Citosol (thiamylal sodium) triggers apoptosis and affects gene
expressions of murine leukemia RAW 264.7 cells
RS-C Wu1, C-S Yu2, K-C Liu3, H-Y Huang4, S-W Ip4,J-P Lin5, F-S Chueh6, J-S Yang7 and J-G Chung8,9
Abstract
Citosol (thiamylal sodium) is one of generally used anesthetic–sedative agents for clinical patients, and it has not been reported to show induction of cytotoxic effects in cancer cells, especially in mice leukemia RAW 264.7 cells in vitro. In the present study, we investigated the cytotoxic effects of citosol on mice leukemic RAW
264.7 cells, including the effects on protein and gene expression levels which are determined by Western blotting and DNA microarray methods, respectively. Results indicated that citosol induced cell morphological changes, cytotoxic effect, and induction of apoptosis in RAW264.7 cells.Western blotting analysis demonstrated
that citosol promoted the levels of Fas, cytochrome c, caspase 9 and 3 active formand Bax levels, but it suppressed Bcl-xl protein level thatmay lead to apoptotic death inRAW264.7 cells. Furthermore,DNAmicroarray assay indicated that citosol significantly promoted the expression of 5 genes (Gm4884, Gm10883, Lce1c, Lrg1, and LOC100045878) and significantly inhibited the expression of 24 genes (Gm10679, Zfp617, LOC621831, Gm5929, Snord116, Gm3994, LOC380994, Gm5592, LOC380994, LOC280487, Gm4638, Tex24, A530064D06Rik, BC094916, EG668725, Gm189, Hist2h3c2, Gm8020, Snord115, Gm3079, Olfr198, Tdh, Snord115, andOlfr1249). Based on these observations, citosol induced cell apoptosis and influenced gene expression in mice leukemia RAW264.7 cells in vitro.
Keywords
citosol, apoptosis, cytochrome c, caspase 3, cDNA microarray, RAW 264.7 cells
Introduction
Cancer is the major cause of death in human populations worldwide and leukemia is one of the lethal cancers. 1,2 In Taiwan, about 4 of 100,000 people die of leukemia each year. Based on the
reports in 2009 from the Department of Health, R.O.C. (Taiwan), leukemia is the 11th most
common malignancy in Taiwan.3 Current therapies for leukemia include intensive
chemotherapy and/or hematopoietic stem cell transplantation. 4,5 In addition, imatinib for
chronic myelogenous leukemia and all-trans retinoid acid for acute promyelocytic leukemia, both of which have markedly
improved the prognosis by inhibiting the activities of key molecules critical for leukemogenesis.6,7 However, these treatments for leukemia are still
unsatisfactory due to low cure rate and occurrence of numerous side effects
transcription factors play important roles in leukemogenesis.8–10 A large number of studies have focused on trying new agents or novel targets for leukemic patients.11,12 Citosol (thiamylal
sodium) has been used as an intravenous hypnotic drug for anesthesia.13–16 However, no
information regarding the effects of citosol on the cytotoxic effects, apoptosis and gene expressions associated with apoptosis inmice leukemia cells is available. Therefore, in the present study, we investigated the effects of citosol on the mice leukemia RAW 264.7 cells in vitro by examining the cytotoxic effects by measuring the gene expressions and associated proteins involved in the
process of apoptosis and cell viability.
