台灣金線連有效分劃抑制thioacetamide誘發小鼠肝臟纖維化; Inhibitory effect of Anoectochilus formosanus effective fraction on thioacetamide- induced liver fibrosis in mice
72
0
0
全文
(2) 台灣金線連有效分劃抑制 thioacetamide 誘發小鼠肝臟纖維化 莊謹如 摘. 要. 本研究主要探討台灣金線連有效分劃(AFEF)對 thioacetamide (TAA) 所誘發小鼠肝臟纖維化的效果。BABL/c 雄性小鼠隨機分成四 組,即控制組、TAA 組及 AFEF 兩組。除了控制組,所有小鼠每週 三次由腹腔注射 TAA (200 mg/kg body weight),持續八週。AFEF 組 小鼠的整個實驗過程,每天一次經口投予 AFEF (0.2 或 1.0 g/kg body weight)。TAA 誘發小鼠肝纖維化,最終的血清 GPT 活性上升、肝臟 hydroxypoline 含量增加、肝臟相對重量增加、血清膽固醇含量下降。 AFEF 之 處 理 能 降 低 血 清 GPT 活 性 、 肝 臟 相 對 重 量 及 肝 臟 hydroxypoline 含量,增加血清膽固醇含量。肝臟組織學的分析也顯示 有改善作用。RT-PCR 的分析顯示 AFEF 能降低 TAA 處理的肝臟 collagen ( α1 )( I )、 insulin-like growth factor binding protein 及 lipopolysaccharidebinding protein mRNA 表現。結論,小鼠口服 AFEF 能減弱 TAA 所誘發的肝臟纖維化,可能是 AFEF 具有抗肝臟發炎的 作用而保護肝細胞不會壞死。. 1.
(3) Inhibitory effect of Anoectochilus formosanus effective fraction on thioacetamide- induced liver fibrosis in mice Jin-Ru Chuang Abstract To investigate the effects of Anoectochilus formosanus effective fraction (AFEF) on liver fibrosis induced by thioacetamide (TAA) in mice. Male BABL/c mice were divdied randomely into 4 groups: control, TAA, and two AFEF groups. Except for mice in control group, all mice were administered intraperitoneally with TAA (200 mg/kg body weight) three time a week for 8 weeks . Mice in AFEF groups were treated daily with AFEF (0.2 or 1.0 g/kg body weight) via gastrogavage throught the whole experimental period. TAA caused liver fibrosis, featuring increase in serum GPT activity, hepatic hydroxyproline content, and liver relative weight; and decrease in serum cholesterol level. Compared with TAA group, AFEF treatment significantly decreased the activity of GPT, liver relative weight and hepatic hydroxyproline content; but increased serum cholesterol concentration. Liver histology in the AFEF-treated mice was also improved. RT-PCR analysis showed that AFEF treatment decreased the expression of collagen (α1)(I), lipopolysaccharide binding protein and insulin-like growth factor binding protein. In conclusion, oral administration of AFEF significantly reduces TAA-induced hepatic fibrosis in mice, probably by exerting a protective effect against hepatocellular necrosis by its anti-inflammatory ability.. 2.
(4) 致 謝 辭 感謝恩師 林教授文川博士之悉心教導與啟發,充實我的專業知 識,並引導我以積極、謹慎的態度面對任何人、事、物。本論文於撰 稿之初,感謝恩師您逐字詳細的閱讀與修正,才使得論文得以更完 整。師恩浩瀚,永銘於心,謹誌卷首致上由衷的謝意。 感謝藥理科蔡輝彥老師、陳玉芳老師、吳介信老師、謝文聰老師、 譚思濰老師及柯毅文助理,於就學期間給予的指導與協助。感謝實驗 室焄蓮學姊、岳文學長、宏柏學長於實驗及課業上的教導與鼓勵,謝 謝小嬋學妹、宛臻學妹、孟宗學弟、蓓諭、及錦芳於實驗上及生活上 的關心與陪伴。 感謝臺大醫院雲林分院全體藥師,在我就學期間給予支持與鼓 勵,更由衷感謝藥劑部廖玲巧主任在我面對工作與學業抉擇時,提供 寶貴的意見,並給予支持、關心及照顧。感念諸位同事的厚愛,在此 限上最深的謝意。 兩年的求學過程,感謝家人對我的支持與鼓勵,我將此論文獻給 關心我、支持我的父親 莊塭溪先生及母親 王淑美女士,還有在我求 學時,不辭辛苦接送我的浩賢,謝謝他們無所求的付出及支持,讓我 無後顧之憂的完成學業。. 3.
(5) 目 第一章. 緒. 第一節 第二節. 錄. 論. 頁次. 研究背景及目的…………………………………..…..…10 金線連文獻考察…………………………….…………12. 一、 金線連種原、分布及植物型態…………………...….12 二、 民間入藥用法…….………………………...…..……..13 三、 金線連成分………………..………………….……….13 四、 金線連藥理活性…………………..……………..……14 第三節. 肝纖維化成因……………………………..……..….....18. 一、 星狀細胞(Hepatic stellate cell)………..…...……..18 二、 膠原蛋白……………………………………….…...…20 三、 間質金屬蛋白酶 (Matrix metalloproteinase, MMP) 與組織金屬蛋白酶抑制劑 (Tissue inhibitors of metalloproteinases, TIMP)………………..…......…21 四、 脂多糖結合蛋白 (Lipopolysaccharide binding protein)………………………………………………….22 五、 轉化生長因子 (Transforming growth factor β1)…..23 六、 胰島素樣生長激素結合蛋白(Insulin-like growth factor binding protein)……………………………….24 七、 肝細胞生長因子(Hepatocyte growth factor)……26 4.
(6) 第四節 第二章. Thioacetamide 誘導肝纖維化……………….….……..27 研究材料與方法. 第一節 研究材料………………………………………………..28 第二節 研究方法………………………………………………..30 一、 AFEF 的製備…………………………….………….…30 二、 動物實驗……………………………………….…..…..30 三、 血液生化值實驗…………………………………..…...30 四、 肝臟組織病理學觀察……………………………..…...31 五、 肝臟 Hydroxypoline 含量測定…………………….....31 六、 RT-PCR 分析………………………………..……..….32 七、 Real time PCR 分析…………….………………....….32 八、 統計方法………………..………………………..…….32 第三章. 研究結果. 一、 對體重的影響………………………….……..………...34 二、 對肝臟重量及含水量影響………………………….….34 三、 對血清生化值影響……………………..……..…….….35 四、 Hydroxylpoline 含量………………..………………....35 五、 病理檢驗…………………………………….………….35 六、 RT-PCR 結果…………………………….………….....36. 5.
(7) 七、 Real-time PCR 結果…………………..……………...37 第四章. 討論與結論…………………………………………….53. 參考文獻…………………………………………………….…....59. 6.
(8) 表目錄 Table 1. Oligonuleotide sequences used in RT-PCR and Real-time PCR reactions…………………………………………………....38 Table 2. The changes of body weight of mice………………..…….....39. 7.
(9) 圖 目 錄 Fig. 1. Effect of AFEF on the absolute weight of liver in TAAtreated mice……………………………………………….….40 Fig.2. Effect of AFEF on the relative weight of liver in TAA-treated mice……………………………………..………………..….41 Fig.3. Effect of AFEF on the water content( ﹪) in TAA-treated mice………………………………………..…………..….…42 Fig.4. Effect of AFEF on the serum activities of GPT in TAA-treated mice………………………………………..…………..…….43 Fig. 5. Effect of AFEF on the concentration of serum cholesterol in TAA-treated mice……………………………………..….….44 Fig. 6. Effect of AFEF on hepatic hydroxypoline contents in TAA-treated mice…………………………………..………………..……45 Fig. 7. The photomicrographs of liver section taken from mice and stained with Hematoxylin-eosin……………………..……....46 Fig. 8. The photomicrographs of liver section taken from mice and stained with Sirus red………………………………….…….47 Fig. 9. RT-PCR analysis of Collagen (α1)(I), Tgfβ1 and MMP13 expressions in mice………………………………………….48 Fig. 10. RT-PCR analysis of LBP, CD14 and TLR4 expressions in mice……………………………………..……………..…….49 Fig. 11. RT-PCR analysis of IGF-I, IGF-R, IGFBP-1 and IGFBP-3 expressions in mice……………………………………….…50 Fig. 12. RT-PCR analysis of HGF and c-Met expressions in mice………………………………..…….………….……….51 8.
(10) Fig.13. Real Time PCR analysis of IGFBP-1 and LBP levels in TAAtreated mice liver tissue………………………….………..52. 9.
(11) 第一章 緒 第一節. 論. 研究背景及目的. 根據流行病學統計,在台灣每年因肝病致死者約有一萬人,其中 肝硬化約四千人,肝癌約五千人,其餘肝病約一千人。在十大癌症中, 肝癌更是高居首位;肝硬化也居十大死因之第六位。因此肝病在台灣 是相當普遍而且嚴重的疾病,所以肝病又有”國病”之稱。 近年來衛生單位極力推廣「預防重於治療」的觀念,因而「健康 食品」逐漸受到社會大眾的重視。自從民國八十八年二月三日總統令 公佈「健康食品管理法」,同年八月三日該法正式執行後,健康食品 的開發如雨後春筍般的蓬勃發展,市面上對於保肝、療肝的健康食品 亦不勝枚舉,本研究室從事台灣金線連保健功效的研究多年,確認其 具有保肝作用,投與台灣金線連水粗萃取物可以改善四氯化碳所引起 的 慢 性 肝 損 傷 (Shih et al., 2005) , 台 灣 金 線 連 水 粗 萃 取 物 對 dimethylnitrosamine(DMN)誘發大鼠肝纖維化後,所造成的肝臟萎 縮、脾臟腫大、血清白蛋白及肝臟膠原蛋白的作用是有利的,且台灣 金線連水粗萃取物對於 DMN 肝損傷後切肝再生,可以顯著增加肝臟 重量再生率(Shih et al,2004)。我們繼而利用四氯化碳誘發小鼠肝纖 維化的實驗模式,分離製備出台灣金線連保肝有效分劃 (Anoectochilus formosanus effective fraction;AFEF)。此分劃在工業. 10.
(12) 上已可量產,其品管方法已建立,技術也已轉移給業界,已有相關產 品在販售。 TAA 是近年來常用的肝纖維化誘導藥物作用,已有蛋白質體學 的研究,TAA 的肝毒性及誘發肝纖維化的機轉已有相當瞭解(Low et al., 2004)。本研究之目的是探討 AFEF 對 thioacetamide (TAA)誘發小 鼠慢性肝炎的效果,並利用 RT-PCR 的技術,分析肝臟相關基因的表 現來說明 AFEF 保肝可能的作用機轉。. 11.
