(Title page)
Gestational ingestion of oxidized frying oil by C57BL/6J mice differentially affects the susceptibility of the male and female offspring to diet-induced obesity in adulthood1,2,3
Hui-Ching Chuang4, Chin-Fang Huang4, Yi-Chun Chang4, Yu-Shun Lin4, Pei-Min Chao4*
4 Institute of Nutrition, China Medical University, Taichung, Taiwan
1 Financial support for this study was provided by the National Science Council of Taiwan [grant number NSC98-2320-B039-039-MY3].
2 Author disclosure: H.C. Chuang, C.F. Huang, Y.C. Chang, Y.S. Lin, and P.M. Chao have no conflicts of interest.
3Supplemental Tables 1-2 and Supplemental Fig 1 are available as Online Supporting Material with the online posting of this paper at http://jn.nutrition.org
* To whom correspondence should be addressed, e-mail: [email protected]
Keywords:
Oxidized frying oil ; PPAR ; Gestation ; Obesity ; Offspring
Running title: OXIDIZED FRYING OIL AND OFFSPRING OBESITY Word count: 5371; Number of figures: 2; Number of tables: 2
Supplementary material: Online Supporting Materials: 3 Author list for indexing: Chuang, Huang, Chang, Lin, Chao 5Abbreviations used:
BAT, brown adipose tissue; DEHP, di-(2-ethylhexyl) phthalate; DIO, diet-induced obesity; FGF21, fibroblast growth factor 21; OFO, oxidized frying oil; OFO-f, female offspring of dams fed the oxidized frying oil-diet; OFO-m, male offspring of dams fed the oxidized frying oil-diet; PP, peroxisome proliferator; SO, fresh soybean oil; SO-f, female offspring of dams fed the fresh soybean oil-diet; SO-m, male offspring of dams fed the fresh soybean oil-diet; WAT, white adipose tissue.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33
Abstract
The aim of this study was to investigate whether maternal ingestion of oxidized frying oil (OFO) during pregnancy influences the susceptibility to diet-induced obesity (DIO) of the adult offspring. Pregnant C57BL/6J mice were fed either a control diet (10% fresh soybean oil; SO) or an OFO-containing diet (10 % OFO) throughout the entire gestational period. After parturition, all pups were lactated by SO-fed dams for 3 wk, weaned onto a non-purified standard diet for 4 wk, and shifted to a high-fat diet (29% butter + 1% SO) for 5 wk. Consequently, four groups of offspring were obtained, consisting of the male (m) or female (f) offspring of dams fed the OFO diet (OFO-m and OFO-f) or the SO diet (SO-m and SO-f). At pregnancy d18, higher levels (P<0.05) of mRNA for PPAR target genes were found in the liver of the OFO-fed dams and their fetuses than in their SO controls. Although all pups were raised under the same conditions in postnatal life, a comparison of pups from dams on the different diets based on gender showed that adult OFO-f mice were prone to DIO, while adult OFO-m mice were resistant. The adult OFO-m mice also showed higher expression of PPAR target genes in the liver and white adipose tissue (WAT) and of thermogenic genes in the WAT than adult SO-m mice, whereas no such difference was seen between adult OFO-f and SO-f mice. We conclude that uterine PPAR activation caused by maternal OFO ingestion affects hepatic PPAR activity and adipose thermogenic capacity and contributes to the differential susceptibility to DIO in the male and female offspring in adulthood.
34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55
Introduction
Although frequent consumption of fried food, inevitably accompanied by high fat intake, is causally related with obesity, cardiovascular disease and cancer, we and others have shown that, both in in vitro and in vivo, oxidized frying oil (OFO)5 is more potent than fresh oil in activating PPAR (1-4). Accordingly, when rodents fed diets with equal amounts of OFO or fresh oil are compared, lower plasma and liver lipid concentrations (1, 5, 6), less adiposity (7, 8), and lower pro-inflammatory responses in the vasculature (9) are observed in the OFO-fed group. PPAR is a transcription factor that plays a pivotal role in regulating lipid homeostasis (10, 11). The target genes regulated by PPAR are mainly a homogeneous group participating in lipid catabolism, lipoprotein transport and clearance, and thermogenesis (11). The PPAR activators in OFO were shown to be oxidized fatty acids, such as hydroxyl and hydroperoxy fatty acids (12-14), which can be absorbed from the intestine and incorporated into lipoproteins for circulation (12, 13).
According to Barker’s hypothesis (15), intrauterine nutrition determines propensity to the later onset of metabolic diseases. Studies in experimental animals in which intrauterine nutrition was manipulated by subjecting the pregnant mothers to an energy-restricted or protein-deficient diet, dexamethasone injection or intrauterine artery ligation to produce intrauterine growth retarded pups have shown that the pups are predisposed to be obese or to suffer from metabolic disorders in later life (16-19). Such effects appear to be gender specific, although no consistent pattern is apparent. Notably, this fetal programming effect can be amplified by exposure to a hypercaloric diet during adulthood, which accelerates the etiology of adult-onset disease (16). The postulated underlying mechanisms for fetal programming are mainly focused on hyperphagia/sedentary behavior and the epigenetic regulation of genes associated with 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80
energy homeostasis, which have been shown to be determined by the prenatal environment (20, 21).