Materials and methods
Materials, chemicals, and reagents
Thiamylal sodium (citosol) was purchased from
Shinlin Sinseng Pharmaceutical Co. Ltd (Taoyuan, Taiwan). Stock solution of citosol was dissolved in phosphate-buffered saline (PBS), and an equal volume of PBS (1%) was added to the well as controls. RNaseA, propidium iodide (PI), and
4,6-diamidino-2-phenylindole (DAPI) were purchased from Sigma- Aldrich Corp. (St Louis, MO, USA). RPMI 1640 medium, fetal calf serum (FBS), penicillin and streptomycin
antibiotic mixture were purchased from Invitrogen Life Technologies (Grand Island, NY, USA). These primary antibodies against Fas, cytochrome c, apoptosis-inducing factor (AIF), endonuclease G (Endo G), caspase 9 and 3, Bax and Bcl-2, and secondary antibodies were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Cell culture
The RAW 264.7 mouse leukemia cell line was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). The RAW 264.7 cells were incubated in 5% CO2 and at 37_C in RPMI 1640 medium with 2 mM L-glutamine, supplemented with 10% heat-inactivated FBS and 1% antibiotic/ antimycotic (100 Units/ml penicillin and 100 mg/ml streptomycin).17
Determination of morphology and percentage of viable cells
To examine the cell morphological changes, the cells from each treatment were examined and photographed under a phase-contrast microscope at _200 magnification as described
elsewhere.18,19 The quantitative analysis of cell viability was performed
previously described.19,20 RAW 264.7 cells at a density of 2 _ 105 cells/ml were cultured in
12-well plates and treated with 0, 0.375, 0.75, 1.5, 3, and 6 mg/ml of citosol followed by incubation for 24 h. At the end of incubation, cells were harvested from each treatment and stained with PI (4 mg/ml). Percentage of viable cells was measured by flow cytometry. DAPI nuclear staining for chromatin condensation in apoptosis
RAW 264.7 cells at a density of 2 _ 105 cells/well were seeded in 12-well plates, were exposed to 3 mg/ml of citosol and fixed in 4% paraformaldehyde solution (Sigma-Aldrich Corp.) for 15 min followed by staining with DAPI (300 nmol/L; DAPI, Invitrogen) for 30 min at 25_C. Then the nuclear morphology or apoptotic body were visualized, examined, and photographed by fluorescence microscopy. Apoptotic cells were recognized by the condensed, fragmented, degraded nuclei and apoptotic body.21,22
Western blotting assay for examining the changes in apoptosis-associated protein levels RAW 264.7 cells were placed into 6-well plates at a density of 5 _ 105 cells/ml in RPMI 1640 medium with 10% FBS for 24 h. Cells in each well were treated without and with 3 mg/ml of citosol for 48 and 72 h. Cells were collected and total protein from each treatment was extracted into a high-salt buffer (PROPREP TM protein extraction solution, iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea) and
centrifuged at 12,000 rpm for 10 min at 4_C. The quantitated total protein from each treatment was determined by Bradford assay (Bio-Rad protein assay kit; Bio-Ray Laboratories,
Richmond, CA, USA).23,24
Proteins from each treatment were resolved on an SDS-polyacrylamide gel via electrophoresis (SDSPAGE) and transferred to nitrocellulose membranes (GE Healthcare, Piscataway, NJ, USA). The membranes were incubated with a blocking buffer of 5% nonfat dry milk in Tris-buffered saline (TBS) containing TBS-Tween-20 (TBS-T) for 1 h at room temperature and then incubated with the specific primary
antibodies (anti-Fas, -cytochrome c, -AIF, -Endo G, -caspase 9 and -caspase 3, -Bax, and -Bcl-2). The membranes were washed, treated with appropriate horseradish peroxidase–conjugated secondary antibodies and visualized by a chemiluminescence detection kit (GE Healthcare, Princeton, NJ, USA).18,24
cDNA microarray assay for gene expression of RAW 264.7 cells after exposure to citosol RAW 264.7 cells (5 _ 105 cells/ml) were seeded in 6- well plates containing RPMI 1640 medium with 10% FBS for 24 h. Cells in each well were treated for 48 h with or without3 mg/ml of citosol. Then the cells from each treatment were harvested and the total RNA was
total RNA were used for cDNA synthesis and labeling, microarray hybridization, and followed by flour-labeled cDNA hybridized their complements on the chip (Affymetrix GeneChip Human Gene 1.0 ST array,
Affymetrix, Santa Clara, CA, USA). Finally the resulting localized concentrations of
fluorescent molecules were detected and quantitated (Asia BioInnovations Corporation). The resulting data were analyzed by Expression Console software (Affymetrix) with default RMA parameters. Genes regulated by
citosol were determined with a twofold change.26
Statistical analysis
Significance of mean values between citosol-treated group and control group was obtained using Student’s t test. Data were expressed as mean + SD. The p value <0.05 was considered significant.