(13) 第二節. 金線連文獻考察. 一、金線連種原、分布及植物型態 台灣金線連學名 Anoectochilus formosanus HAYATA,別名藥王、藥 虎、金蠶、烏蔘、石松、金石松、本山石松、虎頭蕉、金線屈腰、金 線蕨龍、金線連、樹草連、雉雞草、金錢草和黃花粘子蘭。台灣金線 連為蘭科(Orchidaceae)金線連屬(Anoectochilus)之小型地生蘭,屬多年 生藥用植物,分佈於台灣全島海拔約 500-1500 公尺的闊葉林床(林, 1994;周,1983;劉,1987)。其植物型態株高 20-30 公分,花序高 可達 14.5-26.5 公分;主根肥厚,長約 4-5 公分,不定根數平均 3-4 條; 莖基部直徑 0.28-0.5 公分,匍匐圓筒狀,肉質;葉互生;全株葉片數 約八枚,成株則剩約 4-6 片,全缘或具為波浪狀、卵型或卵圓形,最 大葉長約 4.0 公分,寬約 2.5-3.2 公分,銳頭,基部較圓,葉片絨布狀, 綠中兼具金黃色;葉脈中只有中肋脈較明顯,葉面上舖有精美網狀支 脈,葉背泛紅色;花數少,頂生有 3-4 朵之穗狀花序,花序柄紅褐色, 有白色柔毛,柄上具 2-3 片披針型鞘狀苞;萼片數約 3 個,微紅色, 外密披柔毛,倒卵圓形長約 0.6 公分,寬約 0.5 公分;唇瓣長伸,已 相當特化,外型與花瓣迥異,基部有一短距長約 0.5 公分,懼內有二 腺體,唇瓣前端裂成 2 尖瓣,每片長約 0.6 公分,其中段爪部有細絲 狀構造,兩邊各約 6 條,長短不一,外型似梳子;花柱較短,雙柱頭,. 12.
(14) 花藥帽為卵型,花粉塊微黃色,粉質。每年九月抽苔,十月至十一月 開花,未授粉的小花可持續到十二月底才凋謝(甘,1979;劉,1987; 蘇,1975)。. 二、民間入藥用法 「台灣植物藥材誌」及「藥用植物學」記載,台灣金線連味甘性 涼,全草皆可入藥,其藥效十分廣泛,為台灣極為珍貴之民間藥材 (甘,1965、1986) ; 「綱目拾遺」亦述虎頭蕉性熱,治血淋、白帶, 一切吐血。民間亦常用於滋養強壯、清涼退火、涼血固肺、袪傷解鬱 及開中氣,主治肺病、遺精、遺漏、腎虧、高血壓、蛇傷及小兒發育 不良(甘等,1983;謝及李,1983;蔡,1986)。. 三、金線連成分 台灣金線連所含之營養成分除水分含量約 87﹪外,還有脂質 2.7 ﹪、蛋白質 2.2﹪、醣類 6.4﹪、灰份 10﹪、粗纖維 0.7﹪及維生素, (劉等,1987);另外,台灣金線連中亦含有 Ca、P、K、Na 及 Ma 等礦物質,還含有 Fe、Mn、Zn 及 Cu 等微量金屬,其所含的維生素 B、C、E、菸鹼酸及胡蘿蔔素亦豐富。Anoectochilus formosanus 的化 學成分主要為配醣體、固醇類和醣類,及有效之藥理成份如天麻素 gastrodin,kinsenoside (Ito et al., 1993),quercetin、dihydroquercetin、 13.
(15) isorhamnetin、kinsenone (Wang et al., 2002)。. 四、金線連藥理活性 (一) 心血管作用 目前對於金線連藥理的研究顯示,台灣金線連具有明顯鎮痛、降 血糖、利尿、降血壓、細胞貪食及消炎作用(梁,1990);而以組織 培養培育的金線連萃取液研究中發現,金線連萃取液具有抑制血小板 中 前 列 凝 素 (thromboxanes) 合 成 和 促 進 動 脈 內 皮 組 織 中 前 列 環 素 (prostacyclin)的產生,主要是透過抑制前列凝素合成酵素及活化環化 加氧酵素(cyclooxygenase)所產生的作用,且與濃度成現正相關,此研 究結果也顯示金線連對於心血管疾病具有潛在的治療效果(Hung, 1991)。. (二) 降血脂及抗氧化作用 金線連甲醇抽出物中分離出的有效萃取物對高血脂大鼠有改善 作用(Takeshita, 1995)。亦有研究證實台灣金線連具有抗氧化作用,可 以清除自由基和抑制低密度脂蛋白(LDL)的氧化(Wang et al., 2002; Shih et al., 2003;Chen et al., 2004)。可見金線連對於動脈粥狀硬化所 引起的各種疾病具有預防及治療的效果。金線連水萃物也可以保護 HL-60細胞因H2O2產生的氧化壓力所造成凋亡(Wang, 2005)。 14.
(16) (三) 降血糖 金線連甲醇抽出物中分離出的有效萃取物對大鼠血糖上升有抑 制作用(Takeshita., 1995)。金線連能抑制腎上腺素促進糖元的分解, 而且對於四氧嘧啶引起的小鼠血糖升高具有顯著的防治作用(陳, 2000)。對streptozotocin誘發高血糖大鼠,金線連據降血糖作用,可 降低使血清糖化蛋白(serum fructosamine),且抗氧化酵素有顯著性的 回升(Shih et al., 2002)。. (四) 調節免疫作用 近期的研究亦證實,台灣金線連所含的多糖具有重要的免疫功能 (楊,2001)。利用皮下移植方式將CT-26癌化細胞移植到BABL/c小 鼠體內發現,金線連水萃取物可促進巨噬細胞的存活率及吞噬作用, 及增加脾臟細胞的增殖作用(Tseng et al., 2006)。本實驗室亦證實鼷鼠 口服台灣金線連多醣分劃二週後,其腹腔巨噬細胞作用明顯上升,體 外實驗也顯示該多糖分劃能活化RAW264.7巨噬細胞的吞噬作用及釋 出NO、IL-2及γ干擾素(徐,2004) ;其可能的機轉推測金線連本身的 多糖成分會和吞噬細胞表面的接受器結合,造成內在訊息傳導並導致 NO的生成量增加,繼而影響到細胞骨架上蛋白質的變化,使得吞噬 細胞生成偽足而增加吞噬活性(廖,2005);另外,藉由誘導氣喘小 15.
(17) +. +. 鼠模式,我們亦證實AFEF對於免疫抑制作用可能透過CD25 /CD4 T 細胞作用的影響而達免疫抑制效果(蕭,2005). (五) 抗癌作用 以cDNA的微陣分析技術與生物資訊探討台灣金線連萃取液的研 究發現,台灣金線連萃取液可藉由調控乳癌細胞基因的表現,誘引乳 癌細胞產生凋亡及抑制其生長(徐,2001;Yang et al., 2004)。利用 皮下移植方式將CT-26 Colon cancer cell 移植到BABL/c小鼠體內發 現,金線連水萃取物可以抑制腫瘤生長(Tseng et al., 2006)。. (六) 抗疲勞作用 日本學者利用游泳方式,測試小鼠忍耐力,發現金線連萃取物明 顯降低脂肪堆積,推測金線連萃取物可以增加體內對於脂肪的運用 (Ikeuchi, 2005)。. (七) 保肝作用 在保肝作用的研究中,台灣金線連水抽出物可顯著降低 CCl4 所 引起的 GOT、GPT 急速上升,並顯著改善因 CCl4 所導致中心靜脈周 圍之病變(Lin et al., 1993;Lin et al., 2000)。日本學者也指出,台灣金 線連水萃物可保護大鼠初代肝細胞對抗四氯化碳引起的損傷(Du et 16.
(18) al., 2003)。本實驗室研究也顯示,台灣金線連水萃物能不但可抑制四 氯化碳引起的肝臟纖維化 (Shih et al., 2005),對於 DMN 所造成的肝 損傷,有促肝臟再生的作用(Shih et al., 2004)。. (八) 改善學習記憶力 蘭科的天麻所含的天麻素(gastrodin)可治療癲癇等神經症狀,並 具有增強學習記憶的能力(Hsieh et al., 1993)。日本學者也從高雄金線 連成功純化出天麻素(Ito et al., 1993),利用HPLC的比對,我們亦確認 台灣金線連水萃物亦含有相當量的天麻素及天麻元(gastrodigenin) (江,2003),並證實台灣金線連水萃物確能減弱東莨菪鹼誘發的健 忘作用(Cheng et al., 2003)。. (九) 抗骨質疏鬆 本實驗室亦利用金線連水萃物餵食去卵巢大鼠,實驗結果發現, 金 線 連 水 萃 物 有 效 增 加 大 鼠 重 量 及 減 少 血 清 膽 固 醇 與 alkaline phosphatase含量,並有效改善去卵巢大鼠之腦下垂體減輕、右腿骨及 第四腰椎骨的骨密度及鈣含量。證實台灣金線連水萃物有可改善去卵 巢引起之骨質疏鬆症(Shih et al., 2001)。. 17.
(19) 第三節 肝臟纖維化的成因 慢性肝臟疾病例如:病毒性 B、C 型肝炎或非酒精性脂肪肝,最 後會造成肝纖維化的產生(Poynard, 1997、2000;Brunt, 2004)。肝臟 纖維化即細胞外間質(extracellular matrix)過度沉積。 慢性肝炎導致肝纖維化的形成其機轉非常複雜,目前所瞭解的 是多種造成肝損傷的因子引起肝細胞損傷、壞死、凋亡及肝組織炎 症反應,激活 Kupffer 細胞分泌多種細胞因子;以及肝細胞、血小 板及竇內皮細胞等分泌的細胞因子、脂質過氧化等化學媒介物質共 同作用於肝星狀細胞(Hepatic stellate cell),使之激活轉變成肌纖維母 細胞,經由旁分泌及自分泌的機轉,使纖維母細胞增殖,合成大量 的膠原及蛋白多糖等細胞外基質成分(Tsukada et al., 2006 )。. 一、星狀細胞(Hepatic stellate cell) 早期認為肝纖維化是一不可逆的過程(Popper., 1970;Schaffner, 1968),1976 年一份臨床的研究報告指出,肝纖維化是一具有潛在可 逆的特性的過程(Soyer et al., 1976)。肝臟星狀細胞被發現後,肝纖維 化可逆的特性才被重視(Friedman et al., 1985)。 肝臟星狀細胞 , 又稱為 Ito cell、lipocyte、hepatic pericytes、 fat-storing、vitamin A-rich cell 等。位於肝細胞和內皮細胞之間的迪氏. 18.
(20) 腔中,星狀細胞約佔肝臟非實質細胞(non-parenchymal cells)的三分之 一,或肝臟中細胞總數的 15﹪。星狀細胞在正常肝臟具有許多重要 功能,包括:(1) retinoids 的代謝;(2) 細胞激素(cytokine)的分泌,例 如 : 表 皮 生 長 因 子 (epidermal growth factor) 、 肝 細 胞 生 長 因 子 (hepatocytes growth factor)等,這些因子可協助維持肝細胞的質與量; (3) 調控血流,由於星狀細胞所在位置接近神經,故星狀細胞可能受 到神經的刺激而收縮。 星狀細胞的活化主要因肝臟環境的改變,例如:庫氏細胞、內皮 細胞活化釋放激素,或活性氧化物、氧化壓力增加…等,星狀細胞一 旦活化,便會由休止(quiescent)狀態,轉變為活化態的星狀細胞。起 始活化期的星狀細胞會產生更多活性氧化物刺激第一型膠原蛋白基 因表現並減弱抗氧化劑的作用,導致慢性肝損傷。當活化態星狀細胞 進入永久期後,便開始一連串不同於休止態的特徵,與肝纖維化最直 接關係的便是致纖維化(fibrogenesis)特性。在這個過程中,星狀細胞 產生膠原蛋白的量明顯增加,另一方面,星狀細胞也能產生分解細胞 外間質的基質金屬蛋白酶及其抑制因子,而這些物質的產生和活性調 節也隨星狀細胞的活化而發生相應變化,使細胞外間質不能被酶分解 而致逐漸沉積。. 19.
