REGULAR ARTICLE
Salicylic acid-mediated hydrogen peroxide accumulation
and protection against Cd toxicity in rice leaves
Yun-Yang Chao&Chao-Yeh Chen&
Wen-Dar Huang&Ching Huei Kao
Received: 23 June 2009 / Accepted: 26 August 2009 # Springer Science + Business Media B.V. 2009
Abstract The role of H2O2 in salicylic acid (SA)-induced protection of rice leaves against subsequent Cd toxicity was investigated. SA pretreatment resulted in an increase in the contents of endogenous SA, as judged by the expression of OsWRKY45 (a SA responsive gene), and H2O2in rice leaves. Dipheny-leneiodonium (DPI) and imidazole (IMD), inhibitors of NADPH oxidase, prevented SA-increased H2O2 production, suggesting that NADPH oxidase is a H2O2-generating enzyme in SA-pretreated rice leaves. DPI and IMD also inhibited SA-increased activities of superoxide dismutase (SOD), ascorbate peroixdase (APX), and glutathione reductase (GR) activities, but had no effect on SA-increased catalase (CAT) activity. Moreover, SA-induced protection against subsequent Cd toxicity could also be pre-vented by DPI and IMD. The inhibitory effect of DPI and IMD on SA-induced protection against subsequent Cd toxicity could be reversed by exoge-nous H2O2. All these results suggested that SA-induced protection against subsequent Cd toxicity is mediated through H2O2. This conclusion is sup-ported further by the observations that exogenous H2O2 application resulted in an increase in SOD,
APX, and GR activities, but not CAT activity and a protection against subsequent Cd toxicity of rice leaves.
Keywords Cadmium . Hydrogen peroxide . Oryza sativa L. . Oxidative stress . Salicylic acid Abbreviations
APX Ascorbate peroxidase AsA Ascorbate
CAT Catalase
DAB 3,3-Diaminobenzidine DPI Diphenyleneiodonium DW Dry weight
FW Initial fresh weight GR Glutathione reductase IMD Imidazole
IRT Iron-regulated transporter MDA Malondialdehyde ROS Reactive oxygen species SA Salicylic acid
SOD Superoxide dismutase
Introduction
Cadmium (Cd) is a widespread nonessential heavy metal, classified as a human carcinogen, and the uptake and accumulation of Cd in plants represent the main entry pathway into human and mammal food.
DOI 10.1007/s11104-009-0161-4
Responsible Editor: Fangjie J. Zhao.
Y.-Y. Chao
:
C.-Y. Chen:
W.-D. Huang:
C. Kao (*) Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of Chinae-mail: [email protected]
Cd contamination is a vital concern because of its known toxicity to human health (Nawrot et al.2006). In plants, Cd causes severe physiological and mor-phological effects such as chlorosis and growth reduction. Cd is a non-redox metal unable to participate in Fenton-type reactions, but it causes oxidative stress by generating reactive oxygen species (ROS) (Sanitá di Toppi and Gabbrielli1999; Garnier et al.2006). ROS react with lipids, proteins, pigments and nucleic acids and cause lipid peroxidation, membrane damage and inactivation of enzymes, thus resulting toxic effects. To minimize and /or to protect against the toxic effects of these damaging ROS, plants have evolved highly regulated enzymatic and non-enzymatic mechanisms to keep a balance be-tween ROS production and destruction in order to maintain cellular redox homeostasis. Plants use enzymes like superoxide dismutase (SOD), ascorbate peroxidase (APX), glutathione reducatse (GR), and catalase (CAT) as well as low molecular weight antioxidants like ascorbate (AsA) and glutathione to scavenge ROS (Noctor and Foyer1998).
Salicylic acid (SA) is an endogenous growth regulator of phenolic nature, which participates in the regulation of physiological processes in plants, such as growth, photosynthesis, nitrate metabolism, ethylene production, heat production and flowering (Hayat et al.2007). SA is also known to be involved in abiotic stress signaling, including plant response to heavy metals. SA pretreatment alleviates Cd toxicity in barley (Metwally et al.2003), soybean (Drazic and Mihailovic 2005), pea (Popova et al. 2009), maize (Krantev et al. 2008), and rice (Guo et al.2007a,b,
2009). However, SA application has been shown to aggravate the symptoms of cadmium stress (Pál et al.
2002). Zawoznik et al. (2007) also demonstrated that endogenous SA may function in Arabidopsis thaliana as a signaling molecule necessary to potentiate Cd-induced oxidative damage. In a recent review, Horváth et al. (2007) claimed that the effect of exogenous SA depends on numerous factors such as the species and developmental stage of the plant, the mode of application, and the concentration of SA and its endogenous level in the given plant.
Because H2O2 is relatively stable and diffusible through membranes (in contrast with superoxide), it is now considered as an alarm signal that triggers acclamatory/defense mechanisms in plant cells (Chen et al.1993; Prasad et al.1994a,b; Cruz de Carvalho
2008). H2O2 treatment has been shown to enhance tolerance against oxidative stress generated by chill-ing, heat salinity, drought, and high light intensities (Prasad et al. 1994a, b; Dat et al. 1998; Lopez-Delgado et al.1998; Gong et al.2001; Gechev et al.