Results
Citosol induced cell morphological changes and decreased the percentage of RAW 264.7 viable cells
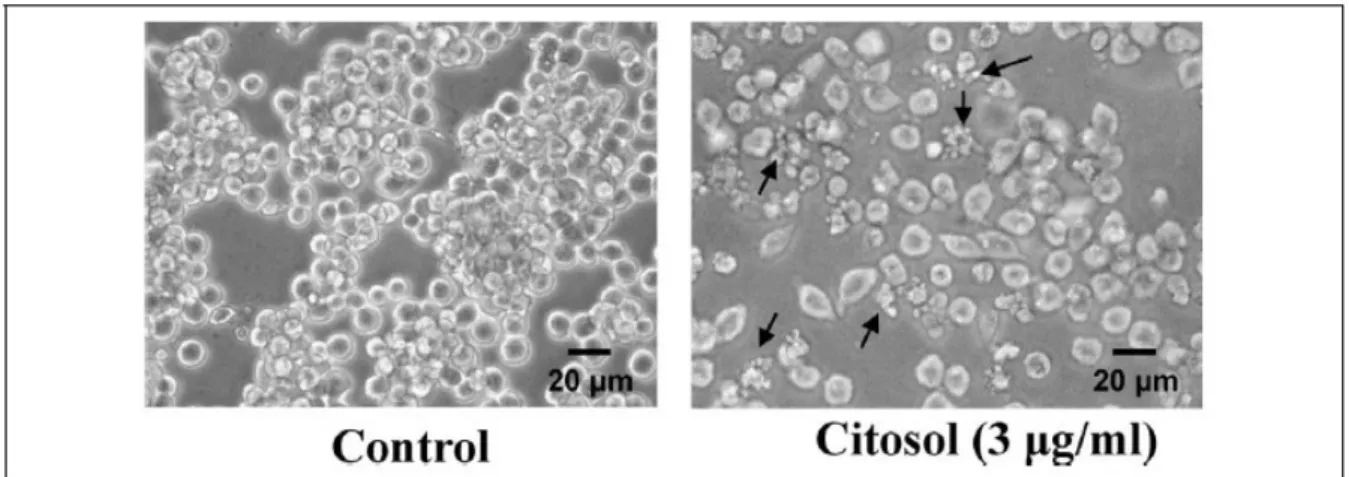
Morphological changes of RAW 264.7 cells after treatment with citosol were examined and investigated. The untreated cells (control) were maintained
in original good shape. The incubation for 24 h at 3 mg/ml of citosol revealed condensation both in cytoplasm and nucleus and also apoptotic body appeared
in the majority of cells (Figure 1). Citosol treatment led to cellular morphological changes and more cells floated on the well compared to the control. Results from flow cytometric assay are shown in Figure 2, and indicated that citosol decreased the percentage of viability and citosol at a concentration of 3–6 mg/ml led to a decrease of 55% and 81%, respectively. Also, the
effective concentration for half-maximal response (EC50) values at 48 h exposure for citosol is 2.74 mg/ml using a PI exclusion method and flow cytometry. Hence, the dose of 3 mg/ml of citosol is close to EC50 and used for further examinations.
Figure 1. Citosol affected the cell morphology of RAW 264.7 cells. Cells cultured for 48 h with or without 3 mg/ml of citosol were examined for cell morphological changes and were
photographed by a phase-contrast microscope as described in the section on Materials and methods. Scale bar, 20 mm.
Citosol induced apoptotic bodies and DNA damage in RAW 264.7 cells
Citosol-induced cell morphological changes and the reduction in the total viable cells in RAW 264.7 cells were further investigated using fluorescence photomicrographs.
RAW 264.7 cells were stained with DAPI after exposure to 3 mg/ml citosol for 48 h. The results are shown in Figure 3, which indicated that control cells were round with homogeneous
nuclei, whereas citosol-treated cells showed apoptotic body (arrows; Figure 3).