(21) 二、膠原蛋白 膠 原 纖 維 (Collagen fibers) 是 由 次 單 位 膠 原 微 纖 維 (Collagen fibrils)所構成,膠原微纖維由膠原分子組構成,膠原分子是一個三 條多胜鏈所構成,稱這些多胜鏈為α−鏈,它們排成三股螺旋。這些 α−鏈並不完全相同,根據α−鏈的不同,組成不同的膠原,稱為第一 型膠原蛋白、第二型膠原蛋白等,目前有 19 種。肝臟中存在 10 種 膠原蛋白,以第一型膠原蛋白最主要,在正常肝臟中佔 40-50%,當 肝硬化時,第一型膠原蛋白佔肝臟 60-70 % (Schuppan and Gressner, 1998)。 膠原合成時先在核糖體合成α−鏈,進一步 hydroxylation 及 glycosylation,在 rough-surfaced endoplastimic reticulum 內形成膠原 三螺旋分子,此時兩端有未形成三螺旋的部分稱 procollagen, procollagen 送出細胞外,進一步將膠原已定未形成螺旋端切斷,合 成膠原微纖維。第一型膠原蛋白由兩條α1 及一條α2 所組成寫成 Collagen [α1(I)]2α2(I),因此合成第一型膠原蛋白的基因有二,即 Collagen α1(I)、Collagen α2(I)。 α−鏈的氨基酸順序為 glycine-Xaa-Yaa,Xaa 及 Yaa 可為任何氨 基酸,但通常 proline 在 Xaa 或 Yaa 的位置,hydroxyproline 在 Yaa 的 位 置 。 典 型 的 膠 原 蛋 白 含 33 % 的 glycine 、 10-13% 的. 20.
(22) hydroxyproline 及 10-15% 的 proline。hydroxypoline 為膠原蛋白特有 的成分,測定 hydroxypoline 的量可以反應膠原蛋白的量,可用來表 示肝臟纖維化的程度(Hanauske., 1996)。. 三、間質金屬蛋白酶 (Matrix metalloproteinase, MMP) 與組織金屬 蛋白酶抑制劑 (Tissue inhibitors of metalloproteinases, TIMP) 細胞外間質的合成增加或降解減少會導致細胞間質的堆積,形成 肝纖維化,細胞外間質平衡主要透過 MMP 與 TIMP 兩者的調控 (Tsukada et al., 2006)。MMP 是 Zn-及 Ca-依賴性的 endopeptidase 能降 解種種細胞外間質蛋白,這些酵素依據其受質可分成 collagenases、 gelatinases,stromelysins、matrilysins 及 membrane-type-MMPs 等。 間隙性膠原分解酵素(interstitial collagenases)如 MMP-1、-8 及-13 可以 降解第一、二、三型膠原蛋白 (Visse and Nagase, 2003)。MMP 可以 受到細胞激素及生長因子的調控,如星狀細胞受到 Tgfβ1 的作用可釋 放出 MMP-13 (Lechuga et al., 2004),IGF 可以增加各種 MMP 的 mRNA 表現(Zhang et al., 2004)。除此之外,MMP 也可以受到細胞外 蛋白−蛋白相互作用的調控,組織中存在抑制 MMP 的酶稱 TIMPs, 目前已知有四種 TIMP。TIMP 會與 MMP 形成共價性結合,而抑制 MMP 降解膠原蛋白的能力,兩者的表現可使的細胞外間質的堆積形. 21.
(23) 成一個動態的平衡(Arthur, 2000)。在急性 CCl4 所誘導的肝損傷及膽道 結紮動物實驗中,發現 TIMP mRNA 的表現有顯著的提高(Iredale et al., 1996),而在患有慢性肝臟疾病的患者血清中發現,其血清的 TIMP 值及其 TIMP 蛋白質表現有上升的傾向,推論應該與肝纖維化形成有 關(Benyon et al., 1996;Murawaki et al., 1997)。另有體外及體內研究 指出,TIMP 抑制肝臟纖維化回覆的原因可能是透過阻止 HSC 的凋亡 (Murphy et al., 2002),後來亦有利用 TIMP-1 基因鼠證實,TIMP-1 過 度表現確可抑制 MMP 的活性及抑制 HSC 的凋亡,因而減少肝臟纖 維化自發性回覆(Yoshuji et al., 2000)。. 四、脂多糖結合蛋白 (Lipopolysaccharide binding protein, LBP) 由腸道細菌來的lipopolysaccharide (LPS)作用於肝臟的Kupffer細 胞,會釋放出發炎前細胞激素(proinfalmmatory cytokines)如腫瘤壞死 因子-α (tumor necrosis factor-α) 對肝臟造成傷害。LPS的作用機轉是 先與Kupffer細胞膜的CD14結合,而後經由Toll-like 4 receptor的途徑 引發訊息傳遞的產生,最後nuclear factor-κB進入細胞核內促使發炎前 細胞激素的產生(Su, 2002)。 當血中LPS增加時會促使肝臟合成LPS binding protein (LBP) (Su, 2002),LBP與LPS結合可增強LPS與CD14的作用(Su, 2002)。在很多. 22.
(24) 肝損傷的模式,如酒精性肝炎、內毒素中毒,LBP就如acute phase protein會被誘發出來。有文獻指出TAA誘發急性肝衰竭時血中內毒素 (LPS)明顯上升(Jia and Turek, 2005; Chen, 2005),因此TAA也應會使肝 臟合成LBP。. 五、轉化生長因子 (transforming growth factor β1, Tgfβ1) Tgfβ1 是一種多功能的 peptide,有許多同形異構物,例如: Tgfβ1、2、3…等;可以調控發炎、防禦、組織修復、胚胎生成與癌 細胞生成,刺激或抑制細胞的增生及分化,也可以調節其他生長因 子的作用及平衡氧化壓力。當組織受損而發生發炎反應時,發炎的 肝細胞會釋放 Tgfβ1,來調控白血球的補給與活性。而 Tgfβ1 的釋 放與活性的表現取決於星狀細胞內的結合蛋白的數目,即 Tgfβ1 接 受器的多寡,當 Tgfβ1 與其接受器結合後,星狀細胞的表現會加強, 促使 Tgfβ1 生成增加,進而刺激星狀細胞生成更多膠原蛋白。 Tgfβ1 會促進星狀細胞形成纖維母細胞以及生成細胞外間質來 修復受損的部位。藉由免疫染色結果得知,Tgfβ1 指出現在纖維化 的區域,肝臟未受損的部位並無 Tgfβ1 的反應(Nagy et al., 1991)。在 慢性肝病患者的肝切片中亦發現,Tgfβ1 mRNA、procollagen type I mRNA 的表現會依組織受損程度增加(Castilla et al., 1991)。在肝纖維. 23.
(25) 化的過程中,Tgfβ1 扮演很重要的角色,肝臟星狀細胞受 Tgfβ1 刺 激活化成肌纖維母細胞,而表現出纖維化之特性,造成肝纖維化。. 六、胰島素樣生長激素結合蛋白(insulin-like growth factor binding protein, IGFBP) 胰島素樣生長激素(Insulin like growth factors, IGFs)是生長所必 需的荷爾蒙因子,包括 IGF-I 和IGF-Ⅱ。IGF 普遍存在於肝臟、脂 肪、肌肉、腎臟、大腦…等處,其中IGF-I在肝臟的含量遠大於其他 組織,且隨著個體的成長增加,IGF-Ⅱ的含量於個體出生後則會降低。 IGF單鏈是由70個氨基酸所組成的polypeptide,40﹪的氨基酸組 成與胰島素類似,也具有類似胰島素的作用,即增加氨基酸進入細 胞、增進葡萄糖進入細胞、促進蛋白質合成並減少蛋白質降解,還有 與 成 長 因 子 相 類 似 的 成 長 、 分 化 及 細 胞 增 殖 作 用 (Cohick et al., 1993)。在體內IGF-I的作用遠大於IGF-Ⅱ。 IGF-I在肝纖維化中的角色不明之處仍多。外給IGF-I可以改善大 鼠的營養狀態及肝功能(Picardi et al., 1997),減少氧化傷害及肝纖維 化(Svegliati et al., 1999)。然而在體外實驗中,IGF-I卻有促進星狀細 胞増生(Pinzani et al., 1990;Scharf et al., 1998;Bachem et al., 1993; Zindy et al., 1992)及膠原蛋白合成作用(Svegliati et al., 1999)。以活化 態星狀細胞過度表現IGF-I的體外實驗發現,IGF-I刺激星狀細胞釋放 24.
(26) 出肝細胞成長因子而促進肝再生(Sanz et al., 2005)。 IGF的接受體有兩種,即第一型IGF接受體與第二型IGF接受體, 此二種接受體存在於大多數的細胞,以增殖細胞(proliferating cells)為 最多。第一型IGF接受體與IGF-I的親合力大於與IGF-Ⅱ,而第二型IGF 接受體則相反。 IGF-I受腦下垂體前葉所分泌的成長因子(growth factors;GH)調 控,GH會刺激肝臟及肝外組織產生IGF-I,釋放至循環系統,作用至 目標組織的IGF-I接受體。在此過程中IGF-I的運送需依賴結合蛋白 (IGF binding protein, IGFBP)。 IGFBP家族共有6個成員(IGFBP-1~6),主要負責IGF的攜帶,控 制IGF血液中的濃度,並調控其半衰期長短(Firth et al., 2002;Jones et al., 1995)。IGFBP-1,-2,-4主要由肝細胞分泌,IGFBP-3則主要表現在 Kupffer 細胞、竇內皮細胞(sinusoidal endothelial cells), 星狀細胞 (Scharf et al., 2001)及肝臟的纖維母細胞(Novosyadlyy et al., 2004)。 IGFBP-1被視為肝臟早期發炎的指標(Mohn et al., 1991;Scharf et al., 2004),IGFBP-1在急性四氯化碳肝損傷時會大量表現,並且能影 響IGF作用於星狀細胞(Scharf et al., 2004),促使纖維化的產生。 血液中有90%的IGF-I分子會與IGFBP-3及一個醣蛋白(acid labile subunit, ALS)鍵結,組成150-kDa的三級複合物。150-kDa的三級複合. 25.
(27) 物因無法穿過血管的內皮細胞,所以可增加IGF-I在血液的半衰期 (Twigg and Baxter, 1998)。在TAA誘發的大鼠肝硬化中,肝組織整體 IGFBP-3 mRNA的表現上升,但在Kupffer 細胞、竇內皮細胞的表現 下降,在活化態星狀細胞的表現增加(Novosyadlyy et al., 2005)。 IGFBP-3在肝纖維化過程中的角色功能仍不明。. 七、肝細胞生長因子(hepatocyte growth factor, HGF) HGF 是促進肝細胞增殖最強的細胞因子(Fausto et al., 2003)。 HGF 的受體 c-Met 在肝細胞和膽管上皮細胞高度表達,而非實質細 胞中,特別是星狀細胞中 c-Met mRNA 的表現卻相當低,因此,HGF 被認為是由非實質細胞產生,從而促進實質細胞的增殖。HGF 的作 用點是肝細胞,HGF 可促進肝細胞再生,改善肝功能和減輕損害。 在肝炎、肝硬化和肝癌症患者中發現液中 HGF 增加,可能是因肝細 胞壞死引致 HGF 合成增加。可見,在肝損害時,HGF 的表現會有 增加的趨勢,藉以修補受損之肝組織 (Schmandra, 2001)。. 26.
