Running Head: antinociceptive and anti-inflammatory activities of asiatic acid 1
2
Antinociceptive activities and the mechanisms of anti-inflammation of
3asiatic acid in mice
45
Shyh-Shyun Huang1, Chuan-Sung Chiu1,2,#, Hsien-Jung Chen3,#, Wen-Chi Hou4, 6
Ming-Jyh Sheu5, Ying-Chih Lin6, Pei-Hsin Shie1, Guan-Jhong Huang1,*
7 8
1
School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, College 9
of Pharmacy, China Medical University, Taichung 404, Taiwan 10
2
Nursing Department, Hsin Sheng College of Medical Care and Management, Taoyuan 11
325, Taiwan 12
3
Department of Biological Sciences, National Sun Yat-sen University, Kaohsiung 804, 13
Taiwan 14
4
Graduate Institute of Pharmacognosy, Taipei Medical University, Taipei, Taiwan 15
5
School of Pharmacy, China Medical University, Taichung 404, Taiwan 16
6
Department of Optometry, Jen-Teh Junior College of Medicine, Nursing and
17 Management,Miaoli, Taiwan 18 19 * Corresponding author 20 Dr. Guan-Jhong Huang 21
Tel.: +886-4-2205-3366 ext 5508; fax: +886-4-2208-3362. 22
E-mail address: [email protected] 23
24 #
These authors are equal to this work 25
26 27
Abstract
28
Asiatic acid (AA), a pentacyclic triterpene compound identified in the medicinal plant 29
Centella asiatica, was evaluated for the antinociceptive and anti-inflammatory effects.
30
Treatment of male ICR mice with AA (1, 5, and 10 mg/kg) significantly inhibited the 31
numbers of acetic acid-induced writhing response in 10 minutes. Also, our result showed 32
that AA (10 mg/kg) significantly inhibited the formalin-induced pain in the late phase (p 33
< 0.001). In the anti-inflammatory test, AA (10 mg/kg) decreased the paw edema at the 34
fourth and fifth h after λ-carrageenan (Carr) administration and increased the activities of 35
catalase (CAT), superoxide dismutase (SOD) and glutathione peroxidase (GPx) in the 36
liver tissue. We also demonstrated that AA significantly attenuated the malondialdehyde 37
(MDA) level in the edema paw at the fifth h after Carr injection. AA decreased the nitric 38
oxide (NO), tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β) levels on serum 39
level at the fifth h after Carr injection. Western blotting revealed that AA (10 mg/kg) 40
decreased Carr-induced inducible nitric oxide synthase (iNOS), cycloxyclase (COX-2) 41
and nuclear factor-κB (NF-κB) expressions at the fifth h in the edema paw. An 42
intraperitoneal (i.p.) injection treatment with AA also diminished neutrophil infiltration 43
into sites of inflammation as did indomethacin (Indo). The anti-inflammatory 44
mechanisms of AA might be related to the decrease in the level of MDA, iNOS, COX-2, 45
and NF-κB in the edema paw via increasing the activities of CAT, SOD, and GPx in the 46
liver through the suppression of NO, TNF-α, and IL-1β. 47
48
Key words: Chinese medicine- asiatic acid-anti-inflammation-NO-TNF-α.
49 50
Introduction
51
Triterpenes are biosynthesized in plants by the cyclization of squalene, and are widely 52
distributed in the plant kingdom. Moreover, their biological activities have attracted much 53
attention. Many triterpenoids have shown promising effects when applied as 54
anti-inflammatory agents (1). In particular, AA is a member of the ursane-type 55
triterpenoids and is derived from the medicinal plant Centella asiatica, which is used as a 56
medicine in tropical regions (2). AA has been found to prevent UVA-mediated 57
photoaging, to inhibit β-amyloid-induced and glutamate-induced neurotoxicity, and to 58
possess anti-ulcer and anti-hepatofibric activities (3). In addition, it has been reported to 59
exhibit a cytotoxic effect on liver, colon and breast cancer cells (4), and neuroprotective
60
in a mouse model of focal cerebral ischemia (5).
61
Carr-induced paw edema is a useful model to assess vascular changes associated with 62
inflammation. Subplantar injections of Carr in mice induce a biphasic edema. The first 63
phase peaks at 3 h and the delayed phase peaks at 48 h after Carr injection. In the early 64
phase, there is a diffuse cellular infiltrate with polymorphonuclear leukocytes (PMNs), 65
whereas the infiltrate of the delayed phase is composed by macrophages, eosinophils and 66
lymphocytes (6). The inflammatory effect induced by Carr could be associated with free 67
radical on. Free radical, prostaglandin and NO will be released when administrating with 68
Carr for 1~5 h. The edema effect was raised to maximum at the third h and its MDA 69
production was due to free radical attack plasma membrane (6). Thus, inflammatory 70
effect would result in the accumulation of MDA. Therefore, in this paper we examined 71
the analgesic effects of AA on nociception induced by acetic acid and formalin. We also 72
evaluated the anti-inflammatory effects of AA on paw edema induced by Carr in mice, 73
and we detected the levels of MDA, NO, TNF-α, iNOS and COX-2 in either paw edema 74
or serum. Also, the activities of CAT, SOD and GPx in the liver at the fifth h after Carr 75
injection were investigated to understand the relationship between the anti-inflammatory 76
mechanism of the AA and antioxidant enzymes. 77 78 Methods 79 Chemicals 80
Asiatic acid, Carr and indomethacin (Indo) were obtained from Sigma (St. Louis, MO,
81
USA). Acetic acid was purchased from Merck (Darmstadt, Germany). Formalin was 82
purchased from Nihon Shiyaku Industries (Japan). TNF-α and IL-1β were purchased 83
from Biosource International Inc. (Camarillo, CA, USA). Anti-iNOS, anti-COX-2, 84
anti-NF-κB (p50), and anti-β-actin antibody (Santa Cruz, USA) and a protein assay kit 85
(Bio-Rad Laboratories Ltd., Watford, Herts, U.K.) were obtained as indicated. Poly 86