Figure 3. Citosol induced chromatin condensation and DNA fragmentation in RAW 264.7 cells. Cells were treated with 3 mg/ml concentration of citosol for 48 h, and chromatin condensation and DNA fragmentation (a characteristic of apoptosis) were examined by 4,6-diamidino-2-phenylindole (DAPI) staining then cells were photographed by fluorescence
microscopy as described in the section on Materials and methods. Arrows indicated damaged cells with apoptotic cells. Scale bar, 10 mm.
Citosol changed the levels of proteins associated with apoptosis in RAW 264.7 cells
To examine whether citosol-induced apoptosis is related to the response of apoptosis-associated proteins, we investigated the effects of citosol on the levels
of proteins associated with apoptosis in RAW 264.7 cells and the results were shown in Figure 4. The results indicated that citosol increased the protein levels of Fas/CD95, cytochrome c, AIF, Endo G, active form of caspase 9 and 3 and Bax, but it decreased the level of Bcl-xl in RAW 264.7 cells. Hence, we suggest that cytosol-triggered apoptotic death of RAW 264.7 cells is carried out through
cDNA microarray analysis for citosol-induced gene expression in RAW 264.7 cells The microarray analysis showed that citosol increased 5 genes which were upregulated including Gm4884, Gm10883, Lce1c, Lrg1, and LOC100045878 and it
inhibited 24 genes which were downregulated including Gm10679, Zfp617, LOC621831, Gm5929, Snord116, Gm3994, LOC380994, Gm5592, LOC380
994, LOC280487, Gm4638, Tex24, A530064D06Rik, BC094916, EG668725, Gm189, Hist2h3c2, Gm8020, Snord115, Gm3079, Olfr198, Tdh, Snord115, and
Olfr1249 in RAW 264.7 cells that were expressed at least by a twofold change when compared with the untreated control cells (Table 1). Moreover, the gene and mRNA descriptions,
respectively, can be seen in Table 1. Overall, the results from cDNA microarray available information addressing whether citosol affects cellular processes related to cancer. However, there are no reports on the effects of citosol on apoptosis and associated gene expressions in mice leukemia cells. Herein, the present study is
the first to show that citosol can induce cytotoxic effect including induction of apoptosis and the associated gene expression changes in mice leukemia cells
In this study, our results demonstrated that citosol induced cell morphological changes, decreased the percentage of total viable cells, induced apoptotic bodies as well as changed the apoptosis-associate protein levels, and gene expressions in mice leukemia RAW 264.7 cells in vitro. The EC50 of citosol for
cytotoxic effects is 2.74 mg/ml of cytosol for 48-h exposure (Figure 1). This is the first study to show that citosol at a concentration of 3–6 mg/ml can decrease
the percentage of viable RAW 264.7 cells
Much evidence revealed that agent- or chemical compound-induced cell death would have two characteristics: necrosis and apoptosis.28,29 Furthermore, the
character of apoptosis included chromatin condensation and DNA damage (DNA fragmentation), apoptotic body formation, and so on.30,31 Thus, we used DAPI
staining to show whether citosol could induce chromatin condensation and DNA damage, and the result showed that citosol triggered apoptosis in RAW 264.7 cells (Figure 3)
It is well known that the two apoptotic pathways are extrinsic and intrinsic pathways.32,33 The extrinsic pathway involved the agent that induced Fas/CD95
and Fas-associated protein with death domain, which led to the activation of caspase 8 and 3, causing cell apoptosis. The intrinsic pathway involved the increase in Bax (a pro-apoptotic protein) and the decrease in Bcl-2 (an antiapoptotic protein) that
cause mitochondrial dysfunction (decrease the levels of mitochondrial membrane potential) with consequent release of cytochrome c release and activation
of caspase 9 and 3, causing cell apoptosis.32,33 Therefore, we used Western blotting to examine
whether citosol would affect apoptotic protein levels, and the results indicated that citosol promoted the levels of AIF and Endo G, and it also stimulated the level of cytochrome c in RAW 264.7 cells (Figure 4). These observations indicated that citosol induced apoptosis through a mitochondria-dependent pathway.