(28) 第四節. Thioacetamide 誘導肝纖維化. 口服或腹腔注射 TAA 在二至三個月內,可以穩定的誘發動物產 生不可逆的肝纖維化,且組織病理變化與人類的肝纖維化類似,因此 TAA 肝纖維化是近年來常被使用的動物模式(Li et al., 2002)。 TAA是含thiono-sulfur的化合物,其經過肝臟cytochrome P-450代 謝後會為 TAA -sulfoxide 、TAA-sulfdioxide (Wang et al., 2001; Ramaiah et al., 2001)這些代謝物會與肝小葉結合,而造成中央請靜脈 周圍肝組織壞死(Ramaiah et al., 2001;Mangipudy et al., 1995)。 另外也有研究指出,在代謝過程中,TAA 受到cytochrome P450 及flavin-containing monooxygenase 作用,能將dioxygen還原成超氧陰 離子,超氧陰離子進一步被摧化成過氧化氫,因此在TAA代謝過程中 會產生氧化壓力對肝臟造成損傷,早期階段會有glutathione排空及脂 質過氧化的產生 (Diez-Fernandez et al., 1993)。 蛋白質體學的研究顯示,TAA抑制了一些初級代謝途徑如fatty acid β-oxidation、branched chain amino acids 及methionine的斷裂,導 致succinyl-CoA排空,使heme的生合成減少,儲存的鐵釋出,游離鐵 增加,引發Fenton及Haber-Weiss 反應增強了氧化壓力的傷害 (Low et al., 2004)。. 27.
(29) 第二章 研究材料與方法 第一節. 研究材料. 一、藥品試劑: 編號. 名稱. 廠牌. 1.. Carbon Tetrachloride. OSAKA. 2. Thioactamide. Sigma-Aldrich. 3. Olive oil. Wako. 4. P-dimethylaminobenzaldehyde. Sigma-Aldrich. 5. TriSolution(trizol). GeneMark. 6. Chloroform. Showa. 7. Isopropanol. Scharlau Chemie S.A.. 8. PCR Master Mix kit. GeneMark. 9. MMLV RT Buffer. GeneMark. 10 dNTP Mix. GeneMark. 11 MMLV RT. GeneMark. 12 oligo(dT). GeneMark. 13 6 × DNA Loading Dye. GeneMark. 14 Gen-50 DNA Ladder. GeneMark. 15 Agarose Ι. Amresco. 16 Sodium hydroxide. Wako. 17 Hydrogen peroxide. Showa. 18 Tween 20. Sigma-Aldrich. 19 ALT kit(GPT 分析套組). Roche 28.
(30) 20 Colesterol kit(CHOL 分析套組). Roche. 21 Trizma base. Sigma-Aldrich. 22 Subcellular Proteome Extraction Kit. Merck. 23 IGFBP-1 antibody. Abcam. 23 Actin antibody. Santa Cruz Biotechnology. 24 Glycine. Amresco. 25 TITANIUM TaqPCR kit. BD. 二、設備器材 編號. 名稱. 廠牌. 1.. Spectrophotometer U-2001. Hitachi. 2.. PCR system 9700. A&B(Applied Biosystems). 3. AlphaDigDocTM(影像擷取與分析系統) Alpha Innotech Coporation. 4. CoBAS MIRA PLUS(生化儀). Roche. 5. KUBOTA 3500(離心機). KUBOTA. 29.
(31) 第二節 研究方法 一、AFEF 的製備 七至八月生的新鮮台灣金線連購自埔里有容農業生技有限公 司,以水萃取後,水萃液再經有機溶煤萃取,水容部分經減壓濃縮至 適當濃度後,儲存於-300C 備用。臨用時以去離子水配成適當濃度使 用。. 二、動物實驗 實驗動物選用 21-25 g 重之雄性 BALB/c 小白鼠(購自財團法人 國家實驗動物中心,台北),將實驗小鼠分成兩組:控制組及 TAA 組,TAA 組每週三次腹腔注射 2 ﹪TAA (溶於生理食鹽水;0.1ml/10g 體重)。TAA 組又分成三組分別經口投予去離子水或 AFEF 0.2、1.0 g/kg body weight,為期八週後犧牲。犧牲時,小鼠在乙醚麻醉下由 腹腔下腔靜脈採血,取出肝臟。肝臟以冰冷生理食鹽水洗淨後,吸 乾水分,稱重後分成四份,(1) 稱重後乾燥供 hydroxyproline 定量, (2) 浸於中性 10%福馬林供病理切片用. (3) 儲存於液態氮中供. RNA 抽取,(4) 儲存於 -800C 備用。. 三、血液生化值實驗 實驗動物以乙醚麻醉後,以 1 ml 針筒於下腔靜脈採血。全血收 30.
(32) 集於微量試管中,以 4700 rpm 離心 10 分鐘,吸取上清液。利用自動 分析生化儀檢測 ALT 及 Cholestrol。. 四、肝臟組織病理學觀察 取肝臟最大葉的三分之一浸泡 10﹪福馬林,進行石臘包埋及切 片製作,再以蘇木青與伊紅染色法(Hematoxylin and eosin stain)和天狼 猩紅(Sirus red)染色,供病理檢查。. 五、肝臟 Hydroxypoline 含量測定 肝臟 Hydroxypoline 含量測定的方法乃參考 Neuman RE 及 Logan MA (Neuman and Logan, 1950) 之方法。將肝組織秤取約 0.2 克於 100℃烘箱中隔夜乾燥。秤取乾燥肝組織織重量放入試管中,加入 2 ml 的 6 N HCl,於 100℃烘箱中放置 16 小時水解。冷卻後取 1 ml 水解 物以 10000 rpm 離心 30 分鐘。取 0.1 ml 上清液加少量的 1 N NaOH 中和水解產物。各加 1 ml 新鮮配置 0.01 M 的 CuSO4、 2.5 N NaOH、 H2O2 促進氧化還原反應,反應 5 分鐘後,以 80℃水浴鍋破壞過多之 過氧化物。冷卻至室溫,再加入 2 ml 之 5﹪ P-dimethylaminobenzaldehyde 及 4 ml 之 3 N H2SO4,再放入 70℃水浴 16 分鐘反應呈色,以分光光度計於 540nm 測量吸光值。其表示單位 為 μg hydroxypoline/g 肝臟重。 31.
(33) 六、RT-PCR 分析 抽取肝臟組織 RNA 取 0.1 g 的肝組織加入 1ml triSolution,利用 18 號及 23 號針頭 粉碎組織,於 4℃、6400 rpm 條件下離心 10 分鐘。取上清液加入 0.2ml 的 chloroform 萃取 RNA,震盪 15 秒後靜置冰上 3 分鐘,於 4℃、12700 rpm 離心 15 分鐘,取上清液加入 0.5 ml Isopropanol 使 RNA 沉澱, 上下搖晃後靜置冰上 10 分鐘,於 4℃、12700 rpm 離心 10 分鐘。倒 掉上清液,留下沉澱物,再加入 75 ﹪酒精(99.9 ﹪絕對酒精以 DEPC 水稀釋)後,靜置於-30℃冰箱內 30 分鐘。於 4℃、10000 rpm 離心 5 分鐘,再倒掉清液,加入 DEPC 水 20- 60 μl 溶解。利用分光光度器 於 260 nm /280 nm 測定吸光度。 合成 cDNA cDNA 的 製 備 主 要 採 用 MMLV Reverse Transcriptase (first-strand cDNA synthesis)。取 1 μg mRNA 加入 2.5 μl 10×MMLV RT buffer、0.6 μl MMLV RT、5 μl dNTP、2.5 μl 0.1M DTT、1 μl oligo dt primer,混合後於 37℃反應 60 分鐘。 Polymerase Chain Reaction(PCR) 利用 PCR Master Mix 套組進行 specific DNA fragments amplification。採用的 primer 序列如 Table 1。. 32.
(34) 七、Real time PCR 分析 由於 RT-PCR 只可得知影像分析之相對定量,為了更準確分析肝 臟 mRNA 的表現,我們利用 Real time PCR 再次驗證。 將 cDNA 稀釋 100 倍,取 DNA template 5 μl 加入 10× TITANIUM Taq PCR buffer 2.5 μl、50μ dNTP Mix (10mM) 0.5 ul、Sybr- Green (1:1000) 1 μl、50 × TITANIUM Taq Polymerase 0.5 μl 及欲分析之 forward 及 reverse primer 各 1 μl,混合好之液體放至 Real time q-PCR box 中作用,最後分析數據。primer 序列採用 Table 1 之 RT-PCR 序列。. 八、統計方法 數據結果以平均值(mean) ±標準誤(Standard error, SE)表示,各組 與對照組間的數據以單因子變異數分析(One-way analysis of variance;One-way ANOVA)方法分析,並進行 Dunnett 測試,以 P 值 小於 0.05 表示在統計學上具有顯著差異。. 33.
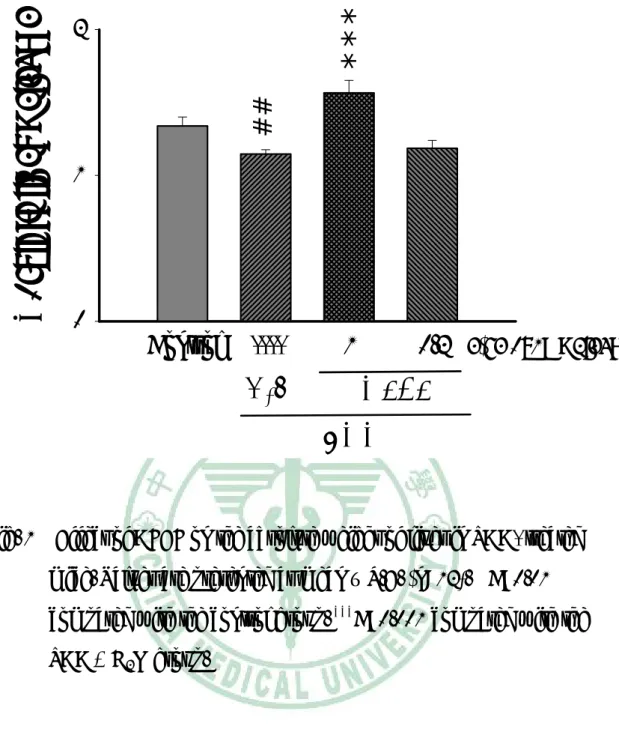
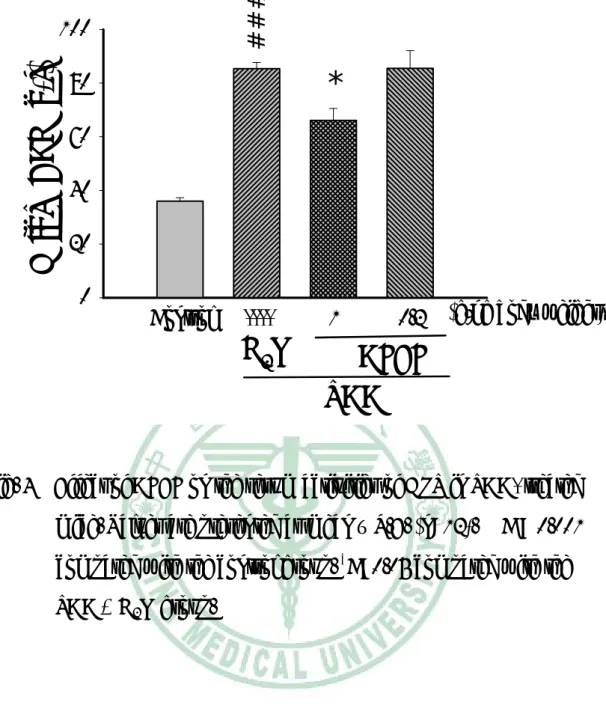
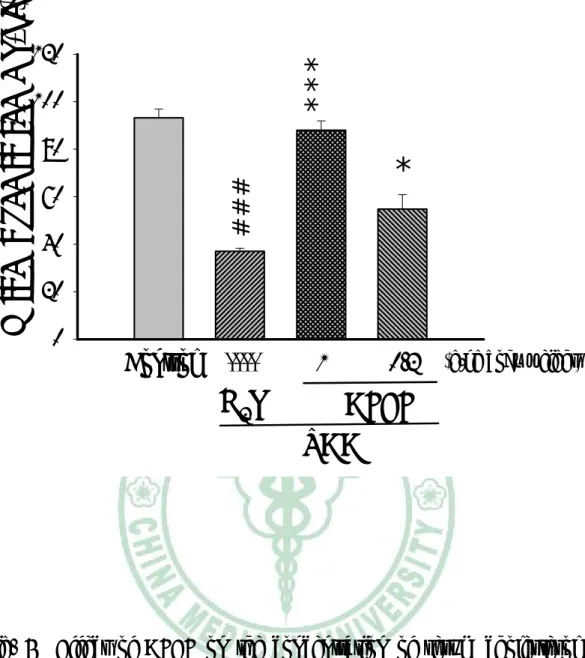
(35) 第三章. 實驗結果. 一、 對體重的影響 TAA 投予前各組間小鼠平均體重沒有差異,約為 23- 24 g。小 鼠一週三次給予 TAA,為期八週,第五週小鼠體重有減輕的傾向, 第六週至第八週,小鼠體重明顯低於控制組。AFEF 0.2 g/kg 組小鼠 體重於第 5 週至第 7 週高於 TAA 組。AFEF 1.0 g/kg 組小鼠體重於 第 5 週至第 8 週高於 TAA 組(Table 2)。. 二、 對肝臟重量及含水量影響 TAA 組小鼠最後的肝臟絕對重量明顯比控制組輕,AFEF 1.0 g/kg 組的小鼠,其肝臟重量明顯比 TAA 組重,而 AFEF 0.2 g/kg 組 的肝臟重量與控制組沒有顯著差異(Fig. 1)。TAA 組小鼠的肝臟相對 重量較控制組顯著增加,給予 AFEF 的小鼠肝臟相對重量 TAA 組輕 (Fig. 2)。 小鼠犧牲時秤取肝臟 0.2 g,放入烘箱中,經 1000C、16 小時乾 燥後,測量肝臟含水量。TAA 組小鼠肝臟含水量比控制組顯著增 加,而投與 AFEF 的組別,其肝臟含水量皆比 TAA 組顯著降低(Fig. 3)。. 34.
