國 立 交 通 大 學
生 物 科 技 學 院
生物科技學系
碩士論文
小兒心室中膈缺損與第二型和第九型基質金屬蛋白
酶基因多型性及酵素活性的相關性
Associations of matrix metalloproteinase-2 and -9 gene
polymorphisms and enzyme activity in the children with
ventricular septal defects
研 究 生: 陳睦元
指導教授: 林志生
博士
小兒心室中膈缺損與第二型和第九型基質金屬蛋白酶基因多型性及酵素活
性的相關性
Associations of matrix metalloproteinase-2 and -9 gene polymorphisms and enzyme activity in the children with ventricular septal defects
研 究 生 : 陳睦元 Student : Mu-Yuan Chen
指導教授 : 林志生 博士 Advisor : Chih-Sheng Lin Ph.D.
國 立 交 通 大 學 生 物 科 技 學 院 生 物 科 技 學 系
碩 士 論 文
A Thesis
Submitted to Department of Biological Science and Technology College of Biological Science and Technology
National Chiao Tung University In partial fulfilment of the requirements
for the degree of master in
Biological Science and Technology July 2012
Hsinchu, Taiwan, Republic of China
Acknowledgement
跑得好快阿,那時間!當我煩惱如何洋洋灑灑的撰寫此頁內容時,才驚覺兩年的研 究所時光已將結束,痛苦、歡樂、掙扎、辛苦的日子不是過得很慢嗎?怎麼轉眼間我已 跨過了人生中一大階段,點滴記憶頓時在心頭湧現。很慶幸我這兩年待在了一個充滿歡 樂又不失專業的實驗室,遇到一群很棒的朋友以及一位非常照顧學生的指導老師。 首先最要感謝的是心臟組終於即將要畢業的深不可測之鐵漢柔情飆仔乖乖大學長 -阿關學長,稱號很長但也描繪出我心目中的你,這兩年讓你擔心了,謝啦!感謝你在 實驗方面的指導,以及生活瑣事的相挺,你認真負責的態度讓我太佩服了,實驗室少了 你真的不行,祝福你事事順心,畢業後娶個美嬌娘。千雅學姊,實驗室的犀利女王,這 個嘴巴真的真的是太厲害了,謝謝妳讓實驗室充滿笑聲,也謝謝妳帶領我們征戰沙場, 大殺四方。耶豆學長,學識一流、才高八斗、電腦高手,沒看過這麼厲害的人耶,羨慕 又忌妒。感謝在我電腦壞掉的期間有你的挺力相助。戴樂,能再工作歷練後還保有這份 赤子之心實屬珍貴,而且你那口才真的太強了就像不死怒火般環繞在實驗室中,等我有 資金在向你討教投資秘訣吧! 子庭,又高又帥又有才真的一定要再提一遍的,你真的是我見過理智線最容易斷的 文青耶!品萱,少根筋行動派少女,很認真的一個女生,不過真的不用對每件事都這麼 憂心忡忡,祝福妳之後工作順利。感謝同屆實驗室兩位同學,陪我一起度過這兩年的生 活,畢業後大家各奔東西,祝你們都能完成自己的夢想。衝阿,好好加油啦! 燕秋,特殊情況會啟動少根筋模式的學妹,妳是稱職的小助手,謝謝妳實驗的幫忙, 也謝謝妳那會停不下來的笑聲響徹了沉悶的實驗室,升上碩二會更加辛苦,加油囉!芳 沅,聰明且落落大方同時又是深藏不露的大胃王。感謝妳會找我討論 Sensor 組的實驗問 題,讓我從中也受益良多,也謝謝妳 office 上的幫忙,格式小天后不是叫假的。 也要感謝大學部的學弟妹的幫忙,佩衡、一華及明慧,你們能在大學活采多姿的生 活之餘,抽空來實驗室學習研究,此用心已勝過大多數人,前除似錦、無可限量,祝福 你們。 相處不久的碩一學妹們葛麗、意涵、碧珊以及采郁,我還來不及與大家熟悉就要離 開,但我很榮幸能教導各位和妳們解說我的實驗,碩班和大學有很大不同,好好適應好好加油! 還要感謝所有球場上的球友陪我流汗、奔跑;謝謝博愛校區的大樹、綠地讓我在實 驗之餘可以散步放鬆身心;謝謝校狗貝蒂、大王的衷心的陪伴。 當然,我最感謝的還是我的指教教授-林志生教授。老師的教導不只是在研究的想 法或觀念上對我有重要的啟發,更多的是在做人做事的態度、方法和責任感上有深深地 影響。也感謝老師在忙綠之餘還抽空對我的論文進行修改及建議。由衷的感謝老師的包 容與指導,對老師的感謝已不是一句感恩就能道盡。到目前的訓練使我羽翼漸豐,即將 翱翔,但對於未來未知且更多險惡的環境顯然是不足的,將來我也將謹記老師的教誨, 更上一層樓。 最後,感謝我親愛的家人,謝謝你們在這兩年對我的支持與鼓勵,以及讓我在這個 物價頗高的新竹生活衣食無缺,使我能無後顧之憂地順利完成學業,謝謝。 陳睦元 謹誌 交通大學生物科技學系碩士班 中華民國一○一年八月
小兒心室中膈缺損與第二型和第九型基質金屬蛋白酶基因多型性
及酵素活性的相關性
研究生 : 陳睦元
指導教授 : 林志生 博士
國 立 交 通 大 學
生 物 科 技 學 院
生 物 科 技 學 系
碩 士 論 文
中文摘要
心室中膈缺損(ventricular septal defect ,VSD)為常見的先天性心臟病之一。其VSDs 佔所有先天性心臟病的20%,多數病童的中膈缺損會隨著年紀增長而逐漸變小或完全閉 合。目前有許多因素被認為與VSD形成有關,其中特定基因的單核苷酸基因多型性(single nucleotide polymorphism, SNP)被指出是造成VSD疾病主要原因之一。基質金屬蛋白酶 (matrix metalloproteinases, MMPs)是一類活性依賴於Zn2+等金屬離子且參與降解細胞外 基質(extracellular matrix, ECM)的蛋白酶家族,其精密調控著組織中ECM的代謝平衡。 MMPs已被報導在胚胎發育、心肌分化,以及心臟組織的重塑扮演著重要的角色,且許 多心臟疾病也與MMPs的基因多型性有關。據此,我們檢測VSDs的病患的MMPs基因 SNPs與其血漿中MMPs活性,來探討MMPs基因SNP和血液中MMPs活性與VSDs之嚴重 程度。 自2010年9月起,我們收集了101位7.4 3.8歲VSDs病童的血液樣品,自白血球萃取 其基因組DNA,利用聚合酶連鎖反應-限制酶片段長度多型性(polymerase chain
reaction-restriction fragment length polymorphism, PCR-RFLP)來檢測這些病童的MMP-9 SNPs,包括 -1562C>T、R279Q及R574P,也檢測MMP-2 -735C>T SNP。我們亦分離病 童的血漿,以明膠酶電泳(gelatin zymography)測定其MMP-2和MMP-9活性。實驗結果顯 示,在所檢測的四個SNPs中,僅有MMP-9 R279Q基因型的分布與VSD的發生有統計上 的顯著意義(p < 0.05),其VSD組中GA基因型的發生頻率明顯高於控制組(分別為52.2%
及37.2%)。另一方面,我們依病患超音波檢查之VSD/aortic root (Ao)比值判定病童VSD 嚴重程度,我們將之分成輕微缺損(VS;VSD/Ao 0.2)、中度缺損(VM;0.2 < VSD/Ao 0.3)及嚴重缺損(VL;VSD/Ao > 0.3) 三組。Gelatin zymography分析結果顯示,在VSD病 童血漿中皆可測得較高活性之MMP-2與MMP-9,特別在VSD嚴重缺損病童中,與控制 組相較,其MMP-2與MMP-9活性表現分別顯著提升60% (p < 0.001) 與50% (p < 0.01)。 本研究結果指出MMP-9 SNP R279Q可能與VSD的發生有關,而隨著VSD趨於嚴重 時,其血液中MMP-2與MMP-9的活性有增加的趨勢。我們推測MMPs的活性可能影響 VSD的嚴重程度或其癒合率,此提供未來繼續探討MMPs影響VSD病理機制十分有價值 的線索。
Associations of matrix metalloproteinase-2 and -9 gene polymorphisms and
enzyme activity in children with ventricular septal defects
Graduate student: Mu-Yuan Chen
Advisor: Chih-Sheng Lin Ph.D.
Department of Biological Science and Technology
College of Biological Science and Technology
National Chiao Tung University
Abstract
Ventricular septal defect (VSD) is the most common form of congenital heart disease. Approximately 20% of patients in congenital heart disease registries have VSD as a solitary lesion. Rates of spontaneous closure for membranous and muscular VSDs in infant and children were 37% and 50%, respectively, during a mean follow-up of 12 months. Single nucleotide polymorphism (SNP) is considered one of the main factors to VSD formation. Matrix metalloproteinases (MMPs) are a family of zinc-dependent endopeptidases involved in breakdown, and physiological homeostasis of extracellular matrix. MMPs have been reported that play the important roles in embryonic development, cardiomyocyte differentiation and causing cardiac tissue remodeling. Also, many cardiac related diseases are associated to the SNP of MMPs gene. Therefore, we tried to determine the SNPs of MMPs genes and plasma MMPs activity in the patients with VSD and investigate the relation between MMPs and VSD in the aspect of incidence and severe level.