The effect of OFO ingestion during pregnancy on the susceptibility to diet-induced obesity (DIO) of the adult offspring has never been studied. It has been suggested that PPAR activators in OFO can be transferred from the maternal circulation to the fetus across the placenta, as prominent peroxisome proliferator (PP) responses including upregulation of PPAR target genes expression and a lower TG concentration in the liver, are observed in the fetuses of OFO-fed mothers (4). The reduced TG availability caused by gestational OFO consumption might restrict the growth of the fetus. In addition, it is not known whether PPAR activation during early life permanently affects hepatic lipid metabolism and/or plasticity of adipose development. Recently, hepatic fibroblast growth factor 21 (FGF21) was found to be involved in the browning of white adipose tissue (WAT) and proposed as a thermogenic hormone (22-24). Interestingly, Fgf21 has been identified as a PPAR target gene (25). It has been reported that feeding thermoxidized oil to female rats throughout rearing, pregnancy and lactation results in pups with a lower birth weight, but a greater body fat mass at weaning (26). In the present study, pregnant mice were subjected to an OFO or fresh soybean oil (SO) diet during the gestational period and the susceptibility of the male and female adult offspring to DIO and the expression of genes participating in lipid catabolism and thermogenesis in the liver and WAT of the offspring were investigated.
81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101
Materials and Methods
Preparation of oxidized frying oil
OFO was prepared by frying dough sheets in soybean oil (President, Tainan, Taiwan) at 205 5℃ for four 6-h periods as described previously (1-3). The extent of oxidation was evaluated by the acid value (OFO, 2.841; SO, 0.056 KOH mg/g) and conjugated diene levels (OFO, 3560; SO, 504 OD233 units/g of oil), values very similar to those obtained in our previous studies (1-3,7,8).
Animals and diets
Female and male C57BL/6JNarl mice purchased from the National Laboratory Animal Center of the National Applied Research Laboratories, Taipei, Taiwan were used for breeding. Females with a parity of at least 1 and not more than 4 were used in this study. Pregnant females were fed either the SO or the OFO diet from conception until delivery. Pregnancy was dated by the presence of a vaginal plug (assigned as d1). After spontaneous delivery (d19.5), litters were cross-fostered to lactating dams fed the SO diet using a litter size of 7 for 3 wk, then were weaned onto a non-purified standard diet (Altromin 1320) for 4 wk, then switched to a high-fat diet for 5 wk. Thus, four groups of offspring were obtained, i.e. SO-m, OFO-m, SO-f, and OFO-f, named according to the maternal diet (SO or OFO) and the gender of the offspring (m or f). All mice were kept in a room maintained at 23 ± 2℃, with a controlled 12 h light:dark cycle with free access to food and drinking water. Body weight and food intake were recorded weekly. The protocols for animal care and handling were approved by the Institutional Animal Care and Use Committee (IACUC) of the China Medical University. The 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126
compositions of the test diets used in this study are shown in Supplemental Table 1.
Tissue sampling and biochemical analysis
The offspring in the four groups (SO-m, OFO-m, SO-f, and OFO-f) were killed at 12 wk of age by carbon dioxide asphyxiation. The retroperitoneal, inguinal, epididymal, ovarian peripheral fat, and interscapular brown adipose (BAT) were excised and weighed. Aliquots of the liver and inguinal fat were stored at -80℃ for RNA extraction. Randox Laboratory commercial kits for cholesterol (CHOD-PAP), TG (GPO-PAP), or non-esterified fatty acids (NEFA) were used to measure serum and liver lipids. Liver lipids were extracted as described previously (1). TBARS levels in the liver homogenate were measured using the method of Oteiza et al (27). The leptin concentration in the serum was assayed using commercial kits (mouse leptin ELISA, Millipore).
At pregnancy d18 [embryonic d18 (E18)], four dams fed the OFO diet and four fed the SO diet were killed and the number and body weight of the fetuses recorded. The livers of the mothers and their fetuses were stored at -80℃ for RNA extraction.
Gender identification of the fetus
For accurate sexing of the fetuses, the presence of the Sry and DXNds3 genes was examined by PCR. The Sry gene is present in a particular sex-determining region of the Y chromosome, while DXNds3 as a polymorphic microstatellite locus located on mouse X chromosome (28). Genomic DNA was extracted from the fetal liver using FavorPrepTM Tissue genomic DNA extraction mini kits (Favorgen Biotech). The sequences of the primers used were 5’-GCCATGTCAAGCGCCCCATGAA-3’ (forward) and GTGGGGATATCAACAGGCTGCCA-3’(reverse) for Sry and 5’-127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151
ATGCTTGGCCAGTGTACATAG-3’ (forward) and 5’-TCCGGAAAGCAGCCATTGGAGA-3’ (reverse) for DXNds3.
RNA isolation and mRNA detection
Total RNA was extracted from homogenized tissue using TRIZOL reagent (Invitrogen) according to the manufacturer’s instructions. Total RNA (1 g) was reverse-transcribed into first-strand cDNA using 200 units of MMLV-RT (Promega) in a total volume of 20 l. For real-time PCR, a TaqMan® system with inventory primers and probes (Applied Biosystems) or an SYBR system with self-designed primers was used. The assay ID of the inventory primers and probes and the sequence of the self-designed primers are shown in Supplemental Table 2. Amplification using forty cycles of two steps (95℃ for 15 s and 60℃ for 1 min) was performed on an ABI Prism 7900HT sequence detection system.
Statistical analysis
Data are expressed as the mean SD. To test the significance of the effects of maternal diet (SO vs. OFO), sex (male vs. female) and their interaction, data from the four groups at the same time point were analyzed by two-way ANOVA. When a significant interaction (P<0.05) was found between maternal diet and sex, the significance of differences among the four groups was further analyzed statistically by Duncan’s multiple range test. The significance of differences between SO- and OFO-fed dams at the same time point was analyzed using Student’s t test. The significance of differences in the survival rate of pups from SO- and OFO-fed dams was analyzed using the Chi-squared test. If the variances were not homogeneous, the data were transformed to log values for the statistical analysis. The SAS System (SAS institute, 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173 174 175 176
Cary, NC) was employed for the statistical analysis and differences were considered significant at P<0.05.
