ARTICLE NO.RC988272
Physiological Role of the Association Complexes of
a
-Crystallin and Its Substrates on the Chaperone Activity
Jiahn-Shing Lee,* Tatsuya Samejima,† Jiahn-Haur Liao,‡
Shih-Hsiung Wu,‡ and Shyh-Horng Chiou
Ø,‡
,1*Department of Ophthalmology, Chang-Gung Memorial Hospital; ‡Laboratory of Crystallin Research, Institute of Biochemical Sciences, National Taiwan University;Ø
Institute of Biological Chemistry, P.O. Box 23-106, Academia, Taipei, Taiwan; and †Department of Chemistry, College
of Science and Engineering, Aoyama Gakuin University, Tokyo, Japan
Received January 24, 1998
other lens structural proteins such as b- and
g-crys-Previous reports on the chaperone activity of a-crys- tallins (5). The mechanism of this protection may in-tallin to prevent protein denaturation and thermal ag- volve preferential binding of the partially denatured gregation have suggested that partially denatured protein toa-crystallin complex via hydrophobic interac-proteins can bind a-crystallin in its central region.
tion (6-8).
Likewise, b- and g-crystallin can also be localized to
Whether the chaperone-like activity of a-crystallin
the central cavity of a-crystallin particle in vivo, which
has any functional significance in the eye lens remains
provides indirect evidence that a-crystallin can
func-conjectural and speculative.a-Crystallin was shown to
tion as a chaperone in the intact lens. In this study,
inhibit UV-induced protein aggregation with a
chaper-we have further demonstrated that the binding of the
onin effect analogous to that seen in thermal
aggrega-substrate proteins to a-crystallin by short-term
prein-tion experiments (9). It can also decrease the light
scat-cubation may mimic the in vivo conditions of crystallin
tering and thiol oxidation of other crystallins under
association. Preheating of a-crystallin with its
sub-conditions of various oxidative stress (10). Because
UV-strate proteins at 607C for 20 min resulted in the
forma-irradiation and oxidative stress are thought to be
pres-tion of stable complexes between a-crystallin and its
ent under normal physiological conditions and are
con-substrates (8.0% of insulin or 5.3% of g-crystallin was
involved in complex formation). Under such condi- sidered as two major risk factors for cataractogenesis
tions, the chaperone activity of a-crystallin to inhibit (11,12), it is conceivable thata-crystallin may therefore
dithiothreitol-, ultraviolet-, or oxidation-induced pro- contribute to some mechanisms that maintain the lens
tein aggregation can be greatly enhanced. Since UV- in a transparent state in vivo.
irradiation and oxidative stress are common insults to However, the protective ability of a-crystallin, in eye lenses under normal physiological conditions, the terms of stoichiometry, is much lowered in UV-irradia-presence of a/g and a/b complex in vivo may play an tion and oxidative stress than in thermal aggregation important role to maintain the lens in a transparent experiments (1,9,10). Upon heating, a-crystallin may state. q 1998 Academic Press undergo an irreversible conformational transition, with a marked increase in surface hydrophobicity, which correlates well with its increased chaperone activity (8,13). Up to now, we do not know whether such struc-a-Crystallin, the major protein of the ocular lens, is
tural changes actually occur under in vivo conditions. believed to play a prominent role in the maintenance
Nevertheless, it was demonstrated that, by immuno-of the transparency and refractive properties immuno-of the lens
electron microscopy, part ofa-crystallin of water solu-(1,2). It belongs to the family of small heat-shock
pro-ble lens proteins contained either g- or b-crystallin, teins (3), consisting of two types of homologous
sub-bound to the putative chaperone binding site in the units to form a dynamic multimeric complex (4). In
central region of the high-molecular-weight
a-crys-vitro, it can function like a molecular chaperone to
pre-tallin complexes (14,15). This binding is similar to that vent thermal aggregation of a number of enzymes and
seen in vitro whena-crystallin is incubated in the pres-ence of partially denatured forms ofg-crystallin (16).