Since September 2010, 101 children with VSD aging from 8 to 13 years old were studied and sampled the blood for genomic DNA preparation. The SNPs of MMP-2 (-735C>T) and MMP-9 (-1562C>T, R279Q, and R574P) were determined using polymerase chain reaction followed by restriction fragment length polymorphism (PCR-RFLP) analysis. Plasma MMP-2 and MMP-9 activity were detected by gelatin zymography.
From the four SNPs assay in this study, only the MMP-9 R279Q SNP showed a statistically significant correlation with VSD formation (p < 0.05). The frequency of GA
On the other hand, according to the definition of VSD/aortic root (Ao) ratio, the patients were classified into three groups: VS (VSD/Ao ratio 0.2), VM (0.2 < VSD/Ao 0.3), and VL (VSD/Ao > 0.3) which stand for small, medium and large defect, respectively. The MMP-2 activity in the VS, VM and VL groups is about 40%, 50 and 60% higher than that in the Control group, respectively. MMP-9 has the similar trend of MMP-2. MMP-9 activity of VL group is about 50% higher than that in Control group (p < 0.01).
In conclusion, MMP-9 R279Q may participate VSD pathogenesis; however, there is need to study further for clarifying the relationship between MMP-9 R279Q polymorphism and VSD. The most valuable found out in this research is that MMP-2 and MMP-9 enzyme activity increases with the defect serious level of VSD increase. Our results hint that MMPs expression may affect the severe level of VSD. It is very valuable to further investigate the exact mechanism of MMPs cause effect on VSD in the future.
Content
Acknowledgement ... i
Chinese Abstract ... iii
English Abstract ... v Content ... vii List of Tables ... x List of Figures ... xi I. Literature review 1-1. Single-nucleotide polymorphisms ... 1
1-2. Congenital heart defects ... 2
1-2-1. Ventricular septal defects ... 2
1-2-2. Atrial septal defects ………. 3
1-3. Matrix metalloproteinase ………... 4
1-3-1. Structures and functions of MMPs ……….. 4
1-3-2. Gelatinase A (MMP-2, Type II collagenase) ……….. 5
1-3-3. Gelatinase B (MMP-9, Type V collagenase) ……….. 6
1-4. The roles of MMPs in cardiac development ……….. 6
II. Materials and Methods 2-1. Patients and sample collection ……….……….. 9
2-2. Genomic DNA extraction ……….. 9
2-3. Genetic analysis ………. 10
2-3-1. MMP-2 -735C>T polymorphism ……… 10
2-3-2. MMP-9 -1562C>T polymorphism ……….. 10
2-3-3. MMP-9 R279Q polymorphism ………...… 11
2-4. Gelatin zymography assay ………..……….. 11
2-5. Statistical analysis ……….. 12
III. Results 3-1. The information of patients ……….……….. 13
3-2. Genotype distribution and allele frequency of MMP-2 -735C>T ………... 13
3-3. Genotype distribution and allele frequency of MMP-9 -1562C>T, R279Q and R574P ………. 14
3-3-1. MMP-9 -1562C>T polymorphism ……….. 14
3-3-2. MMP-9 R279Q polymorphism ………... 14
3-3-3. MMP-9 R574P polymorphism ……… 15
3-4. Plasma MMPs activity in the VSD children with different MMP polymorphisms ……….. 16
3-4-1. MMP-2 activity ……….……….. 16
3-4-2. MMP-9 activity ……….………….. 16
3-5. The relationship between plasma MMPs activity and VSD severity ……… 17
3-5-1. MMP-2 activity ………...……….... 17
3-5-2. MMP-9 activity ………...……….... 17
3-6. Investigating the relationship between MMPs activity and spontaneous closure rate of VSD ………...……….…... 18 IV. Discussion ………. 20 V. Conclusions ……… 26 VI. References ………. 28 Tables ……….. 40 Figures ………. 49
Appendixes
Appendix 1 ………... 64 Appendix 2 ………...……….... 68
List of Tables
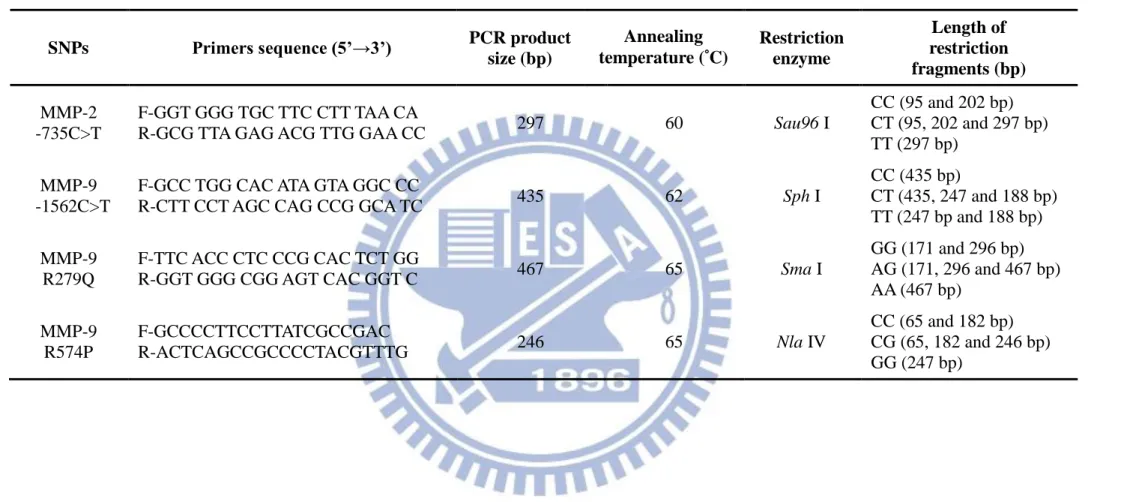
Table 1. Outlines of the information of MMPs and TIMPs SNPs defined and
relationship with particular diseases ………... 40 Table 2. Sequences of the primers used for MMP-2 and MMP-9 genotyping by
PCR-RFLP ……….. 42 Table 3. Genotype distribution and allele frequency of MMP-2 -735C>T
polymorphism ………. 43 Table 4. Genotype distribution and allele frequency of MMP-9 -1562C>T
polymorphism ………. 44 Table 5. Genotype distribution and allele frequency of MMP-9 R279Q
polymorphism ………....………. 45 Table 6. Genotype distribution and allele frequency of MMP-9 R574P
polymorphism ………. 46 Table 7. Incidence of spontaneous closure with respect to type and size of defect ….. 47 Table 8. References of VSDs related gene polymorphisms ………. 48
List of Figures
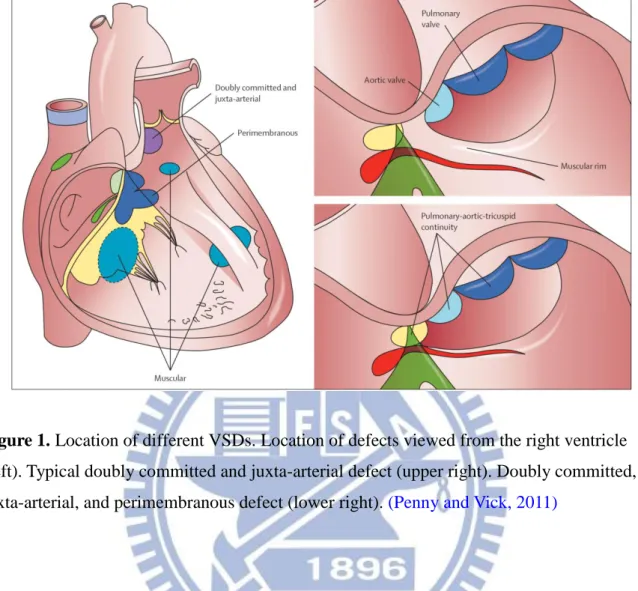
Figure 1. Location of different VSDs ……….. 49
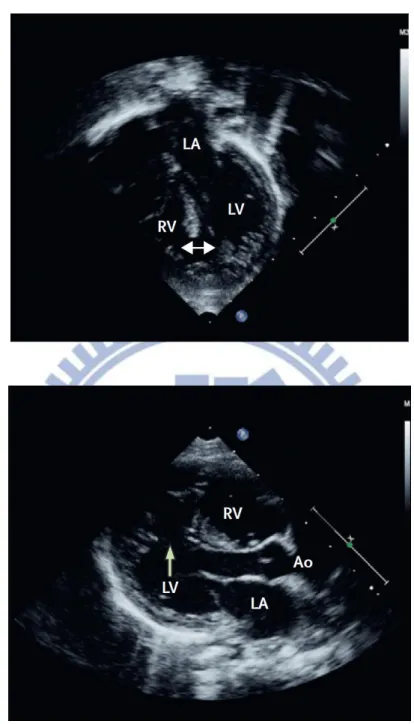
Figure 2. Echocardiograms of VSDs ………..………. 50
Figure 3. Genomic organization and localization of the SNP rs2285053 of human MMP-2 gene ………..………... 51
Figure 4. Detection of the RFLP for MMP-2 polymorphism rs2285053 …... 52
Figure 5. Genomic organization and localization of the SNP rs3918242 of human MMP-9 gene ………..…………... 53
Figure 6. Detection of the RFLP for MMP-9 polymorphism rs3918242 …... 54
Figure 7. Genomic organization and localization of the SNP rs17576 of human MMP-9 gene ………....…………... 55
Figure 8. Detection of the RFLP for MMP-9 polymorphism rs17576 ……... 56
Figure 9. Genomic organization and localization of the SNP rs2250889 of human MMP-9 gene ………..………... 57
Figure 10. Detection of the RFLP for MMP-9 polymorphism rs2250889 …... 58
Figure 11. Plasma MMP-2 activity in the children with VSDs ………... 59
Figure 12. Plasma MMP-9 activity in the children with VSDs ………... 60
Figure 13. Echocardiographic examination for the valuation of VSDs severity ……... 61
Figure 14. Plasma MMP-2 and MMP-9 activity of the children with different VSD severity ………... 62
Figure 15. Relation of MMPs activity and spontaneous closure in different severe levels of VSD patients ………... 63
I. Literature review
1-1. Single-nucleotide polymorphisms
Polymorphism represents natural sequence variants (alleles), which may occur in more than one form. These appear in at least 1% of a population and are considered biologically normal. Approximately 90% of DNA polymorphisms are single-nucleotide polymorphisms (SNPs) due to a single base exchange (Collins et al., 1998). A SNP represents an alternate nucleotide in a given and defined genetic location at a frequency exceeding 1% in a given population. This definition does not include other types of genetic variability like insertions and deletions, and variability in copy number of repeated sequences. They occur once in every 300 nucleotides on average, which means there are roughly 10 million SNPs in the human genome. Although the majority of DNA polymorphisms are probably functionally neutral, a proportion of them can exert allele-specific effects on the regulation of gene expression or function of the coded protein, which underlie individual differences in various biological traits and in susceptibility to disease (Brookes, 1999).