2002; Uchida et al. 2002; Yu et al. 2002, 2003; Azevedo Neto et al.2005; Wahid et al.2007). Recent work also demonstrated that H2O2 pretreatment protects against the subsequent Cd stress of rice seedlings (Hsu and Kao 2007). Several lines of evidence indicate that SA pretreatment results in an accumulation of H2O2(Chen et al. 1993; Kauss and Jeblick 1995; Rao et al. 1997; Dat et al. 1998; Ganesan and Thomas 2001; Agarwal et al. 2005; Harfouche et al. 2008). It has been shown that thermotolerance obtained by exogenous application of SA can be achieved by an early increase in H2O2in mustard seedlings (Dat et al. 1998) and potato (Lopez-Delgado et al.1998).
In the present study, we showed that a SA pretreatment could induce Cd stress tolerance in rice leaves and investigated the possible involvement of H2O2in Cd stress tolerance.
Materials and methods Plant material
Rice (Oryza sativa L., cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed extensively seeds with distilled water. These seeds were then germinated in Petri dishes with wetted filter paper at 37°C under dark conditions. After 48 h incubation, uniformly germi-nated seeds were selected and cultivated in a 500 ml beaker containing half-strength Kimura B solution as described previously (Hsu and Kao 2003). The hydroponically cultivated seedlings were grown for 12 days in a Phytotron (Agricultural Experimental Station, National Taiwan University, Taipei, Taiwan) with natural sunlight at 30°C day/25°C night and 90% relative humidity.
SA pretreatment and Cd stress treatment
The apical 3-cm of the third leaves of 12-day-old seedlings was used for all experiments. Ten leaf segments were floated in a Petri dish containing 10 ml
SA containing 0.01% Triton X-100. Control leaf segments were floated in 0.01% Triton X-100 alone. Control and SA-pretreated rice leaves were then treated with or without CdCl2(5 mM) for 24 h. All pretreatments and treatments were carried out at 27°C and irradiance of 40μmol m−2s−1.
Measurement of H2O2, malondialdehyde (MDA), and Cd
H2O2 was visually detected in the leaves by using 3,3-diaminobenzidine (DAB) as substrate (Orózco-Cárdenas and Ryan1999). Rice leaves were supplied through the cut ends with DAB (1 mg ml−1) solution for 24 h under light at 27°C. Rice leaves were then decolorized in boiling ethanol (95%) for 0.5 h. This treatment decolorized the leaves except for the brown polymerization product produced by DAB with H2O2. After cooling, the leaves were extracted at room temperature with fresh ethanol to visualize the brown spots. The H2O2 staining was repeated four times with similar results. In some experiments, the H2O2 content was measured spectrophotometri-cally after reaction with TiCl4(Tsai et al.2004). The reaction mixture consisted of 2 ml of 50 mM phosphate buffer (pH 6.8) leaf extract supernatant and 1 ml reagent [0.1% (v/v) TiCl4 in 20% (v/v) H2SO4]. The blank probe consisted of 50 mM phosphate buffer in the absence of leaf extract. The absorbance was measured at 410 nm. The amount of H2O2 was calculated by use of a standard curve prepared with known concentration of H2O2. The H2O2 content was expressed on the basis of initial fresh weight (FW).
Malondialdehyde (MDA), routinely used as an indicator of lipid peroxidation, was extracted with 5% (w/v) trichloroacetic acid and determined by the thiobarbituric acid reaction as described by Heath and Packer (1968). MDA content was expressed on the basis of FW.
For determination of Cd, leaves were dried at 65°C for 48 h. Dried material was ashed at 550°C for 20 h. The ash residue was incubated with 31% HNO3 and 17.5% H2O2 at 72°C for 2 h, and dissolved in distilled water. Cd concentration was then quantified using an atomic absorption spectro-photometer (Model AA-6800, Shimadzu, Kyoto, Japan). Amount of Cd is expressed on the basis of dry weight (DW).
Enzyme extraction and assays
For extraction of enzymes, leaf tissues were homog-enized with 0.1 M sodium phosphate buffer (pH 6.8) in a chilled pestle and mortar. For analysis of ascorbate peroxidase (APX) activity, 2 mM ascorbate (AsA) was added to the extraction buffer. The homogenate was centrifuged at 12,000 g for 20 min and the resulting supernatant was used for determi-nation of enzyme activity and protein content. The whole extraction procedure was carried out at 4°C. Superoxide dismutase (SOD) was determined accord-ing to Paoletti et al. (1986). The reaction mixture (2.73 ml) contained 100 mM triethanoamine-diethanolamine buffer (pH 7.4), 7.5 mM NADH, EDTA/MnCl2(100 mM/50 mM, pH 7.0), 10 mM 2-mercaptoethanol, and enzyme extract (0.2 ml). The reaction was started by the addition of NADH. The reaction was allowed to proceed for 10 min. The absorbance was measured at 340 nm. One unit of SOD was defined as the amount of enzyme that inhibits by 50% the rate of NADH oxidation observed in blank sample. Catalase (CAT) activity was assayed according to Kato and Shimizu (1987). The decrease in H2O2 was followed as the decline in the absor-bance at 240 nm, and the activity was calculated using the extinction coefficient (40 mM−1 cm−1 at 240 nm) for H2O2. One unit of CAT was defined as the amount of enzyme which degraded 1 μmol H2O2 per min. APX activity was determined according to Nakano and Asada (1981). The decrease in AsA concentration was followed as a decline in the absorbance at 290 nm and activity was calculated using the extinction coefficient (2.8 mM−1 cm−1 at 290 nm) for AsA. One unit of activity for APX was defined as the amount of enzyme that degraded 1 μmol of AsA per min. Glutathione reductase (GR) was determined by the method of Foster and Hess (1980). One unit of GR was defined as the amount of enzyme that decreased 1 absorbance min−1at 340 nm. The enzyme extracts were used for the determination of protein by the method of Bradford (1976).