177
Results
Body weight and energy intake of, and liver TBARS levels in, the dams and offspring
Throughout the gestational period, there was no difference between the SO- and OFO-fed dams in body weights at the same time point (Supplemental Fig. 1A) or energy intake (data not shown). Using liver TBARS as an index of peroxidation, the values for OFO-fed dams (14.2 ± 3.5 nmol/g) was significantly higher than those for SO-fed dams (8.9 ± 2.1 nmol/g). Although there was no difference in fetus number per litter (7.7±2.4), the birth weight of the pups of the OFO-fed dams (0.88 ± 0.13 g) was significantly lower than that of the pups of the SO-fed dams (0.93 ± 0.13 g). The fetuses from the OFO-fed dams had a 40% lower survival rate than those from the SO-fed controls (P<0.05).
The difference in body weight at birth between pups from the SO- or OFO-fed dams disappeared during postnatal life when the surviving pups from both groups were raised under the same conditions of lactation by SO-fed dams, weaning onto chow, and exposure to high-fat diet for 5 wk (Supplemental Fig. 1B). The growth curve for the pups from 0 to 12 wk of age was not affected by the maternal diet, although the females weighed less than the males from 6 wk onwards. From weaning to 12 wk of age, energy intake was not affected by the maternal diet or sex except at 8 wk of age when females ate less than males (Supplemental Fig. 1C). Liver TBARS levels in the offspring at 12 wk of age did not differ among the four groups (9.4 ± 2.8 nmol/g).
Susceptibility of the adult offspring to DIO
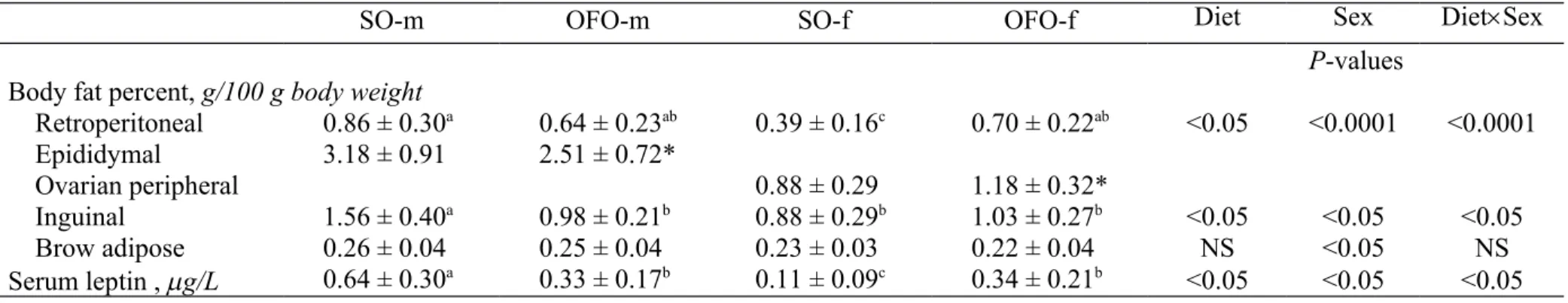
The maternal diet significantly affected the adiposity of the adult offspring exposed to a high-fat diet for 5 wk (Table 1). There were significant effects of maternal diet, sex, 179 180 181 182 183 184 185 186 187 188 189 190 191 192 193 194 195 196 197 198 199 200 201 202 203
and the interactions of the two factors on variables associated with adiposity, such as the body fat percent of the retroperitoneal and inguinal fat and the serum leptin concentration. When offspring from dams fed the SO or OFO diet during gestation were compared within the same gender, the body fat percent of the epididymal and inguinal fat and the serum leptin concentration were significantly lower in the OFO-m group than in the SO-m group, while the body fat percent of the retroperitoneal and ovarian peripheral fat and the serum leptin concentration were significantly greater in the OFO-f group than in the SO-f group. These results show that the adult mice in the OFO-f group were predisposed to DIO, while those in the OFO-m group were protected.
In contrast to WAT, the body fat percent in BAT in the offspring was not affected by maternal diet, but was higher in males than in females.
Serum and liver lipid concentrations in the adult offspring
For serum lipids, a significant interaction of the two factors on serum TG and NEFA concentrations was noticed (Table 2). Compared within the same gender, the OFO-m group had significantly lower serum TG and NEFA concentrations and the OFO-f group had a significantly higher serum TG concentration than the corresponding controls. Neither maternal diet nor sex significantly affected the serum cholesterol concentration.
The liver TG concentration was significantly higher in the male offspring than in the female offspring, while an opposite trend was seen for cholesterol (Table 2). In contrast to the cholesterol concentration, which was not affected by the maternal diet, the TG concentration was significantly reduced by the maternal OFO diet in the male, but not the female, offspring (P-interaction<0.05).
204 205 206 207 208 209 210 211 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228
Hepatic PPAR activation in the pregnant dam and fetus
Hepatic mRNA levels for Cyp4a10 and Acox1, two representative PPAR target genes, were significantly higher in OFO-fed dams than in SO-fed dams at pregnancy d18 (Fig. 1A), showing that PPAR activation was caused by dietary OFO. Levels of the PPAR transcript itself did not differ between the two groups. Consistent with liver enlargement being one of the pleotropic responses to PP, liver weight was significantly greater in the OFO-fed dams (2.4 ± 0.2 g) than in the SO-fed dams (1.7 ± 0.2 g).