A SNP in which both alleles produce the same polypeptide sequence is called a
synonymous polymorphism (sometimes called a silent mutation); which produce a different polypeptide sequence is called a non-synonymous polymorphism (replacement
polymorphism). A non-synonymous polymorphism can differ from missense and nonsense, which the former results in a different amino acid and the latter results in a premature stop codon. Over half of all known disease mutations come from non-synonymous polymorphisms
(Stenson, 2009). If gene expression is affected by a SNP this type is referred to as an eSNP (expression SNP) and may be upstream or downstream from the gene.
Analytical methods to discover novel SNPs and detect known SNPs include DNA sequencing; restriction fragment length polymorphism (RFLP); capillary electrophoresis;
mass spectrometry; single-strand conformation polymorphism (SSCP); electrochemical analysis; denaturating HPLC and gel electrophoresis; hybridization analysis. All of the references information of SNP in this study was obtain from National Center for Biotechnology Information (NCBI)-SNP database.
1-2. Congenital heart defects
1-2-1. Ventricular septal defects
Ventricular septal defects (VSDs) are the most common form of congenital heart disease. The defect can be in any portion of the ventricular septum, and the physiologic consequences can range from trivial to severe. Approximately 20% of patients in congenital heart disease registries have VSD as a solitary lesion (Hoffman and Rudolph, 1965). Although historically the incidence of VSDs is cited as approximately 1.5 to 3.5 per 1,000 term infants and 4.5 to 7 per 1,000 premature infants (Moe and Guntheroth, 1987), recent echocardiographic studies demonstrated an incidence of VSD in newborns to be 5 to 50 per 1,000 (Tikanoja, 1995). Rates of spontaneous closure for membranous and muscular VSDs in infant and children were 37% and 50%, respectively during a mean follow-up of 12 months (Moe and Guntheroth, 1987). The others have no such a fortunate course and the VSD persist. The development of echocardiography has provided insight into the mechanisms by which VSDs close
spontaneously in a gross point of view (Murphy et al., 1986).
Defects can generally be classified according to their location, either within the muscular septum (muscular defects) or at its margins. Ventricular septal defects at the margins of the muscular septum can be related to hinge-points of the leaflets of the atrioventricular valves (perimembranous), those of the arterial valves (juxta-arterial or subarterial), or both (Figure 1)
pouch or aneurysm derived from tissue from the septal leaflet of the tricuspid valve. Muscular defects appear to close by progressive growth of tissue from the right ventricular side of the circumference of the defect. Neither detail molecular insight of spontaneous VSDs closure has been studied, nor studies of association between extracellular matrix (ECM) and spontaneous VSDs closure have been clarified.
1-2-2. Atrial septal defects
Atrial septal defects (ASDs) represent 6 to 10% of all cardiac anomalies and are more frequent in females than males by about 2:1. ASDs occur in 1 child per 1,500 live births
(Samanek, 1992). A prospective echocardiographic study suggested that as many as 24% of newborns have evidence of an opening (3 to 8 mm) in the atrial septum in the first week of life (Fukazawa et al., 1988). However, by a little more than 1 year of age, 92% of the patients were found to have spontaneous closure of the opening, and in most patients, there is
evidence of a valve-like opening of the atrial septum that is believed to contribute to closure. Helgason and Jonsdottir (Helgason and Jonsdottir, 1999) reviewed the medical records of all patients in Iceland with a diagnosis of ASD born between 1984 and 1993. ASD was
confirmed by 2-D echocardiogram, and data only from patients with secundum ASDs were analyzed. A total of 84 children diagnosed at a mean age of 12 months were followed for 4 years. Spontaneous closure or decreased size was observed in 89% with a 4-mm ASD, 79% with a 5- to 6-mm defect, and only 7% with a defect > 6 mm. Occasionally, spontaneous closure will occur as late as 16 years (Brassard et al., 1999). As the same condition mentioned in the paragraph of VSDs, the underlying mechanism of spontaneous closure of ASDs is till mysterious.
1-3. Matrix metalloproteinase
1-3-1. Structures and functions of MMPs
MMPs is a family of extracellular zinc-dependent neutral endopeptidases (Lombard et al., 2005), capable of degrading essentially all ECM components including fibrillar and
non-fibrillar collagens, fibronectin, laminin and basement membrane glycoproteins (Fedarko et al., 2004). MMPs not only play an important role in ECM remodeling in physiologic situations, such as embryonal development, tissue regeneration, and wound repair, also in pathological conditions including rheumatoid arthritis, osteoarthritis, atherosclerotic plaque rupture, tissue ulceration, and in cancer cell invasion and metastasis (Roeb and Matern, 2001; Jones et al., 2003). The MMP family currently consists of 28 enzymes with somewhat
different activities. The members are generally divided into six groups according to either structure or preferred substrates and referred to as interstitial collagenases (MMP-1, -8 and -13), stromelysins (MMP-3, -10, -11, and -12), matrilysins (MMP-7 and MMP-26),
gelatinases (MMP-2 and MMP-9), membrane-type MMPs (MMP-14, -15, -16, -17, -24, and -25) and others (Visse and Nagase, 2003; Bode and Maskos, 2003). Although MMPs are subclassified based on their ability to degrade 4 various proteins of the ECM, they also play other important roles such as the activation of cell surface receptors and chemokines
(Stefanidakis and Koivunen, 2006). In addition, MMP-2 has proteolytic activity to specific targets within the cell to cause acute, reversible contractile dysfunction in cardiac disease
(Schulz, 2007).
The basic structures of MMPs can be approximately divided into three structurally well-preserved domain motifs, including a catalytic domain, an N-terminal domain and a C-terminal domain. Zinc-dependent catalytic domain of MMPs is similar with subtle
N-terminal domain (propeptide domain) contains a unique PRCG(V/N)PD sequence in which the cysteine residue interacts with the catalytic zinc atom in the active site, prohibiting
activity of the MMPs. Thus, the interaction has to be disrupted to “open” the cysteine switch in the process of MMPs activation (van Wart and Birkedal-Hansen, 1990), which is a critical step that leads to ECM breakdown (Carmeli et al., 2004). The C-terminal hemopexin domain of metalloproteinases has a four-bladed propeller structure and contributes to substrate specificity (Wallon and Overall, 1997). In membrane-type MMPs, the hemopexin domain contains a transmembrane domain for anchoring the protein in the membrane; besides, the hemopexin domain in MMP-2 also has a function in the activation of the enzyme (Morgunova et al., 1999; Overall et al., 1999). The regulation of MMPs occurs at many levels, including transcription (the major one), post-transcriptional modulation of mRNA stability, secretion, localization, zymogen (proenzyme) activation and inhibition of activity by natural inhibitors of MMPs, tissue inhibitor of metalloproteinases (TIMPs). In addition, there are more and more researchers turn their focus on emphasizing MMPs protein physical functions to the change of nucleotides and the SNPs of MMPs and TIMP correlated with different cardiac diseases and cancers(Table 1).
1-3-2. Gelatinase A (MMP-2, Type II collagenase)
In 1978, Sellers et al. were the first to separate a gelatinase activity from collagenase and stromelysin in culture medium from rabbit bone (Sellers et al., 1978). A similar enzyme, acting on basement membrane type IV collagen was reported by Liotta et al. the following year (Liotta et al., 1979). Gelatinase was purified from human skin, mouse tumor cells, rabbit bone, and human gingival. The completed sequence of the human enzyme except for the signal peptide was reported by Collier et al. (Collier et al., 2001). Gelatinase A had a triple
repeat of fibronectin type I domains inserted in the catalytic domain; these participate in binding to the gelatin substrates of the enzyme (Lee et al., 1997; Libson et al., 1995).
1-3-3. Gelatinase B (MMP-9, Type V collagenase)
Harrwas and Krane in 1972 detected a gelatinase activity in rheumatoid synovial fluid. Sopata and Wize described a gelatinase from human polymorphonuclear leukocytes (Sopata and Wize, 1979). Rabbit macrophages produce a very similar enzyme which was able to digest type V collagen (Horwitz et al., 1977). The neutrophil collagenase and gelatinase were resolved in 1980 (Murphy et al., 1980). Purification was achieved in 1983 and sequencing of the cDNA in 1989. An interesting phenomenon, still not fully understood, is the binding of TIMP-1 to proMMP-9 to form a complex (Sakyo et al., 1983; Stetler-Stevenson et al., 1989). Human neutrophil MMP-9 commonly occurs as a complex with lipocalin (Fernández et al., 2005). A series of papers concerned a 95 kDa protein in plasma that binds to gelatin
culminated in the identification of this protein as MMP-9 (Makowski and Ramsby, 1998).
