Ginger and Zingerone Ameliorate Lipopolysaccharide-Induced Acute Systemic Inflammation in Mice, Assessed by Nuclear Factor-κB Bioluminescent Imaging
Chien-Yun Hsiang,a,1 Hui-Man Cheng,b,1 Hsin-Yi Lo,c Chia-Cheng Li,d Pei-Chi Chou,b Yu-Chen Lee,e Tin-Yun Ho*,c,g
aDepartment of Microbiology, China Medical University, Taichung 40402, Taiwan bSchool of Chinese Medicine, China Medical University, Taichung 40402, Taiwan
cGraduate Institute of Chinese Medicine, China Medical University, Taichung 40402, Taiwan dGraduate Institute of Cancer Biology, China Medical University, Taichung 40402, Taiwan eGraduate Institute of Acupuncture Science, China Medical University, Taichung 40402,
Taiwan
gDepartment of Health and Nutrition Biotechnology, Asia University, Taichung 41354, Taiwan
*Corresponding author. Telephone: +886 4 22053366 ext. 3302. Fax: +886 4 22032295. E-mail: [email protected]
1 These authors equally contributed to this work.
Short title: ginger and zingerone ameliorate LPS-induced inflammation 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
ABSTRACT
Ginger is a commonly used spice in cooking. In this study, we comprehensively evaluated the anti-inflammatory activities of ginger and its component zingerone in lipopolysaccharide (LPS)-induced acute systemic inflammation in mice via nuclear factor-κB (NF-κB) bioluminescent imaging. Ginger and zingerone significantly suppressed LPS-induced NF-κB activities in cells in a dose-dependent manner, and the maximal inhibition (84.5±3.5% and 96.2±0.6%) was observed at 100 μg/ml ginger and zingerone, respectively. Moreover, dietary ginger and zingerone significantly reduced LPS-induced proinflammatory cytokine production in sera by 62.9±18.2% and 81.3±6.2%, respectively, and NF-κB bioluminescent signals in whole body by 26.9±14.3% and 38.5±6.2%, respectively. In addition, ginger and zingerone suppressed LPS-induced NF-κB-driven luminescent intensities in most organs and the maximal inhibition by ginger and zingerone was observed in small intestine. Immunohistochemical staining further showed that ginger and zingerone decreased interleukin-1β (IL-1β)-, CD11b-, and p65-positive areas in jejunum. In conclusion, our findings suggested that ginger and zingerone were likely to be broad-spectrum anti-inflammatory agents in most organs that suppressed the activation of NF-κB, the production of IL-1β, and the infiltration of inflammatory cells in mice.
Key words: Ginger, zingerone, nuclear factor-κB, inflammation, bioluminescent imaging 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37
INTRODUCTION
Ginger, the rhizome of Zingiber officinale, is one of the most commonly used spices in cooking. It is also a frequently used herb in alternative medicines. Clinical studies have shown that ginger is effective in ameliorating nausea and vomiting caused by anti-retroviral therapy, pregnancy, post-operation, and chemotherapy.1,2 It has add-on effects on reducing knee pain and improving knee function in patients with symptomatic knee osteoarthritis.3,4 It is effective on pain relief in primary dysmenorrhea, eccentric exercise, and migraine.5,6 Moreover, consumption of ginger is useful for patients with type 2 diabetes due to the reduction of glycated hemoglobin and the improvement of insulin resistance.7 These clinical data indicate the pharmacological application of ginger in medicine.
Anti-inflammatory activities of ginger and its ingredients have been suggested in in vitro studies. For example, ginger extract inhibits the production of nitric oxide (NO) and proinflammatory cytokines in lipopolysaccharide (LPS)-stimulated microglial cells, inhibits the activation of macrophages, and reduces the production of LPS-induced proinflammatory chemokines in bronchial epithelial cells.8,9 Moreover, ginger constituents, such as 6-shogaol, gingerol and 6-dehydroginerdione, display anti-inflammatory potentials in LPS-induced microglial cells or macrophages by inhibiting the production of cytokines.10,11 Anti-inflammatory effects of ginger and it ingredients have also been evaluated in individual organs, such as liver, brain, lung, and colon.12 For example, 6-shogaol suppresses the 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56
microglial activation in an in vivo neuroinflammatory model and shows a neuroprotective effect in transient global ischemia.13,14 Zingerone, a phenolic alkanone of ginger extract, attenuates LPS-induced acute lung injury and hepatic injury in mice.15 Moreover, zingerone improves experimental colitis in mice via nuclear factor-κB (NF-κB) activity in our previous study.16 However, these studies raise a question: do ginger extract and its constituents exhibit broad-spectrum anti-inflammatory effects in most organs?