A maternal OFO diet throughout gestation also resulted in a significant increase in hepatic PPAR transcriptional activity in the fetus at E18, as shown by significant higher hepatic mRNA levels for Cyp4a10 and Acox1 in the fetuses of OFO-fed dams regardless of sex (Fig. 1B), but had no effect on liver weight (0.05 ± 0.01 g). The maternal OFO diet also resulted in significantly increased Ppara mRNA levels regardless of sex. Fgf21mRNA levels were increased by the maternal OFO diet only in male fetuses (P-interaction<0.05). For fetus sexing, DNA carrying both the Sry and
DXNds genes was identified as male, while that carrying only DXNds3 was identified
as female (Fig. 1C).
Hepatic and WAT gene expression in the adult offspring
To test whether the PPAR activation in the fetus caused by gestational OFO consumption had a long-term effect on lipid catabolism or energy homeostasis in later life, we measured mRNA levels for genes participating in fatty acid catabolism (mainly Ppara and its targets) in the liver in the male and female offspring at 12 wk of age (Fig. 2A). In contrast to the situation in the fetus, Ppara mRNA levels were 229 230 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250 251 252 253
significantly and independently reduced by a maternal OFO diet and female gender of the offspring. Maternal diet did not affect Cyp4a10 and Ehhadh mRNA levels, but had a significant effect on Acox1 mRNA levels and a marginal effect on Cpt1a mRNA levels with a significant interaction with sex. Compared to their SO counterparts, the male, but not the female, offspring of the OFO-fed dams showed significantly increased Acox1 and Cpt1a mRNA levels. There was no effect of maternal diet on the liver weight of the adult offspring, but there was a significant effect of sex (males: 0.99 ± 0.16, females: 0.84 ± 0.10 g).
mRNA levels for genes associated with fatty acid oxidation and thermogenesis were also measured in the WAT of the adult offspring (Fig. 2B). The observed marginal or significant interaction of the two factors on mRNA levels for Ppara,
Acox1, and Ppargc1a could be attributed to significant upregulation of these genes by
the maternal OFO diet in male, but not in female, offspring. Ppargc1a expression was even reduced in females by a maternal OFO diet. The maternal OFO diet significantly and independently increased Ucp1 mRNA levels in the WAT of the adult offspring regardless of sex, while Lep mRNA levels were significantly lower in the OFO-m group, but not the OFO-f group, compared to their corresponding controls (P-interaction<0.05). 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269 270 271
Discussion
In this study, we showed that gestational OFO consumption produced pups with a lower birth weight and fetuses with PPAR activation in accordance with other studies (4, 26). Unexpectedly, when these pups were exposed to a high-fat diet at adulthood, maternal OFO ingestion predisposed females to obesity, but protected males. The protection observed in the male offspring might be associated with enhanced lipid catabolism, which presumably persisted from the fetus to adulthood, as mRNA levels for PPAR target genes showed a marked activation of hepatic PPAR in the fetus and slight activation in the adult and a lower serum/liver TG concentration in the adult in the OFO-m group compared to the SO-m group. In the female offspring, enhanced lipid catabolism associated with PPAR activation caused by maternal OFO ingestion was detectable in the fetus, but not the adult. The upregulation of PPAR target genes in the liver in the adult OFO-m group could not be attributed to oxidized fatty acids from the maternal diet, since they are assumed to be metabolized by microsomal cytochrome P450 enzymes (29) and excreted over time during postnatal life and residual levels in the tissues of these pups at adulthood are expected to be quite low. The composition of OFO is quite complex (30). Currently, it is impossible to track specific oxidized fatty acids in the circulation and/or tissues. Although TBARS was used as an index of peroxidation in this study, establishment of a specific biomarker for oxidized fatty acids (or their metabolites) from OFO is needed.
In this study, the effects observed in female offspring were in accordance with the expected relationship between low birth weight and late onset obesity (15, 20, 21). Maternal undernutrition induced by energy restriction or protein deficiency results in postnatal hyperphagia and sedentary behavior, contributing to obesity in later life (20, 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288 289 290 291 292 293 294 295 296
21). The use of the plasticizer di-(2-ethylhexyl) phthalate (DEHP), which activates PPAR, might cause maternal malnutrition by decreasing maternal plasma levels of TG, which is a significant source of nutrient for the fetus (31). Hayashi et al (32) reported that exposure to DEHP at the fetal and pup stages results in increased food consumption at adulthood when the offspring are fed a control diet, but not when fed a high-fat diet. However, our study showed that the energy intake of the offspring from either the chow or the high-fat diet was not affected by maternal OFO ingestion. Other mechanisms associated with energy expenditure or tissue redistribution of TG require investigation. In this study, the sex-specific responses in the gene expression profile in the WAT and liver to maternal OFO ingestion might involve a complex interaction of sex hormones and epigenetic modification of the fetal genome.