(vinylidene fluoride) membrane (Immobilon-P) was obtained from Millipore Corp. 87
(Bedford, MA, USA). 88
89
Animals
90
6-8 weeks male ICR mice were obtained from the BioLASCO Taiwan Co., Ltd. The 91
animals were kept in plexiglass cages at a constant temperature of 22 ±1°C, relative 92
humidity 55 ± 5 % with 12 h dark-light cycle for at least 2 week before the experiment. 93
They were given food and water ad libitum. All experimental procedures were performed 94
according to the NIH Guide for the Care and Use of Laboratory Animals. And all tests 95
were conducted under the guidelines of the International Association for the Study of 96
Pain (7). 97
After a 2-week adaptation period, male ICR mice (18-25 g) were randomly assigned 98
to five groups (n=6) of the animals in acetic acid-induced writhing (1%, 0.l mL/10 g i.p.) 99
and formalin-induced licking (5%, 20 µL/per mice i.p.) experiments. These include a 100
pathological model group (received acetic acid or formalin), a positive control (acetic 101
acid or formalin + Indo), and the AA administered groups (acetic acid or formalin+ AA: 102
1, 5, and 10 mg/kg). In the Carr-induced edema experiment, there were randomly 103
assigned to six groups (n=6) of the animals in the study. The control group receives 104
normal saline (i.p.). The other five groups include a Carr-treated, a positive control (Carr 105
+ Indo) and AA administered groups (Carr + AA: 1, 5, and 10 mg/kg). 106
107
Acetic acid-induced writhing response
108
The test was performed as described by Chang et al., (8). Writhing was induced by an 109
intraperitoneal (i.p.) injection of 0.1 mL/10 g acetic acid solution (10 mL/kg). Positive 110
control animals were pretreated with Indo (10 mg/kg, i.p.) 25 min before acetic acid. 111
Each AA administered group was pretreated with 1 mg/kg, 5 mg/kg, or 10 mg/kg 112
(dissolved in 0.5% carboxymethylcellulose) i.p. 25 min before acetic acid. Five minutes 113
after the i.p. injection of acetic acid, the number of writhing and stretching was recorded. 114
115
Formalin test
116
The antinociceptive activity of the drugs was determined using the formalin test (8). 117
Twenty microliters of 5% formalin was injected into the dorsal surface of the right hind 118
paw of mice 30 min after i.p. administration of AA (1, 5, and 10 mg/kg), or Indo. The 119
mice were observed for 30 min after the injection of formalin, and the amount of time 120
spent licking the injected hind paw was recorded. The first 5 min post formalin injection 121
is referred to as the early phase and the period between 15 min and 40 min as the late 122
phase. The total time spent licking or biting the injured paw (pain behavior) was 123
measured with a stop watch. The activity was recorded in 5 min intervals. 124 125 λ λλ λ-carrageenin-induced edema 126
Carr-induced hind paw edema model was used for determination of anti-inflammatory 127
activity (8). Animals were i.p. treated with AA (1, 5, and 10 mg/kg), Indo or normal 128
saline, 30 min prior to injection of 1% Carr (50 µL) in the plantar side of right hind paws 129
of the mice. Paw volume was measured immediately after Carr injection and at 1, 2, 3, 4, 130
and 5 h intervals after the administration of the edematogenic agent using a 131
plethysmometer (model 7159, Ugo Basile, Varese, Italy). The degree of swelling induced 132
was evaluated by the ratio a/b, where is the volume of the right hind paw after Carr 133
treatment, and b is the volume of the right hind paw before Carr treatment. Indo was used 134
as a positive control. After 5 hrs, the animals were sacrificed; the Carr-induced edema 135
feet were dissected and stored at -80 ºC. Also, blood were withdrawn and kept at -80 ºC. 136
The protein concentration of the sample was determined by the Bradford dye-binding 137
assay (Bio-Rad, Hercules, CA). 138
139
MDA assay
140
MDA from Carr-induced edema foot was evaluated by the thiobarbituric acid reacting 141
substances (TRARS) method (8). Briefly, MDA reacted with thiobarbituric acid in the 142
acidic high temperature and formed a red-complex TBARS. The absorbance of TBARS 143
was determined at 532 nm. 144
145
Measurement of Nitric oxide/Nitrite
146
NO production was indirectly assessed by measuring the nitrite levels in serum 147
determined by a colorimetric method based on the Griess reaction (8). Serum samples 148
were diluted four times with distilled water and deproteinized by adding 1/20 volume of 149
zinc sulfate (300 g/L) to a final concentration of 15 g/L. After centrifugation at 10,000×g 150
for 5 min at room temperature, 100 µL supernatant was applied to a microliter plate well, 151
followed by 100 µL of Griess reagent (1% sulfanilamide and 0.1% 152
N-1-naphthylethylenediamine dihydrochloride in 2.5% polyphosphoric acid). After 10
153
min of color development at room temperature, the absorbance was measured at 540 nm 154
with a Micro-Reader (Molecular Devices, Orleans Drive, Sunnyvale, CA). By using 155
sodium nitrite to generate a standard curve, the concentration of nitrite was measured by 156
absorbance at 540 nm. 157
158
Measurement of serum TNF-α and IL-1ββββ by ELISA
159
Serum levels of TNF-α and IL-1β were determined using a commercially available
160
enzyme linked immunosorbent assay (ELISA) kit (Biosource International Inc., 161
Camarillo, CA) according to the manufacturer’s instruction. TNF-α and IL-1β were
162
determined from a standard curve. The concentrations were expressed as pg/mL. 163
Antioxidant enzyme activity measurements
165
The following biochemical parameters were analyzed to check the hepatoprotective 166
activity of AA by the methods given below. 167
Total SOD activity was determined by the inhibition of cytochromec reduction (9). The 168