To address this question, we applied bioluminescent imaging on LPS-induced transgenic mice, which carried NF-κB-driven luciferase genes, to comprehensively monitor the anti-inflammatory effects of ginger and zingerone in whole body and organs. NF-κB bioluminescent imaging has been applied to assess host responses to the implantation of biomaterials and the exposure of ionizing radiation.17,18 It has been used to evaluate the anti-inflammatory potentials of vanillin and ginger extract on experimental colitis.16 It also has been utilized to monitor the anti-inflammatory effects of medicinal herbs on LPS-induced acute systemic inflammation and carbon tetrachloride-induced chronic hepatitis.19,20 Immunohistochemical (IHC) staining was further performed to elucidate the anti-inflammatory mechanisms of ginger and zingerone. Our findings suggested that ginger and zingerone were likely to be broad-spectrum anti-inflammatory agents in most organs that suppressed the activation of NF-κB, the production of interleukin-1β (IL-1β), and the infiltration of inflammatory cells.
57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75
MATERIALS AND METHODS
Chemicals. LPS (from Escherichia coli 055:B5), zingerone, and 3-(4,5-dimethylthiazol-2-yl-2,5-diohenyl tetrazolium bromide (MTT) were purchased from Sigma (St. Louis, MO). MG-132, a NF-κB inhibitor, was purchased from Santa Cruz (Dallas, TX). D-Luciferin was purchased from Xenogen (Hopkinton, MA). Mouse monoclonal antibody against p65 was purchased from Chemicon (Temecula, CA). Rabbit polyclonal antibodies against IL-1β and CD11b were purchased from Santa Cruz (Dallas, TX) and Abcam (Cambridge, UK), respectively.
Preparation of Ginger Extract. Dried ginger was purchased from Xin Lung Chinese Herbal Medicine Pharmacy (Taichung, Taiwan). The voucher specimen has been deposited in Graduate Institute of Chinese Medicine, China Medical University. Ginger was ground to a fine powder and extracted by mixing 20 g powder with 100 ml ethanol at room temperature with shaking. Three days later, the supernatant was collected and stored at -30C for further analysis.
Cell Culture. Recombinant HepG2/NF-κB cells, which carried NF-κB-driven luciferase genes, were constructed previously.19 HepG2/NF-κB cells were maintained in Dulbecco's modified Eagle's medium (Life Technologies, Gaithersburg, MD) supplemented with 10% 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95
fetal bovine serum (Hyclone, Logan, Utah) and incubated at 37C with 5% CO2.
Luciferase Assay and Cell Viability Assay. HepG2/NF-κB cells (2×107 cells) were seeded in a 96-well plate and incubated at 37C overnight. LPS (100 ng/ml), MG-132 (5 μM), or various amounts of ginger and zingerone were then added to cells and incubated at 37C for 24 h. Cell viability was analyzed by MTT colorimetric assay as described previously.19 Luciferase assay was performed as described previously.19 Relative NF-κB activity was calculated by dividing the relative luciferase unit (RLU) of compound-treated cells by the RLU of solvent-treated cells.
Animal Experiments. Transgenic mice, carrying the luciferase genes driven by NF-B-responsive elements, were constructed as described previously.17 Mouse experiments were conducted under ethics approval from China Medical University Animal Care and Use Committee (Permit No. 97-28-N).
Six-week-old female transgenic mice were randomly divided into four groups of five mice: (1) mock, no treatment; (2) LPS, (3) LPS/ginger, and (4) LPS/zingerone. Mice were challenged intraperitoneally with 1 mg/kg LPS and then orally with 100 mg/kg ginger extract or zingerone 10 min later. Four hours later, mice were imaged for the luciferase activity and subsequently sacrificed for ex vivo imaging and IHC staining.
96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115
In Vivo and Ex Vivo Bioluminescence Imaging. Bioluminescence imaging was
performed as described previously.17 Briefly, for in vivo imaging, mice were injected intraperitoneally with 150 mg/kg D-luciferin, placed in the IVIS Imaging System® 200 Series chamber (Xenogen, Hopkinton, MA) 5 min later, and imaged for 1 min. Photons emitted from bodies were quantified using Living Image® software (Xenogen, Hopkinton, MA).The intensity of the signal from bodies was quantified as the sum of all photon counts per second and presented as photon/sec. For ex vivo imaging, mice were injected with D -luciferin and sacrificed 5 min later. The organs were removed immediately, placed in the IVIS chamber, and imaged for 1 min. The intensity of signal was quantified as the sum of all detected photon counts per second with the region of interest and presented as photon/sec/cm2/steradian (sr).