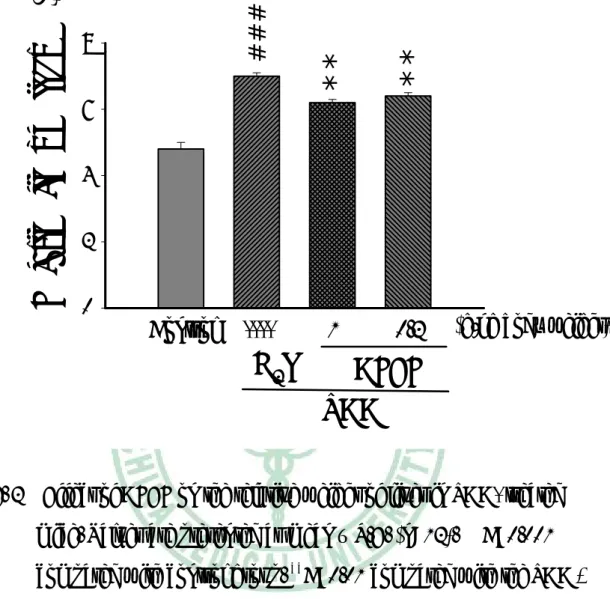
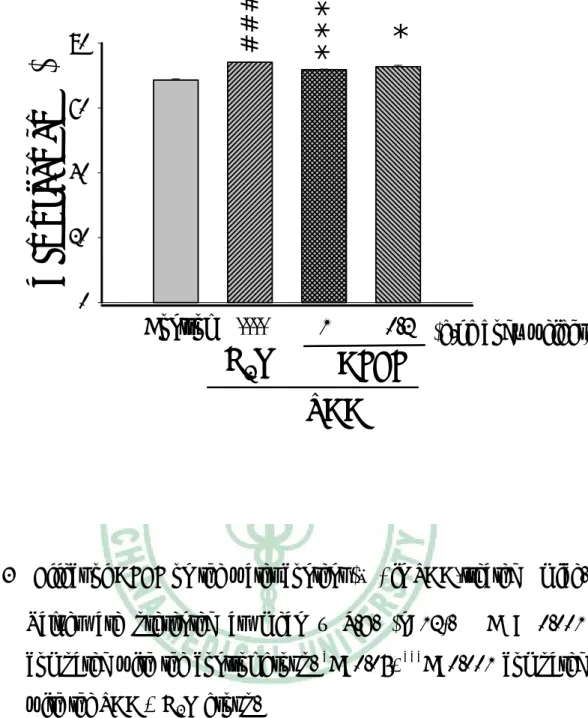
(36) 三、 對血清生化值影響 實 驗 第 八 週 後 犧 牲 採 血 , 取 血 清 部 份 測 定 ALT 活 性 及 Cholesterol 濃度。如 Fig. 4、Fig. 5 所示,第八週的 ALT 值,TAA 組小鼠較控制組顯著增加,AFEF 1.0 g/kg 組的小鼠其 ALT 值較 TAA 組顯著降低,低劑量組則無明顯差異。第八週血清 Cholesterol 濃度, TAA 組較控制組顯著降低,給予 AFEF 的組別,無論高劑量或低劑 量,其血清膽固醇皆顯著增加。. 四、 Hydroxylpoline 含量 TAA 組肝臟 hydroxyproline 含量較控制組顯著增加,AFEF 1.0 g/kg 組的小鼠其肝臟 hydroxylpoline 含量較 TAA 組低,低劑量組 無明顯差異(Fig. 6)。. 五、 病理檢驗 肝臟病理切片的觀察顯示,投與 TAA 八週之小鼠,其組織細胞 有浸潤的現象,且細胞核有破裂的情形(Fig. 7B),投予高劑量的 AFEF 則可減少細胞死亡發生(Fig. 7C)。以 Sirus red 染色發現,單獨 給予 TAA 八週之小鼠肝組織出現膠原蛋白纖維束(Fig. 8B),而高劑 量 AFEF 組可減少纖維束的產生(Fig. 8C)。. 35.
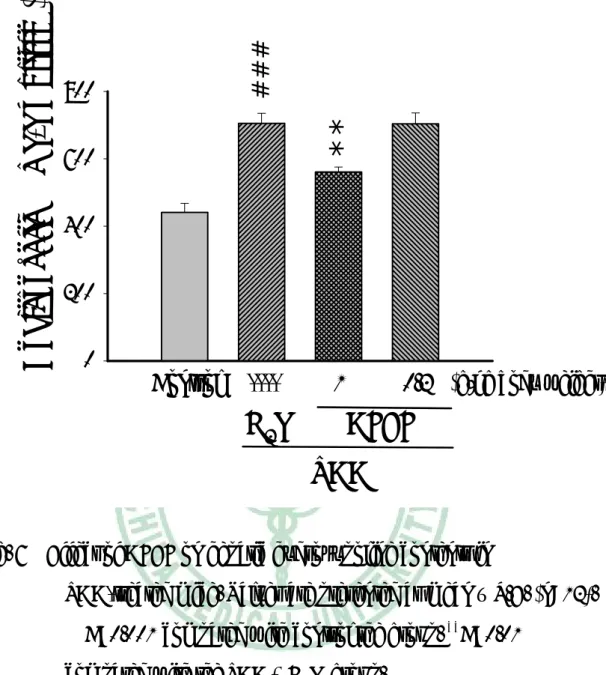
(37) 六、 RT-PCR 結果 如 Fig. 9 所示,RT-PCR 的分析顯示,TAA 組肝臟 Tgfβ1、Collagen (α1)(I)的表現較控制組分別提高 33、44%。AFEF 1.o g/kg 組的肝臟 Collagen (α1)(I)的表現低於控制組,Tgfβ1 的表現與控制組比較沒有 差異。TAA 組肝臟 MMP13 的表現較控制組低約 46%。AFEF 1.o g/kg 組的肝臟 MMP13 的表現高於控制組。 如 Fig. 10 所示,TAA 組肝臟 LBP、CD 14、TLR-4 及 TNF-α Rcepter 的表現較控制組分別提高 37.5、23、23 及 65﹪。AFEF 1.0 g/kg 組的 肝臟 LBP、CD 14、TLR4 及 TNF-α Rcepter 的表現低於控制組。 如 Fig. 11 所示,TAA 組肝臟 IGF-I 的表現與控制組比較沒有顯著 差異,AFEF 組的肝臟 IGF-I 的表現與 TAA 組比較也沒有差異。TAA 組肝臟 IGF-R 及 IGFBP-1 的表現較控制組分別提高 44、77﹪。AFEF 1.0 g/kg 組的肝臟 IGF-R 及 IGFBP-1 的表現低於控制組。TAA 組肝臟 IGFBP-3 的表現較控制組低約 43%。AFEF 1.0 g/kg 組的肝臟 IGFBP-3 的表現高於控制組。 如 Fig. 12 所示,TAA 組肝臟 HGF 及 c-Met 的表現較控制組分別 提高 56、44﹪。AFEF 1.0 g/kg 組的肝臟 HGF 及 c-Met 的表現低於控 制組。. 36.
(38) 七、 Real-time PCR 結果 IGFBP-1、LBP 的表現進一步利用 Real-Time PCR 來分析,將 cDNA 稀釋 100 倍後,以 Titanium TaqPCR kit 進行定量分析。TAA 組小鼠,IGFBP-1 及 LBP 的表現約為控制組的 58、54%。AFEF 1.0 g/kg 組的肝臟 IGFBP-1 及 LBP 表現較 TAA 組分別減少約 40、 65 %(Fig. 13)。. 37.
(39) Table 1 Oligonuleotide sequences used in RT-PCR and Real-time PCR* reactions mRNA. size. GAPDH. 76 bp. Collagen(αⅠ)(Ⅰ) 174 bp MMP13. 135 bp. Tgfβ1. 176 bp. IGF-I. 99 bp. IGF recepter-1. 90 bp. IGFBP-1*. 81 bp. IGFBP-3. 139 bp. LBP*. 127 bp. CD 14. 375 bp. TLR4. 85 bp. TNF recepter. 96 bp. HGF. 146 bp. c-Met. 370 bp. Oligomers Sense 5′-TGTGTCCGTCGTGGATCTGA-3′ Antisense 5′-CCTGCTTCACCACCTTCTTGA-3′ Sense 5′-GGTCCCAAAGGTGCTGATGG-3′ Antisense 5′-GACCAGCCTCACCACGGTCT-3′ Sense 5′-GGTGGAGATATCAGGGGAGA-3′ Antisense 5′-TGGGGTTCTCAATTGCATTA-3′ Sense 5′-CAAGGGCTACCATGCCAACTTC-3′ Antisense 5′-TTGCGACCCACGTAGTAGACGA-3′ Sense 5′-GCTCTGCTTGCTCACCTTCAC-3′ Antisense 5′-CTCGGTCC ACACACGAACTG-3′ Sense 5′-GCTGAGGGAGGAGGCGGC-3′ Antisense 5′-GGCGAGGGGCAGAAACGC-3′ Sense 5′-ACTCGGAATTCCTCATCGTC-3′ Antisense 5′-GGTTACACAATCAGCATCGG-3′ Sense 5′-ATTTGCGACATCTCTGAACG-3′ Antisense 5′-GTTCTAGGGCACTCTGGGAG-3′ Sense 5′-ACCCTTGACCTGGACTTGAC-3′ Antisense 5′-ACAGTGCCCGCTCTTAAAGT-3′ Sense 5′-CCTAGTCGGATTCTATTCGGAGCC-3′ Antisense 5′-AACTTGGAGGGTCGGGAACTTG-3′ Sense 5′-ATGGAACACATGGCTGCTAA-3′ Antisense 5′-TCATACCCCTGGAAAGGAAG-3′ Sense 5′-GAGATGGTTGTTTCCGTGTG-3′ Antisense 5′-GCCACTCCCTGAGTTTTAGC-3′ Sense 5′-GGGCTGAAAAGATTGGATCA-3′ Antisense 5′-TCGAACAAAAATACCAGGACG-3′ Sense 5′-GAATGTCGTCCTACACGGCC-3′ Antisense 5′-CACTACACAGTCAGGACACTGC-3′. 38.