Figure 4 also showed that citosol promoted the level of Bax and decreased the level of Bcl-xl, which implied that citosol decreased the level of mitochondrial membrane potential (DCm). In the present study, our results showed that citosol decreased the
level of Bcl-xl and promoted the level of Bax in RAW 264.7 cells, which affected the ratio of Bax/ Bcl-2, leading to AIF and Endo G release from mitochondria. It was reported that some inducer of apoptosis could induce apoptosis through mitochondria
and led to AIF and Endo G release, causing apoptosis. 34 It was also reported that the inducer of
apoptosis decreased the ratio of Bax and Bcl-xl and led to mitochondrial dysfunction thereby
To confirm the cytotoxic effects of citosol on apoptotic proteins involved in the associated gene expression in RAW 264.7 cells, we used Cdna microarray to examine the gene expression from RAW 264.7 cells after exposure to citosol, and our results showed that citosol
significantly promoted the expression of 5 genes, including Gm4884, Gm10883, Lce1c, Lrg1, and LOC100045878, but it significantly inhibited the expression of 24 genes, including Gm10679, Zfp617, LOC621831, Gm5929,
Snord116, Gm3994, LOC380994, Gm5592, LOC38 0994, LOC280487, Gm4638, Tex24, A530064D06Rik, BC094916, EG668725, Gm189, Hist2h3c2, Gm8020,
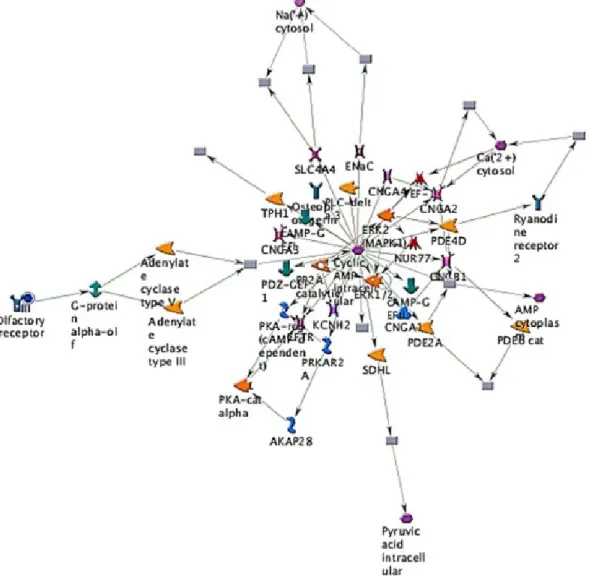
Snord115, Gm3079, Olfr198, Tdh, Snord115, and Olfr1249 in RAW 264.7 cells as can be seen in Figure 5 and Table 1.
Figure 5. The representative possible canonical pathways from RAW 264.7 cells after exposure to citosol. Cells were exposed to 3 mg/ml of citosol, were harvested and total RNA were extracted for cDNA microarray assay. Thick cyan lines indicate the fragments of canonical pathways. Upregulated genes are marked with red circles; downregulated with blue
circles. The ‘checkerboard’ color indicates mixed expression for the gene between files or between multiple tags for the same gene (AKAP28: A-kinase anchor protein, 28 kD; CNGA1: cyclic nucleotide–gated channel alpha 1; CNGA2: cyclic nucleotide–gated channel alpha 2; CNGA3: cyclic nucleotide–gated channel alpha 3; CNGA4: cyclic nucleotide–gated
channel alpha 4; KCNH2: potassium voltage–gated channel, subfamily H (eag-related), member 2; SDHL: L-serine ammonia-lyase; SLC4A4: solute carrier family 4, sodium bicarbonate cotransporter, member 4; PDE4D: phosphodiesterase 4D, cAMP-specific; PDE2A: phosphodiesterase 2A, cGMP-stimulated; PRKAR2A: protein kinase, cAMP-dependent, regulatory, type II, alpha; TPH1: tryptophan hydroxylase 1).