1-4. The roles of MMPs in cardiac development
MMP-2 is the earliest MMP known to be expressed during heart development. The process of cardiac looping which converts the single, straight tubular heart into a S-shaped tube and re-positions the primitive heart chambers into their adult anatomical positions before cardiac septation is complete. During the process of making a single heart tube, MMP-2 is expressed in the endocardium, early differentiating cardiomyocytes, and dorsal mesocardium but is soon lost within the myocardium. Blocking MMP-2 activity prevents midline fusion of the primitive heart tubes leading to cardiac bifida (Cai et al., 2000; Linask et al., 2005).
Cell proliferation is more pronounced within the left splanchnic mesoderm and left dorsal mesocardium. Blocking MMP-2 activity not only disrupts this asymmetric pattern of proliferation, it also randomizes the direction of cardiac looping (Linask et al., 2005) and increases the incidence of dextrocardia (reversal of the normal anatomical position of the heart, i.e., right-sided heart). In other words, blocking MMP-2 activity prevents midline fusion of the primitive heart tubes leading to cardiac bifida (Linask et al., 2005). Therefore, MMP-2 mediated growth appears to be involved in the direction of cardiac looping.
Defects in cardiogenesis during the first three weeks of gestation are usually dangerous. However, anomalous events occurring later in embryonic development often permit the embryo and fetus to make it to term. These anomalies most often manifest themselves as great vessel or cardiac septal defects in neonates. The septation of the atria and ventricles and
division of the cardiac outflow tract into the aorta and pulmonary artery requires the migration, proliferation, and differentiation of two distinct mesenchymal populations,
endocardial-derived cushion cells and invading neural crest (NC) cells (Creazzo et al.,1998). MMPs have been implicated in regulating epithelial-to-mesenchymal transitions (EMTs) which responsible for forming both populations of cells. MMP-2 is expressed by endocardial cells prior to and during the EMT of the endocardium in both the atrioventricular and outflow tract regions of the developing heart (Cai et al., 2000, Alexander et al., 1997).
Hyaluronan is abundant in the ECM of the pre-mesenchymal heart (Markwald et al., 1979). Hyaluronan is an esssential mediator of cell migration and invasion for proper heart development (Camenisch et al., 2002). Hyaluronan relate to regulate the expression of
multiple MMPs in several cell types and directly induces EMT of the endocardium. Therefore, MMPs play pivotal roles for EMT of the endocardial and cardiac septation. Cells of septum primum adjacent ostium primum express MMP-2, TIMP-2, and TIMP-3 during the formation
and remodeling of the muscular septa. MMPs also involve in remodeling events which are responsible for transfiguring the primitive ventricular myocardial wall into a compact layer and inner trabecular layer. MMPs release bioactive VEGFs from the ECM (Belotti et al., 2003; Sounni et al., 2004) and active VEGF-A increases the expression of several MMPs (Pufe et al., 2004; Wang and Keiser ,1998). Therefore, if MMP processing is required for erbB signaling in the developing heart, the specific temporal and tissue-specific expression of MMPs and TIMPs could dictate where and when particular growth factors modulate cardiac remodeling events.
In a nutshell, the heart undergoes remodeling events eliciting changes in MMP activity and ECM turnover as a part of the response to functionally compensate for the extra load. Such responses may reflect MMP-driven remodeling events that occur during embryonic cardiac morphogenesis. All the evidences imply the MMPs play a vital role in cardiac developing.
II. Materials and Methods
2-1. Patients and sample collection
The blood of 101 children of congenital heart defects will be recruited in this study of Taipei and Hsin-Chu Maykay memorial hospital from the outpatient base in one year period. All the patients of congenital heart defects are enrolled in study when they are already
documented by 2-D echocardiography (Figure 2). The 152 control subjects contain no genetic diseases patients or the people who did physical examination in hospital, besides, the plasma of 12 health people were classified as control group in MMPs activity assays.
The blood samples will be obtained by nontraumatic needle aspiration from the
antecubital vein, with no hemolysis occurring in any of the samples when they receive clinical follow-up at 0, 6, and 12 months. The plasma and white blood cells were separated by
centrifuge 2,000 rpm for 10 min, and stored to -80°C.
2-2. Genomic DNA extraction
Genomic DNA for PCR-RFLP genotyping was isolated from the blood of 101 children of congenital heart defects using the genomic DNA Mini kit. Up to 200 L of whole blood add 30 L of Proteinase K (10 mg/mL) and mix briefly. After that, incubate the mixture at 60°C for 15 min. Following then add 200 L of GB Buffer, mix by shaking vigorously and incubate the mixture in a 60°C water bath for 15 min. Add 200 L of absolute ethanol to the sample lysate and immediately mix by shaking vigorously. Transfer the entire mixture (including any precipitate) to the GD Column. Centrifuge at 14,000 x g for 5 min. The collections add 400 L of W1 Buffer to the GD Column and centrifuge at 14,000 x g for 1 min. Then add 600 L of Wash Buffer (ethanol added) to the GD Column and centrifuge at
14,000 x g for 1 min. Displace into new collection tube and centrifuge again at 14,000 x g for 3 min to dry the column matrix. Finally, add 30 L of pre-heated Elution Buffer (10 mM Tris-HCl, pH 8.5) to the matrix and last for 5 min; after that, centrifuge at 14,000 x g for 1 min to elute the purified DNA. The maximum yield is up to 50 μg. Purified genomic DNA were Stored in the -80°C.
2-3. Genetic analysis
2-3-1. MMP-2 -735C>T polymorphism
To analyze the -735C>T polymorphism, we amplified a region of the MMP-2 gene with the primers pair were shown in Table 2. The target sequence was amplified in a 50 µL
reaction volume containing 1 μL of genomic DNA, 1 µL each primer (10µM), 5 µL of 10X PCR buffer, 4 µL of 2.5 mM dNTPs, 0.5 µ L of Taq Polymerase (Promega, Madison, WI, USA), and 37.5 µL distilled water. The PCR started with an initial activation step of 94°C for 2 min followed by 40 cycles of denaturation at 94°C for 30 sec, annealing at 60°C for 30 second, and extension at 72°C for 30 sec, and a final step at 72°C for 7 min. The 297-bp fragment was purified with a purification kit (Invitrogen, Grand Island, NY, USA). Purified PCR products were then digested with Sau96 I (New England BioLabs, Ipswich, MA, USA) at 37°C overnight and separated on a 2% agarose gel. Sau96 I does not digest the T allele but generates 202-bp and 95-bp fragments for the C allele.
2-3-2. MMP-9 -1562C>T polymorphism
The primers used to detect this polymorphism were shown in table, which generated a 435-bp fragment containing the -1562C>T site in a 50 µL reaction volume, and the annealing temperature were shown in Table 2. The PCR product was then digested with Sph I (New
England BioLabs) at 37°C overnight and separated on a 2% agarose gel. Sph I does not digest the C allele but generates 247-bp and 188-bp fragments for the T allele.
2-3-3. MMP-9 R279Q polymorphism
The primers used to detect this polymorphism were sense 5’-GGT GGG TGC TTC CTT TAA CA-3’ and antisense 5’ -GCG TTA GAG ACG TTG GAA CC-3’ (Ikeda et al., 2008), which generated a 467-bp fragment containing the R279Q site in a 50 µL reaction volume, and the annealing temperature were shown in Table 2. The PCR product was then digested with Sma I (New England BioLabs) at 25°C overnight and separated on a 2% agarose gel.
Sma I does not digest the A allele but generates 296-bp and 171-bp fragments for the G allele. 2-3-4. MMP-9 R574P polymorphism
The primers used to detect this polymorphism were shown in Table 2, which generated a 246-bp fragment containing the R574P site in a 50 µL reaction volume, and the annealing temperature were shown in table. The PCR product was then digested with Nla IV (New England BioLabs) at 25°C overnight and separated on a 2% agarose gel. Three genotypes could be distinguished: GG (246-bp), GC (246, 182 and 65-bp) and CC (182 and 65-bp).
2-4. Gelatin zymography assay
The activity of MMP-2 and MMP-9 in the plasma was measured by gelatin gel zymography as previously has described (Chen et al., 2008). Plasma was mixed with 2× zymography sample buffer (0.125 M Tris-HCl, pH 6.8, 20% [v/v] glycerol, 4% [w/v] SDS, 0.005% bromophenol blue) incubated for 10 min at room temperature, and then loaded into SDS-PAGE which was performed in 7% acrylamide gels containing 0.1% (w/v) gelatin (Sigma-Aldrich, St. Louis, MO, USA). After electrophoresis under power supply of 100 V,
the gel was washed twice for 30 min in zymography renaturing buffer (2.5% Triton X-100) with gentle shake at room temperature to remove SDS, then incubated 18 h at 37°C in reaction buffer (50 mM Tris-HCl, pH 7.4, 200 mM NaCl, and 5 mM CaCl2). The gels were
then stained with Coomossie blue for 30 min prior to destain with destain buffer (50%
methanol, 10% acetic acid, and 40% ddH2O). The presence of enzyme activity was evident by
clear or unstained zones, indicating the action of the enzyme on the gelatin substrate
(Stawowy et al., 2004). Gelatinase activities in the gel slabs were quantified by Scion Image software (NIH, Bethesda, MD, USA), which quantifies the area of bands hydrolyzed by gelatinase. A MMP-2 or MMP-9 positive controls (Chemicon, Temecula, CA, USA) was contained in each gel as a standard inensity value to normalize smaple intensityand express in arbitrary units.