reduction of cytochrome c was mediated by superoxide anions generated by 169
xanthine/xanthine oxidase system and monitored at 550 nm. One unit of SOD was 170
defined as the amount of enzyme requiredto inhibit the rate of cytochrome c reduction by 171
50%. Total CAT activity was based on that of Aebi (10). In brief, the reduction of 10 mM 172
H2O2 in 20 mM of phosphate buffer (pH 7.0) was monitored by measuring the absorbance 173
at 240 nm. The activity was calculated using a molar absorption coefficient, and the 174
enzyme activity was defined as nmoles of dissipating hydrogen peroxide per mg protein 175
per min. Total GPx activity in cytosol was determined according to Paglia and 176
Valentine’s method (11). The enzyme solution was added to a mixture containing 177
hydrogen peroxide and glutathione in 0.1 mM Tris buffer (pH 7.2) and the absorbance at 178
340 nm was measured. Activity was evaluated from a calibration curve, and the enzyme 179
activity was defined as nmoles of NADPH oxidized per mg protein per min. 180
181
Western blot analysis of iNOS, COX-2, and NF-κκκκB
182
Soft tissues were removed from individual mice paws and homogenized in a solution 183
containing 10 mM CHAPS, 1mM phenylmethylsulphonyl fluoride (PMSF), 5 µg/mL, 184
aprotinin, 1 µM pepstatin and 10 µM leupeptin. The homogenates were centrifuged at 185
12,000g for 20 min, and 30 µg of protein from the supernatants was then separated on 186
10% sodium dodecylsulphate–polyacrylamide gel and transferred to polyvinylidene 187
difluoride membranes. Following transfer, the membrane was blocked for 2 h at room 188
temperature with 5% skim milk in Tris-buffered saline-Tween (TBST; 20 mM Tris, 500 189
mM NaCl, pH 7.5, 0.1% Tween 20). The membranes were then incubated with mouse 190
monoclonal anti-iNOS, anti-COX-2 or anti-NF-κB (p50) antibody in 5% skim milk in 191
TBST for 2 h at room temperature. The membranes were washed three times with TBST 192
at room temperature and then incubated with a 1 : 2000 dilution of anti-mouse IgG 193
secondary antibody conjugated to horseradish peroxidase (Sigma, St Louis, MO, U.S.A.) 194
in 2.5% skim milk in TBST for 1 h at room temperature. The membranes were washed 195
three times and the immunoreactive proteins were detected by enhanced 196
chemiluminescence (ECL) using hyperfilm and ECL reagent (Amersham International 197
plc., Buckinghamshire, U.K.). The results of Western blot analysis were quantified by 198
measuring the relative intensity compared to the control using Kodak Molecular Imaging 199
Software (Version 4.0.5, Eastman Kodak Company, Rochester, NY) and represented in 200
the relative intensities. 201
202
Histological examination
203
For histological examination, biopsies of paws were taken 5 h following the 204
interplanetary injection of Carr. The tissue slices were fixed in (1.85% formaldehyde, 1% 205
acetic acid) for 1 week at room temperature, dehydrated by graded ethanol and embedded 206
in Paraffin (Sherwood Medical). Sections (thickness 5 µm) were deparaffinized with 207
xylene and stained with H & E stain. All samples were observed and photographed with 208
Nikon microscopy. Every 3~5 tissue slices were randomly chosen from Carr, Indo and 209
AA-treated (10 mg/kg) groups. Histological examination of these tissue slices revealed an
excessive inflammatory response with massive infiltration of PMNs by microscope. The 211
numbers of neutrophils were counted in each scope (400 x) and thereafter obtain their 212
average count from 5 scopes of every tissue slice. 213
214
Statistical analysis
215
Data are expressed as mean ± S.E.M. Statistical evaluation was carried out by one-way 216
analysis of variance (ANOVA followed by Scheffe's multiple range test). Statistical 217
significance is expressed as *p < 0.05, **p < 0.01, and ***p < 0.001.
218 219
Results
220
Effects of AA on acetic-induced writhing response
221
The cumulative amount of abdominal stretching correlated with the level of acetic acid 222
induced pain (Fig. 2). AA treatment (1 mg/kg) significantly inhibited the number of 223
writhing in comparison with the normal controls (p <0.05). AA (5 or 10 mg/kg) further 224
reduced the number of writhing (p < 0.01 or p < 0.001) and AA (10 mg/kg) demonstrates 225
more inhibition than Indo (10 mg/kg). 226
227
Formalin test
228
AA (1 mg/kg) significantly (p < 0.05) inhibited formalin-induced pain in the late phase
229
(Fig. 3); however, it did not show any inhibition in the early phase. The positive control 230
Indo (5 or 10 mg/kg) also significantly (p < 0.01 or p < 0.001) inhibited the formalin 231
induced pain in the late phase. 232
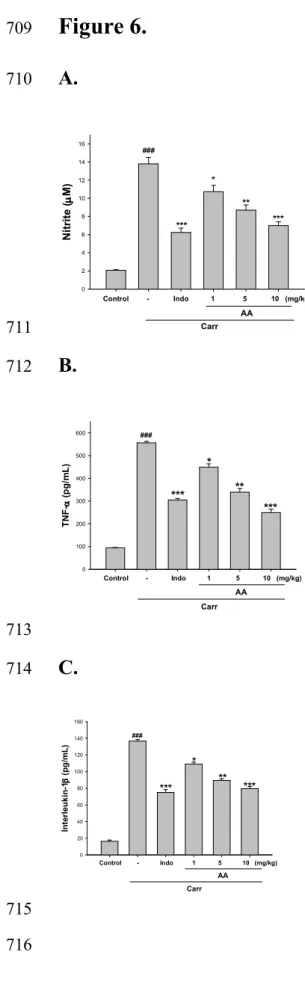
Effects of AA on λλλλ-Carrageenan-induced mice paw edema 234
As shown in Fig. 4, Carr induced paw edema. AA (5 or 10 mg/kg) inhibited (p < 0.01 or 235
p < 0.001) the development of paw edema induced by Carr after 4 and 5 h of treatment,
236
significantly. Indo (10 mg/kg) significantly decreased the Carr induced paw edema after 4 237
and 5 h of treatment (p < 0.001). 238
239
Effects of AA on MDA level
240
MDA level increased significantly in the edema paw at the 5 h after Carr injection (p < 241
0.001). However, MDA level was decreased significantly by treatment with AA (5 mg/kg) 242
(p < 0.001), as well as 10 mg/kg Indo (Fig. 5). 243
244
Effects of AA on NO level
245
In Fig. 6A, the NO level increased significantly in the edema serum at the 5 h after Carr 246
injection (p < 0.001). AA (5 or 10 mg/kg) significantly decreased the serum NO level (p 247
< 0.01 or p < 0.001). The inhibitory potency was similar to that of Indo (10 mg/kg) at 5th 248
h after induction. 249
250
Effects of AA on TNF-α and IL-1ββββ levels.