The gene expression pattern in the WAT showed that a maternal OFO diet led to an increase not only in lipid catabolism, but also in thermogenic ability in adult males, and the latter effect might also contribute to their resistance to DIO. The higher mRNA levels for Ppara, Ppargc1a, and Ucp1 in the inguinal fat of adult OFO-m mice compared to SO-m mice suggests that the “browning response” of WAT occurred in the offspring of the OFO-fed dams. Subcutaneous fat, such as inguinal fat, is a thermogenically competent WAT depot with a high propensity to accumulate UCP-1-positive, brown fat-like cells (i.e. “beige” or “brite” cells) (33). The induction of brown-like adipocytes in the WAT is believed to help reduce obesity by increasing heat expenditure (34-36). The plasticity of the adipose organ is a well-known phenomenon in which reversible transformation of the adipocytes between the white and brown phenotypes is seen in response to physiological stimuli. Cold exposure or β-adrenergic stimulation is known to increase the number of brown-like adipocytes in the WAT (37). The findings in this study suggest that FGF21 might be an important 297 298 299 300 301 302 303 304 305 306 307 308 309 310 311 312 313 314 315 316 317 318 319 320 321
molecular signal linking WAT browning in adulthood and uterine OFO exposure. Consistent with the finding that dietary moderately oxidized oil induces Fgf21 expression in the liver of pigs (38), Fgf21mRNA levels were markedly increased in the liver of male fetuses from OFO-fed dams compared to their SO controls. Hondares et al (22) showed that, in rodents, milk intake during suckling (i.e. a high fat diet) is responsible for induction of hepatic Fgf21 expression via PPAR, and that the FGF21 released by the liver contributes not only to metabolic adaptation to a high-fat diet, such as milk, but also to the thermogenic activation of the BAT during the fetal-to–neonatal transition. Although the thermogenic activity of BAT was not investigated in the present study, it is plausible that the maternal OFO diet influences the thermogenic activity of the BAT as early as the embryonic stage, since the BAT is the first adipose tissue detectable at E14.5 (39). After parturition, the upregulated
Fgf21 in pups caused by OFO might also contribute to the browning of the WAT, as
FGF21 has been shown to induce thermogenic gene expression in the WAT and BAT via an increase in PGC-1 protein (24).
In contrast to interscapular BAT, which shows no strain variability, large differences between strains are seen in induction of Ucp1 expression in thermogenically competent WAT as a result of adrenergic signal transduction (40). When mice are reared thermoneutrally, Ucp1 expression in the WAT is transiently seen at postnatal wk 1-2, then declines over time and is maintained at only a low level (41). It should be noted that the genetic variability in the ability of -adrenergic stimulation to induce brown adipocytes (or Ucp1 expression) in the WAT of adult mice can be recapitulated by the transient expression of Ucp1 during early life (41). Our findings inspire us to explore whether the increased browning potential of the WAT in the adult OFO-m group compared to the SO-m group is related to changes, at 322 323 324 325 326 327 328 329 330 331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346
the neonatal stage, in hormone (e.g. catecholamine or FGF21) concentrations or to a higher recruitment of brown adipocytes in the newly-formed WAT. In addition, we would like to know if these events are causally attributable to PPAR activation at the prenatal stage.
In this study, a significant perinatal mortality associated with gestational OFO ingestion was seen. Using oxidized fat prepared at a lower temperature than ours (48 vs. 205℃), Bransch et al (26) reported there was no difference in the number of live pups of Sprague-Dawley rats subjected to oxidized oil during rearing, pregnancy, and lactation. Moreover, no overt reproductive or developmental toxicity was observed in rats or mice subjected to fibrate-class hypolipidemic drugs during pregnancy (42). However, environmental contaminants, such as perfluoroalkyl derivatives or phthalates, have been found to cause reproductive and developmental toxicity in rodents partly through PPAR (42, 43). Differences in the reproductive or developmental toxicity related to dietary OFO or other PPAR activators might be associated with the test compounds, exposure time, dosage, and animal species used.
In summary, gestational OFO ingestion in C57BL/6J mice predisposes the adult female offspring to DIO, but protects the adult male offspring. Increased expression of PPAR target genes in the liver and of thermogenic genes in the WAT was seen in the male adult offspring of OFO-fed dams, along with a reduced TG concentration in the serum and liver, suggesting that increased lipid catabolism and heat expenditure contribute to the resistance to DIO in these mice. In the female adult offspring, no hepatic PPAR activation was induced by maternal OFO ingestion and TG accumulation shifted to the adipose tissue instead of the liver, resulting in a propensity to obesity. Prenatal activation of PPAR might provide an opportunity for study the underlying mechanisms governing the capacity of WAT from an adult to express 347 348 349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368 369 370 371
brown adipocyte developmental program, which is postulated as a way to reduce obesity.
Acknowledgements
P.M.C. designed the study and wrote the paper; H.C.C., C.F.H., Y.C.C., and Y.S.L performed the research; H.C.C. and C.F.H analyzed the data; and P.M.C. had primary responsibility for the final content. All authors have read and approved the final manuscript. 372 373 374 375 376 377 378 379 380 381
Literature Cited
1. Chao PM, Chao CY, Lin FJ, Huang C. Oxidized frying oil up-regulates hepatic acyl-CoA oxidase and cytochrome P450 4 A1 genes in rats and activates PPAR. J Nutr. 2001;131:3166-74.
2. Chao PM, Yang MF, Tseng YN, Chang KM, Lu KS, Huang CJ. Peroxisome proliferation in liver of rats fed oxidized frying oil. J Nutr Sci Vitaminol (Tokyo). 2005;51:361-8.
3. Sulzle A, Hirche F, Eder K. Thermally oxidized dietary fat upregulates the expression of target genes of PPAR in rat liver. J Nutr. 2004;134:1375-83.
4. Ringseis R, Gutgesell A, Dathe C, Brandsch C, Eder K. Feeding oxidized fat during pregnancy up-regulates expression of PPAR-responsive genes in the liver of rat fetuses. Lipids Health Dis. 2007;6:6.
5. Eder K, Suelzle A, Skufca P, Brandsch C, Hirche F. Effects of dietary thermoxidized fats on expression and activities of hepatic lipogenic enzymes in rats. Lipids. 2003;38:31-8.
6. Eder K, Kirchgessner M. The effect of dietary vitamin E supply and a moderately oxidized oil on activities of hepatic lipogenic enzymes in rats. Lipids. 1998;33:277-83.
7. Chao PM, Huang HL, Liao CH, Huang ST, Huang CJ. A high oxidised frying oil content diet is less adipogenic, but induces glucose intolerance in rodents. Br J Nutr. 2007;98:63-71.