(40) Table 2. The changes of body weight of mice Group. g/kg. Week0. Week1. Week2. Week3. Week4. Week5. Week6. Week7. Week8. 24.3±0.4. 25.2±0.6. 25.5±0.7. 25.8±0.5. 26.0±0.6. 25.3±0.3. 26.8±0.4. 28.0±0.5. 27.8±0.7. 25.2±0.3. 24.2±0.4. 24.3±0.4. 24.6±0.6. 24.9±0.5. 23.0±1.0. 19.4±0.8###. 16.9±0.2###. 16.8±0.2###. 1.0. 25.5±0.4. 24.3±0.3. 24.3±0.3. 24.5±0.3. 24.8±0.4. 25.5±0.3*. 26.1±0.3***. 24.4±0.7****. 22.9±0.9***. 0.2. 25.5±0.3. 24.0±0.5. 24.0±0.4. 24.2±0.3. 25.1±0.3. 25.6±0.4*. 23.8±0.8**. 20.5±0.9**. 17.7±0.7. Control TAA + H2O + AFEF. body weight(g). Dose. Values are presented as mean ± S.E.(n=12).. ###. P<0.001 compared with. the control group. **P<0.01 and ***P<0.001 compared with the TAA + H2O group.. 39.
(41) *** ##. Absolute liver weight (g). 2. 1. 0. Control. ----. 1. H2O. 0.2 (g/kg body weight). AFEF TAA. Fig. 1 Effect of AFEF on the absolute weight of liver in TAA- treated mice. Values are presented as mean ± S.E. (n=12). ##P<0.01 compared with the control group. ***P<0.001 compared with the TAA + H2O group.. 40.
(42) **. **. ###. Relative liver weight (%). 8. 1. 0.2. 6 4 2 0. Control. ----. H2O. (g/kg body weight). AFEF TAA. Fig. 2 Effect of AFEF on the relative weight of liver in TAA- treated mice. Values are presented as mean ± S.E. (n=12). ###P<0.001 compared with control group. **P<0.01 compared with the TAA + H2O group.. 41.
(43) ###. ***. water content (%). 80. ----. 1. *. 60 40 20 0. Control. H2O. 0.2 (g/kg body weight). AFEF TAA. Fig. 3 Effect of AFEF on the water content (﹪) in TAA-treated Values are presented as mean ± S.E. (n=12). compared with the control group. *P<0.05, with the TAA + H2O group.. 42. ***. mice.. ###. P < 0.001. P<0.001 compared.
(44) Serum ALT (u/l). ###. 100 80. *. 60 40 20 0. Control. ----. 1. 0.2. H2O. AFEF TAA. (g/kg body weight). Fig. 4 Effect of AFEF on the serum activities of GPT in TAA- treated mice. Values are presented as mean ± S.E. (n=12). ###P < 0.001 compared with the control group. *P<0.05 compared with the TAA + H2O group.. 43.
(45) Serum Cholesterol (mg/dl). ***. 120 100 80. *. ###. 60 40 20 0. Control. ----. 1. H2O. 0.2. (g/kg body weight). AFEF TAA. Fig. 5 Effect of AFEF on the concentration of serum cholesterol in TAA-treated mice. Values are presented as mean ± S.E. (n=12). ###. P<0.001 compared with the control group. *P<0.05, ***P<0.001. compared with the TAA + H2O group.. 44.
(46) ###. **. Hydroxypoline ( mg/g wt tissue). 800 600 400 200 0. Control. ----. 1. H2O. 0.2 (g/kg body weight). AFEF TAA. Fig. 6 Effect of AFEF on hepatic hydroxypoline contents in TAA-treated mice. Values are presented as mean ± S.E. (n=12). ###. P<0.001 compared with control the group. **P<0.01. compared with the TAA + H2O group.. 45.
(47) (A). (B). (C). Fig. 7. The photomicrographs of liver section taken from mice and stained. with. hematoxylin-eosin.. (A). Control. group.. (B). TAA+H2O, Note that tissue infiltration and necrosis was observed. (C) TAA+AFEF 1.0 g/kg body weight (Original magnification : left ×40, right ×100). 46.
(48) (A). (B). (C). Fig. 8. The photomicrographs of liver section taken from mice and stained with Sirus red. (A) Control group. (B) TAA+H2O, Note that displaying fibrin of collagen was observed. (C) TAA+AFEF 1.o g/kg body weight. (Original magnification ×40). 47.
(49) 174 bp. Collagen(α1)(I). 176 bp. Tgfß1. 135 bp. MMP13. 76 bp. GAPDH Control. ----. 1.0. 0.2. H2O. (g/kg body weight). AFEF. C o TA TA TA. 3. n tro l A +H 2O A + A F E F 1 g /k g A + A F E F 0 .2 g /k g. *. *. 2. ##. ###. mRNA expression(ratio). TAA. 1. 0. C o lla g e n ( α 1 ) ( I). T g fβ 1. M M P 13. Fig. 9 (A) RT-PCR analysis of Collagen (α1)(I), Tgfβ1 and MMP13 expressions in control mice, in TAA-treated mice, and in mice treated with TAA + AFEF (1.0 or 0.2 g/kg body weihgt). Expression of the amplified fragement corresponding to GAPDH is shown for comparison. (B) Densitometric analyses of Collagen (α1)(I), Tgfβ1 and MMP13 expressions after normalization against GAPDH. Values are presented as mean ratio ± S.E. (n=4). ##. P<0.01,. ###. P<0.001 compared with the control group. *P<0.05. compared with the TAA + H2O group.. 48.
(50) LBP. 127 bp. CD 14. 375 bp. TLR4. 85 bp. TNF-R. 96 bp. GAPDH. 76 bp Control. ----. 1.0. 0.2. H2O. (g/kg body weight). AFEF TAA C T T T. o n tro l A A + H 2O A A + A F E F 1 g /k g A A + A F E F 0 .2 g /k g. 3. 1. ##. 0 L B P. * **. #. **. **. 2. #. mRNA expression(ratio). #. T L R 4. C D 14. T N F -R. Fig. 10 (A) RT-PCR analysis of LBP, CD14 ,TLR4 and TNF-R expressions in control mice, in TAA-treated mice, and in mice treated with TAA + AFEF (1.0 or 0.2 g/kg body weight). Expression of the amplified fragement corresponding to GAPDH is shown for comparison. (B) Densitometric analyses of LBP, TLR4, CD14 and TNF-R expressions after normalization against GAPDH. Values are presented as mean ratio ± S.E. (n=4). #P<0.05, ##. P<0.01 compared with the control group.. with the TAA + H2O group.. 49. **. P<0.01 compared.
(51) IGF-I. 99 bp. IGF-R. 90 bp. IGFBP-1. 260 bp bp. IGFBP-3. 139 bp 76 bp. GAPDH Control. ----. 1.0. H2O. 0.2. (g/kg body weight). AFEF TAA. ###. 5. o A A A. n tro l A + H 2O A + A F E F 1 g /k g A + A F E F 0 .2 g /k g. **. 4. ##. 3 2. *. **. 1 0. #. mRNA expression(ratio). C T T T. IG F -1. IG F B P -1. IG F -R. IG F B P -3. Fig. 11 (A) RT-PCR analysis of IGF-I, IGF-R, IGFBP-1 and IGFBP-3 expressions in control mice, in TAA-treated mice, and in mice treated with TAA + AFEF (1.0 or 0.2 g/kg body weight). Expression of the amplified fragement corresponding to GAPDH is shown for comparison. (B) Densitometric analyses of. IGF-R,. IGFBP-1. and. IGFBP-3. expressions. after. normalization against GAPDH. Values are presented as mean ratio ± S.E. (n=4). #P<0.05, #P<0.05,. ##. P<0.01 compared with. the control group. *P<0.05, **P<0.01 compared with the TAA + H2O group.. 50.
(52) 146 bp. HGF. 375 bp. c-Met. 76 bp. GAPDH Control. ----. 1.0. H2O. 0.2. (g/kg body weihgt). AFEF TAA Control TAA+H2O TAA+AFEF 1g/kg TAA+AFEF 0.2g/kg. #. ** **. 2. **. 3. ###. mRNA expression(ratio). 4. 1 0. HGF. c-Met. Fig. 12 (A) RT-PCR analysis of HGF and c-Met expressions in control mice, in TAA-treated mice, and in mice treated with TAA + AFEF (1.0 or 0.2 g/kg body weight). Expression of the amplified fragement corresponding to GAPDH is shown for comparison. (B) Densitometric analyses of HGF and c-Met expressions after normalization against GAPDH. Values are presented as mean ratio ± S.E. (n=4).. #. P<0.05,. #. P<0.05. compared with the control group. **P<0.01 compared with the TAA + H2O group.. 51.
(53) Control TAA+H2O TAA+AFEF 1g/kg TAA+AFEF 0.2g/kg. 3 #. 2 1. *. mRNA fold increase. 4. 0. IGFBP-1. Fig. 13. LBP. Real Time PCR analysis of IGFBP-1 and LBP levels in TAA-treated mouse liver tissue. Results are expressed in arbitrary units, and Data are mean mean ± S.E. (n=3). #P<0.05 compared with the control group.*P<0.05 compared with the TAA + H2O group.. 52.
(54) 第四章 討論與結論 本研究以小鼠同時投與AFEF及TAA,探討AFEF對TAA誘發小鼠 肝纖維化是否有減輕的作用,結果發現AFEF可以減輕TAA誘發的小 鼠肝臟纖維化,此減輕作用可能經由抑制發炎相關的LBP及IGFBP-1 的表現。 文獻指出TAA會使小鼠體重減輕(Laleman, 2006),在本研究也發 現TAA 使小鼠體重明顯減輕,AFEF的處理能明顯提升小鼠的體重。 TAA的體重下降作用與血中的leptin 及IGF-I經由中樞神經系統的作 用有密切關係(LaPaglia, 1998)。本實驗重點在抗纖維化的研究,僅分 析肝臟 IGF-1基因的表現,沒有測定血中的leptin 及IGF-1的含量, 無法說明AFEF的減輕體重下降作用是否與leptin 及IGF-1有關。 肝臟受到損傷時,肝會腫大,當損傷嚴重時,肝臟會萎縮(Bissell, 1990)。TAA雖使肝臟絕對重量減輕,但肝臟相對重量卻增加,顯示 在本試驗條件下,TAA對肝臟的損傷還不是很嚴重。AFEF對TAA肝 臟重量的影響有反轉的效果。肝臟生長因子HGF具有促進肝臟再生的 作用(Skrtic et al, 1999),在肝臟受到傷害時,由於修複的作用,會使 肝臟HGF的表現增加 (Sanz et al., 2005)。TAA誘使肝臟的HGF及其作 用的接受體 c-Met mRNA的表現增強,此應與肝臟的相對重量增加有 關,AFEF可以減弱TAA所提升的HGF及c- Met mRNA的表現。這些結. 53.
