxygenase-1 can determine cardiac xenograft survival.
37 .Cozzi E. Bhatti F. Schmoeckel M. Chavez G. Smith KG. Zaidi A. Bradley JR. Thiru S. Goddard M. Vial C. Ostlie D. Wallwork J. White DJ. Friend PJ. Long-term survival of nonhuman primates receiving life-supporting transgenic porcine kidney xenografts. Transplantation. 70(1):15-21, 2000;
38.Vial CM. Ostlie DJ. Bhatti FN. Cozzi E. Goddard M. Chavez GP. Wallwork J. White DJ. Dunning JJ. Life supporting function for over one month of a transgenic porcine heart in a baboon.
Journal of Heart & Lung Transplantation. 19(2):224-9, 2000;
39. Ramirez P, Chavez R, Majado M, Munitiz V, Munoz A, Hernandez Q, Palenciano CG, Pino-Chavez G, Loba M, Minguela A, Yelamos J, Gago MR, Vizcaino AS, Asensi H, Cayuela MG, Segura B, Marin F, Rubio A, Fuente T, Robles R, Bueno FS, Sansano T, Acosta F, Rodriguez JM, Navarro F, Cabezuelo J, Cozzi E, White DJ, Calne RY, Parrilla P Life-supporting human complement regulator decay accelerating factor transgenic pig liver xenograft maintains the metabolic function and coagulation in the nonhuman primate for up to 8 days. Transplantation 70: 989-98, 2000.
40. Gong, J. H., Maki, G. & Klingemann, H. G. (1994). Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 8, 652-8.
Cytotoxicity assay of the NK cell line, K562 Fig1: Target cells w/o stain
Fig 2: Target cells stained with DIOC18 (3)/PI
Fig 3: E:T 1:5
Fig 5: E:T 1: 1
Fig 8: Xenogenic MLC for the HLA-DRw15 transgenic pig using PBMC from HLADRw15 (+) human (representive of one of thre experiments).
Fig 7 Responder: DR15(+)&DQ6(+) 0.0 500.0 1000.0 1500.0 2000.0 2500.0 3000.0 3500.0 Medium S1 S2 TG1 TG2 NTG Stimulator cp m Fig 7: E:T 1: 0.2
Fig 8: Xenogenic MLC for the HLA-DRw15 transgenic pig using PBMC from HLADRw15 (+) human (representive of one of thre experiments).
Fig 10 Responder: DR15(-)&DQ6(-) 0.0 500.0 1000.0 1500.0 2000.0 2500.0 3000.0 3500.0 4000.0 4500.0 5000.0 Medium S1 S2 TG1 TG2 NTG
Stimulator(SLA antibody pretreatment)
cp