251
TNF-α and IL-1β levels increased significantly in serum at the 5th h after Carr injection 252
(p < 0.001). However, AA (5 or 10 mg/kg) decreased the TNF-α and IL-1β levels in 253
serum at the 5th h after Carr injection (p < 0.01 or p < 0.001), as well as 10 mg/kg Indo 254
(Fig. 6B and 6C). 255
Effects of AA on activities of antioxidant enzymes
257
The acute inflammatory response is associated with the production of reactive oxygen 258
species (ROS) such as superoxide anions, hydrogen peroxide and peroxynitrite. In a 259
number of pathophysiological conditions associated with inflammation or oxidant stress, 260
these ROS have been proposed to mediate cell damage in the liver (1). At the 5th h 261
following the intrapaw injection of Carr, liver tissues were analyzed for the biochemical 262
parameters such as CAT, SOD and GPx activities (Table 1). CAT, SOD and GPx 263
activities in liver tissue were significantly decreased by Carr administration. CAT, SOD, 264
and GPx activity were increased significantly after treated with 10 mg/kg AA and 10 265
mg/kg Indo (P<0.01) (Table 1). 266
267
Effects of AA on λλλ-Carrageenan-induced iNOS, COX-2, λ and NF-κκκB κ protein
268
expressions in mice paw edema
269
Transcription of pro-inflammatory mediators such as iNOS, COX-2, TNF-α, and IL-1β is
270
regulated by activation of transcription factor NF-κB (Kubes and McCafferty, 2000). The 271
effect of AA on iNOS, COX-2, and NF-κB protein expression was studied by western 272
blot. Equal amounts of protein (30 µg/lane) were resolved by SDS-PAGE and then 273
transferred to a nitrocellulose membrane and iNOS, COX-2, and NF-κB were detected 274
using a specific antibody. The results showed that injection AA (10 mg/kg) on 275
Carr-induced for 5 h inhibited iNOS, COX-2 and NF-κB proteins expression in mouse 276
paw edema (Fig. 7A). The detection of β-actin was also performed in the same blot as an 277
internal control. The intensity of protein bands was analyzed using Kodak Quantity 278
software (Molecular Imaging Software System, Kodak) in three independent experiments 279
and showed an average of 77.6%, 72.4%, and 62.8% down-regulation of iNOS, COX-2, 280
and NF-κB protein, respectively, after the treatment with AA at 10 mg/kg compared with 281
the Carr-induced alone (Fig. 7B). And the protein expression showed an average of 282
43.6%, 41.1%, and 36.4% down-regulation of iNOS, COX-2, and NF-κB protein after 283
treatment with Indo at 10 mg/kg compared with the Carr-induced alone (Fig. 7B). The 284
down-regulation of iNOS, COX-2, and NF-κB activity of AA (10 mg/kg) was better than 285 Indo (10 mg/kg). 286 287 Histological examination 288
Paw biopsies of Carr model animals showed marked cellular infiltration in the connective 289
tissue. The infiltrates accumulated between collagen fibers and into intercellular spaces. 290
Paw biopsies of animals treated with AA (10 mg/kg) showed a reduction in inflammatory 291
response Carr-induced. Inflammatory cells were actually reduced in number and confined 292
to near the vascular areas. Intercellular spaces did not show any cellular infiltrations. 293
Collagen fibers were regular in shape and showed a reduction of intercellular spaces. 294
Moreover, the hypoderm connective tissue was not damaged (Fig. 8). Neutrophils were 295
notified increased with Carr treatment (P < 0.001). As Indo and AA (10 mg/kg) could 296
significantly decrease the neutrophils numbers as compared to the Carr-treated group (P 297 < 0.001) (Fig. 8E). 298 299 Discussion 300
We have evaluated the putative analgesic and anti-inflammatory activities of AA to 301
clarify the pain and inflammation relieving effects. Two different analgesic testing 302
methods were employed with the objective of identifying possible peripheral and central 303
effects of the test substances. The acetic writhing test is normally used to study the 304
peripheral analgesic effects of drugs. Although this test is nonspecific (e.g., 305
anticholinergic, antihistaminic and other agents also show activity in the test), it is widely 306
used for analgesic screening (12). In our study, we found that AA (1, 5, and 10 mg/kg) 307
exhibited antinociceptive effect in acetic acid-induced writhing response (Fig. 2). This 308
effect may be due to inhibition of the synthesis of the arachidonic acid metabolites (13). 309
The in vivo model of pain, formalin-induced paw pain has been well established as a 310
valid model for analgesic study. It is well known that the formalin test produces a distinct 311
biphasic nociception, a first phase (lasting the first 5 min) corresponding to acute 312
neurogenic pain, and a second phase (lasting from 15 to 30 min after injection of formalin) 313
corresponding to inflammatory pain responses (14). Therefore, the test can be used to 314
clarify the possible mechanism of an antinociceptive effect of a proposed analgesic. 315
Centrally acting drugs such as opioids inhibit both phases equally, but peripherally acting 316
drugs such as aspirin, Indo and dexamethasone only inhibit the late phase (15). The 317
inhibitory effect of AA on the nociceptive response in the late phase of the formalin test 318
suggested that the anti-nociceptive effect of AA could be due to its peripheral action (Fig. 319
3). 320
The injection of Carr in mice produces a typical biphasic edema associated with the 321
production of several inflammatory mediators, such as bradykinin, prostaglandins, nitric 322
oxide, and cytokines. The Carr test is highly sensitive to nonsteroidal antiinflammatory 323
drugs, and has long been accepted as a useful phlogistic tool for investigating new drug 324
therapies (16). The degree of swelling of the Carr-injected paws was maximal at 3 th after 325
injection. Statistical analysis revealed that AA (10 mg/kg) and Indo significantly 326
inhibited the development of edema at 4 th after treatment (p<0.001) (Fig. 4). They both 327
showed anti-inflammatory effects in Carr-induced mice edema paw. It is well known that 328
the third phase of the edema-induced by Carr, in which the edema reaches its highest 329
volume, is characterized by the presence of prostaglandins and other compounds of slow 330
reaction (17) found that the injection of Carr into the rat paw induces the liberation of 331
bradykinin, which later induces the biosynthesis of prostaglandin and other autacoids, 332
which are responsible for the formation of the inflammatory exudates. In addition, the 333
classification of antinociceptive drugs is usually based on their mechanism of action 334
either on the central nervous system or on the peripheral nervous system (18). 335
NO plays an important role in Carr induced paw edema. iNOS is expressed in this 336
model within 4 h after injection of Carr. The subsequent production of NO maintains the 337
edema. In the studies of mechanism on the inflammation, L-arginine–NO pathway has 338
been proposed to play an important role in the Carr-induced inflammatory response (19). 339
Our present results also confirm that Carr-induced paw edema model results in the 340
production of NO. The expression of the inducible isoform of NO synthase has been 341
proposed as an important mediator of inflammation (20). In our study, the level of NO 342
was decreased significantly by treatment with 1, 5 and 10 mg/kg AA. We suggest the 343
mechanism of anti-inflammatory of AA may be through the L-arginine–NO pathway 344
since AA significantly inhibits the NO production (Fig. 6A). 345
TNF-α is a major mediator in inflammatory responses, inducing innate immune 346
responses by activating T cells and macrophages, and stimulating secretion of other 347
inflammatory cytokines (21). Also, TNF-α is a mediator of Carr-induced inflammatory 348
incapacitation, and is able to induce the further release of kinins and leukotrienes, which 349
is suggested to have an important role in the maintenance of long-lasting nociceptive 350
response. IL-1β is also important in the regulation of the inflammatory response.