(55) 果顯示AFEF有減輕TAA肝損傷的效果。 當肝細胞受到損傷時,細胞內的GPT會釋放至血液中,所以GPT 最常被用來當作肝臟損傷時初期的生化指標(Sturgill and Lambert, 1997),而血清GPT在肝臟中比GOT更具專一性(Bissell, 1990)。投與 TAA八週後發現,高劑量AFEF明顯降低血清GPT值,顯示高劑量 AFEF對於TAA肝臟損傷具有減輕效果。病理切片檢查的結果也顯 示,AFEF可以減輕TAA引起的肝組織發炎及壞死的情形。 膽固醇主要在肝臟合成(Bissell, 1990),合成完成的膽固醇會釋放 至血液中,因此血中膽固醇的濃度可以顯示肝藏的合成功能。TAA使 血中膽固醇濃度下降,顯示TAA的肝損傷影響到肝臟的合成功能。 AFEF對此有改善的效果,支持了AFEF可以減輕TAA的肝毒性。 所有慢性肝炎皆會引起肝纖維化的產生(Poynard, 1997、2000; Brunt, 2004),小鼠經 TAA 八週的投予,在組織病理的檢查可見現纖 維狀組織出現(Ramaiah et al., 2001;Mangipudy et al., 1995),即結締 組織增生。結締組織主要由膠原蛋白構成,hydroxypoline 是膠原蛋白 特有的成分,測定 hydroxypoline 可以反應膠原蛋白的量,可用來表 示肝纖維化的程度(Hanauske-Abel, 1996)。在本實驗 TAA 誘發肝臟纖 維化,其肝臟 hydroxypoline 含量明顯增加,而高劑量 AFEF 可使 hydroxypoline 含量減少,此情形在組織病理檢驗也得到證實。膠原蛋. 54.
(56) 白種類很多,已知在肝臟纖維化時以膠原蛋白型 I 的增加最主要 (Iredale, 1996) , 因 此 在 本 實 驗 也 利 用 RT-PCR 的 技 術 分 析 肝 臟 Collagen (α1)(I)的表現,TAA 明顯增加肝臟 Collagen (α1)(I)的表現, 此作用可被 AFEF 減弱。顯示 AFEF 可以減輕 TAA 誘發肝纖維化的 作用。 肝臟纖維化是因為細胞外間質(extracellular matrix;ECM)過度 沉積(Friedman, 2003)。Matrix metalloproteinases (MMPs)是一種蛋白分 解酵素,可以降解細胞外間質。MMPs有多種,與肝纖維化有關的以 MMP13較主要,可降解膠原蛋白型I (Yoshiji, 2002)。肝臟MMP13的 表現曾強,可以降解肝纖維化,反之則肝纖維化較重。TAA可使肝臟 的MMP13表現減弱,AFEF的處理可以增強MMP13的表現,此更支持 了AFEF可以減輕TAA誘發肝纖維化的作用。 Lipopolysaccharide (LPS) 是 一 眾 所 皆 知 的 發 炎 因 子 , 其 會 與 Kupffer 細胞作用引起發炎反應,造成肝損傷。已有文獻指出,TAA 的肝損傷會使肝臟的 LPS 增加(Chen, 2005)。LPS 會刺激肝臟合成 LPS 結合蛋白金(LPS-binding protein;LBP),血中的 LBP 與 LPS 結合, 可將 LPS 運送至 kupffer 細胞與 Toll 接受體及 CD14 結合,進而活化 kupffer 細胞釋放出 TNF-α 等發炎因子造成發炎反應(Su, 2002)。在本 實驗,TAA 造成的肝臟發炎反應,其 LBP、TL-4 接受體及 CD-14 的. 55.
(57) mRNA 的表現顯著提高,投與 AFEF 則有效降低這些 mRNA 的表現。 高劑量的 AFEF 也可以減少 TNF-α Rcepter 的表現,此結果表示 AFEF 可以有效抑制 LPS 所引起的發炎反應。 胰島素成長因子-I (insulin growth factor 1;IGF-I) 在肝硬化的病 理角色還有很多爭議,肝硬化的病人血中 IGF-1 濃度下降,此與肝硬 的併發症營養不良有關(Mendenhall et al., 1989; Inaba et al., 1999)。因 此 有 研 究 指 出 , 外 給 IGF-I 對 實 驗 肝 硬 化 有 改 善 的 效 果 (Castilla-Cortazar et al., 1997; Mirpuri et al., 2002; Canturk et al., 2003)。雖然如此,局部作用的 IGF-1 具促細胞分裂增生的作用,能 活化星狀細胞產生膠原蛋白會促進肝臟纖維化(Scharf et al., 1998)。 在血液中,IGF-I是與其結合蛋白結合的,結合蛋白有6種之多, 其中肝細胞可以產生的IGFBP-1 (IGF binding protein 1),被視為肝臟 早期發炎的指標之一(Mohn et al.,1991;Scharf et al.,2004)。IGFBP-1 在急性肝損傷時會大量表現,IGFBP-1會藉由釋放出磷酸根轉而與 IGF-1結合,當此結合體(IGFBP-1/IGF-I complex)與星狀細胞接觸後, IGFBP-1會與星狀細胞表面的integrin結合而釋放IGF-I,游離的IGF-I 則轉與星狀細胞膜表面的IGF recepter結合,一旦 IGF-I 與其受體結 合後,便會刺激加速星狀細胞cDNA合成(Scharf et al., 2004)。在本實 驗 , TAA 雖 沒 有 使 肝 臟 IGF mRNA 表 現 增 強 , 但 明 顯 的 提 升. 56.
(58) IGFBP-1、IGF接受體mRNA的表現,由這些可推測TAA使星狀細胞 活化引起肝藏纖維化與IGF-I的作用有關。這些也說明了AFEF減輕 TAA引起肝臟纖維化的機轉之一,是經由抑制IGFBP-1、IGF接受體 mRNA 的表現。 IGFBP-3 主 要 表 現 在 非 實 質 細 胞 如 Kupffer cell 、 sinusoidal endothelial cells 及星狀細胞等(Scharf et al., 2001)。在 TAA 誘發的大 鼠肝硬化,Kupffer cell、sinusoidal endothelial cells 的 IGFBP-3 mRNA 表現下降,活化的星狀細胞表現增強,整體肝臟的表現上升 (Novosyadlyy et al., 2005)。在本研究的結果與前述文獻不同,TAA 的 處理使小鼠肝臟整體 IGFBP-3 mRNA 表現下降,而 AFEF 能提升 IGFBP-3 mRNA 的表現。二者的差異可能與動物品種或誘發時間的長 短有關。 Tgfβ1一直被當作造成肝臟纖維化的一個重要因子,主要因為 Tgfβ1會活化肝臟星狀細胞,進而造成collagen的產生。本實驗也證 實,TAA確實會造成小鼠肝臟Tgfβ1表現增加,但AFEF卻無法抑制 Tgfβ1的mRNA表現,由此推論AFEF減輕肝臟纖維化的機轉與Tgfβ1 無關。. 57.
(59) 結. 論. 小鼠以TAA每週三次腹腔注射,連續八週,誘發小鼠肝纖維化, 同時經口投予AFEF。TAA使小鼠體重減輕、肝臟相對重量增加、血 清GPT值上升及膽固醇降低,增加肝組織膠原蛋白含量,提升發炎相 關的因子如LBP、IGFBP-1的mRNA表現。高劑量AFEF可以改善這些 現象,顯示AFEF對TAA誘發肝纖維化有改善的作用。. 58.
(60) 參考文獻 林學詩,珍貴藥草金線連的栽培法,花蓮區農技報導,1994。 周惠慈、謝萬全、張其善,台灣金線連組織培養繁殖法,理工學報, 1982;19:155-160。 蘇洪傑,台灣野生蘭,豐年叢書,豐年社,1975;79- 81。 劉新裕、蔡新聲、徐原田、劉柏煙,金線連之繁殖與栽培技術,科學 農業,1987;35:125- 129。 甘偉松,藥用植物學,國立中國醫藥研究所出版,1979,p. 900。 甘偉松,台灣植物藥材誌,第二輯,國立中國醫藥出版社,1965,p. 85。 甘偉松、程兆熊、張清標,金線連的藥用植物學考察,行政院衛生署 中醫藥年報,1983;3:46- 55。 蔡東,名貴的藥用植物-金線連,興農,1986;196:83- 84。 甘偉松,藥用植物學,國立中國醫藥研究所出版,1986,p.647。 梁文俐、陳榮進、江育仁、蘇慶華、楊玲玲、顏焜棾,台灣金線連之 研究 Ι,金線連之生理活性研究,台灣科學,1990;47- 58。 陳卓、黃自強,金線連降血糖作用的初步研究,福建醫藥雜誌,2000; 22:207- 208。 楊旻璟、林順福、盧美光、鄭可大,台灣金線連多糖品系遺傳標誌篩. 59.
(61) 選,”生物多樣性與天然藥物之開發”國際研討會,台北醫學大學 研究所,2001。 徐麗芬、駱秋萍、王升陽、孫秀珍、唐培玲、陳志淮、曾驥孟、楊寜 蓀,台溫金線連萃取物引發人類乳癌之凋亡:利用 DNA 微陣技 術、細胞生物化學與代謝產物圖譜分析之研究,”生物多樣性與 天然藥物之開發”國際研討會,中央研究院生物農業科學研究 所,2001。 羅妙華,以大鼠肝細胞與星狀細胞為模式探討抗纖維化之保肝中藥, 國立陽明大學醫學院碩士論文,2002。 林文川,台灣金線連的研究近況,國際藥用植物產業發展研討會專 刊,2004。 徐致芬,台灣金線連有效分劃對小鼠非特異性免疫調節,中國醫藥大 學碩士論文,2004。 蕭宏柏,台灣金線連有效分劃對卵蛋白誘發小鼠呼吸道過度反應之免 疫調節作用,中國醫藥大學碩士論文,2005。 廖思穎,台灣金線連多醣體成分對先天免疫反應的影響,中國醫藥大 學碩士論文,2005。 江芙美,台灣金線連成份—天麻素及天麻元及尿嘧啶之藥物動力學研 究. 中國醫藥大學碩士論文,2003。. 60.
(62) Arthur MJ Fibrogenesis II liver fibrosis.. Metalloproteinases and their inhibitors in. Am J Physiol Gastrointest Liver Physiol. 2000;279:G245–G249 Bachem MG, Meyer D, Schafer W. The response of rat liver. perisinusoidal lipocytes to polypeptide growth regulator changes with their trans-differentiation into myofibroblast- like cells in culture.. J Hepatol 1993;18:40–52. Benyon RC, Iredale JP, Goddard S, Winwood PJ, Arthur MJ Expression of tissue inhibitor of metalloproteinases 1 and 2 is increased in fibrotic human liver.. Gastroenterology. 1996;110:821-831 Bissell DM, Roll J. Connective tissue metabolism and hepatic fibrosis.. Hepatology:A Textbook of liver Disease. 2nd ed Brunt EM. Nonalcoholic steatohepatitis.. 1990;424-444. Liver Dis. 2004;24:3–20. Caballero ME, Berlanga J, Ramirez D, Lopez-Saura P, Gozalez R, Floyd RN, Marchbank T, Playford RJ Epidermal growth factor reduces multiorgan failure induced by thioacetamide Gut 2001;48:34–40 Castilla A, Prieto J, Fausto N. Transforming growth factor beta1 and. alpha in chronic liver diease. Effects of interferon alpha therapy. New England Journal of Medicine 1991;324:933-940 Castilla-Cortazear I, Garcia M, Muguuerza B, Quiroga J, Perez R, Santidrian S, Prieto J. Hepatoprotective effects of. insulin-like growth factor I in rats with carbon tetrachloride - induced cirrhosis.. Gastroenterology. 1997;113:1682-1691 Canturk NZ, Canturk Z, Ozden M, Dalcik H, Yardimoglu M, 61.