2-5. Statistical analysis
Data analysis was performed using GraphPad Prism 5 statistical software. Continuous variables were expressed as mean ± standard deviation (SD), and categorical variables were shown as frequencies and percentages. All polymorphisms were tested for confirmation with Hardy–Weinberg expectations in both groups separately. Differences between patients with and without VSDs were examined with the χ2
test for categorical variables or the Student's t-test and analysis of covariance (ANCOVA) for continuous variables. The association between the MMPs polymorphisms and the risk of VSDs was estimated by odds ratios (ORs) and their 95% confidence intervals (CIs), which were calculated by binary logistic regression models. A 2-tailed p < 0.05 was considered statistically significant.
III. Results
3-1. The information of patients
The 101 congenital heart defects patients contained 44 boys and 57 girls were studied at mean age of 8.2 ± 5.0 years old. The blood of CHD patients recruited from hospital
containing 90 patients of type 2 VSD, 6 patients of other type VSD, and 5 patients of ASD. All the congenital heart defects patients enrolled in this study were already documented by 2-D echocardiography.
3-2. Genotype distribution and allele frequency of MMP-2 -735C>T
The genomic organization of the human MMP-2 gene and localization of the studied SNPs were shown in Figure 3A. In this study, -735C>T polymorphism of MMP-2 was analyzed by PCR-RFLP. The sequence of the human MMP-2 gene and position of the PCR primers which were used to detect MMP-2 -735C>T were shown in Figure 3B. Restriction enzyme digestion was performed on MMP-2 -735C>T PCR products using the restriction enzyme Sau96 I and generate three different length bands between different genotype (Figure 4A). We also used direct sequencing to detect MMP-2 polymorphism in patients with VSD, and there were three genotypes of direct sequencing map for MMP-2 -735C>T shown in
Figure 4B.
The genotype and allele frequencies of MMP-2 -735C>T between groups were reported in Table 3. The frequencies of MMP-2 -735C>T genotypes and alleles in the VSD, and control subjects conformed to the Hardy-Weinberg equilibrium. As showed in Table, the allele frequencies of C and T allele were 66.3% and 33.7%; however, the results were similar to control group (p = 0.92). The genotype frequency of MMP-2 -735C>T were CC: 47.4%, CT
37.9%, TT 14.7%, but no significant differences was found in the genotype frequencies between the VSDs and the control group (p = 0.1554).
3-3. Genotype distribution and allele frequency of MMP-9 -1562C>T,
R279Q and R574P
Localizations of the studied SNPs of human MMP-9 gene were shown in Figure 5A, 7A and 9A. MMP-9 gene contains 13 exons. MMP-9 -1562C>T, R279Q and R574P
polymorphisms of MMP-9 are located on promoter region, exon 6 and exon 10 of MMP-9 gene, respectively. PCR-restriction fragment length polymorphism analysis was used for the genotyping of the MMP-9 -1562C>T, R279Q and R574P polymorphism in this study.
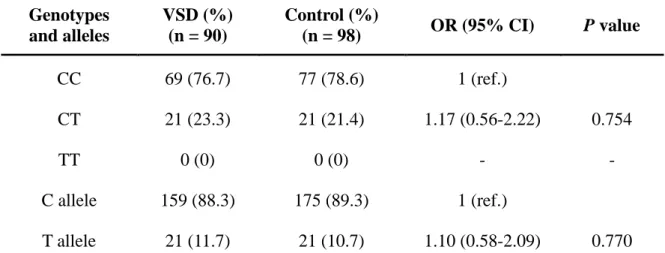
3-3-1. MMP-9 -1562C>T polymorphism
The sequence of the human MMP-9 gene and position of the PCR primers which were used to detect MMP-9 -1562C>T were shown in Figure 5B. PCR product digestion by restriction enzyme Sph I and generate two different length bands between different genotype (Figure 6A). And there were two genotypes of direct sequencing map for MMP-9 -1562C>T were shown in Figure 6B.
The distribution of the MMP-9 -1562C>T genotype were described in Table 4. However, no MMP-9 T/T genotype was detected in patients with VSD and control group. The
distribution of the genotype and allelic status were analyzed by the χ2 value test.
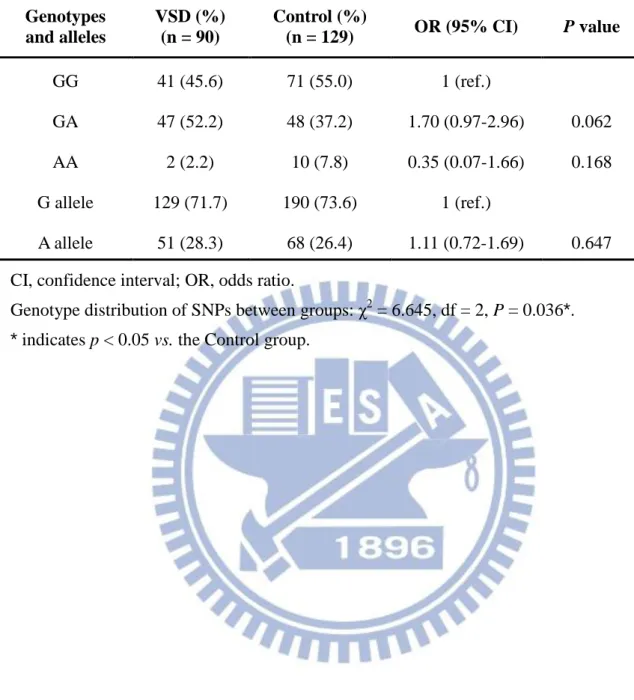
3-3-2. MMP-9 R279Q polymorphism
The sequence of the human MMP-9 gene and position of the PCR primers which use to detect MMP-9 R279Q were shown in Figure 7B. PCR product digestion by restriction
467-bp, 296-bp, and 171-bp, GG genotype: 296-bp and 171-bp) between different genotype (Figure 8A). And there were three genotypes of direct sequencing map for MMP-9 R279Q
were shown in Figure 8B.
The genotype and allele frequencies of MMP-9 R279Q between VSD and control group
were summarized in Table 5. As shown in Table 5, the genotype frequency of MMP-9 R279Q was GG: 45.6%, GA: 52.2% and AA: 2.2%. The results indicate that the genotype distribution MMP-9 R279Q have statistically significant differences compared to control group which the samples of patients without VSD (p = 0.0361); however, there were no significant differences was found in the genotype frequencies between the VSD and the control group (p = 0.6635).
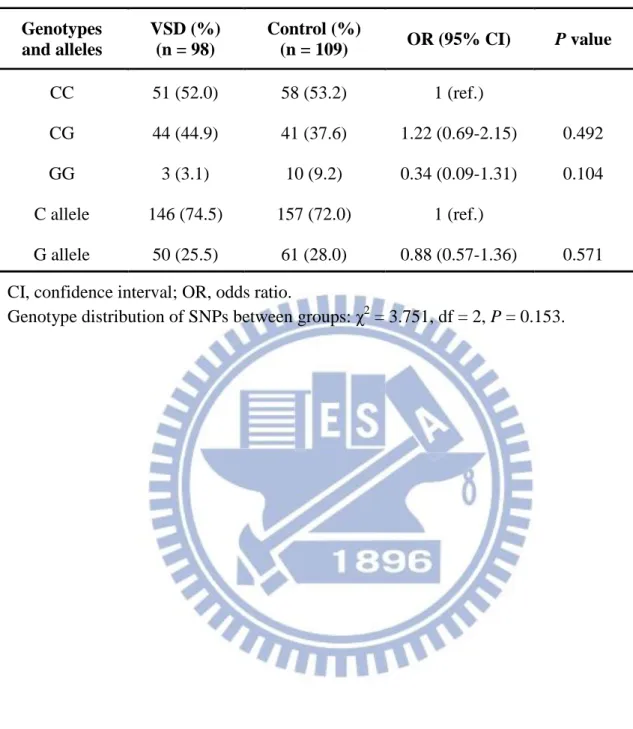
3-3-3. MMP-9 R574P polymorphism
The sequence of the human MMP-9 gene and position of the PCR primers which were used to detect MMP-9 R574P were shown in Figure 9B. PCR product digestion by restriction enzyme Nla IV and generate three different length bands between different genotype (Figure 10A). And there were three genotypes of direct sequencing map for MMP-9 R574P were shown in Figure 10B. The genotypes and allele frequencies of MMP-9 R574P that found in patients with VSD were reported in Table 6. The results shows that the homozygous C/C, heterozygous C/G and homozygous G/G genotypes of MMP-9 R574P were 52.0%, 44.9% and 3.1%, respectively, in the VSD group and 53.2%, 37.6% and 9.2%, respectively, in the control group. There were no significant differences in genotypes (p = 0.1533) and allele frequencies (p = 0.1664) between groups.
3-4. Plasma MMPs activity in the VSD children with different MMP
polymorphisms
In order to investigate the differences of each MMPs activity in VSD patients that have different genotype of each MMPs polymorphism, we used the gelatin zymography to detect MMP-2 and MMP-9 activity in plasma from congenital heart defects children.