351
Moreover, IL-1β increases the expression of adhesion factors on endothelial cells to
352
enable transmigration of leukocytes, and is associated with hyperalgesia and fever (22).
353
In this study, we found AA decreased the TNF-α and IL-1β levels in serum after Carr
354
injection by treatment with 1, 5, and 10 mg/kg AA, significantly (Fig. 6B and 6C).
355
AA is one of the most common triterpenes and has a variety of pharmacological
356
activities (23). Nonetheless, little information is available with respect to the molecular 357
mechanisms underlying the anti-inflammatory effect of AA. The inhibitory effects of AA 358
and asiaticoside on the LPS-induced pro-inflammatory molecules, including NO and 359
prostaglandin E2, and found that AA is a more potent inhibitor than asiaticoside. These 360
results suggest that the anti-inflammatory properties of AA might be the results from the 361
inhibition of iNOS, COX-2, interleukin-6, IL-1β and TNF-α expression through the 362
down-regulation of Nuclear factor-kappa B activation via suppression of IκB kinase and 363
mitogen-activated protein kinase (p38, ERK1/2 and JNK) phosphorylation in RAW264.7 364
cells (24). 365
The Carr-induced inflammatory response has been linked to neutrophils infiltration 366
and the production of neutrophils-derived free radicals as well as the release of other 367
neutrophils-derived mediators (8). Some researches demonstrate that inflammatory effect 368
induced by Carr is associated with free radicals. Free radicals, prostaglandin and NO will 369
be released when administrating with Carr for 1-6 h. The edema effect was raised to the 370
maximum at the third h. MDA production is due to free radical attack plasma membrane. 371
Thus, inflammatory effect would result in the accumulation of MDA. GSH is a known 372
oxyradical scavenger. Enhancing the level of GSH conducive toward favor reduces MDA 373
the production. Endogenous GSH plays an important role against Carr-induced local 374
inflammation. In a number of pathophysiological conditions associated with 375
inflammation or oxidant stress, these ROS have been proposed to mediate cell damage 376
via a number of independent mechanisms including the initiation of lipid peroxidation, 377
the inactivation of a variety of antioxidant enzymes and depletion of glutathione. Giving 378
the importance of the oxidative status in the formation of edema, the anti-inflammatory 379
effect exhibited by drug in this model might be related to its antioxidant properties (8). In 380
this study, there are significantly increases in CAT, SOD and GPx activities with AA 381
treatment (Table 1). Furthermore, there are significant decreases in MDA level with AA 382
treatment (Fig. 5). We assume the suppression of MDA production is probably due to the 383
increases of CAT, SOD and GPx activities. 384
During inflammatory processes, large amounts of the proinflammatory mediators, NO 385
and PGE2, are generated by inducible iNOS and COX-2, respectively (25). INOS, is 386
generally not present in resting cells, but is induced by various stimuli, which include 387
bacterial LPS, TNF-α, IL-1β and interferon-γ (26). However, COX-2 is induced by 388
pro-inflammatory stimuli, including LPS and cytokines in cells in vitro and in inflamed 389
sites in vivo. Furthermore, COX-2 is believed to be the isoform responsible for the 390
production of pro-inflammatory prostaglandins (PGs) in various models of inflammation 391
(27). In this study, there are significantly decreased in iNOS and COX-2 activities with 392
AA treatment (Fig. 7A). We assume the suppression of NO production is probably due to
393
the decreases of iNOS and COX-2 activities. An inflammatory response implicates 394
macrophages and neutrophils, which secrete a number of mediators (eicosinoids, oxidants, 395
cytokine and lytic enzymes) responsible for initiation, progression and persistence of 396
acute or chronic state of inflammation (28). NO is the most important among these 397
mediators and is produced in macrophages by COX-2 and iNOS, respectively (29). COXs
398
are pro-inflammatory enzymes that are involved in arachidonic acid metabolism and
399
influence biological reactions such as tissue repair and immune responses, all of which
400
are associated with inflammation. COX-1 and COX-2 are the rate-limiting enzymes in the
401
synthesis of PGE2. COX-1 is constitutively expressed and involved in the acute
402
inflammatory response, whereas COX-2 is expressed in specific cells (i.e., macrophages,
403
monocytes, and neutrophils) after stimulation COX-2-dependent PGE2 is produced by
404
inflammatory cells and increased in disease (30).
405
NF-κB is known to be a major transcription factor to regulate the expressions of
406
pro-inflammatory enzymes and cytokines, such as iNOS, COX-2, and TNF-α (31).