(63) Tulubas F Protective effect of IGF-1 on experimental liver cirrhosis-induced common bile duct ligation. Hepatogastroenterology 2003;50:2061-2066 Chen IC, Chang HC, Yang HW, Chen GI. Evaluation of total. antioxidant activity of several popular vegetables and chinese herbs:a fast approach with ABTS/H2O2/HRP system in microplates.. J of Food and Drug Analysi 2004;12:29-33. Cheng HY, Lin WC, Kiang FM, Wu LY, Peng WH. Anoectochilius. formosanus attenuates amnesia induced by scopolamine in rats. Chin Med. J. 2003;14:235-245. Chen CT, Chu CJ, Wang TF, Lu RH, Lee FY, Chang FY, Lin HC, Chan CC, Wang SS, Huang HC, Lee SD. Evidence against a role for. endotoxin in the hepatic encephalopathy of rats with thioacetamide-induced fulminant hepatic failure.. Journal of. Gastroenterology and Hepatology 2005;20:450–455 Cohick WS, Clemmons DR. The insulin- like growth factors.. Annu Rev Physiol 1993;55:131–153 Diez-Fernandez C, Bosca L, Fernandez-Simon L, Alvarez A, Cascales M Relationship between genomic DNA ploidy and parameters of liver damage during necrosis and regeneration induced by thioacetamide. Hepatology 1993;18:912–918 Du XM, Sun NY, Hayashi J, Chen Y, Sugiura M, Shoyama Y Hepatoprotective and antihyperliposis activities of in vitro cultured Anoectochilius formosanus.. Phytother Res 2003;17:30-33. Fausto N, Campbell JS, Riehle1 KJ Liver Regeneration. 62. Hepatology.
(64) 2006;43:S45-S53 Firth SM, Baxter RC. Cellular actions of the insulin-like growth factor. binding proteins.. Endocr Rev 2002;23:824–854. Friedman SL, Roll FJ, Boyles J, Bissell DM. Hepatic lipocytes: the. principal collagen-producing cells of normal rat liver. Proc Natl Acad Sci U.S.A. Friedman SL.. 1985;82:8681–8685. Liver fibrosis - from bench to bedside.. J Hepatol. 2003;38:S38–S53 Hanauske-Abel HM. Fibrosis:representive molecular elements, a basic. concept, and emerging targets for suppressive tretment. In: Hepatology:A textbook of liver disease. 1996;465-506 Hsieh MT, Wu CR, Chen CF. Gastrodin and p-hydoxybenzyl alcohol. facilitate memory consolidation and retrieval, but not acquisition, on the passive, avoidance tasks in rats. J Ethnopharmacol. 1997;56:45-54. Huang DD, Law RCS, Mak OT Effects of tissue- cultured Anoectochilus formosanus HAY.Extracts on the arachidonate metabolism, Bot. Bull. Academia Sinica. 1991;32:113-119 Ikeuchi M, Yamaguchi K, Nishimura T, Yazawa K. Effects of. Anoectochilus formosanus on endurance capacity in mice. J Nutr Sci Vitamiol. 2005;51:40-44. Inaba T, Saito H, Han I, Furukawa S, Matsuda T, Ikeda S, Muto T Growth hormone/insulin-like growth factor 1 axis alterations contribute to disturbed protein metabolism in cirrhosis patients after hepatectomy. J. Hepatol 1999 ;31:271-276 63.
(65) Iredale JP, Benyon RC, Arthur MJ, Ferris WF, Alcolado R, Winwood PJ, Clark N. Tissue inhibitor of metalloproteinase-1 messenger RNA. expression is enhanced relative to interstitial collagenase messenger RNA in experimental liver injury and fibrosis. Hepatology 1996;24:176-184 Ito K, Kasai R, Yamasaki K, Sugimoto H. Alphatic and aromatic. glucosides from Anoextochilus koshunensis.. Phytochemistry. 1993;33:1133-1137 Jia Y, Turek JJ. Altered NF-kappaB gene expression and collagen. formation induced by polyunsaturated fatty acids.. J Nutr Biochem. 2005;16:500-506 Jones JI, Clemmons DR Insulin-like growth factors and their binding proteins: biological actions. LaPaglia N, Steiner J, Kirsteins L. Endocr Rev 1995;16:3–34 Leptin alters the response of the. growth hormone releasing factor–growth hormone– insulin- like growth factor- 1 axis to fasting. J Endocrinol 1998;159:79 Laleman W, Vander-Elst I, Zeegers M, Servaes R, Libbrecht L, Roskams T, Fevery J, Nevens F. A stable model of cirrhotic. portal hypertension in the rat: thioacetamide revisited. European Journal of Clinical Investigation 2006;36:242–249 Lechuga CG, Herna´ndez-Nazara ZH,Rosales JD, Morris ER, Rinco´n AR, Rivas-Estilla AM, Esteban-Gamboa A, Rojkind1 M. TGF-β1. modulates matrix metalloproteinase-13 expression in hepatic stellate cells by complex mechanisms involving p38MAPK, PI3-kinase, AKT, and p70S6k. Am J Physiol Gastrointest Liver Physiol 64.
(66) 2004;287:G974–G987 Lin CC, Huang PC, Lin JM. Antioxidant and Hepatoprotective Effective. of Anoectochilus formosanus,Ganoderma lucidum and Gynostemma pentaphyllum.. Am J Chin Med 2000;28:87-96. Lin JM, Lin CC, Chih HF, Yang JJ, Lee SG Evalution of the antiinflammatory and liver- protective effects of Anoectochilus formosanus,Ganoderma lucidum and Gynostemma pentaphyllum in rats.. Am J Chin Med 1993;21:59-69. Li X, Benjamin IS, Alexander B. Reproducible production of. thioacetamide-induced macronodular cirrhosis in the rat with no mortality. J Hepatol 2002;36:488-493 Low YL, Leow CK, Salto-Tellez M, Chung MCM.. A proteomic. analysis of thioacetamide-induced hepatoxicity and cirrhosis in rat livers.. Proteomics. 2004;4:3960-3974.. Mangipudy RS, Chanda S, Mehendale HM. Hepatocellular regeneration:. key to thioacetamide autoprotection. Pharmacol Toxicol. 1995;77:182–188 Mendenhall CL, Chernausek SD, Ray MB, Gartside PS, Roselle GA, Grossman CJ, Chedid A. The interactions of insulin-like growth. factor I(IGF-I) with protein-calorie malnutrition in patients with alcoholic liver disease: V.A. Cooperative Study on Alcoholic Hepatitis VI.. Alcohol 1989;24:319-29. Mirpuri E, Garcia-Trevijano ER, Castilla-Cortazear, Berasain C, Quiroga J, Rodriguez-Ortigosa C, Mato JM, Prieto J, Avila MA. Altered. liver gene expression in CCl4-cirrhotic rats is partially normalized by insulin-like growth factor-I. Int J Biochem Cell Biol 65.
(67) 2002;34:242-52 Mohn KL, Melby AE, Tewari DS, Laz TM, Taub R. The gene encoding. rat insulin-like growth factor-binding protein 1 is rapidly and highly induced in regenerating liver.. Mol Cell Biol 1991;11:1393–1401. Murawaki Y, Ikuta Y, Idobe Y, Kitamura Y,Kawasaki H. Tissue. inhibitor of metalloproteinase-1 in the liver of patients with chronic liver disease.. J Hepatol 1997;26:1213-1219. Murphy FR, Issa R, Zhou X, Ratnarajah S, Nagase H, Arthur MJ, Benyon C. Inhibition of apoptosis of activated hepatic stellate cells by. tissue inhibitor of metalloproteinase-1 is mediated via effects on matrix metalloproteinase inhibition. Implications for reversibility of liver fibrosis.. J Biol Chem. 2002;277:11069-11076. Nagy P, Schaff Z, Lapis K, Immunohistochemical detection of transforming growth factor-β1 in fibrotic liver disease.. Hepatology. 1991;14:269-273 Novosyadlyy R, Tron K, Dudas J, Ramadori G Scharf JG. Expression. and regulation of the insulin-like growth factor axis components in rat liver myobroblasts. J Cell Physiol Novosyadlyy R, DargelR, Scharf JG. 2004;199:388–398. Expression of insulin-like growth. factor-I and insulin-like growth factor binding proteins during thioacetamide- induced liver cirrhosis in rats.. Growth Hormone &. IGF Research 2005;15:313–323 Picardi A, Deoliveira AC, Muguerza B. Low doses of insulin-like. growth factor-I improve nitrogen retention and food efficiency in rats with early cirrhosis.. J Hepatol 1997;26:191–202. Pinzani M, Abboud HE, Aron DC Secretion of insulin-like growth 66.
(68) factor-I and binding proteins by rat liver fat-storing cells: regulatory role of platelet-derived growth factor.. Endocrinology. 1990;127:2343–2349 Popper H, Uenfriend S. Hepatic fibrosis. Correlation of biochemical and. morphologic investigations. Poynard T. Am J Med 1970;49:707–721. Natural history of HCV infection. Baillieres Best Pract. Res Clin Gastroenterol 2000;14:211–228 Poynard T, Bedossa P, Opolon P. Natural history of liver fibrosis. progression in patients with chronic hepatitis C.. Lancet. 1997;349:825–832 Ramaiah SK, Apte U, Mehendale HM. Cytochrome P4502E1 induction. increases thioacetamide liver injury in diet- restricted rats.. Drug. Metab Dispos 2001;29:1088–1095 Rechler MM, Insulin-like growth factor binding proteins. Vitam Horm. 1993;47:1–114. Sanz S, Pucilowska JB, Liu S, Rodriguez-Ortigosa CM, Lund PK, Brenner DA, Fuller CR, Simmons JG, Pardo A, Martinez- Chantar ML, Fagin JA, Prieto J. Expression of insulin-like growth factor I. by activated hepatic stellate cells reduces fibrogenesis and enhances regeneration after liver injury. Gut Schaffner F, Klion FM. 2005;54:134–141. Chronic hepatitis.. Annu Rev Med. 1968;19:25–38 Scharf JG, Knittel T, Dombrowski F. Characterization of the IGF axis. components in isolated rat hepatic stellate cells. Hepatology 1998;27:1275–1284 Scharf JG, Dombrowski F, Ramadori G 67. The IGF axis and.
數據



+7

相關文件
vs Functional grammar (i.e. organising grammar items according to the communicative functions) at the discourse level2. “…a bridge between
Myers effect condensation of mean field D(-1) Chern Simons term is induced. Fuzzy sphere is
The growth in the number of vanco- mycin-induced thrombocytopenia cases presently seen may be associated with the increased use of the drug, especially in multiresistant patients
Higher immunoexpression of HIF-1 a, NOTCH1, ADAM-12, and heparin-binding epidermal growth factor like growth factor (HB-EGF) in epidermoid cells in compari- son with mucous cells
Population migration, the other factor in determining population growth, recorded an estimated inflow of 21 811 persons in 2005, including legal and illegal immigrants from
Since the assets in a pool are not affected by only one common factor, and each asset has different degrees of influence over that common factor, we generalize the one-factor
台灣金線連( Anoectochilus formosanus Hayata )在民間是一種極珍貴的藥材。在「2004 國際藥用
Parry,“ Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures, ” Aquaculture, Vol.211, No.1-4,