Cytokine Enzyme-Linked Immunosorbent Assay (ELISA). The amounts of proinflammatory cytokines, including IL-1β and tumor necrosis factor-α (TNF-α), in sera were quantified using Quantikine® Mouse ELISA kits (R&D Systems, Minneapolis, MN). Briefly, mouse sera were added to wells coated with anti-IL-1β or anti-TNF-α antibodies and incubated at room temperature for 2 h. Biotinylated anti-mouse IL-1β or TNF-α antibodies, and avidin-horseradish peroxidase were added sequentially to wells. After a final wash, chromogenic substrate (tetramethylbenzidine) was added and the reaction was stopped with 2 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134
N H2SO4. The absorbance at 450 nm was measured using an ELISA reader (Multiskan GO, Thermo Scientific, Waltham, MA).
IHC Staining. Parafilm-embedded small intestines were cut into 5-µm-thick sections, deparaffinized in xylene, and rehydrated in graded ethanol. Sections were incubated with anti-p65, anti-IL-1β, or anti-CD11b antibodies overnight at 4C and then incubated with a biotinylated secondary antibody (Zymed Laboratories, Carlsbad, CA) for 20 min at room temperature. Finally, the sections were incubated with avidin-biotin complex reagent and stained with 3,3'-diaminobenzidine according to manufacturer’s protocol (Histostain®-Plus, Zymed Laboratories, Carlsbad, CA).
Statistics Analysis. Data were presented as mean ± standard error. Student’s t-test was used for the comparison between two experiments. A value of p < 0.05 was considered statistically significant.
RESULTS
Ginger and Zingerone Suppressed LPS-Induced NF-κB Activities in Cells. Zingerone is a phenolic alkanone of ginger extract (Figure 1), and the content of zingerone was approximately 0.01 mg/ml in ethanolic extract of ginger by high-performance liquid 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154
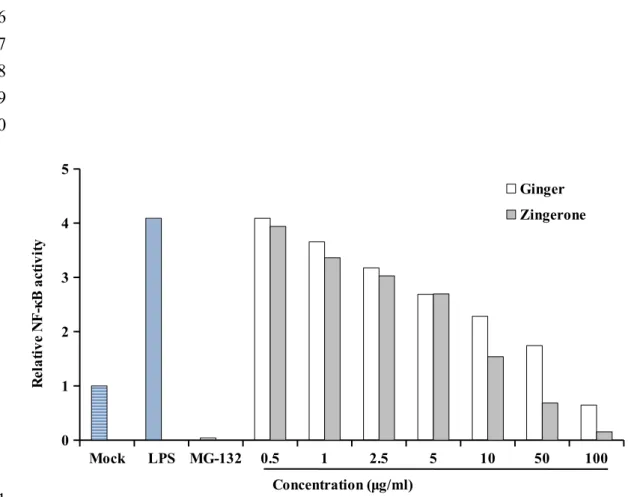
chromatography analysis (see Supplementary Figure 1 of the Supporting Information). We first analyzed the effects of ginger and zingerone on LPS-induced NF-κB activation in cells. HepG2/NF-κB cells were treated with LPS, followed by MG-132 or various amounts of ginger and zingerone. As shown in Figure 2, LPS increased the NF-κB activity by 4-fold, compared with mock. MG-132, a well-known NF-κB inhibitor, significantly suppressed LPS-induced NF-κB activities. Ginger and zingerone decreased NF-κB activities LPS-induced by LPS, and the decrease displayed a dose-dependent manner. The maximal inhibition (84.5±3.5% and 96.2±0.6%) was observed at 100 μg/ml ginger and zingerone, respectively. Moreover, zingerone was more effective than ginger on the inhibition of LPS-induced NF-κB activity. No visible cytotoxic effects were observed, judged by MTT assay (data not shown). These findings suggested that ginger and zingerone significantly suppressed NF-κB activities induced by LPS in cells.
Ginger and Zingerone Suppressed LPS-Induced Inflammation in Mice. The in vivo anti-inflammatory effects of ginger and zingerone were then analyzed by NF-κB bioluminescent imaging. Ginger extract or zingerone was orally given to transgenic mice, which has been challenged by 1 mg/kg LPS. The NF-κB-dependent bioluminescence was monitored 4 h later. As shown in Figure 3, LPS induced an approximately 4-fold increase in NF-κB-driven luminescent intensity, compared with mock. The induced luminescence was 155 156 157 158 159 160 161 162 163 164 165 166 167 168 169 170 171 172 173
observed over the whole body, and the strongest luminescence appeared in the abdominal region. Ginger and zingerone significantly decreased the LPS-induced luminescent intensity by 26.9±14.3% and 38.5±6.2%, respectively. These findings suggested that ginger and zingerone suppressed LPS-induced NF-κB-dependent luminescence in mice.