The presence of the a/gand a/b complexes in vivo
1To whom correspondence should be addressed. Fax:
play a functional role as a molecular chaperone in the intact lens (14). In this paper, we further demonstrate that in vitro short-term preincubation of substrate pro-teins and a-crystallin engenders the crystallin com-plexes similar to those observed under physiological conditions. The protective abilities ofa-crystallin to in-hibit protein denaturation and aggregation associated with dithiothreitol (DTT)-reduction, UV-irradiation, or H2O2-oxidation are found to enhance greatly. Probably through this association mechanism, a-crystallin can provide more protection for other lens proteins against various stress under physiological conditions (9,10). MATERIALS AND METHODS
Lens crystallins were isolated from extracts of bovine lenses using
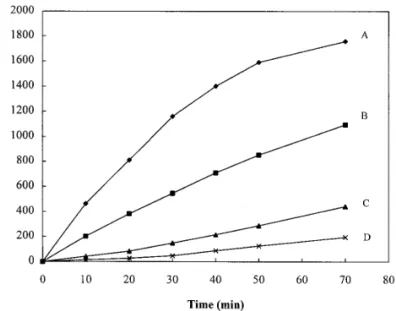
a TSK HW-55(S) gel permeation column as described previously (17). FIG. 1. DTT-induced aggregation of insulin B chains at room The concentration of lens protein was determined by absorbance temperature in the absence and presence ofa-crystallin under native measurements using extinction coefficients of 0.83 and 2.26 at 280 and preheating conditions. (A) insulin B chains with DTT in the nm fora- andg-crystallins, respectively (based on 1 mg/ml). Dithio- absence ofa-crystallin; (B) insulin B chains with DTT in the presence threitol (DTT) was purchased from Research Organics, Inc. (Cleve- of nativea-crystallin; (C) insulin B chains with DTT in the presence land, Ohio). Insulin and other chemicals were of the highest quality ofa-crystallin preincubated at 607C for 20 min; and (D) both insulin and purchased from Sigma Chemical Co. (St. Louis, MO). B chains anda-crystallin preincubated at 607C for 20 min before
assays. Chaperone Activity Assay for DTT-Induced
Aggregation of Insulin B Chain
a-crystallin to inhibit the aggregation of UV-irradiatedg-crystallin. Chaperone activity ofa-crystallin under DTT reduction was
mea-Briefly, 0.2 mg ofg-crystallin, in 1 ml of 0.1 M phosphate buffer, pH sured as described previously (8). This assay measures the ability of
7.0 (in the absence or presence of 0.2 mg ofa-crystallin), was irradi-a-crystallin to protect against the aggregation of insulin B chain in
ated at 295 nm with the excitation bandpass of 20 nm for a fixed time the presence of DTT. Briefly, 0.2 mg of insulin in 1 ml of 0.1 M
period in a Jasco-FP777 spectrofluorometer, and after irradiation the phosphate buffer, pH 7.0 (in the absence or presence of 0.2 mg of
a-excitation and emission of the monochrometer were set at 600 nm crystallin) was reduced with 20 mM freshly prepared DTT and the
with a bandpass of 1.5 nm to measure the relative scattering. Four kinetics of B chain aggregation was followed by measuring the
appar-different conditions were compared in this assay, which include: 1. ent absorption at 360 nm due to light scattering or turbidity
forma-g-crystallin only, 2.g-crystallin and nativea-crystallin, 3. g-crys-tion. The measurements were performed at room temperature using
tallin and preheated a-crystallin, 4. bothg- anda-crystallin were a Jasco 7800 UV/VIS spectrophotometer and a 1 cm pathlength
cu-preheated. The preheated samples were carried out at 607C for 20 vette. Four different conditions were compared in this assay, which
min and then allowed to equilibrate at room temperature for 12 h include: 1. insulin only, 2. insulin and nativea-crystallin, 3. insulin
prior to experiments. and preheateda-crystallin, 4. both insulin anda-crystallin were