3-4-1. MMP-2 activity
MMP-2 activity in sample plasma was detected by gelatin zymography and samples were classified according to genotype of patients on MMP-2-735, which has CC, CT, TT genotype and each group contained 45, 39, and 13 samples, respectively. Used standard MMP-2 as positive controls (Chemicon, Temecula, CA, USA) to calculate MMP-2 activity of each group and collected the statistic as shown in Figure 11. MMP-2 activities have no significant difference in MMP-2 subgroups, but TT allele has the highest MMP-2 activity in MMP-2 -735C>T group (p = 0.052).
3-4-2. MMP-9 activity
For investigating the relation of MMP-9 activity in the VSDs children and MMP-9 SNP, three SNP were chosen, MMP-9 -1562C>T, MMP-9 R279Q and MMP-9 R574P. According to different genotype of patients underwent classification. Figure 12A shows the part of MMP-9 R279Q and is departed in GG, GA, AA and each group have 42, 50, 2 samples, sequentially.
Figure 12B shows the part of MMP-9 R574P and is departed in CC, CG, GG and recruit 51,
44, 3 samples, respectively. Figure 12C shows the part of MMP-9 -1562C>T and is classified in CC, CT and includes 69 and 21 patients, respectively. We used standard MMP-9 as positive controls (Chemicon, Temecula, CA, USA) to calculate MMP-9 activity of each group. As the data showed, MMP-9 activities have no significant difference in the subgroups of MMP-9 genotypes.
3-5. The relationship between plasma MMPs activity and VSD severity
Anatomically, VSDs are classified in to three defect levels: large, medium and small. The large defect is defined as the size is larger than two-thirds of the aortic root diameter; the medium size is in between two-thirds and one-third of the aortic root diameter; and small if the size is less than one-third of the aortic root diameter(Hornberger et al., 1989; Kabra and Srivastava, 2010).Figure 13 shows the Echocardiograms of normal, small defect and medium defect. The sample of patients with VSD conducted in this study are relative not severe
population, which the severe population means surgery needed. According to the definition of VSD/aortic root (Ao) ratio, the patients were classified into three groups: VSD/Ao ratio 0.2 is VS group; 0.2 < VSD/Ao 0.3 is VM group; and VSD/Ao > 0.3 is considered to VL group. The VS, VM and VL stand for small, medium and large defect, respectively; each group includes 12, 45, 25 and 15 patients, respectively. The MMPs activity was detected by zymography, and the results were shown in Figure 14.
3-5-1. MMP-2 activity
According to figureFigure 14A, the MMP-2 activity of VS group is 1.4 fold higher than control group (p < 0.01), besides, VM and VL group MMP-2 activity are 1.6 fold higher than control group (p < 0.01, p < 0.001, respectively).
3-5-2. MMP-9 activity
In the part of MMP-9 activity, the patients also were separately assigned into VS, VM and VL group, and three of them were compared to control group (no VSD). Figure 14B
shows that there is no significantly difference between MMP-9 activity of VS defect level and control group; but when defect level rises to VM defect level, the MMP-9 activity
significantly increases (p < 0.05); as defect level considered large also performs same trend which MMP-9 activity of VL group is about 1.5 fold higher than control group.
3-6. Investigating the relationship between MMPs activity and spontaneous
closure rate of VSD
The patients with VSD were total diagnosed with echocardiographic examination twice with the interval of six months. Patients were initially assigned by first echocardiographic examination result based on t VSD/Ao ratio and then all samples were classified into VS, VM and VL groups. For the purpose of investigating the relation between MMPs activity and spontaneous closure rate of VSD, the ratio of VSD to aortic root from second
echocardiographic examination result were compared to first ratio. If the compared ratio value were above 15 % , then the patient with VSD were viewed as happened spontaneous closure of VSD; if the value below 15%, then the patient with VSD were considered as happened no spontaneous closure or personal equation. There total were 73 patients conducted in this experiment, the statistic table of spontaneous closure classification were shown at Table 7. If the patient with VSD were operated or had only one time echocardiographic examination, then will not include in this study. The relative MMPs activity was defined as the spontaneous closure MMPs activity to the basic of non-spontaneous closure MMPs activity.
Figure 15A denotes that the MMP-2 activity in spontaneous closure subgroup of VS group is significantly lower than non-spontaneous closure subgroup (p < 0.05), indicating that the mechanism of spontaneous closure might suppress the activity of MMP-2. However, the unusual point is that the phenomenon presented in VM and VL subgroups are opposite to VS, which non-spontaneous closure subgroup owns not significantly but higher MMP-2 activity
than spontaneous closure subgroup. The study of MMP-9 also demonstrates same trend in MMP-2 as showed in Figure 15B.
IV. Discussion
The first description of MMP-like activity was related to the metamorphosis of the tadpole (Brinckerhoff and Matrisian, 2002). This and many other subsequent discoveries strongly suggest that this well-characterized family of metalloproteinases is also intimately involved in normal tissue growth and ECM remodeling (Stamenkovic, 2003). Morphogenesis and tissue growth, remodeling, and repair are sentinel features of childhood and adolescence. Heart morphogenesis involves the characteristic mechanisms of cell proliferation, cell
rearrangements, cell to cell, cell to matrix interactions, and tissue remodeling. Linask et al. documented MMP activity regulates the coordination of early heart organogenesis by
affecting ventral closure of the heart and gut tubes, asymmetric cell proliferation in the dorsal mesocardium to drive looping direction, and ECM degradation within the dorsal mesocardium allowing looping to proceed toward completion (Linask et al., 2005), although little is known about the production, secretion, and clearance of these important proteinases throughout normal growth and development in human heart
At present, information suggests that the septum has both mesenchymal and muscular components (Contreras-Ramos et al., 2008). The mesenchymal element originates mainly from fusion of the conotruncal and atrioventricular endocardial cushions. Mechanisms that initiate development of the muscular septum are less well defined, but at least two processes have been proposed. The first process is that the muscular septum forms from coalescence of the part of the ventricular wall is interposed between the enlarging free walls of the
developing right and left ventricles; therefore, as the ventricular cavities become deeper the septum grows passively inwards ( Goor et al., 1970). The second process is an alternative hypo thesis suggesting that the muscular septum originates from a cluster of cells and the
so-called primitive inter ventricular septum, which expands actively towards the cushions of the atrioventricular canal (De La Cruz and Moreno-Rodriguez, 1998).
Gene polymorphism also is considered an essential cause of VSD formation; especially the TBX5 and GATA4 SNP are frequently discussed. These gene both express in the heart and their interaction is vital for normal cardiac septation (Maitra et al.,2009). TBX5 is a vital gene during embryonic differentiation which affects cardiac and upper limbs development. When TBX5 occurs mutation may cause the loss of this gene function of and therefore cause cardiac and upper limbs malformations (Basson et al.,1997). A TBX5 polymorphismis also associated with ventricular septal defect in the Chinese Han population (Liu et al., 2009).
There are many researches point out that GATA4 gene mutationis related to the happen of ASD and VSD (Zhang et al., 2009; Chen et al., 2010b; Rajagopal et al.,2007). The other VSD related genes were list in Table 8. According to above, this research is based on studying the association of MMPs SNP and MMPs activity to the formation of VSDs and may participate the mechanism of spontaneous closure of VSDs.
To realize MMPs polymorphism from National Center for Biotechnology Information (NCBI)-SNP database, which had registered over 700 SNPs of MMP-2 and 300 SNPs of MMP-9. The relatively important and correlative literatures were organized and shown in
Appendix 1 for MMP-2and Appendix 2 for MMP-9. As MMPs play important role in many physiological functions, MMP polymorphism also has proven to be relative to many diseases. Gene transcription is the primary point of regulation of MMPs, which implicates that
sequence changes in the promoter may be important for the transcription and cause effect in protein levels and cell physiology.
Two of MMPs polymorphisms in promoter region were detected in this study: MMP-2 -735C>T and MMP-9 -1562 C>T. The reason of chosen these two promoter will described as
following. The C>T polymorphism, located at nucleotide -735, destroys a Sp1-binding element and the T allele is associated with significantly diminished promoter activity (Yu et al., 2004). The -1562 C>T polymorphism in the promoter region of MMP-9 exerts an T allelic effect on MMP-9 promoter strength and results in losing a nuclear repressor protein binding site which decreasing MMP- 9 expression as the T allele is present, and thus increasing the enzyme expression compared to the C allele (Morgan et al., 2003; Zhang et al., 1999).
Besides, if SNPs occurred at gene exon sequence, the structure of enzyme may change which further affects itself activity. So, this study also examined SNP in MMP-9 rs17576 (Q279R) and rs2250889 (R574P). SNP in MMP-9 rs17576 (Q279R) is a non-conservative amino acid substitution that modifies an amino acid residue within the highly conserved gelatinase-specific fibronectin type II domain(FN2) (Allan et al., 1995). Although the specific functional consequences of the substitution of proline for arginine coded by
rs2250889 in MMP-9 (R574P) have not been studied yet, the location of SNP rs2250889 in the C-terminal hemopexin-like domain could lead to an important protein structural
modification. This structural modification could either decrease the affinity of MMP-9 for its specific inhibitors, resulting in a higher level of protein activity, or enhance the affinity binding for gelatin which might increase its gelatinase activity (Rodríguez-Pla et al.,2008).
All SNPs above have more potential in ruling and causing MMP-2 and MMP-9 to significant change than other SNPs, which making them to be the target SNP in this research. In this study, the SNPs of the MMPs genes, such as MMP-2 (-735C>T) and MMP-9
(-1562C>T, R279Q, and R574P) were determined by the polymerase chain reaction, restriction fragment length polymorphism method (Table 2).The results showed that genotype distribution and allele frequencies of MMP-2 -735C>T between the VSD and the control group had no significant differences (Table 3). This result suggest that C>T
polymorphism located at nucleotide -735 may not influence the incidence of VSD or not the core factor in Taiwan individuals.