In conclusion, based on these observations, citosol induced cytotoxic effects and the protein levels and gene expressions in mice leukemia RAW 264.7 cells
in vitro, and our study is the first to show the cytotoxic and gene regulatory effects of citosol on mice leukemia RAW 246.7 cells
Funding
This work was supported by the grant DOH100-TDC- 111-005 from Taiwan Department of Health, China Medical University Hospital Cancer Research Center
of Excellence.
References
1. Anand P, Kunnumakkara AB, Sundaram C, et al. Cancer is a preventable disease that requires major lifestyle changes. Pharm Res 2008; 25: 2097–2116.
2. Wiemels JL, Cazzaniga G, Daniotti M, et al. Prenatal origin of acute lymphoblastic leukaemia in children. Lancet 1999; 354: 1499–1503.
3. Lin CC, Kuo CL, Lee MH, et al. Extract of Hedyotis diffusa willd influences murine
leukemia WEHI-3 cells in vivo as well as promoting T- and B-cell proliferation in leukemic mice. In Vivo 2011; 25: 633–640.
4. Nau KC and Lewis WD. Multiple myeloma: diagnosis and treatment. Am Fam Physician 2008; 78: 853–859.
5. Fotoohi AK, Assaraf YG, Moshfegh A, et al. Gene expression profiling of leukemia T-cells resistant to methotrexate and 7-hydroxymethotrexate reveals
alterations that preserve intracellular levels of folate and nucleotide biosynthesis. Biochem Pharmacol 2009; 77: 1410–1417.
6. Yoshimi A and Kurokawa M. Key roles of histone methyltransferase and demethylase in leukemogenesis. J Cell Biochem 2011; 112: 415–424.
7. Yang JS, Wu CC, Kuo CL, et al. Solanum lyratum extract affected immune response in normal and leukemia murine animal in vivo. Hum Exp Toxicol 2010; 29: 359–367.
8. Lorsbach RB and Downing JR. The role of the AML1 transcription factor in leukemogenesis. Int J Hematol 2001; 74: 258–265.
proteins-implication for MLL-induced leukemogenesis. Leuk Lymphoma 2001; 42: 595– 602.
10. Ayton PM and Cleary ML. Molecular mechanisms of leukemogenesis mediated by MLL fusion proteins. Oncogene 2001; 20: 5695–5707.
11. Gratacap MP, Martin V, Valera MC, et al. The new tyrosine-kinase inhibitor and anticancer drug dasatinib reversibly affects platelet activation in vitro and in vivo. Blood 2009; 114: 1884–1892.
12. Matsunaga T, Fukai F, Miura S, et al. Combination therapy of an anticancer drug with the FNIII14 peptide of fibronectin effectively overcomes cell adhesion-mediated drug resistance of acute myelogenous leukemia. Leukemia
2008; 22: 353–360.
13. Hori Y, Hiraga K, and Watanabe S. The effects of thiamylal sodium on the tonic vibration reflex in man. Brain Res 1989; 497: 291–295.
14. Shimoji K, Kano T, Nakashima H, et al. The effects of thiamylal sodium on electrical activities of the central and peripheral nervous systems in man. Anesthesiology 1974; 40: 234–240.
15. Gaynor JS, Bednarski RM, and MuirWW, III. Effect of hypercapnia on the arrhythmogenic dose of epinephrine in horses anesthetized with guaifenesin,
thiamylal sodium, and halothane. Am J Vet Res 1993; 54: 315–321.
16. Mokriski BK, Nagle SE, Papuchis GC, et al. Electroconvulsive therapy-induced cardiac arrhythmias during anesthesia with methohexital, thiamylal, or thiopental sodium. J Clin Anesth 1992; 4: 208–212.
17. Chiou WF, Don MJ, Liao JF, et al. Psoralidin inhibits LPS-induced iNOS expression via repressing Syk-mediated activation of PI3K-IKK-IkappaB signaling
pathways. Eur J Pharmacol 2011; 650: 102–109.
18. Yu FS, Yang JS, Yu CS, et al. Safrole induces apoptosis in human oral cancer HSC-3 cells. J Dent Res 2011; 90: 168–174.