pre-heated. The preheated samples were carried out at 607C for 20 min
Chaperone Activity Assay for Oxidation-Induced and then allowed to equilibrate at room temperature for 12 h prior
to experiments. Aggregation ofg-Crystallin
Chaperone activity ofa-crystallin under oxidative stress was mea-Gel-Permeation HPLC for Analysis of Substrate sured similar to a previous report (10). This assay measures the
Binding ability of a-crystallin to protect against the aggregation of
g-crys-tallin under oxidation. Briefly, 0.5 mg ofg-crystallin in 1 ml of 0.1 A Jasco Familic-300S HPLC pump, a UVIDEC-100V detector, and
M phosphate buffer, pH 7.0 (in the absence or presence of 0.5 mg of a Hitachi D2500 Chromato-Integrator were used in conjunction with a-crystallin) was incubated with 0.5 mM sodium ascorbate, 0.1 mM a SynChropak GPC 300 (250 1 4.6 mm) size exclusion column for FeCl
3, 0.3 mM EDTA, and 1 mM H2O2. The kinetics ofg-crystallin
analysis of protein interaction witha-crystallin. The mobile phase aggregation was followed by measuring the apparent absorption at was 0.1 M sodium phosphate pH 7.0 with an isocratic flow rate of
360 nm due to light scattering (similar to that used in assays for DTT-0.25 ml/min. Absorbance was monitored at 280 nm.a-Crystallin was
induced aggregation). Four different assay conditions were compared mixed with insulin org-crystallin and analyzed at room temperature similar to those described for UV-induced aggregation ofg-crystallin. before and after incubation at 607C for 20 min. The samples after
thermal incubation were also allowed to equilibrate at room
tempera-RESULT ture for at least 12 h. The elution profiles of mixtures with or without
heating were compared regarding relative changes in area for each
peak. In Fig. 1, upon reducing the disulfide bond between
the two chains of insulin with reducing agent DTT,
Chaperone Activity Assay for UV-Induced the B chain of insulin aggregates and precipitates out
Aggregation ofg-Crystallin (curve A), leaving the A chain in solution (8). Also
shown in curve B of Fig. 1, this process can be sup-Chaperone activity ofa-crystallin under UV-irradiation was
ag-gregation-prone protein (19). The extent of the protec-tion for B chain from aggregaprotec-tion depends on the rela-tive concentration of insulin and the chaperone, with a complete prevention of aggregation at a weight ratio of a-crystallin to insulin of more than 6:1 (19). In Fig. 1 (curve C), the preheateda-crystallin was shown to be more potent in suppressing the aggregation of insulin B chain than the native one. This increased chaperone activity of the preheated a-crystallin was explained previously by the increase in its surface hydrophobicity as assessed by the ANS (1-anilino-8-naphthalene sulfo-nate) binding assay (8,17,20). In the present study, we have further found that if both insulin anda-crystallin were preincubated at the same condition as that of preheated a-crystallin, the extent of apparent absorp-tion due to light scattering would be much more de-pressed (curve D).
Preincubation of both insulin and a-crystallin may lead to heat-induced interaction between these two pro-teins and thermal perturbation upon either one respec-tively. The temperature-induced changes of insulin or a-crystallin alone can not explain the difference be-tween curves C and D. In order to explore the interac-tion of insulin anda-crystallin under thermal incuba-tion, we have thus performed and compared gel-perme-ation HPLC of these two proteins before and after heating.