Quantitative RT-PCR analysis of OsWRKY 45 gene Total RNA was isolated from rice leaves treated with or without SA using the TRIZOL reagent (Invitrogen, CA, USA), accroding to supplier’s recommendations. To remove contaiminating genomic DNA, RNA was
treated with Turbo DNase I (Ambio, TX, USA) for 30 min at 37°C before the cDNA systhesis system for RT-PCR (Invitrogen, CA, USA). cDNA was amplified by PCR in 7500 Real-Time PCR System (Applied Biosystems ) with SYBR Advantage qPCR Premix (Clontech, USA) with 1 cycle of 95°C for 10 min and 40 cycles of 95°C for 15 s, 60°C for 1 min.
The gene-specific primers were designed from the 3′-UTR of the rice WRKY gene. OsWRKY45 (AY870611) forward primer (ACG ATCGAA AGAAGATGGAT) and reverse primer (TCGTGTTGTTACTTGCT AGCATG). The mean value of the three replicates was normalized using OsUbiquitin as the internal control. OsUbiqutin (D12629.1) forward primer (CGCAAGTACAACCAGGACAA) and reverse primer (TGGTTGCTGTGACCACACTT). The primer sets were tested by dissocitation curve analysis and verified for the absence of nonspecific amplification. The relative expression level was then normalized to the treatment without SA.
Statistical analysis
Statistical differences between measurements (n=4) for different treatments or different times were analyzed following Student’s t-test or Duncan’s multiple range test. A P<0.05 was considered statistically significant.
Results
H2O2content and antioxidant enzyme activities in response to SA
In the present study, DAB staining was used to examine whether SA induces H2O2production in rice leaves. To
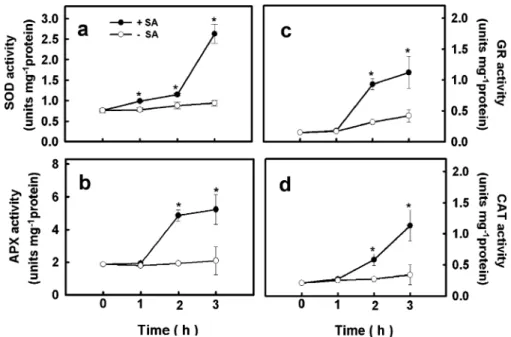
verify the specificity of brown polymerization product produced by DAB with H2O2, some leaves were immersed for 1 h in solution containing 2 mM AsA, a scavenger of H2O2. It was observed that the development of DAB-H2O2 reaction product by SA in rice leaves could be reduced by AsA (Fig. 1), indicating that the DAB staining method for H2O2 is specific. Little DAB staining was observed in control leaves (Figs. 1 and 2a). In our preliminary work, we observed that increasing concentration of SA from 1 to 3 mM for 3 h progressively increased DAB staining in rice leaves and no further increase was observed at 5 mM SA. Thus, in the present study, 3 mM SA was used for all experiments. The increase in DAB-H2O2 reaction product was evident 1 h after SA treatment (Fig.2a). SA-responsive rice gene WRKY45 has been reported (Ryu et al. 2006; Shimono et al. 2007). To examine the effect of SA on the expression of OsWRKY45, rice leaves were treated with 3 mM SA for 1, 2, and 3 h. The expression of OsWRKY45 was notably increased by SA 1 h after treatment (Fig.2b). SOD, APX, GR, and CAT activities in SA-treated rice leaves were higher than their respective control leaves (Fig.3a, b, c, d). The increase in SOD activity was evident 1 h after SA treatment (Fig.3a). However,
Treatment ( 1 h )
SA ( 3 mM ) AsA ( 2 mM )
– – + + – + – +
Fig. 1 Histochemical detection of H2O2with DAB staining in
rice leaves treated with SA, AsA, and SA + AsA. The concentrations of SA and AsA were 3 mM and 2 mM, respectively
Fig. 2 Histochemical detection of H2O2with DAB staining (a)
and change in mRNA level of OsWRKY45 (b) in rice leaves treated with or without SA. Rice leaves were incubated in 3 mM SA in 0.01% Triton X-100 for 1, 2, and 3 h in the light. Control leaves (−SA) were incubated in Triton X-100 alone. Bars indicate standard error (n=4). Asterisk represents values that are significantly different between −SA and +SA treat-ments at P<0.05
the increase in APX, GR, and CAT activities occurred 2 h after SA treatment (Fig.3b, c, d).