NF-κB plays a crucial role in the regulation of immunity. We wondered whether the intensity of NF-κB-driven luminescence was correlated with inflammation. The amount of proinflammatory cytokines, including IL-1β and TNF-α, in sera were therefore quantified by ELISA. As shown in Figure 4, LPS significantly increased the amount of IL-1β and TNF-α in sera by 55.6±7.6 and 172±57.5 fold, respectively. However, ginger and zingerone significantly decreased LPS-induced IL-1β and TNF-α production in sera. Ginger reduced the production of IL-1β and TNF-α by 73.5±23% and 63.9±18.2%, respectively, while zingerone decreased IL-1β and TNF-α production by 79.9±10.4% and 81.3±6.2%, respectively. These data suggested that ginger and zingerone suppressed LPS-induced systemic inflammation in mice. Moreover, the correlation between NF-κB-dependent luminescent intensity and cytokine production indicated the representative of NF-κB-driven luminescence on the degree of inflammation.
Ginger and Zingerone Suppressed LPS-Induced Inflammation in Most Organs. Previous studies have shown that ginger or zingerone displayed anti-inflammatory efficacies 174 175 176 177 178 179 180 181 182 183 184 185 186 187 188 189 190 191 192
in individual organs, such as liver, lung, brain, and colons. We would like to know whether ginger and zingerone exhibited broad-spectrum anti-inflammatory actions in most organs. Transgenic mice were therefore challenged with LPS and orally administered with ginger extract and zingerone, and NF-κB-driven luminescence in organs was monitored 4 h later. As shown in Figure 5, luminescent intensities of organs were increased by LPS, suggesting that intraperitoneal injection of LPS induced inflammation in most organs. The maximal induction of luminescence by LPS was observed in kidney (15.9±8 fold), followed by intestine (15.2±6 fold), brain (10.4±1.4 fold), heart (5.7±1.5 fold), liver (4.7±1.9 fold), spleen (4.7±1.2 fold), lung (3.7±1.4 fold), and stomach (2.7±0.7 fold). Administration of ginger and zingerone significantly decreased LPS-induced NF-κB-driven luminescence in most organs. The maximal inhibition of LPS-induced luminescence by ginger and zingerone was observed in intestine, followed by kidney, liver, heart, and brain. Zingerone significantly decreased LPS-induced luminescence in lung, while ginger slightly decreased luminescent signals in lung. Moreover, both ginger and zingerone slightly decreased LPS-induced luminescent intensities in spleen.
We further analyzed the anti-inflammatory effects of ginger and zingerone in different segments of small intestine. We divided the small intestine into 35 segments and the length ratio of duodenum, jejunum, and ileum was 1:3:2. The intensity of signal from each segment was quantified as photon/sec. The suppression of LPS-induced luminescent signal by ginger 193 194 195 196 197 198 199 200 201 202 203 204 205 206 207 208 209 210 211
or zingerone was further represented as the inhibitory percentage. As shown in Figure 6, LPS increased NF-κB-driven luminescence in whole small intestine, and the strong luminescence was observed from segment 17 to 30, which corresponded to the region between mid-jejunum and mid-ileum. Both ginger and zingerone suppressed LPS-induced bioluminescent intensity in whole small intestine, and the inhibition from segment 14 to 35 was > 50% by ginger and zingerone. Overall, these data suggested that ginger and zingerone displayed broad-spectrum anti-inflammatory activities in most organs. Additionally, ex vivo imaging first showed that LPS induced a more severe inflammation in the region spanning from jejunum to mid-ileum. Moreover, LPS-induced luminescent signal in the junction of jejunum and ileum was suppressed efficiently by ginger and zingerone.
Ginger and Zingerone Inhibited LPS-Induced NF-κB Activation, IL-1β Production, and Inflammatory Cell Infiltration in Small Intestine. IHC staining was further performed to analyze the anti-inflammatory effects and mechanisms of ginger and zingerone in small intestine. As shown in Figure 7, the number of IL-1β-positive cells was increased by LPS, compared with mock. However, the expression of IL-1β-positive area was decreased by ginger and zingerone. In addition, LPS increased the number of CD11b-positive cells, including monocytes and granulocytes, while ginger and zingerone inhibited the expression of CD11b-positive area. These data suggested that ginger and zingerone suppressed the 212 213 214 215 216 217 218 219 220 221 222 223 224 225 226 227 228 229 230
production of IL-1β and the infiltration of inflammatory cells, resulting in the amelioration of LPS-inflammation in small intestine.