19. Wu PP, Liu KC, Huang WW, et al. Triptolide induces apoptosis in human adrenal cancer NCI-H295 cells through a mitochondrial-dependent pathway. Oncol Rep 2011; 25: 551–557. 20. Wu SH, Hang LW, Yang JS, et al. Curcumin induces apoptosis in human non-small cell
lung cancer NCI-H460 cells through ER stress and caspase
cascade-and mitochondria-dependent pathways. Anticancer Res 2010; 30: 2125–2133.
21. Lu CC, Yang JS, Huang AC, et al. Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells.
Mol Nutr Food Res 2010; 54: 967–976.
22. Lu HF, Chie YJ, Yang MS, et al. Apigenin induces apoptosis in human lung cancer H460 cells through caspase- and mitochondria-dependent pathways. Hum
23. Chen HY, Lu HF, Yang JS, et al. The novel quinolone CHM-1 induces DNA damage and inhibits DNA repair gene expressions in a human osterogenic sarcoma cell line. Anticancer Res 2010; 30: 4187–4192.
24. Chiang JH, Yang JS, Ma CY, et al. Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human
gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chem Res Toxicol 2011; 24: 20–29.
25. Lin CC, Chen JT, Yang JS, et al. Danthron inhibits the migration and invasion of human brain glioblastoma multiforme cells through the inhibition of mRNA expression of focal adhesion kinase, Rho kinases-1 and metalloproteinase-9. Oncol Rep 2009; 22: 1033–1037. 26. Jacobs AT and Marnett LJ. HSF1-mediated BAG3 expression attenuates apoptosis in
4-hydroxynone nal-treated colon cancer cells via stabilization of anti-apoptotic Bcl-2 proteins. J Biol Chem 2009; 284: 9176–9183.
27. Orallo MO and Eather KF. Sodium methohexital as a rectal agent in pediatric anesthesia: a controlled comparison with sodium thiamylal. Anesth Analg 1965; 44: 97–103.
28. Madge LA, Sierra-Honigmann MR, and Pober JS. Apoptosis-inducing agents cause rapid shedding of tumor necrosis factor receptor 1 (TNFR1). A nonpharmacological explanation for inhibition of TNF-mediated activation. J Biol Chem1999; 274: 13643–13649.
29. Jori G and Fabris C. Relative contributions of apoptosis and random necrosis in tumour response to photodynamic therapy: effect of the chemical structure of
Zn(II)-phthalocyanines. J Photochem Photobiol B 1998; 43: 181–185.
30. Lu C, Fu W, and Mattson MP. Caspase-mediated suppression of glutamate (AMPA) receptor channel activity in hippocampal neurons in response to DNA damage promotes apoptosis and prevents necrosis: implications for neurological side effects of cancer therapy and neurodegenerative disorders. Neurobiol Dis 2001; 8: 194–206.
31. Fenech M, Crott J, Turner J, et al. Necrosis, apoptosis, cytostasis andDNAdamage in human lymphocytesmeasured simultaneously within the cytokinesis-block micronucleus assay: description of the method and results for hydrogen peroxide. Mutagenesis 1999; 14: 605– 612.
32. Tsuchiya M, Asada A, Arita K, et al. Induction and mechanism of apoptotic cell death by propofol in HL-60 cells. Acta Anaesthesiol Scand 2002; 46: 1068–1074.
33. Chien SY, Wu YC, Chung JG, et al. Quercetin-induced apoptosis acts through
mitochondrial- and caspase-3- dependent pathways in human breast cancer MDA-MB-231 cells. Hum Exp Toxicol 2009; 28: 493–503.
34. Liu KC, Huang YT, Wu PP, et al. The roles of AIF and Endo G in the apoptotic effects of benzyl isothiocyanate on DU 145 human prostate cancer cells via the mitochondrial signaling pathway. Int J Oncol 2011; 38: 787–796.
beyond. Nat Rev Mol Cell Biol 2010; 11: 621–632.
36. Lu HF, Wang HL, Chuang YY, et al. Danthron induced apoptosis through mitochondria- and caspase-3- dependent pathways in human brain glioblastoma multiforms GBM 8401 cells. Neurochem Res 2010; 35: 390–398.