Effect of SA pretreatment on subsequent Cd toxicity of rice leaves
In the present study, Cd toxicity in rice leaves caused by Cd was assessed by an increase in MDA content. Increasing concentration of Cd from 0.1 to 5 mM progressively increased MDA content in rice leaves in the light and no further increase was observed at 10 mM CdCl2(data not shown). Thus, 5 mM CdCl2 was used in the present study. To test if SA pretreatment would affect the subsequent Cd-induced toxicity, rice leaves were pretreated with 3 mM SA for 3 h in the light and then treated with 5 mM CdCl2for 24 h in the light. It was observed that a 3-h SA pretreatment exhibited a significant reduc-tion of Cd-increased MDA content (Fig.4a).
The effect of NADPH oxidase inhibitors
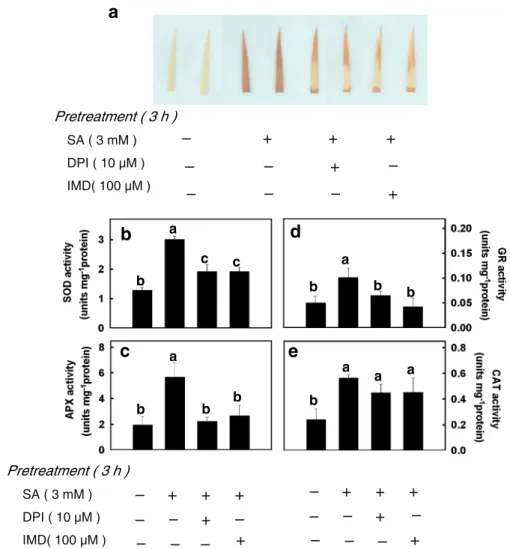
The role of NADPH oxidase in the SA-induced H2O2production was investigated by using NADPH oxidase inhibitors such as diphenyleneiodonium (DPI) and imidazole (IMD). When rice leaves were
treated with a solution of DPI (10 μM) or IMD (100 μM), SA-induced accumulation of H2O2 was reduced by DPI or IMD (Fig.5a). DPI or IMD also inhibited SA-increased activities of SOD, APX, GR (Fig.5b, c, d). However, DPI or IMD had no effect
Pretreatment ( 3 h ) SA ( 3 mM ) H2O2 ( 10 mM ) DPI ( 10 µM ) IMD ( 100 µM ) Treatment ( 24 h ) CdCl2 ( 5 mM ) – – + + + + + + – – – – – – + + – – – – + – + – – – – – – + – + – + – + + + + + – – + + – – – – – – – – – + – + – – – – a b d a cd c b b c c c c a b MDA (nmol g-1FW) 0 20 40 60 80
Fig. 4 Effect of CdCl2 on MDA content in rice leaves
pretreated with SA, SA + DPI, SA + IMD, SA + DPI + H2O2, or SA + IMD + H2O2(a) or pretreated with or without
H2O2(b) for 3 h in the light. The concentrations of CdCl2, SA,
DPI, IMD, and H2O2were 5 mM, 3 mM, 10μM, 100 μM, and
10 mM, respectively. MDA was determined 24 h after CdCl2
treatment. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
Fig. 3 Changes in the activities of SOD (a), APX (b), GR (c), and CAT (d) in rice leaves in response to SA. Rice leaves were incubated in 3 mM SA in 0.01% Triton X-100 for 1, 2, and 3 h in the light. Control leaves (−SA) were incubated in Triton
X-100 alone. Bars indicate standard error (n = 4). Asterisk represents values that are significantly different between−SA and +SA treatments at P<0.05
on SA-induced CAT activity (Fig. 5e). DPI or IMD alone had no effect on MDA content in rice leaves (Table 1). Content of MDA in Cd-exposed DPI-pretreated rice leaves was similar to that in the Cd-treated control, whereas that in Cd-exposed IMD-pretreated rice lee leaves was higher than in Cd-treated controls (Table 1). SA-induced protection against subsequent increase in MDA content by Cd was reduced by DPI or IMD (Fig. 4a). The inhibitory effect of DPI or IMD on SA-induced protection against subsequent increase in MDA content by Cd could be reversed by exogenous H2O2(10 mM) (Fig.4a).
Effect of H2O2pretreatment
To test if exogenous H2O2acts similarly as SA on the activities of antioxidant enzymes, rice leaves were pretreated with 10 mM H2O2for 3 h. Figure6ashows
that H2O2 pretreatment resulted in an increase in endogenous H2O2content in rice leaves. Furthermore, rice leaves pretreated with H2O2showed an enhance-ment in SOD, APX, and GR activities (Fig.6b, c, d) but had no effect on CAT activity in rice leaves (Fig.6e). When H2O2-pretreated rice leaves were then transferred to solution with or without CdCl2for 24 h. It was observed that pretreatment of rice leaves with H2O2 greatly improved tolerance of rice leaves exposed to Cd stress (Fig.4b).