Because NF-κB plays a critical role in inflammation, we analyzed the level of NF-κB activity by IHC staining. The monoclonal antibody used here was against p65 nuclear localization sequence, which was blocked by inhibitory IκB when NF-κB was inactivated. LPS increased the number of p65-positive cells, while ginger and zingerone decreased the p65-positive area. These findings suggested the inhibition of ginger and zingerone on LPS-induced inflammation might be through NF-κB signaling pathway.
DISCUSSION
In this study, we comprehensively evaluated the anti-inflammatory effects of ginger and zingerone by NF-κB bioluminescent imaging. Because of the light absorption by pigmented molecules and the low spatial resolution of bioluminescent imaging, we performed ex vivo imaging to monitor the effects of ginger and zingerone on individual organs. Previous studies have shown that ginger extract and zingerone display anti-inflammatory activities in specific organs. For example, ginger extracts ameliorate LPS-induced hepatic injury and experimental colitis in mice.16,22 Administration of ginger extracts significantly represses paw and joint swelling in rats with severe chronic adjuvant arthritis.23 Ginger also exhibits a protective role on the diabetic brain by modulating the astroglial response to the injury in rats.24 Moreover, 231 232 233 234 235 236 237 238 239 240 241 242 243 244 245 246 247 248 249 250
zingerone attenuates LPS-induced lung injury in mice.15 By NF-κB bioluminescent imaging, we found that administration of ginger and zingerone inhibited LPS-induced NF-κB-driven luminescence in brain, lung, liver, and colon However, we newly identified that ginger and zingerone rreduced LPS-induced luminescent intensities in heart, stomach, kidney, and small intestine. In addition, the maximal inhibition of LPS-induced luminescent signal by ginger and zingerone was observed in intestines, followed by kidney. The in vivo metabolism or pharmacokinetics of ginger and zingerone has been reported. After oral administration of 2 g ginger extract in human, the glucuronide and sulfate metabolites of ginger components, such as gingerols and shogaol, are detected in plasma and gastrointestinal tract.25 Oral dosage (100 mg/kg) of zingerone in rats results in the urinary excretion of glucuronide and/or sulfate conjugates of zingerone.26 The distribution of ginger extract and zingerone in gastrointestinal tract and kidney might explain their anti-inflammatory effects in small intestine and kidney. Because of the correlation between NF-κB-driven luminescent signals and inflammation, we speculated that ginger and zingerone exhibited broad-spectrum anti-inflammatory activities that suppressed the LPS-induced inflammation in various organs.
Anti-inflammatory mechanisms of ginger and its constituents have been analyzed in in vitro and in vivo studies. For instance, ginger extract ameliorates LPS-induced hepatic injury via inhibiting the production of proinflammatory cytokines and attenuating the mitogen-activated protein kinases and NF-κB signaling pathways.22 It also inhibits the activities of 251 252 253 254 255 256 257 258 259 260 261 262 263 264 265 266 267 268 269
cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS), and thus inhibits the synthesis of prostaglandins and NO, mediators of inflammation.27 Ginger derivatives, such as 6-shogaol, reduce osteoarthritis symptom by inhibiting toll-like receptor 4 (TLR-4)-mediated innate immunity and cathepsin-k activity.28 6-Shogaol also displays a neuroprotective effect via inhibiting iNOS, COX-2, proinflammatory cytokines, and NF-κB activities in LPS-treated microglial cells.14 In addition, 1-dehydro-10-gingerdione inhibits TLR-4-mediated signaling cascades and cytokine expression via blockade of LPS binding to myeloid differentiation protein 2, a co-receptor of TLR-4 in macrophages.29 In this study, we found that ginger and zingerone inhibited LPS-induced NF-κB activities in various organs in mice. Ginger and zingerone also suppressed the nuclear translocation of NF-κB subunit p65, the production of IL-1β, and the infiltration of granulocytes in small intestine. Overall, this study suggested that ginger and zingerone shared a common anti-inflammatory mechanism by inhibiting NF-κB activities and proinflammatory cytokine production in LPS-induced systemic inflammation. Moreover, ginger exhibits a non-steroid anti-inflammatory activity via inhibiting the activities of COX-2 and iNOS in other studies, probably explaining why ginger displayed a broad-spectrum anti-inflammatory activity in various organs in this study.