MMP-2 activity in VSD patient plasma were also examined by zymography, categorized by their different genotypes (CC, CT, and TT) of MMP-2 -735 polymorphism and conducted in statistic. As mentioned earlier that C>T polymorphism located at nucleotide -735 should significantly diminish promoter activity(Yu et al., 2004); however, the statistical results in this study showed that there was no significant correlation between MMP-2 -735 genotypes and their activity. The result also showed that the T allele of MMP-2 -735 polymorphism has the highest MMP-2 activity which is different from the results of Yu C et al, suggesting C allele enhanced MMP-2 protein transcription which corresponds with higher MMP-2 activity
(Yu et al., 2004).
Polymorphisms in the promoter of MMP-9 have been implicated in the regulation of gene expression and susceptibility to various diseases. The results of genotype distribution and allele frequencies of MMP-9 -1562 C>T between the VSD and the control group were shown in Table 4. The results show that polymorphism in the promoter of MMP-9 -1562 C>T
has no correlation to VSD. However, there is no VSD patient have TT genotype of MMP-9 -1562 C>T in our study. We speculate that lack of T allelic genotype of MMP-9
polymorphism is relative to population and region, which is correspond toother disease studiesconducted in Asian population also had few or even no T allelic genotype of MMP-9 polymorphism in their samples (Chen et al., 2010a; Lacchini et al., 2010; Buss et al., 2009).
The polymorphism at position -1562 were expected to change the promoter activity of MMP-9 (Zhang et al., 1999; Van den Steen et al., 2002); however, our results showed no relationship between genotypes of MMP-9 -1562 polymorphism and their activity. Our results
communicates with Demacq et al. whose plasma samples were recruited from healthy subjects (Demacq et al., 2006).
In detection of MMP-9 codon polymorphisms: MMP-9 Q279R and R574P, our data demonstrated that there is an association of the MMP-9 Q279R polymorphism and risk of VSD (Table 5 and Table 6). The importance of MMP-9 Q279R is based on the ability of changing MMP-9 protein structure and their substrate binding affinity which related to many diseases, such as melanoma, lumbar-disc herniation, type 2 diabetes, and pelvic organ prolapse (Ahluwalia et al., 2009; Cotignola et al., 2007; Hirose et al., 2008; Chen et al., 2010a). The GA genotype distribution in VSD group is much higher than control group. Although there were few subjects of AA genotype in VSD group, there is an obvious difference in genotype distribution to control group.
Blankenberg et al. found that the G allele of MMP-9 R574P polymorphism was overrepresented in patients with histologically confirmed giant cell arteritis (GCA)
(Blankenberg et al., 2003); however, we do not find any significant differences in both the genotype and allele distribution between the VSD and control groups in Taiwanese population. According to VSD patients different genotypes of MMP-9 Q279R and R574P polymorphism, there is no significant difference showed in MMP-9 activity detection, which indicates these SNPs may cause effect in protein structure and substrate binding efficiency but no notable effect in enzyme activity.
The VSD patients according to their serious level were further categorized to four groups. All groups were conducted of the study that whether the serious level is correlative to MMPs activity. The most valuable find out in this research is that MMP-2 and MMP-9 enzyme activity increases with the defect serious level of VSDs increase (Figure 14). Indicating that
MMPs expression may affect the severe level of VSDs, and interfere the closure rate of VSDs.
The rate of spontaneous closure of VSDs has been reported to be between 11% and 70.8% in various researches (Kidd et al., 1993; Mehta and Chidambaram, 1992; Ahunbay et al., 1999; Eroğlu et al., 2003). In our study, spontaneous closure was detected in 23% of perimembranous defect (Table 7). Commonly, patient with small defect is considered more easy to happen spontaneous closure for not violent blood stream; but in our study, our samples showed that patients with severe defect are more possible to happen spontaneous closure, which the possibility of spontaneous closure are 38% and 14% for severe and slight VSD, respectively.
In several studies, it was proposed that ventricular septal aneurysm is an important mechanism of closure and shows a more favorable prognosis in perimembranous defects
(Freedom et al., 1974; Ramaciotti et al., 1995). This study proposes that in vivo proteases are very possible relative to spontaneous closure of VSD. Figure 15 demonstrates that
spontaneous closure subgroup of VS group owns lower MMPs activity than the other
subgroup, and the results are opposite to VM and VL group, which their spontaneous closure subgroup own higher MMPs activity. Combined the results from Figure 14 and Figure 15, the reasons of VM and VL group have higher MMPs activity expressed in spontaneous closure subgroup are suspected to their severe defect level and the role of MMPs participating in spontaneous closure of VSD. However, the role of MMPs participating in spontaneous closure of VSD is advantage to closure or not still unknown. The further research of mechanism about MMPs in VSD closure is needed in the future.
V. Conclusions
Ventricular septal defect is one of the most common form of congenital heart diseases, and there are about 50% children patients will spontaneous closure in 12 months; however, it still remains vague about the mechanisms of VSD occurrence and its’ spontaneous closure. VSD children patients’ blood were collected and processed with extracting their genome for MMPs genotyping and separating plasma for MMPs activity evaluation. The genotypes of the MMP2 -735C>T polymorphisms and MMP9 -1562C>T, R279Q and R574P polymorphisms were determined using PCR-RFLP. Plasma MMPs activity was determined by zymography in VSDs children with different MMP polymorphisms.
The results in this study indicate that the R279Q polymorphism of MMP-9 gene is significantly associated with VSD risk in Taiwan children. However, both enzyme activity and genotype distribution of another MMPs SNP in this study were not associated with VSD. Furthermore, VSD patients according to echocardiography were classified into different severe group for investigating the relationship between plasma MMPs activity and VSD severity and the relationship between MMPs activity and spontaneous closure rate of VSD. We find that when the severity of VSD rises then MMP-2 and MMP-9 activities increase. And the experiments results show that MMPs participate in spontaneous closure mechanism of different severity VSD.
This study finds out the potential role of MMP SNPs in regulation of ECM effector and the recovery of the congenital defects in children. Also help physician to elucidate the different activities among different MMPs members in these common clinical situation. The work may offer the predictable factor about which group of cardiac defects will recovery spontaneously. Besides, the results may point-out the novel targets which are significantly in
the process of heart recovery, and the researches might modulate the natural remodeling process by regulating the target through pharmacological therapy.
VI. References
Ahluwalia TS, Khullar M, Ahuja M, Kohli HS, Bhansali A, Mohan V, Venkatesan R, Rai TS, Sud K, Singal PK. 2009. Common variants of inflammatory cytokine genes are associated with risk of nephropathy in type 2 diabetes among Asian Indians. PLoS One 4:e5168. Ahunbay G, Onat T, Celebi A, Batmaz G. 1999. Regression of right ventricular pressure in
ventricular septal defect in infancy: a longitudinal color-flow Doppler echocardiographic study. Pediatr Cardiol. 20:336-342.
Alexander SM, Jackson KJ, Bushnell KM, McGuire PG. 1997. Spatial and temporal
expression of the 72-kDa type IV collagenase (MMP-2) correlates with development and differentiation of valves in the embryonic avian heart. Dev Dyn. 209:261-268.
Allan JA, Docherty AJ, Barker PJ, Huskisson NS, Reynolds JJ, Murphy G. 1995. Binding of gelatinases A and B to type-I collagen and other matrix components. Biochem J.
309:299-306.
Ban HJ, Heo JY, Oh KS, Park KJ. 2010. Identification of type 2 diabetes-associated combination of SNPs using support vector machine. BMC Genet. 11:26.
Basson CT, Bachinsky DR, Lin RC, Levi T, Elkins JA, Soults J, Grayzel D, Kroumpouzou E, Traill TA, Leblanc-Straceski J, Renault B, Kucherlapati R, Seidman JG, Seidman CE. 1997. Mutations in human TBX5 [corrected] cause limb and cardiac malformation in Holt-Oram syndrome. Nat Genet. 15:30-35.
Beeghly-Fadiel A, Lu W, Long JR, Shu XO, Zheng Y, Cai Q, Gao YT, Zheng W. 2009. Matrix metalloproteinase-2 polymorphisms and breast cancer susceptibility. Cancer Epidemiol Biomarkers Prev. 18:1770-1776.
Beeghly-Fadiel A, Lu W, Shu XO, Long J, Cai Q, Xiang Y, Gao YT, Zheng W. 2011. MMP9 polymorphisms and breast cancer risk: a report from the Shanghai Breast Cancer Genetics Study. Breast Cancer Res Treat. 126:507-513.
Belo VA, Souza-Costa DC, Luizon MR, Lanna CM, Carneiro PC, Izidoro-Toledo TC, Ferraz KC, Tanus-Santos JE. 2012. Matrix metalloproteinase-9 genetic variations affect MMP-9 levels in obese children. Int J Obes (Lond). 36:69-75.
Belotti D, Paganoni P, Manenti L, Garofalo A, Marchini S, Taraboletti G, Giavazzi R. 2003. Matrix metalloproteinases (MMP9 and MMP2) induce the release of vascular endothelial growth factor (VEGF) by ovarian carcinoma cells: implications for ascites formation. Cancer Res. 63:5224-5229.
Blankenberg S, Rupprecht HJ, Poirier O. 2003. Plasma concentrations and genetic variation of matrix metalloproteinase 9 and prognosis of patients with cardiovascular disease. Circulation 107:1579–1585.