Effect of SA or H2O2pretreatment on Cd concentration in rice leaves
To test if SA- or H2O2 pretreatment affects subsequent Cd uptake, rice leaves were first treated with SA or H2O2 for 3 h and then transferred to solution with or without 5 mM CdCl2. It was
a b b b a a a b Pretreatment ( 3 h ) SA ( 3 mM ) DPI ( 10 µM ) IMD( 100 µM )
d
e
–
–
+
–
– + + +
–
–
–
+
a
Pretreatment ( 3 h ) SA ( 3 mM ) DPI ( 10 µM ) IMD( 100 µM )– + + +
– +
–
–
– +
–
–
a b b bc
a b c cb
–
–
+
–
– + + +
–
–
–
+
Fig. 5 Effect of DPI and IMD on SA-induced H2O2
-DAB reaction product (a) activities of SOD (b), APX (c), GR (d), and CAT (e) in rice leaves. Rice leaves were incubated in SA, SA + DPI, and SA + IMD in 0.01% Triton X-100 for 1, 2, and 3 h in the light. Control leaves (−SA) were incubated in Triton X-100 alone. Con-centrations of SA, DPI, and IMD were 3 mM, 10μM, and 100μM, respectively. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
observed that rice leaves pretreated with 3 mM SA had higher Cd concentration than those pretreated without SA (Fig. 7a). Interestingly, pretreatment of rice leaves with H2O2 also increased Cd accumula-tion (Fig.7b).
Discussion
The expression of OsWRKY45 is known to increase in rice leaves treated with SA (Ryu et al.2006; Shimono et al. 2007). Thus, the level of SA reported in this study was judged by the transcript level of OsWRKY45. The expression of OsWRKY45 was enhanced in rice leaves by 3 mM SA (Fig. 2b), indicating that exogenous SA treatment indeed resulted in an increase in SA content in rice leaves.
In previous work, it has been demonstrated that Cd can induce oxidative stress in rice leaves, character-ized by an increase in the content of MDA (an indicator of lipid peroxidation) (Kuo and Kao 2004; Hsu and Kao 2005). Thus, in the present study, Cd toxicity was evaluated by the increase in MDA content of rice leaves. On the basis of this criterion, we demonstrated that rice leaves pretreated with SA protected against subsequent Cd stress (Fig. 4a). The protective effect of SA against subsequent Cd toxicity has also been described previously (Metwally et al. 2003;
Table 1 Effect of DPI and IMD on MDA content in rice leaves treated with or without CdCl2for 3 h in the light
Pretreatment (3 h) Treatment (24 h) MDA (nmol g−1FW) H2O H2O 28.4±4.9c DPI H2O 31.2±4.0c IMD H2O 24.8±4.3c H2O CdCl2 57.4±1.4b DPI CdCl2 54.6±7.5b IMD CdCl2 70.0±4.3a
The concentrations of CdCl2, DPI, and IMD were 5 mM, 10μM,
and 100μM respectively. MDA was determined 24 h after CdCl2
treatment. Bars indicate standard error (n=4). Values with the same letter are not significantly different at P<0.05
*
*
*
*
Pretreatment ( 3 h ) H2O2 ( 10 mM )– +
a
Pretreatment ( 3 h ) H2O2 ( 10 mM )– +
– +
b
c
d
e
Fig. 6 Effect of exogenousH2O2on the content of H2O2
(a) and the activities of SOD (b), APX (c), GR (d), and CAT (e) in rice leaves. The concentration of H2O2was
10 mM. Bars indicate stan-dard error (n=4). Asterisk represents values that are significantly different be-tween−H2O2and +H2O2
Drazic and Mihailovic2005; Guo et al.2007a,b,2009; Krantev et al.2008; Popova et al.2009).
Several lines of evidence indicate that SA pretreat-ment results in an accumulation in H2O2 content (Chen et al.1993; Kauss and Jeblick1995; Rao et al.
1997; Dat et al. 1998; Ganesan and Thomas 2001; Agarwal et al. 2005; Harfouche et al. 2008). Using DAB staining method, we have shown that SA induced the accumulation H2O2 in rice leaves (Fig. 2a). Figure 2b shows that OsWRKY45 expres-sion was increased in rice leaves treated with SA. The negligible H2O2detected in control leaves suggested that H2O2observed in SA-treated rice leaves is indeed caused by SA applied.
SA-induced accumulation of H2O2 has been suggested to be due to SA-inhibited CAT and APX activities (Chen et al. 1993; Sánchez-Casas and Klessig 1994; Klessig et al. 2000; Horváth et al.
2007). This does not seem to be the case in rice leaves, because SA increased APX and CAT activities (Fig. 3b, d). Agarwal et al. (2005) reported that SA-induced H2O2 production in wheat seedlings is mediated through a plasma membrane NADPH oxidase activity. The fact that SA-induced H2O2 production in rice leaves can be inhibited by DPI and IMD, inhibitors of NADPH oxidase (Fig. 5a), suggests that SA-dependent H2O2 generation in rice leaves originated, at least in part, from plasma membrane NADPH oxidase.
The present study indicated that H2O2 was in-volved in SA-induced protection against subsequent Cd stress of rice leaves. This conclusion was based on the observations that (1) endogenous H2O2 was increased in rice leaves pretreated with SA (Fig.2a); (2) SA-induced H2O2 production (Fig. 5a) and
protection against subsequent Cd stress (Fig. 4a) could be counteracted by DPI or IMD, inhibitor of NADPH oxidase; (3) the effect of DPI or IMD on SA-induced protection against subsequent Cd stress could be reversed by exogenous application of H2O2 (Fig. 4a), and (4) pretreatment of rice leaves with exogenous H2O2 greatly improved tolerance of rice leaves to Cd (Fig.4b).