Detailed anti-inflammatory effects of ginger and zingerone in small intestine were evaluated here. It is interesting to find that LPS increased NF-κB-driven luminescence in the entire small intestine, especially from the middle portion of jejunum to the end of ileum. It is 270 271 272 273 274 275 276 277 278 279 280 281 282 283 284 285 286 287 288
known that LPS activates macrophages, neutrophils, and dendritic cells via binding to TLR-4 and activating downstream NF-κB activity. The activated immune cells then initiate the inflammatory response and present antigens to lymphocytes in lymph nodes.30 In comparison with duodenum, jejunum and ileum have abundant Peyer's patches, organized lymphoid nodules, in mice. Thus, we speculated that LPS induced maximal NF-κB-driven luminescence in jejunum and ileum might result from the abundance of Peyer's patches. We also found that ginger and zingerone suppressed LPS-induced NF-κB-dependent bioluminescent signals in the entire small intestine, especially in the region between mid-jejunum and mid-ileum. Zingerone possesses the vanillyl moiety, which is considered important for the activation of vanilloid receptor 1 (VR1) expressed in nociceptive sensory neurons.31 Recent study shows that activated VR1 protects against LPS-mediated renal injury possibly via reducing renal inflammation responses.32 In addition, sensory VR1 has been found to modulate cytokine response to LPS and thereby induce the subsequent anti-inflammatory effect in the gut mucosa.33 VR1 nerve fibers are observed within enteric ganglia of jejunum and ileum,34 probably explaining why zingerone displayed more activities in jejunum and ileum than in duodenum.
In conclusion, we comprehensively evaluated the anti-inflammatory effects of ginger and zingerone on LPS-induced systemic inflammation via NF-κB bioluminescent imaging. Our data showed for the first time that ginger and zingerone suppressed NF-κB-drive 289 290 291 292 293 294 295 296 297 298 299 300 301 302 303 304 305 306 307
luminescence in most organs. In addition, our findings suggested that ginger and zingerone were likely to be broad-spectrum anti-inflammatory agents in most organs that suppressed the activation of NF-κB, the production of IL-1β, and the infiltration of inflammatory cells.
ABBREVIATIONS USED
COX-2, cyclooxygenase-2; ELISA, enzyme-linked immunosorbent assay; IHC, immunohistochemical; iNOS, inducible nitric oxide synthase; IL-1β, interleukin-1β; LPS, lipopolysaccharide; MTT, 3-(4,5-dimethylthiazol-2-yl-2,5-diohenyl tetrazolium bromide; NO, nitric oxide; NF-κB, nuclear factor-κB; RLU, relative luciferase unit; sr, steradian; TLR-4, toll-like receptor 4; TNF-α, tumor necrosis factor-α; VR1, vanilloid receptor 1
SUPPORTING INFORMATION
HPLC profile of ginger extract (Supplementary Figure S1).
FUNDING SOURCES
This work was supported by grants from Ministry of Science and Technology (NSC101-2320-B-039-034-MY3, NSC102-2632-B-039-001-MY3, and MOST 104-2815-C-039-012-B), China Medical University (CMU102-NSC-04 and CMU103-SR-44), and CMU under the Aim for Top University Plan of the Ministry of Education, Taiwan.
308 309 310 311 312 313 314 315 316 317 318 319 320 321 322 323 324 325 326 327
NOTES
The authors declare no competing financial interest. 328
329 330
REFERENCES
(1) Ryan, J. L.; Heckler, C. E.; Roscoe, J. A.; Dakhil, S. R.; Kirshner, J.; Flynn, P. J.; Hickok, J. T.; Morrow, G. R. Ginger (Zingiber officinale) reduces acute chemotherapy-induced nausea: a URCC CCOP study of 576 patients. Support. Care Cancer 2012, 20, 1479-1489.
(2) Dabaghzadeh, F.; Khalili, H.; Dashti-Khavidaki, S.; Abbasian, L.; Moeinifard, A. Ginger for prevention of antiretroviral-induced nausea and vomiting: a randomized clinical trial. Expert Opin. Drug Saf. 2014, 13, 859-866.
(3) Chopra, A.; Saluja, M.; Tillu, G.; Sarmukkaddam, S.; Venugopalan, A.; Narsimulu, G.; Handa, R.; Sumantran, V.; Raut, A.; Bichile, L.; Joshi, K.; Patwardhan, B. Ayurvedic medicine offers a good alternative to glucosamine and celecoxib in the treatment of symptomatic knee osteoarthritis: a randomized, double-blind, controlled equivalence drug trial. Rheumatology 2013, 52, 1408-1417.
(4) Bartels, E. M.; Folmer, V. N.; Bliddal, H.; Altman, R. D.; Juhl, C.; Tarp, S.; Zhang, W.; Christensen, R. Efficacy and safety of ginger in osteoarthritis patients: a meta-analysis of randomized placebo-controlled trials. Osteoarthr. Cartil. 2015, 23, 13-21.