The gel-permeation HPLC analysis of the mixture of a-crystallin (0.2 mg/ml) and insulin (0.2 mg/ml) is shown in Fig. 2A. Similar result of the elution profile
for the mixture ofa-crystallin (0.5 mg/ml) andg-crys- FIG. 2. Gel permeation HPLC ofa-crystallin and substrate pro-teins, (A)a-crystallin plus insulin, (B)a-crystallin plusg-crystallin. tallin (0.5 mg/ml) is shown in Fig. 2B. Previously
a-The sample proteins were preheated at 607C for 20 min (dotted line) crystallin was shown to act as a chaperone interacting
and without preheating (solid line). with other proteins to prevent heat-induced protein
in-solubilization (5). Both mixtures consisting of the stud-ied proteins before and after incubation at 607C for 20
min respectively, showed little change in light scatter- cal stress, we have performed similar experiments against UV-irradiation and H2O2-oxidation using g-ing and no apparent pellet formation after
centrifuga-tion. However, comparison of the elution profiles of the crystallin, instead of insulin, as substrate. In Fig. 3, UV-irradiation ofg-crystallin leads to aggregation and mixtures clearly indicate that there is complex
forma-tion between a-crystallin and insulin or g-crystallin this process can be suppressed bya-crystallin (compare curves A and B of Fig. 3). Similar to previous reports after heating. The patterns of unheated mixtures (solid
lines in Fig. 2) give the identical elution profiles as (13,17), the preheated a-crystallin exhibits a higher chaperone activity than the native one to protect g-those based on independent runs with each individual
protein alone. However after heating at 607C for 20 crystallin against photoaggregation (curve C of Fig. 3). However, if the preincubation included both a-crys-min (dot lines in Fig. 2), the elution time of the first
peaks decrease and the area increases, indicating the tallin and g-crystallin, the extent of UV-induced pro-tein aggregation decreased most (curve D of Fig. 3). formation of complex between native a-crystallin and
substrate protein. The areas of the second peaks de- In Fig. 4, we have also confirmed a previous report (10) thata-crystallin can act as a chaperone under oxi-crease by about 8.0% and 5.3% for insulin and
g-crys-tallin respectively. dative stress, decreasing the light scattering of g-crys-tallin (curves A and B of Fig. 4). Furthermore, the pre-The short-term incubation of protein mixtures
cre-ates a condition that part of the aggregation-prone sub- heateda-crystallin showed a higher ability to protect g-crystallin against oxidative stress (curve C of Fig. 4) strates are bound toa-crystallin, which mimics the in
vivo condition ofa-crystallin. To test whether such en- even though the effect is not so prominent like that under UV-irradiation (curve C of Fig. 3). We have again hanced chaperone-like activity under DTT-induced
ity to protect against heat-induced aggregation of other proteins. It is apparent that the limited amounts of binding site may enable only a fixed number of partially denatured proteins to bind to thea-crystallin particles. In the present study, using a fixed number of substrate proteins, we have otherwise demonstrated that pre-binding of substrate proteins to a-crystallin showed higher chaperone efficiency against various types of in-sults. It is possible that the entrapped substrate pro-teins become more resistant to UV-irradiated and oxi-dative stress or the decreased concentration of the un-bound aggregation-prone peptides makes the stress-induced aggregation less prominent.
Although the binding pattern of b- or g-crystallin to a-crystallin in vivo may be similar to that of in vitro by thermal incubation, it is physiologically impossible that such high temperature (607C) exist in vivo to account for
FIG. 3. UV-irradiated aggregation ofg-crystallin at room
tem-the formation ofa/ganda/bcomplexes in the intact lens. perature in the absence and presence ofa-crystallin under native
One of the alternative mechanisms to account for these and preheating conditions. (A) UV-irradiation ofg-crystallin in the
complex formation is mediated by UV-irradiation. a-absence ofa-crystallin; (B) UV-irradiation ofg-crystallin in the
pres-ence of nativea-crystallin; (C) UV-irradiation ofg-crystallin in the Crystallin can bind with its substrate under UV-irradia-presence of a-crystallin preincubated at 607C for 20 min; and (D) tion, however the reaction intermediates are different in bothg-crystallin anda-crystallin preincubated at 607C for 20 min
UV-irradiated versus heat-induced aggregation. UV irra-before assays.