407
NF-κB subunits (p65 and/or p50) are normally sequestered in the cytosol as an inactive
408
complex by binding to inhibitory factor IκB-α in un-stimulated cells. Upon stimulation of
409
pro-inflammatory signals including LPS, IκB-α is phosphorylated by IκB kinase (IKK)
410
and inactivated through ubiquitin-mediated degradation. The resulting free NF-κB is
411
translocated into the nucleus and acts as a transcription factor. As shown in Fig. 7A, the
412
treatment with AA blocks the degradation of NF-κB in Carr-induced paw edema.
413
Therefore, these results suggest that AA inhibits the expression of iNOS and COX-2, and
414
thus NO production through inactivation of NF-κB activation.
415
NO also is responsible for vasodilatation, increase in vascular permeability and 416
edema formation at the site of inflammation (32). NO along with superoxide (O2‧−) and 417
the products of their interaction, also initiates a wide range of toxic oxidative reactions 418
causing tissue injury (33). Likewise, the neutrophils produce oxidants and release 419
granular constituents comprising of lytic enzymes performing important role in 420
inflammatory injury (34). In this study, AA inhibition in the release of these mediators is 421
a potential strategy to control inflammation and is implicated in mechanism of action as 422
shown in Fig. 9. 423
In conclusion, these results suggested that AA possessed analgesic and 424
anti-inflammatory effects. The anti-inflammatory mechanism of AA may be related to 425
iNOS and associated with the increase in the activities of antioxidant enzymes (CAT, 426
SOD and GPx). AA may be used as a pharmacological agent in the prevention or 427
treatment of disease in which free radical formation is a pathogenic factor. 428
429
Acknowledgements
430
The authors want to thank the financial supports from the National Science Council (NSC 431
97-2313-B-039-001-MY3) and China Medical University (CMU) (CMU95-PH-11, 432
CMU96-113, CMU97-232, and CMU99-S-29). The authors would like to thank Dr 433
Jeffrey Conrad for criticallyreading the manuscript. 434
435
References
436
1. Huang GJ,Huang SS,Lin SS,Shao YY,Chen CC,Hou WC, et al. Analgesic effects and 437
the mechanisms of anti-inflammation of ergostatrien-3β-ol from Antrodia camphorata 438
submerged whole broth in mice. J Agric Food Chem 2010; 58:7445–7452. 439
2. Coldren CD, Hashim P, Ali JM, Oh SK, Sinskey AJ, Rha C. Gene expression changes 440
in the human fibroblast induced by Centella asiatica triterpenoids. Planta Med 2003; 441
69:725–732. 442
3. Dong MS, Jung SH, Kim HJ, Kim JR, Zhao LX, Lee ES, et al. Structure-related 443
cytotoxicity and anti-hepatofibric effect of asiatic acid derivatives in rat hepatic 444
stellate cell-line, HSC-T6. Arch Pharm Res 2004;27:512–517. 445
4. Lee YS, Jin DQ, Kwon EJ, Park SH, Lee ES, Jeong TC, et al. Asiatic acid, a triterpene, 446
induces apoptosis through intracellular Ca2+ release and enhanced expression of p53 in 447
HepG2 human hepatoma cells. Cancer Lett. 2002;186:83–91. 448
5. Krishnamurthy RG, Senut MC, Zemke D, Min J, Frenkel MB, Greenberg EJ, Yu SW, 449
Ahn N, Goudreau J, Kassab M, Panickar KS, Majid A. Asiatic acid, a pentacyclic 450
triterpene from Centella asiatica, is neuroprotective in a mouse model of focal 451
cerebral ischemia. J Neurosci Res 2009;87:2541-50. 452
6. Janero DR. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of 453
lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990;9: 454
515–540. 455
7. Zimmermann M. Ethical guidelines for investigations of experimental pain in 456
conscious animals. Pain 1983;16:109-110. 457
8. Chang HY, Sheu MJ, Yang CH, Leu ZC, Chang Y, Peng WH, et al. Analgesic effects 458
and the mechanisms of anti-inflammation of hispolon in mice. Evidence-Based Compl. 459
Altern. Med. 2009; doi:10.1093/ecam/nep027. 460
9. Flohe L, Otting F. Superoxide dismutase assays. Methods in Enzymology 1984;105: 461
93–104. 462
10. Aebi H. Catalase in vitro. Methods in Enzymology. 1984;105:121–126. 463
11. Paglia ED, Valentine WN. Studies on the quantitative and qualitative characterization 464
of erythrocytes glutathione peroxidase. J Lab Clin Med 1967; 70:158–169.
465
12. Shibata M, Ohkubo T, Takahashi H, Inoki R. Modified formalin test: characteristic 466
biphasic pain response. Pain 1989;38:347–352. 467
13. Franzotti EM, Santos CV, Rodrigues HM, Mourao RH, Andrade MR, Antoniolli AR. 468
Anti-inflammatory, analgesic activity and acute toxicity of Sida cordifolia L. 469
(Malva-branca). J Ethnopharmacol. 2000;72:273–727. 470
14. Viana AF, Maciel IS, Motta EM, Leal PC, Pianowski L, Campos MM, Calixto JB. 471
Antinociceptive Activity of Trichilia catigua Hydroalcoholic Extract: New Evidence 472
on its Dopaminergic Effects. Evidence-Based Compl. Altern. Med. 473
2009;doi:10.1093/ecam/nep144. 474
15. Tjolsen A, Berge OG, Hunskaar S, Rosland JH, Hole K. The formalin test: an 475
evaluation of the method. Pain 1992; 51:5-17. 476
16. Spector WG, Willoughb DA. The inflammatory response. Bacteriol Rev. 1963;27: 477
117–154. 478
17. Tohda C, Nakayama N, Hatanaka F, Komatsu K. Comparison of anti-inflammatory 479
activities of six Curcuma rhizomes: a possible curcuminoid-independent pathway 480
mediated by Curcuma phaeocaulis extract. Evidence-Based Compl. Altern. Med. 2006; 481
3:255–260. 482
18. Salvemini D, Wang Z, Bourdon DM, Stern MK, Curne MG, Manning PT. Evidence 483
of peroxynitrite involvement in the carrageenan induced rat paw edema. Eur. J. Clin. 484
Pharmacol. 1996; 303:217–220.