Bode W, Maskos K. 2003. Structural basis of the matrix metalloproteinases and their physiological inhibitors, the tissue inhibitors of metalloproteinases. Biol Chem. 384:863-872.
Brassard M, Fouron JC, van Doesburg NH, Mercier LA, De Guise P. 1999. Outcome of children with atrial septal defect considered too small for surgical closure. Am J Cardiol. 83:1552-1555.
Brinckerhoff CE, Matrisian LM. 2002. Matrix metalloproteinases: a tail of a frog that became a prince. Nat Rev Mol Cell Biol. 3:207-214.
Brookes AJ. 1999. The essence of SNPs. Gene 234:177-186.
Buss A, Pech K, Roelver S, Bloemeke B, Klotzsch C, Breuer S. 2009. Functional
polymorphisms in matrix metalloproteinases -1, -3, -9 and -12 in relation to cervical artery dissection. BMC Neurol. 9:40
Cai DH, Vollberg TM Sr, Hahn-Dantona E, Quigley JP, Brauer PR. 2000. MMP-2 expression during early avian cardiac and neural crest morphogenesis. Anat Rec. 259:168-179. Camenisch TD, Schroeder JA, Bradley J, Klewer SE, McDonald JA. 2002. Heart-valve
mesenchyme formation is dependent on hyaluronan-augmented activation of ErbB2-ErbB3 receptors. Nat Med. 8:850-855
Carmeli E, Moas M, Reznick AZ, Coleman R. 2004. Matrix metalloproteinases and skeletal muscle: a brief review. Muscle Nerve 29:191-197.
Chen CL, Huang SK, Lin JL, Lai LP, Lai SC, Liu CW, Chen WC, Wen CH, Lin CS. 2008. Upregulation of matrix metalloproteinase-9 and tissue inhibitors of metalloproteinases in rapid atrial pacing-induced atrial fibrillation. J Mol Cell Cardiol. 45:742-753.
Chen HY, Lin WY, Chen YH, Chen WC, Tsai FJ, Tsai CH. 2010a. Matrix metalloproteinase-9 polymorphism and risk of pelvic organ prolapse in Taiwanese women. Eur J Obstet
Gynecol Reprod Biol. 149:222-224.
Chen Y, Han ZQ, Yan WD, Tang CZ, Xie JY, Chen H, Hu DY. 2010b. A novel mutation in GATA4 gene associated with dominant inherited familial atrial septal defect. J Thorac Cardiovasc Surg. 140:684-687.
Cheong JY, Cho SW, Lee JA, Lee KJ, Wang HJ, Lee JE, Kim JH. 2008. Matrix
metalloproteinase-3 genotypes influence recovery from hepatitis B virus infection. J Korean Med Sci. 23:61-65.
Collier IE, Saffarian S, Marmer BL, Elson EL, Goldberg G. 2001. Substrate recognition by gelatinase A: the C-terminal domain facilitates surface diffusion. Biophys J. 81:2370-2377. Collins FS, Brooks LD, Chakravarti A. 1998. A DNA polymorphism discovery resource for
research on human genetic variation. Genome Res. 8:1229-1231
Contreras-Ramos A, Sánchez-Gómez C, García-Romero HL, Cimarosti LO. 2008. Normal development of the muscular region of the interventricular septum--I. The significance of the ventricular trabeculations. Anat Histol Embryol. 37:344-351.
Cotignola J, Reva B, Mitra N, Ishill N, Chuai S, Patel A, Shah S, Vanderbeek G, Coit D, Busam K, Halpern A, Houghton A, Sander C, Berwick M, Orlow I. 2007. Matrix Metalloproteinase-9 (MMP-9) polymorphisms in patients with cutaneous malignant melanoma. BMC Med Genet. 8:10.
Creazzo TL, Godt RE, Leatherbury L, Conway SJ, Kirby ML. 1998. Role of cardiac neural crest cells in cardiovascular development. Annu Rev Physiol. 60:267-286.
De La Cruz MV, Moreno-Rodriguez R. 1998. Embryological development of the apical trabeculated region of both ventricles: the contribution of the primitive interventricular septum in ventricular septation. Living morphogenesis of the heart. pp 120-130. Demacq C, de Souza AP, Machado AA, Gerlach RF, Tanus-Santos JE. 2006. Genetic
polymorphism of matrix metalloproteinase (MMP)-9 does not affect plasma MMP-9 activity in healthy subjects. Clin Chim Acta 365:183-187.
Eroğlu AG, Oztunç F, Saltik L, Bakari S, Dedeoğlu S, Ahunbay G. 2003. Evolution of
ventricular septal defect with special reference to spontaneous closure rate, subaortic ridge and aortic valve prolapse. Pediatr Cardiol. 24:31-35.
Fatar M, Stroick M, Steffens M, Senn E, Reuter B, Bukow S, Griebe M, Alonso A, Lichtner P, Bugert P, Meitinger T, Wienker TF, Hennerici MG. 2008. Single-nucleotide
polymorphisms of MMP-2 gene in stroke subtypes. Cerebrovasc Dis. 26:113-119. Fedarko NS, Jain A, Karadag A, Fisher LW. 2004. Three small integrin binding ligand
N-linked glycoproteins (SIBLINGs) bind and activate specific matrix metalloproteinases. FASEB J. 18:734-736
Fernández CA, Yan L, Louis G, Yang J, Kutok JL, Moses MA. 2005. The matrix
metalloproteinase-9/neutrophil gelatinase-associated lipocalin complex plays a role in breast tumor growth and is present in the urine of breast cancer patients. Clin Cancer Res. 11:5390-5395.
Freedom RM, White RD, Pieroni DR, Varghese PJ, Krovetz LJ, Rowe RD. 1974. The natural history of the so-called aneurysm of the membranous ventricular septum in childhood. Circulation 49:375-384.
Fukazawa M, Fukushige J, Ueda K. 1988. Atrial septal defects in neonates with reference to spontaneous closure. Am Heart J. 116:123-127.
Gai X, Lan X, Luo Z, Wang F, Liang Y, Zhang H, Zhang W, Hou J, Huang M. 2009.
Association of MMP-9 gene polymorphisms with atrial fibrillation in hypertensive heart disease patients. Clin Chim Acta 408:105-109.
Gai X, Zhang Z, Liang Y, Chen Z, Yang X, Hou J, Lan X, Zheng W, Hou J, Huang M. 2010. MMP-2 and TIMP-2 gene polymorphisms and susceptibility to atrial fibrillation in Chinese Han patients with hypertensive heart disease. Clin Chim Acta 411:719-724.
Goor DA, Edwards JE, Lillehei CW. 1970. The development of the interventricular septum of the human heart; correlative morphogenetic study. Chest 58:453-467.
Gu H, Gong J, Qiu W, Cao H, Xu J, Chen S, Chen Y. 2011. Association of a tandem repeat polymorphism in NFATc1 with increased risk of perimembranous ventricular septal defect in a Chinese population. Biochem Genet. 49:592-600.
Han DH, Kim SK, Kang S, Choe BK, Kim KS, Chung JH. 2008. Matrix Metallopeptidase 2 Gene Polymorphism is Associated with Obesity in Korean Population. Korean J Physiol Pharmacol. 12:125-129.
Haq I, Chappell S, Johnson SR, Lotya J, Daly L, Morgan K, Guetta-Baranes T, Roca J, Rabinovich R, Millar AB, Donnelly SC, Keatings V, MacNee W, Stolk J, Hiemstra PS, Miniati M, Monti S, O'Connor CM, Kalsheker N. 2010. Association of MMP-2
polymorphisms with severe and very severe COPD: a case control study of MMPs-1, 9 and 12 in a European population. BMC Med Genet. 11:7.
Helgason H, Jonsdottir G. 1999. Spontaneous closure of atrial septal defects. Pediatr Cardiol. 20:195-199.
Hirose Y, Chiba K, Karasugi T, Nakajima M, Kawaguchi Y, Mikami Y, Furuichi T, Mio F, Miyake A, Miyamoto T, Ozaki K, Takahashi A, Mizuta H, Kubo T, Kimura T, Tanaka T, Toyama Y, Ikegawa S. 2008. A functional polymorphism in THBS2 that affects alternative splicing and MMP binding is associated with lumbar-disc herniation. Am J Hum Genet. 82:1122-1129.
Hoffman JI, Rudolph AM. 1965. The natural history of ventricular septal defects in infancy. Am J Cardiol. 16:634-653.
Hornberger LK, Sahn DJ, Krabill KA, Sherman FS, Swensson RE, Pesonen E, Hagen-Ansert S, Chung KJ. 1989. Elucidation of the natural history of ventricular septal defects by serial Doppler color flow mapping studies. J Am Coll Cardiol. 13:1111-1118
Horwitz AL, Hance AJ, Crystal RG. 1977. Granulocyte collagenase: selective digestion of type I relative to type III collagen. Proc Natl Acad Sci USA 74:897-901.
Hua Y, Song L, Wu N, Lu X, Meng X, Gu D, Yang Y. 2009a. Polymorphisms of MMP-2 gene are associated with systolic heart failure risk in Han Chinese. Am J Med Sci. 337:344-348. Hua Y, Song L, Wu N, Xie G, Lu X, Fan X, Meng X, Gu D, Yang Y. 2009b. Polymorphisms of
MMP-2 gene are associated with systolic heart failure prognosis. Clin Chim Acta 404:119-123.
Ikeda K, Ihara K, Yamaguchi K, Muneuchi J, Ohno T, Mizuno Y, Hara T. 2008. Genetic analysis of MMP gene polymorphisms in patients with Kawasaki disease. Pediatr Res.