(5) Black, C. D.; Herring, M. P.; Hurley, D. J.; O'Connor, P. J. Ginger (Zingiber officinale) reduces muscle pain caused by eccentric exercise. J. Pain 2010, 11, 894-903.
331 332 333 334 335 336 337 338 339 340 341 342 343 344 345 346 347 348
(6) Rahnama, P.; Montazeri, A.; Huseini, H. F.; Kianbakht, S.; Naseri, M. Effect of Zingiber officinale R. rhizomes (ginger) on pain relief in primary dysmenorrhea: a placebo randomized trial. BMC Complement. Altern. Med. 2012, 12, 92.
(7) Mozaffari-Khosravi, H.; Talaei, B.; Jalali, B. A.; Najarzadeh, A.; Mozayan, M. R. The effect of ginger powder supplementation on insulin resistance and glycemic indices in patients with type 2 diabetes: a randomized, double-blind, placebo-controlled trial. Complement. Ther. Med. 2014, 22, 9-16.
(8) Jung, H. W.; Yoon. C. H.; Park, K. M.; Han, H. S.; Park, Y. K. Hexane fraction of Zingiberis Rhizoma Crudus extract inhibits the production of nitric oxide and proinflammatory cytokines in LPS-stimulated BV2 microglial cells via the NF-kappaB pathway. Food Chem. Toxicol. 2009, 47, 1190-1197.
(9) Podlogar, J. A.; Verspohl, E. J. Antiinflammatory effects of ginger and some of its components in human bronchial epithelial (BEAS-2B) cells. Phytother. Res. 2012, 26, 333-336.
(10) Li, F.; Nitteranon, V.; Tang, X.; Liang, J.; Zhang, G.; Parkin, K. L.; Hu, Q. In vitro antioxidant and anti-inflammatory activities of 1-dehydro-[6]-gingerdione, shogaol, 6-dehydroshogaol and hexahydrocurcumin. Food Chem. 2012, 135, 332-327.
(11) Huang, S. H.; Lee, C. H.; Wang, H. M.; Chang, Y. W.; Lin, C. Y.; Chen, C. Y.; Chen, Y. H. 6-Dehydrogingerdione restrains lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages. J. Agric. Food Chem. 2014, 62, 9171-9179.
349 350 351 352 353 354 355 356 357 358 359 360 361 362 363 364 365 366 367 368
(12) Rahmani, A. H.; Shabrmi, F. M.; Aly, S. M. Active ingredients of ginger as potential candidates in the prevention and treatment of diseases via modulation of biological activities. Int. J. Physiol. Pathophysiol. Pharmacol. 2014, 6, 125-136.
(13) Ha, S. K.; Moon, E.; Ju, M. S.; Kim, D. H.; Ryu, J. H.; Oh, M. S.; Kim, S. Y. 6-Shogaol, a ginger product, modulates neuroinflammation: a new approach to neuroprotection. Neuropharmacology 2012, 63, 211-223.
(14) Moon, M.; Kim, H. G.; Choi, J. G.; Oh, H.; Lee, P. K.; Ha, S. K.; Kim, S. Y.; Park, Y.; Huh, Y.; Oh, M. S. 6-Shogaol, an active constituent of ginger, attenuates neuroinflammation and cognitive deficits in animal models of dementia. Biochem. Biophys. Res. Commun. 2014, 449, 8-13.
(15) Xie, X.; Sun, S.; Zhong, W.; Soromou, L. W.; Zhou, X.; Wei, M.; Ren, Y.; Ding, Y. Zingerone attenuates lipopolysaccharide-induced acute lung injury in mice. Int. Immunopharmacol. 2014, 19, 103-109.
(16) Hsiang, C. Y.; Lo, H. Y.; Huang, H. C.; Li, C. C.; Wu, S. L.; Ho, T. Y. Ginger extract and zingerone ameliorated trinitrobenzene sulphonic acid-induced colitis in mice via modulation of nuclear factor-κB activity and interleukin-1β signalling pathway. Food Chem. 2013, 136, 170-177.
(17) Ho, T. Y.; Chen, Y. S.; Hsiang, C. Y. Noninvasive nuclear factor-κB bioluminescence imaging for the assessment of host-biomaterial interaction in transgenic mice. Biomaterials 2007, 28, 4370-4377. 369 370 371 372 373 374 375 376 377 378 379 380 381 382 383 384 385 386 387 388
(18) Chang, C. T.; Lin, H.; Ho, T. Y.; Li, C. C.; Lo, H. Y.; Wu, S. L.; Huang, Y. F.; Liang, J. A.; Hsiang, C. Y. Comprehensive assessment of host responses to ionizing radiation by nuclear factor-κB bioluminescence imaging-guided transcriptomic analysis. PLoS ONE 2011, 6, e23682.