8. Liao CH, Shaw HM, Chao PM. Impairment of glucose metabolism in mice induced by dietary oxidized frying oil is different from that induced by conjugated linoleic acid. Nutrition. 2008;24:744-52.
9. Kammerer I, Ringseis R, Eder K. Feeding a thermally oxidised fat inhibits 382 383 384 385 386 387 388 389 390 391 392 393 394 395 396 397 398 399 400 401 402 403 404 405 406
atherosclerotic plaque formation in the aortic root of LDL receptor-deficient mice. Br J Nutr. 2011;105:190-9.
10. Issemann I, Green S. Activation of a member of the steroid hormone receptor superfamily by peroxisome proliferators. Nature. 1990;347:645-50.
11. Mandard S, Muller M, Kersten S. Peroxisome proliferator-activated receptor target genes. Cell Mol Life Sci. 2004;61:393-416.
12. Staprans I, Rapp JH, Pan XM, Feingold KR. Oxidized lipids in the diet are incorporated by the liver into very low density lipoprotein in rats. J Lipid Res. 1996;37:420-30.
13. Staprans I, Rapp JH, Pan XM, Kim KY, Feingold KR. Oxidized lipids in the diet are a source of oxidized lipid in chylomicrons of human serum. Arterioscler Thromb. 1994;14:1900-5.
14. Konig B, Eder K. Differential action of 13-HPODE on PPAR downstream genes in rat fao and human HepG2 hepatoma cell lines. J Nutr Biochem. 2006;17:410-8. 15. Barker DJ, Clark PM. Fetal undernutrition and disease in later life. Rev Reprod.
1997;2:105-12.
16. Vickers MH, Breier BH, Cutfield WS, Hofman PL, Gluckman PD. Fetal origins of hyperphagia, obesity, and hypertension and postnatal amplification by hypercaloric nutrition. Am J Physiol Endocrinol Metab. 2000;279:E83-7.
17. Fernandez-Twinn DS, Ozanne SE. Mechanisms by which poor early growth programs type-2 diabetes, obesity and the metabolic syndrome. Physiol Behav. 2006;88:234-43.
18. Seckl JR, Meaney MJ. Glucocorticoid programming. Ann N Y Acad Sci. 2004;1032:63-84.
19. Neitzke U, Harder T, Schellong K, Melchior K, Ziska T, Rodekamp E, 407 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427 428 429 430 431
Dudenhausen JW, Plagemann A. Intrauterine growth restriction in a rodent model and developmental programming of the metabolic syndrome: A critical appraisal of the experimental evidence. Placenta. 2008;29:246-54.
20. Vickers MH, Breier BH, McCarthy D, Gluckman PD. Sedentary behavior during postnatal life is determined by the prenatal environment and exacerbated by postnatal hypercaloric nutrition Am J Physiol Regul Integr Comp Physiol. 2003;285:R271–3.
21. Vickers MH. Developmental programming of the metabolic syndrome-critical windows for intervention. World J Diabetes. 2011,2:137-48.
22. Hondares E, Rosell M, Gonzalez FJ, Giralt M, Iglesias R, Villarroya F. Hepatic FGF21 expression is induced at birth via PPAR in response to milk intake and contributes to thermogenic activation of neonatal brown fat. Cell Metab. 2010;11:206-12.
23. Hondares E, Iglesias R, Giralt A, Gonzalez FJ, Giralt M, Mampel T, Villarroya F. Thermogenic activation induces FGF21 expression and release in brown adipose tissue. J Biol Chem. 2011;286:12983-90.
24. Fisher FM, Kleiner S, Douris N, Fox EC, Mepani RJ, Verdeguer F, Wu J, Kharitonenkov A, Flier JS, Maratos-Flier E, et al. FGF21 regulates PGC-1 and browning of white adipose tissues in adaptive thermogenesis. Genes Dev. 2012;26:271-81.
25. Badman MK, Pissios P, Kennedy AR, Koukos G, Flier JS, Maratos-Flier E. Hepatic fibroblast growth factor 21 is regulated by PPAR and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007;5:426-37.
26. Brandsch C, Eder K. Effects of peroxidation products in thermoxidised dietary oil in female rats during rearing, pregnancy and lactation on their reproductive 432 433 434 435 436 437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455 456
performance and the antioxidative status of their offspring. Br J Nutr. 2004;92:267-75.
27. Oteiza PI, Olin KL, Fraga CG, Keen CL. Zinc deficiency causes oxidative damage to proteins, lipids and DNA in rat testes. J Nutr. 1995;125:823-9.
28. Kunieda T, Xian M, Kobayashi E, Imamichi T, Moriwaki K, Toyoda Y. Sexing of mouse preimplantation embryos by detection of Y chromosome-specific sequences using polymerase chain reaction. Biol Reprod. 1992;46:692-7.
29. Huang C, Cheung NS, Lu VR. Effects of deteriorated frying oil and dietary protein levels on liver microsomal enzymes in rats. J Am Oil Chem Soc. 1988;65:1796-803.
30. Chang SS, Peterson RJ, Ho CT. Chemical reaction involved in the deep fat frying of foods. JAOCS. 1978;55:718-27.
31. Xu Y, Agrawal S, Cook TJ, Knipp GT. Maternal di-(2-ethylhexyl)-phthalate exposure influences essential fatty acid homeostasis in rat placenta. Placenta. 2008;29:962–9.
32. Hayashi Y, Ito Y, Nakajima T. Relationship of maternal malnutrition caused by di-(2-ethylhexyl) phthalate exposure with lifestyle disease in offspring. Jpn J Hyg. 2012,67:22-5.