It has been shown that various abiotic stresses induce oxidative stress and improvement of stress tolerance is often related to the increase in the activities of antioxidant enzymes (Noctor and Foyer
1998). Cho and Seo (2005) reported that seedlings of Cd-resistant Arabidopsis have higher activities of SOD, APX, and GR and experienced lower oxidative stress from Cd exposure. In the present study, we show that SA pretreatment resulted in higher activities of SOD, APX, and GR activities (Fig. 3a, b, c) and protection against of subsequent Cd stress in rice leaves (Fig.4a). H2O2is now considered as a signal molecule that induces gene expression and activities of antioxidant enzyme (Foyer et al.1997; Morita et al.
1999; Prasad et al. 1994a,b; Hong et al. 2009). The fact that DPI or IMD treatment was effective in reducing SA-increased SOD, APX, and GR activities (Fig. 5b, c, d) but not CAT activity (Fig. 5e) in rice leaves suggests that H2O2can act as a signal molecule in SA- increased SOD, APX, and GR activities. This suggestion was supported further by the observations that exogenous H2O2treatment increased SOD, APX and GR activities (Fig. 6b, c, d) but not CAT activity (Fig. 6e).
Earlier work by Metwally et al. (2003) demon-strated that pretreatment of barley seeds with SA protects Cd toxicity to roots. However, this protection
– – + + – + +– – + + – – – + + a b c c a b c c a b Pretreatment ( 3 h ) SA ( 3 mM ) H2O2 ( 10 mM ) Treatment ( 24 h ) CdCl2 ( 5 mM ) – – – – – – – – Fig. 7 Effect of CdCl2 on Cd concentration in rice leaves pretreated with or without SA (a) or pretreated with or without H2O2(b) for
3 h in the light. The con-centration of CdCl2, SA and
H2O2were 5 mM, 3 mM,
and 10 mM, respectively. Cd was determined 24 h after CdCl2treatment. Bars
indi-cate standard error (n=4). Values with the same letter are not significantly different at P<0.05
was not due to upregulation of antioxidant activity. In soybean seedlings, the influence of SA on the alleviation of toxic effect of Cd is possibly mediated through the regulation of K and Mg distribution (Drazic and Mihailovic 2005). Guo et al. (2007a,
2009) concluded that the SA-elevated enzymatic and non-enzymatic antioxidants contribute to alleviation of Cd toxicity in rice roots. Using detached rice leaves, we showed that early accumulation of H2O2 during SA pretreatment signals the increase in SOD, APX, and GR activities, which in turn prevents rice leaves from oxidative damage by Cd. It appears that the mechanism of SA on the alleviation of Cd toxicity depends on the methods of SA application and plant species examined.
SA-pretreatment increased Cd accumulation in rice leaves (Fig. 7a). Thus, the protective effect of SA against subsequent Cd toxicity of rice leaves is unlikely due to inhibition of Cd uptake. The increase in the Cd uptake by SA has also been described previously (Drazic et al. 2006). However, there are other reports showing that exogenous SA application reduced the uptake of Cd (Pál et al.2002; Krantev et al.2008: Popova et al.2009). Of particular interest is the finding that H2O2 pretreatment resulted in an increase in Cd concentration (Fig.7b). The possibility that H2O2acts as a signal molecule for SA-enhanced Cd uptake remained to be seen.
In the present study, the total leaf Cd concentration was measured. It is not known whether SA-pretreatment alters Cd distribution between the vacu-olar compartment and the rest of the cell. In future studies, it will be important to determine the effect of SA on Cd concentration in different cellular compart-ments. SA pretreatment increased endogenous SA content, as judged by the expression of OsWRKY45 (Fig.2b). Thus, the possibility that formation of stable SA-Cd complexes, which might reduce Cd toxicity after SA pretreatment, cannot be excluded.
It has been suggested that SA-enhanced Cd uptake is mediated through activation of some divalent cation transporters (Drazic et al. 2006). Iron-regulated transporter 1 (IRT1) is a major Fe transporter (Eide et al. 1996). A role for OsIRT1 in Cd transport has been previously investigated in yeast (Nakanishi et al.
2006). Recent experiment using transgenic rice plants over-expressing OsIRT1 confirmed the conclusion from those earlier yeast experiments that OsIRT1 indeed transports Cd (Lee and An 2009). It is not
known whether SA or H2O2pretreatment activates the expression of OsIRT1. Future research on the effect of SA or H2O2on the expression of OsIRT1 is likely to be highly rewarding.
Acknowledgements This work was supported by a research grant of the National Science Council of the Republic of China (NSC2628-B-002-002).