485
19. Cuzzocrea S, Zingarelli B, Calapai G, Nava F, Caputi AP. Zymosanactivated plasma 486
induces paw oedema by nitric oxide and prostaglandin production. Life Sci. 1997;60: 487
215–220. 488
20. Liao H, Banbury LK, Leach DN. Elucidation of Danzhixiaoyao Wan and Its 489
Constituent Herbs on Antioxidant Activity and Inhibition of Nitric Oxide Production. 490
Evidence-Based Compl. Altern. Med. 2007; 4:425–430. 491
21.Saad B, Abouatta BS, Basha W, Hmade A, Kmail A, Khasib S, et al. Hypericum 492
triquetrifolium—Derived Factors Downregulate the Production Levels of LPS-Induced
493
Nitric Oxide and Tumor Necrosis Factor- in THP-1 Cells. Evidence-Based Compl. 494
Altern. Med. 2007; 4:425–430. 495
22. Dawson J, Sedgwick AD, Edwards JC, Lees PA. comparative study of the cellular, 496
exudative and histological responses to carrageenan, dextran and zymosan in the 497
mouse. Int. J. Tissue React. 1991;13:171–185. 498
23. Lee YS, Jin DQ, Beak SM, Lee ES, Kim JA. Inhibition of ultraviolet-A-modulated 499
signaling pathways by asiatic acid and ursolic acid in HaCaT human keratinocytes. 500
Eur J Pharmacol. 2003; 476:173–178.
501
24.Yun KJ, Kim JY, Kim JB, Lee KW, Jeong SY, Park HJ, Jung HJ, et al. Inhibition of 502
LPS-induced NO and PGE2 production by asiatic acid via NF-κB inactivation in 503
RAW 264.7 macrophages: Possible involvement of the IKK and MAPK pathways. 504
Int Immunopharmacol 2008; 8:431–441.
505
25. Lee SH, Soyoola E, Chanmugam P, Hart S, Sun W, Zhong H, et al. Selective 506
expression of mitogen-inducible cyclooxygenase in macrophages stimulated with 507
lipopolysaccharide. J Biol Chem 1992; 267:25934–25938. 508
26. Salvemini D, Ischiropoulos H, Cuzzocrea S. Roles of nitric oxide and superoxide in 509
inflammation. Methods Mol Biol. 2003; 225: 291–303. 510
27. Subbaramaiah K, Dannenberg AJ. Cyclooxygenase 2: a molecular target for cancer 511
prevention and treatment. Trends Pharmacol Sci. 2003;24:96–102. 512
28. Lefkowitz DL, Gelderman MP, Fuhrmann SR, Graham S, Starnes JD, Lefkowitz SS, 513
et al. Neutrophilic lysozyme-macrophage interactions perpetuate chronic 514
inflammation associated with experimental arthritis. Clin Immunol 1999; 515
91:145–155. 516
29. Harris SG, Padilla J, Koumas L, Ray D, Phipps RP. Prostaglandins as modulators of 517
immunity. Trends Immunol 2002;23:144–150. 518
30. Min SW, Ryu SN, Kim DH. Anti-inflammatory effects of black rice,
519
cyanidin-3-O-β-D-glycoside, and its metabolites, cyanidin and protocatechuic acid.
520
Int Immunopharmacol 2010;10:959-966. 521
31. Karin M, Ben-Neriah Y. Phosphorylation meets ubiquitination: the control of NF-κB
522
activity.Annu Rev Immunol 2000;18:621–663. 523
32. Moncada S, Palmer RMJ, Higgs EA. Nitric oxide physiology, pathophysiology and 524
pharmacology. Pharmacol Rev 1991;43:109–142. 525
33. Hogg N. Free radicals in disease. Semin Reprod Endocrinol 1998;16:241–248. 526
34. Yoshikawa T, Naito Y. The role of neutrophils and inflammation in gastric mucosal 527
injury. Free Radical Research 2000;33:785–794. 528 529 530 531 532 533 534 535 536 537 538 539 540 541 542 543 544 545 546 547
Figure Legends
548Figure 1. Chemical structure of
asiatic acid
(AA).549 550
Figure 2. Analagesic effects of AA and indomethacin (Indo) on acetic acid-induced
551
writhing response in mice. Each value represents as mean ± S.E.M. *p < 0.05,
552
**
p < 0.01, and ***p < 0.001 as compared with the pathological model group
553
(Con) (one-way ANOVA followed by Scheffe’s multiple range test). 554
555
Figure 3. Effects of AA and Indo on the early phase and late phase in formalin test in
556
mice. Each value represents as mean ± S.E.M. *p < 0.05, **p < 0.01 and ***p <
557
0.001 as compared with the pathological model group (Con) (one-way ANOVA 558
followed by Scheffe’s multiple range test). 559
560
Figure 4. Effects of AA and Indo on hind paw edema induced by Carr in mice. Each
561
value represents as mean ± S.E.M. ***p < 0.001 as compared with the Carr
562
group (one-way ANOVA followed by Scheffe’s multiple range test). 563
564
Figure 5. Effects of AA and Indo on the tissue MDA concentration of paw in mice.
565
Normal control received 0.9% normal saline. Animals treated with AA (1, 5,
566
and 10 mg/kg) and Indo were assayed for their ability inhibiting MDA
567
production in the right hind paws. The right hind paw tissues were dissected
568
at the 5 h. Then the homogenate was centrifuged and the supernatant was
569
obtained for the MDA assays. Each value represents as mean ± S.E.M. ###p <
0.001 as compared with the control group. **p < 0.01 and ***p < 0.001 as
571
compared with the Carr group (one-way ANOVA followed by Scheffe’s 572
multiple range test). 573
574
Figure 6. Effects of AA and Indo on Carr-induced (A) NO, (B) TNF-α, and (C)
575
interlukin-1β concentrations of serum at 5 h in mice. Normal control received
576
0.9% normal saline. Animals treated with AA (1, 5, and 10 mg/kg) and Indo
577
were assayed in the right hind paws. After 5 h, the animals were sacrificed and
578
blood was withdrawn. Then fresh blood was centrifuged and the supernatant
579
was obtained for measuring NO, TNF-α, and interlukin-1β levels. Each value 580
represents as mean ± S.E.M. ###p < 0.001 as compared with the control group.