33. Seale P, Conroe HM, Estall J, Kajimura S, Frontini A, Ishibashi J, Cohen P, Cinti S, Spiegelman BM. Prdm16 determines the thermogenic program of subcutaneous white adipose tissue in mice. J Clin Invest. 2011;121:96-105.
34. Tsukiyama-Kohara K, Poulin F, Kohara M, DeMaria CT, Cheng A, Wu Z, Gingras AC, Katsume A, Elchebly M, Spiegelman BM, et al. Adipose tissue reduction in mice lacking the translational inhibitor 4E-BP1. Nat Med. 2001;7:1128-32. 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475 476 477 478 479 480 481
35. Hansen JB, Jorgensen C, Petersen RK, Hallenborg P, De Matteis R, Boye HA, Petrovic N, Enerback S, Nedergaard J, Cinti S, et al. Retinoblastoma protein functions as a molecular switch determining white versus brown adipocyte differentiation. Proc Natl Acad Sci U S A. 2004;101:4112-7.
36. Romanatto T, Roman EA, Arruda AP, Denis RG, Solon C, Milanski M, Moraes JC, Bonfleur ML, Degasperi GR, Picardi PK, et al. Deletion of tumor necrosis factor- receptor 1 (TNFR1) protects against diet-induced obesity by means of increased thermogenesis. J Biol Chem. 2009;284:36213-22.
37. Cinti S. Adipocyte differentiation and transdifferentiation: Plasticity of the adipose organ. J Endocrinol Invest. 2002;25:823-35.
38. Varady J, Ringseis R, Eder K. Dietary moderately oxidized oil induces expression of fibroblast growth factor 21 in the liver of pigs. Lipids Health Dis. 2012;11:34. 39. Barak Y, Nelson MC, Ong ES, Jones YZ, Ruiz-Lozano P, Chien KR, Koder A,
Evans RM. PPAR is required for placental, cardiac, and adipose tissue development. Mol Cell. 1999;4:585-95.
40. Guerra C, Koza RA, Yamashita H, Walsh K, Kozak LP. Emergence of brown adipocytes in white fat in mice is under genetic control: effects on body weight and adiposity. J Clin Invest. 1998;102:412-20.
41. Xue B, Rim J, Hogan JC, Coulter AA, Koza RA, Kozak LP. Genetic variability affects the development of brown adipocytes in white fat but not in interscapular brown fat. J Lipid Res. 2007;48:41-51.
42. Peraza MA, Burdick AD, Marin HE, Gonzalez FJ, Peters JM. The toxicology of ligands for peroxisome proliferator-activated receptors (PPAR). Toxicol Sci. 2006;90:269-95.
43. Abbott BD, Wolf CJ, Schmid JE, Das KP, Zehr RD, Helfant L, Nakayama S, 482 483 484 485 486 487 488 489 490 491 492 493 494 495 496 497 498 499 500 501 502 503 504 505 506
Lindstrom AB, Strynar MJ, Lau C. Perfluorooctanoic acid induced developmental toxicity in the mouse is dependent on expression of peroxisome proliferator activated receptor-. Toxicol Sci. 2007;98:571-81.
44. Reeves PG, Nielsen FH, Fahey GC. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr. 1993;123:1939-51.
45. ChenGC, HuangCY, ChangMY, ChenCH, ChenSW, Huang C, ChaoPM. Two unhealthy dietary habits featuring a high fat content and a sucrose-containing beverage intake, alone or in combination, on inducing metabolic syndrome in Wistar rats and C57BL/6J mice. Metabolism. 2011;60:155-164.
507 508 509 510 511 512 513 514 515 516 517 518
Figure legend
Fig. 1 Effect of gestational ingestion of OFO by C57BL/6J mice on hepatic mRNA levels for PPAR and its target genes in the dams at pregnancy d18 (A) and in the fetus at embryonic d18 (B). The gender of the fetus was determined by the presence of DXNds3 and Sry or of DXNds3 alone (C); “f” indicates female and “m” male. In A and B, the data are the mean SD, n=4 for A and n=10 for B. In A, * different from SO-fed dams, P<0.05. In B, results of two-way ANOVA are shown in the inset table. Values not sharing a superscript letter are significantly different, P<0.05. NS, not significant, P≥0.1; OFO, oxidized frying oil; OFO-f, female offspring of dams fed the oxidized frying diet; OFO-m, male offspring of dams fed the oxidized frying diet; SO, fresh soybean oil; SO-f, female offspring of dams fed the fresh soybean oil-diet; SO-m, male offspring of dams fed the fresh soybean oil-diet.
Fig. 2 Effect of gestational ingestion of OFO by C57BL/6J mice on mRNA levels for genes participating in lipid catabolism and thermogenesis in the liver (A) and inguinal fat (B) of the adult offspring at 12 wk of age. Data are mean SD, n=10. Results of two-way ANOVA are shown in the inset table. Values not sharing a superscript letter are significantly different, P<0.05. NS, not significant, P≥0.1; OFO-f, female offspring of dams fed the oxidized frying oil-diet; OFO-m, male offspring of dams fed the oxidized frying diet; SO-f, female offspring of dams fed the fresh soybean oil-diet; SO-m, male offspring of dams fed the fresh soybean oil-diet.