References
Agarwal S, Sairam RK, Srivastava GC, Tyagi A, Meena RC (2005) Role of ABA, salicylic acid, calcium and hydrogen peroxide on antioxidant enzymes induction in wheat seedlings. Plant Sci 169:559–570
Azevedo Neto AD, Prisco JT, Enéas-Filho J, Medeiros J-VR, Gomes-Filho E (2005) Hydrogen peroxide pre-treatment induces salt-stress acclimation in maize plants. J Plant Physiol 162:1114–1122
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254 Chen Z, Silva H, Klessig DF (1993) Active oxygen species in the induction of plant systemic acquired resistance by salicylic acid. Science 262:1883–1886
Cho U-H, Seo N-H (2005) Oxidative stress in Arabidopsis thaliana exposed to cadmium is due to hydrogen peroxide accumulation. Plant Sci 168:113–120
Cruz de Carvalho MH (2008) Drought stsress and reactive oxygen species. Plant Signal Behav 3:156–165
Dat JF, Foyer CH, Scott IM (1998) Changes in salicylic acid and antioxidants during induced thermotolerance in mustard seedlings. Plant Physiol 118:1455–1461
Drazic G, Mihailovic N (2005) Modification of cadmium toxicity in soybean seedlings by salicylic acid. Plant Sci 168:511–517 Drazic G, Mihailovic N, Lojic M (2006) Cadmium accumula-tion in Medicago sativa seedlings treated with salicylic acid. Biol Plant 50:239–244
Eide D, Broderius M, Fett J, Guerinot ML (1996) A novel iron-regulated metal transporter from plants identified by functional expression in yeast. Proc Natl Acad Sci USA 93:5624–5628
Foster JG, Hess JL (1980) Response of superoxide dismutase and glutathione reductase activities in cotton leaf tissue exposed to an atmosphere enriched in oxygen. Plant Physiol 66:482–487
Foyer CH, Lopez-Delgado H, Dat JF, Scott IM (1997) Hydrogen peroxide and glutathione-associated mechanism of acclima-tory stress tolerance and signaling. Physiol Plant 100:241– 254
Ganesan V, Thomas G (2001) Salicylic acid response in rice: influence of salicylic acid on H2O2 accumulation and
oxidative stress. Plant Sci 160:1095–1106
Garnier L, Simon-Plas F, Thuleau P, Angel J, Blein J, Ranjeva R, Montillet J-L (2006) Cadmium affects tobacco cells by a series of three waves of reactive oxygen species that contribute to cytotoxicity. Plant Cell Environ 29:1956–1969
Gechev T, Gadjev I, Van Breusegem F, Inzé D, Dukiandjiev S, Toneva V, Minko I (2002) Hydrogen peroxide protects tobacco from oxidative stress by inducing a set of antioxidant enzymes. Cell Mol Life Sci 59:708–714 Gong M, Chen B, Li X-G, Guo L-H (2001) Heat-shock-induced
cross adaptation to heat, chilling, drought and salt stress in maize seedlings and involvement of H2O2. J Plant Physiol
158:1125–1130
Guo B, Liang YC, Zhu YG, Zhao FJ (2007a) Role of salicylic acid in alleviating oxidative damage in rice roots (Oryza sativa) subjected to cadmium stress. Environ Pollut 147:743–749
Guo B, Liang YC, Li ZJ, Guo W (2007b) Role of salicylic acid in alleviating cadmium toxicity in rice roots. J Plant Nutr 30:427–439
Guo B, Liang YC, Zhu Y (2009) Does salicylic regulate antioxidant defense system, cell death, cadmium uptake and partitioning to acquire cadmium tolerance in rice? J Plant Physiol 166:20–31
Harfouche AL, Rugini E, Mencarelli F, Botondi R, Muleo R (2008) Salicylic acid induces H2O2 production and
endochitinase gene expression but not ethylene biosynthe-sis in Castanea sativa in vitro model system. J Plant Physiol 165:734–744
Hayat S, Ali B, Ahmad A (2007) Salicylic acid: biosynthesis, metabolism and physiological role in plants. In: Hayat S, Ahmad A (eds) Salicylic acid, a plant hormone. Springer, Drodrecht, pp 1–14
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198 Hong C-Y, Chao Y-Y, Yang M-Y, Cheng S-Y, Cho S-C, Kao
CH (2009) NaCl-induced expression of glutathione reduc-tase in roots of rice (Oryza sativa L.) seedlings is mediated through hydrogen peroxide but not abscisic acid. Plant Soil 320:103–115
Horváth E, Szalai G, Janda T (2007) Induction of abiotic stress tolerance by salicylic acid signaling. J Plant Growth Regul 26:290–300
Hsu YT, Kao CH (2003) Role of abscisic acid in cadmium tolerance of rice (Oryza sativa L.) seedlings. Plant Cell Environ 26:867–874
Hsu YT, Kao CH (2005) Abscisic acid accumulation and cadmium tolerance in rice seedlings. Physiol Plant 124:71–80 Hsu YT, Kao CH (2007) Heat shock-mediated H2O2
accumu-lation and protection against Cd toxicity in rice seedlings. Plant Soil 300:137–147
Kato M, Shimizu S (1987) Chlorophyll metabolism in higher plants VII. Chlorophyll degradation in senescing tobacco leaves: phenolic-dependent peroxidative degradation. Can J Bot 65:729–735
Kauss H, Jeblick W (1995) Pretreatment of parsely suspension cultures with salicylic acid enhances spontaneous and elicited production of H2O2. Plant Physiol 108:1171–1178