581
*
p < 0.05, **p < 0.01 and ***p < 0.001 as compared with the Carr group
582
(one-way ANOVA followed by Scheffe’s multiple range test). 583
584
Figure 7. Inhibition of iNOS, COX-2, and NF-κB protein expression by AA induced by
585
Carr in mice paw edema for 5 h. Normal control received 0.9% normal saline.
586
Animals treated with AA (1, 5, and 10 mg/kg) and Indo to injection of Carr
587
right hind paws. The right hind paw tissues were taken at the 5 h. Then the
588
homogenate was centrifuged and tissue suspended were then prepared and 589
subjected to western blotting using an antibody specific for iNOS, COX-2 and 590
NF-κB. β-actin was used as an internal control. (A) Representative western 591
blot from two separate experiments is shown. (B) Relative iNOS, COX-2 and 592
NF-κB protein levels were calculated with reference to Carr-injected mouse. 593
###
compared with sample of control group. The data were presented as mean ± 594
S.D. for three different experiments performed in triplicate. **p < 0.01 and ***p
595
< 0.001 were compared with Carr-alone group. 596
597
Figure 8. Histological appearance of the mouse hind footpad after a subcutaneous
598
injection with Carr stained with H&E stain at the 5 h to reveal hemorrhage, 599
edema and inflammatory cell infiltration in (A) control mice, (B) Carr-treated 600
mice demonstrating hemorrhage with moderately extravascular red blood cells 601
and a large amount of inflammatory leukocyte mainly neutrophils infiltration 602
in the subdermis interstitial tissue of mice, and (C) mice given Indo (10 mg/kg) 603
before Carr. AA significantly shows (D) morphological alterations (100×) and 604
(E) the numbers of neutrophils in each scope (400x) compared to 605
subcutaneous injection of Carr only. ###p < 0.001 as compared with the control
606
group. **P < 0.01, and ***p < 0.001 compared with Carr group. Scale bar =
607
100 µm. 608
609
Figure 9. Propose the mechanism of AA in λ-carrageenan (Carr) -injected mouse. AA
610
inhibit the production of TNF-α, free radicals and lipid peroxidation, which in 611
turn decrease MDA level, iNOS, COX-2, and NF-κB activation in the paw 612
edema and increase the CAT, SOD and GPx activities in the liver. MDA: 613
malondialdehyde; TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β; NO: 614
nitric oxide; CAT: catalase; SOD: superoxide dismutase; GPx: glutathione 615
peroxidase; iNOS: inducible nitric oxide synthase; COX-2: cycloxyclase-2; 616
NF-κB: Nuclear factor- κB. 617 618 619 620 621 622 623 624 625 626 627 628 629 630 631 632 633 634 635 636 637 638 639
640
Table 1: Effects of AA and Indo on the liver CAT, SOD, and GPx activities in mice. 641
642
Groups Catalase(U/mg protein) SOD (U/mg protein) GPx (U/mg protein)
Control 5.12 ± 0.21 24.39 ± 0.18 3.23 ± 0.18 Carr 3.46 ± 0.32### 17.56 ± 0.31### 1.96 ± 0.14### Carr+ Indo 4.53 ± 0.25** 22.13± 0.26** 2.76 ± 0.29** Carr + AA (1 mg/Kg) 3.84 ± 0.17 19.47 ± 0.15 2.14 ± 0.19 Carr + AA (5 mg/Kg) 4.36 ± 0.25* 21.32 ± 0.19* 2.49 ± 0.27* Carr + AA (10 mg/Kg) 4.67 ± 0.36** 23.06 ± 0.33** 2.93 ± 0.14**
Each value represents as mean ± S.E.M. ###p < 0.001 as compared with the control. *p <
643
0.05 and **p < 0.01 as compared with the Carr group (one-way ANOVA followed by
644
Scheffe’s multiple range test). 645 646 647 648 649 650 651 652 653 654 655
Figure 1.
656 657 658 659 660 661 662 663 664 665 666 667668
Figure 2.
669 W ri th in g r es p o n se 0 10 20 30 40 50 *** * ** *** AA Control 10 1 5 10 (mg/kg) Indo 1% Acetic acid 670 671 672 673 674 675 676 677 678679 680
Figure 3.
681 L ic k in g t im e (s e c ) 0 20 40 60 80 100 120 140 Early phase Late phase AA Control 10 1 5 10 (mg/kg) Indo 5% Formalin ** *** *** * 682 683 684 685 686 687 688689
Figure 4.
690Time (Hr)
0 1 2 3 4 5 C h a n g e s o f e d em a v o lu m e ( m L ) 0.00 0.02 0.04 0.06 0.08 Carr Carr and Indo Carr and AA 1.0 mg/kg Carr and AA 5.0 mg/kg Carr and AA 10 mg/kg *** *** *** ** *** ** ** ** * 691 692 693 694 695 696 697 698 699700
Figure 5.
701 T is s u e M D A c o n c e n tr a ti o n ( n m o l/ m g p ro te in ) 0.0 0.3 0.6 0.9 1.2 Control - Indo 1 5 10 (mg/kg) Carr**
***
###*
AA**
702 703 704 705 706 707 708
Figure 6.
709A.
710 *** Control - Indo 1 5 10 (mg/kg) ** ** *** *** ** ** Carr ### N it ri te (µµµµ M ) 0 2 4 6 8 10 12 14 16 ### *** *** AA * ** 711B.
712 T N F -αααα ( p g /m L ) 0 100 200 300 400 500 600 Carr ### Control - Indo 1 5 10 (mg/kg) *** ** * AA *** 713C.
714 In te rl e u k in -1 ββββ ( p g /m L ) 0 20 40 60 80 100 120 140 160 Carr ### Control - Indo 1 5 10 (mg/kg) *** ** * AA *** 715717
Figure 7.
718A.
719 720B.
721 *** ** ** *** Control - 10 10 (mg/kg) Indo AA Carr ** ** *** iN O S , C O X -2 , a n d N F -κκκκ B ( % o f c o n tr o l) 0.0 0.2 0.4 0.6 0.8 1.0 1.2 iNOS COX-2 NF-κB (p50) ######### *** *** *** **** ** 722723
Figure 8.
724 725E.
726 *** Control - Indo 10 (mg/kg) AA ** ** N e u tr o p h il / s c o p e ( c e ll ) 0 20 40 60 80 100 *** *** Carr ### 727728