diation was shown to generate covalently linked aggre-gates which were not dissociated by SDS and reducing agents, whereas heat-induced aggregates could be easily its ability to preserve solution transparency, is the dissociated under the same conditions (9). Another possi-highest (curve D of Fig. 4) when both a-crystallin and bility for the formation of these complexes is through the g-crystallin are preincubated at 607C for 20 min. process of oxidative stress. However, the interaction ofa/ gcomplex formed from in vitro oxidation was too weak to DISCUSSION be isolated with procedures used to obtain the thermally
induced complexes (10). There is a considerable interest in the functional role
ofa-crystallin, especially after it was shown to have in
vitro chaperone activity (5). However, the mechanism
of this chaperone action of a-crystallin in the intact lens is still not very clear. Previous heat denaturation assays (16,21) have demonstrated thata-crystallin can preferentially bind to partially denatured proteins via a central region of the complex. In this process, the polypeptide is protected against further denaturation (16). In vivo,b- andg-crystallins can also be localized to the central region of thea-crystallin particle, strongly suggesting that thea-crystallin can function as a mo-lecular chaperone in the intact lens (14). It was esti-mated that 3.3% and 3.7% ofa-crystallin in lens soluble proteins was bound byb- andg-crystallin respectively, and a higher percentage of binding (7.8% and 9.0%) was noted in the high-molecular-weight aggregate (HMW-a) fraction (15).
However, it was reported that binding of partially FIG. 4. Oxidation-induced aggregation of g-crystallin at room denatured protein toa-crystallin particles results in a temperature in the absence and presence ofa-crystallin under native and preheating conditions. (A) oxidation-induced aggregation of g-subsequent decrease in the ability of the a-crystallin
crystallin in the absence ofa-crystallin; (B) oxidation-induced aggre-to protect against further protein denaturation (22).
gation ofg-crystallin in the presence of nativea-crystallin; (C) oxida-In that study, a constant amount of a-crystallin was tion-induced aggregation ofg-crystallin in the presence of a-crys-preincubated with increasing amounts ofg-crystallin, tallin preincubated at 607C for 20 min; and (D) bothg-crystallin and
a-crystallin preincubated at 607C for 20 min before assays. and the resulting complexes were tested for their
abil-It remains enigmatic howa/ganda/bcomplexes are ACKNOWLEDGMENTS
formed in vivo. However as shown in this study the This work was supported by Academia Sinica and the National formation of such complexes may afford higher effica- Science Council (NSC Grants 83-0203-B-001-086, 83-0418-B-001-cies of protection to preserve the ocular lens in a trans- 020BA, 84-2311-B-001-050-BA, and 86-2311-B-002-031-B15), Taipei,
Taiwan. parent state against various physiological stress. Since
the ocular lens is constantly exposed to visible and UV
REFERENCES light irradiation and there is a significant amount of
1. Horwitz, J. (1993) Invest. Ophthalmol. Vis. Sci. 34, 10 – 22. H2O2(30mM) in the aqueous humor of the normal
hu-2. Groenen, P. J. T. A., Merck, K. B., de Jong, W. W., and Bloemen-man eye (23), it is reasonable to assume that an
effi-dal, H. (1994) Eur. J. Biochem. 225, 1 – 19. cient chaperone-like protective action against
photo-3. Ehrnsperger, M., Graber, S., Gaestel, M., and Buchner, J. (1997) chemical and oxidative insults indeed exists in vivo. In Curr. Eye Res. 16, 221 – 229.
this contexta-crystallin may be the best candidate for 4. van der Quderaa, F., de Jong, W., Hilderink, A., and Blomendal, mediating such a protective role in the lens because H. (1974) Eur. J. Biochem. 49, 157 – 168.
the molecular chaperone activity of a-crystallin have 5. Horwitz, J. (1992) Proc. Natl. Acad. Sci. USA 89, 10449 – 10453. 6. Boyle, D., and Takemoto, L. (1994) Exp. Eye Res. 58, 9 – 16. been correlated with the extents of posttranslational
7. Raman, B., and Rao, C. M. (1994) J. Biol. Chem. 269, 27264 – modifications, such as UV-irradiation (17) and
oxida-27268. tion (24).