519 520 521 522 523 524 525 526 527 528 529 530 531 532 533 534 535 536 537 538 539 540
TABLE 1 Effect of gestational ingestion of OFO by C57BL/6J mice on variables associated with the adiposity of the adult offspring at 12 wk of age.1
SO-m OFO-m SO-f OFO-f Diet Sex DietSex
P-values
Body fat percent, g/100 g body weight
Retroperitoneal 0.86 ± 0.30a 0.64 ± 0.23ab 0.39 ± 0.16c 0.70 ± 0.22ab <0.05 <0.0001 <0.0001 Epididymal 3.18 ± 0.91 2.51 ± 0.72* Ovarian peripheral 0.88 ± 0.29 1.18 ± 0.32* Inguinal 1.56 ± 0.40a 0.98 ± 0.21b 0.88 ± 0.29b 1.03 ± 0.27b <0.05 <0.05 <0.05 Brow adipose 0.26 ± 0.04 0.25 ± 0.04 0.23 ± 0.03 0.22 ± 0.04 NS <0.05 NS Serum leptin , g/L 0.64 ± 0.30a 0.33 ± 0.17b 0.11 ± 0.09c 0.34 ± 0.21b <0.05 <0.05 <0.05 1 Data are mean SD, n=10. Values not sharing a superscript letter are significantly different, P <0.05. * Different from SO counterparts, P< 0.05. NS, not significant, P≥0.1; OFO-f, female offspring of dams fed the oxidized frying oil-diet; OFO-m, male offspring of dams fed the oxidized frying diet; SO-f, female offspring of dams fed the fresh soybean diet; SO-m, male offspring of dams fed the fresh soybean oil-diet. 541 542 543 544 545 546
TABLE 2 Effect of gestational ingestion of OFO by C57BL/6J mice on serum and liver lipid concentrations in the adult offspring at 12 wk of age.1
SO-m OFO-m SO-f OFO-f Diet Sex DietSex
P-values Serum TG, mmol/L 0.98 ± 0.23a 0.68 ± 0.20b 0.55 ± 0.06c 0.67 ± 0.17b 0.05 <0.0001 <.0001 TC, mmol/L 4.24 ± 0.61 3.88 ± 1.46 4.12 ± 0.48 3.76 ± 0.77 NS2 NS NS NEFA, mmol/L 0.93 ± 0.16a 0.77 ± 0.21b 0.82 ± 0.11b 0.89 ± 0.13ab NS NS <0.05 Liver TG, mol 33.2 ± 11.1a 24.3 ± 8.9b 20.2 ± 3.7b 23.4 ± 4.2b NS <0.05 <0.05 TC, mol 8.68 ± 1.09 8.63 ± 2.10 9.56 ± 2.20 11.2 ± 2.77 NS <0.05 NS
1 Data are mean SD, n=10. Values not sharing a superscript letter are significantly different, P <0.05. NS, not significant, P≥0.1; OFO-f, female offspring of dams fed the oxidized frying oil-diet; OFO-m, male offspring of dams fed the oxidized frying oil-diet; SO-f, female offspring of dams fed the fresh soybean oil-diet; SO-m, male offspring of dams fed the fresh soybean oil-diet.
547 548 549 550 551 552
Online Supporting Material
Supplemental Table 1 Compositions of the test diets used in this study1
SO OFO High-fat
g/kg
Corn starch 561 561 160
Sucrose 100 100 160
Casein 140 140 260
Oxidized frying oil -- 100
--Fresh soybean oil 100 -- 10
Butter -- -- 290
Cellulose 50 50 61
Mineral mixture (AIN93G-MX) 35 35 42
Vitamin mixture (AIN93-VX) 10 10 12
L-Cystine 1.8 1.8 3
Choline bitartrate 2.5 2.5 2
tert-Butylhydroquinone 0.008 0.008
--1 The composition of the fresh soybean oil (SO) and oxidized frying oil (OFO) diets is based on the AIN-93M diet (44). The composition of the high-fat diet is based on Chen et al (45). 553 554 555 556 557 558 559 560 561
Online Supporting Material
Supplemental Table 2 Assay ID of the inventory primers and probes and the sequence of the self-designed primers used for qRT-PCR1
Gene Accession number
Product ID or primer sequence
Ppara NM_001113418.1 Mm00440939_ml Cyp4a10 NM_010011.3 Mm01188913_gl Ucp1 NM_009463.3 Mm01244861_m1 Ppargc1a NM_008904.2 Mm00447181_m1 Lep NM_008493.3 Mm00434759_ml Acox1 NM_015729 F:CCAAGATTCAAGACAGAGCC R:TCCCCTCAAGAAAATCCCC Ehhadh NM_023737.3 F: TGTACCTTCGGGGGTCGGGG R: GGCCCATCGTTCCCAAGCCAAG Cpt1a NM_013495 F: CCAAGATTCAAGACAGAGCC R: TCCCCTCAAGAAAATCCCC Fgf21 NM_020013 F: ACCTGGAGATCAGGGAGGAT R: GTCCTCCAGCAGCAGTTCTC 1 Inventory primers and probes were purchased from Applied Biosystems. 562 563 564 565 566 567 568
Online Supporting Material A
B
C
Supplemental Fig. 1 Effect of gestational ingestion of OFO by C57BL/6J mice on the body weight of the dams during pregnancy (A) and the body weight (B) and energy intake (C) of the pups during postnatal life. Data are mean SD, n=20 for A and n=10 for B and C. In B and C, results of two-way ANOVA are shown in the inset table. NS, not significant, P≥0.1; OFO, oxidized frying oil; OFO-f, female offspring of dams fed the oxidized frying oil-diet; OFO-m, male offspring of dams fed the oxidized frying oil-diet; SO, fresh soybean oil; SO-f, female offspring of dams fed the fresh soybean oil-diet; SO-m, male offspring of dams fed the fresh soybean oil-diet.
Diet Sex DietSex 0-5 wk
NS NS NS
6-12 wk
NS <0.0001 NS
Diet Sex DietSex 8 wk
NS <0.05 NS Other time points
NS NS NS 569 570 571 572 573 574 575 576 577 578 579 580 581 582 583 584 585 586 587