Klessig DF, Durnder J, Noad R, Navarre DA, Wendehenne D, Kumar D, Zhou JM, Shah J, Zhang S, Kachroo P, Trifa Y, Pontier D, Lam E, Silva H (2000) Nitric oxide and salicylic and signaling in plant defense. Proc Natl Acad Sci USA 97:8849–8855
Krantev A, Yordanova R, Janda T, Szalai G, Popova L (2008) Treatment with salicylic acid decreases the effect of
cadmium on photosynthesis in maize plants. J Plant Physiol 165:920–931
Kuo MC, Kao CH (2004) Antioxidant enzyme activities are upregulated in response to cadmium in sensitive, but not in tolerant, rice (Oryza sativa L.) seedlings. Bot Bull Acad Sin 45:291–299
Lee S, An G (2009) Over-expression of OsIRT1 leads to increased iron and zinc accumulations in rice. Plant Cell Environ 32:408–416
Lopez-Delgado H, Dat JF, Foyer CH, Scott IM (1998) Induction of thermotolerance in potato microplants by acetylsalicylic acid and H2O2. J Exp Bot 49:713–720
Metwally A, Finkemeier I, Georgi M, Dietz K-J (2003) Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol 132:272–281
Morita S, Kaminaka H, Masumura T, Tanaka K (1999) Induction of rice cytosolic ascorbate peroxidase mRNA by oxidative stress involvement of hydrogen peroxide in oxidative stress; the involvement of hydrogen peroxide by oxidative stress signaling. Plant Cell Physiol 40:417–422 Nakanishi H, Ogawa I, Ishimaru Y, Mori S, Nishizawa NK
(2006) Iron deficiency enhances cadmium uptake and translocation mediated by Fe2+ transporters OsIRR1 and
OsIRT2 in rice. Soil Sci Plant Nutr 52:464–469
Nakano Y, Asda K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867–880
Nawrot T, Plusquin M, Hogervorst J, Roels HA, Celis H, Thijs L, Vangronsveld J, Van Hecke E, Staessen JA (2006) Environmental exposure to cadmium and risk of cancer: a prospective population-based study. Lancet Oncol 7:119–126
Noctor G, Foyer CH (1998) Ascorbate and glutathione: keeping active oxygen under control. Annu Rev Plant Physiol Plant Mol Biol 49:249–279
Orózco-Cárdenas ML, Ryan CA (1999) Hydrogen peroxide is generated systemically in plant leaves by wounding and systemin via the octadecanoid pathway. Proc Natl Acad Sci USA 96:6553–6557
Pál M, Szalai G, Horváth E, Janda T, Páldi E (2002) Effect of salicylic acid during heavy metal stress. Acta Biologica Szegediensis 46:119–120
Paoletti F, Aldinucci D, Mocali A, Capparini A (1986) A sensitive spectrophotometric method for the determination of superoxide dismutase activity in tissue extracts. Anal Biochem 154:536–541
Popova LP, Maslenkova LT, Yordanova RY, Ivanova AP, Krantev AP, Szalai G, Janda T (2009) Exogenous treatment with salicylic acid attenuates cadmium toxicity in pea seedlings. Plant Physiol Biochem 47:224–231 Prasad TK, Anderson MD, Martin BA, Stewart CR (1994a)
Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide. Plant Cell 6:65–74
Prasad TK, Anderson MD, Stewart CR (1994b) Acclimation, hydrogen peroxide, and abscisic acid protect mitochondria against irreversible chilling injury in maize seedlings. Plant Physiol 105:619–627
Rao MV, Paliyath G, Ormrod DP, Murr DP, Watkins CB (1997) Influence of salicylic acid on H2O2production, oxidative
acid-mediated oxidative damage requires H2O2. Plant Physiol
115:137–149
Ryu H-S, Han M, Lee S-K, Cho JI, Ryoo N, Heu S, Lee Y-H, Bhoo SH, Wang G-L, Hahn T-R, Jeon J-S (2006) A comprehensive expression analysis of the WRKY gene superfamily in rice plants during defense response. Plant Cell Rep 25:836–847
Sánchez-Casas P, Klessig DF (1994) A salicylic acid-binding activity and a salicylic acid-inhibitable catalase activity are present in a variety of plant species. Plant Physiol 106:1675–1679
Sanitá di Toppi L, Gabbrielli R (1999) Response to cadmium in higher plants. Environ Exp Bot 41:105–130
Shimono M, Sugano S, Nakayama A, Jiang C-J, Ono K, Toki S, Takatsuji H (2007) Rice WRKY45 plays a crucial role in benzothiadiazole-inducible blast resistance. Plant Cell 19:2064–2074
Tsai Y-C, Hong C-Y, Liu L-F, Kao CH (2004) Relative importance of Na+ and Cl−in NaCl-induced antioxidant sytems in roots of rice seedlings. Physiol Plant 122:86–94
Uchida A, Jagendorf AT, Hibino T, Takabe T, Takabe T (2002) Effects of hydrogen peroxide and nitric oxide on both salt and heat stress tolerance in rice. Plant Sci 163:515–523
Wahid A, Perveen M, Gelani S, Basra SMA (2007) Pretreatment of seed with H2O2improves salt tolerance
of wheat seedlings by alleviation of oxidative damage and expression of stress proteins. J Plant Physiol 164:83– 294
Yu C-W, Murphy TM, Sung W-W, Lin C-H (2002) H2O2
treatment induced glutathione accumulation and chilling tolerance in mung bean. Funct Plant Biol 29:1081–1087 Yu C-W, Murphy TM, Lin C-H (2003) Hydrogen
peroxide-induced chilling tolerance in mung beans mediated through ABA-independent glutathione accumulation. Funct Plant Biol 30:955–963
Zawoznik M, Groppa MD, Tomaro ML, Benavides MP (2007) Endogenous salicylic acid potentiates cadmium-induced oxidative stress in Arabidopsis thaliana. Plant Sci 173:190–197