8. Das, K. P., and Surewicz, W. K. (1995) FEBS Lett. 369, 321 – In this study, we did not include b-crystallin as a 325.
substrate, mainly because its oxidation at room tem- 9. Borkman, R. F., Knight, G., and Obi, B. (1996) Exp. Eye Res. 62, perature produced only a slight increase in light scat- 141 – 148.
tering, in contrast to those observations obtained with 10. Wang, K., and Spector, A. (1995) Invest. Ophthalmol. Vis. Sci. 36, 311 – 321.
g-crystallin (10). On the other hand, the preincubation
11. Spector, A., and Garner, W. H. (1981) Exp. Eye Res. 33, 673 – under higher temperatures and/or for longer periods
681. may increase the amounts of complex formation
be-12. Walker, M. L., and Borkman, R. F. (1989) Exp. Eye Res. 48, 375 – tween a-crystallin and its substrates. However, we 383.
have found that a-crystallin could not provide long- 13. Das, B. K., Liang, J. J. N., and Chakrabarti, B. (1997) Curr. Eye term heating (ú 30 min) protection against thermally Res. 16, 303 – 309.
14. Boyle, D., and Takemoto, L. (1994) Exp. Eye Res. 58, 9 – 16. induced aggregation of g-crystallin, and a/g complex
15. Takemoto, L., and Boyle, D. (1994) Curr. Eye Res. 13, 35 – 44. may gradually become insoluble at temperatures
16. Boyle, D., Gopalakrishnan, S., and Takemoto, L. (1993) Biochem. higher than 607C. Nevertheless, it is interesting to note
Biophys. Res. Commun. 192, 1147 – 1154.
that such short-term preincubation as shown in this
17. Lee, J.-S., Liao, J.-H., Wu, S.-H., and Chiou, S.-H. (1997) J. study gave the percentage of a/g complex formation Protein Chem. 16, 283 – 289.
very close to that seen in vivo (15). 18. Lee, J.-S., Satoh, T., Shinoda, H., Samejima, T., Wu, S.-H., and In conclusion, we have demonstrated that the prein- Chiou, S.-H. (1997) Biochem. Biophys. Res. Commun. 237, 277 –
282. cubation ofg- anda-crystallin mixtures at 607C for 20
19. Farahbakhsh, Z. T., Huang, Q. L., Ding, L. L., Altenbach, C., min resulted in the formation of a/g complex, which
Steinhoff, H. J., Horwitz, J., and Hubbell, W. L. (1995) Biochem-may mimic the formation of such complexes in vivo. istry 34, 509 – 516.
Under such conditions of complex formation, the chap- 20. Raman, B., Ramakrishna, T., and Rao, C. M. (1995) FEBS Lett. erone-like activity of a-crystallin against UV-irradia- 365, 133 – 136.
tion or H2O2-induced oxidation on lens proteins was 21. Rao, P. V., Horwitz, J., and Zigler, J. S., Jr. (1993) Biochem.
Biophys. Res. Commun. 190, 786 – 793.
shown to be more effective than that of native or
pre-22. Takemoto, L., and Boyle, D. (1994) Arch. Biochem. Biophys. 315, heated a-crystallin. It is apparent that in addition to
133 – 136. the inherent chaperone activity of a-crystallin, some
23. Ramachandran, S., Morris, S. M., Devamanoharan, P., Henein, other factors which induce the formation ofa/ganda/ M., and Varma, S. D. (1991) Exp. Eys. Res. 53, 503 – 506. b complexes in vivo may also work in a cooperative 24. Cherian, M., and Abraham, E. C. (1995) Biochem. Biophys. Res. manner to protect the lens against physiological stress. Commun. 208, 675 – 679.