(19) Li, C. C.; Hsiang, C. Y.; Lo, H. Y.; Pai, F. T.; Wu, S. L.; Ho, T. Y. Genipin inhibits lipopolysaccharide-induced acute systemic inflammation in mice as evidenced by nuclear factor-κB bioluminescent imaging-guided transcriptomic analysis. Food Chem. Toxicol. 2012, 50, 2978-2986.
(20) Li, C. C.; Hsiang, C. Y.; Wu, S. L.; Ho, T. Y. 2012. Identification of novel mechanisms of silymarin on the carbon tetrachloride-induced liver fibrosis in mice by nuclear factor-κB bioluminescent imaging-guided transcriptomic analysis. Food Chem. Toxicol. 2012, 50, 1568-1575.
(21) Badr, C. E. Bioluminescence imaging: basics and practical limitations. Methods Mol. Biol. 2014, 1098, 1-18.
(22) Choi, Y. Y.; Kim, M. H.; Hong, J.; Kim, S. H.; Yang, W. M. Dried ginger (Zingiber officinalis) inhibits inflammation in a lipopolysaccharide-induced mouse model. Evid. Based Complement. Alternat. Med. 2013, 2013, 914563.
(23) Sharma, J. N.; Srivastava, K. C.; Gan, E. K. Suppressive effects of eugenol and ginger oil on arthritic rats. Pharmacology 1994, 49, 314-318.
389 390 391 392 393 394 395 396 397 398 399 400 401 402 403 404 405 406 407
(24) El-Akabawy, G.; El-Kholy, W. Neuroprotective effect of ginger in the brain of streptozotocin-induced diabetic rats. Ann. Anat. 2014, 196, 119-128.
(25) Yu, Y.; Zick, S.; Li, X.; Zou, P.; Wright, B.; Sun, D. Examination of the pharmacokinetics of active ingredients of ginger in humans. AAPS J. 2011, 13, 417-426.
(26) Monge, P.; Scheline, R.; Solheim, E. The metabolism of zingerone, a pungent principle of ginger. Xenobiotica 1976, 6, 411-423.
(27) Yu, Y. S.; Hsu, C. L.; Yen, G. C. Anti-inflammatory effects of the roots of Alpinia pricei Hayata and its phenolic compounds. J. Agric. Food Chem. 2009, 57: 7673-7680.
(28) Villalvilla, A.; da Silva, J. A.; Largo, R.; Gualillo, O.; Vieira, P. C.; Herrero-Beaumont, G.; Gómez, R. 6-Shogaol inhibits chondrocytes' innate immune responses and cathepsin-K activity. Mol. Nutr. Food Res. 2014, 58, 256-266.
(29) Park, S. H.; Kyeong, M. S.; Hwang, Y.; Ryu, S. Y.; Han, S. B.; Kim, Y. Inhibition of LPS binding to MD-2 co-receptor for suppressing TLR4-mediated expression of inflammatory cytokine by 1-dehydro-10-gingerdione from dietary ginger. Biochem. Biophys. Res. Commun. 2012, 419, 735-740.
(30) Morris, M. C.; Gilliam, E. A.; Li, L. Innate immune programing by endotoxin and its pathological consequences. Front Immunol. 2015, 5, 680.
(31) Dedov, V. N.; Tran, V. H.; Duke, C. C.; Connor, M.; Christie, M. J.; Mandadi, S.; Roufogalis, B. D. Gingerols: a novel class of vanilloid receptor (VR1) agonists. Br. J. Pharmacol. 2002, 137, 793-798. 408 409 410 411 412 413 414 415 416 417 418 419 420 421 422 423 424 425 426 427
(32) Wang, Y.; Wang, D. H. Deletion of the transient receptor potential vanilloid type 1 channel exacerbates renal inflammation induced by lipopolysaccharide in mice. FASEB J.
2013, 27, 721.10
(33) Assas, B. M.; Miyan, J. A.; Pennock, J. L. Cross-talk between neural and immune receptors provides a potential mechanism of homeostatic regulation in the gut mucosa. Mucosal Immunol. 2014, 7, 1283-1289.
(34) Vinuesa, A. G.; Sancho, R.; García-Limones, C.; Behrens, A.; ten Dijke, P.; Calzado, M. A.; Muñoz, E. Vanilloid receptor-1 regulates neurogenic inflammation in colon and protects mice from colon cancer. Cancer Res. 2012, 72, 1705-1716.
428 429 430 431 432 433 434 435 436
FIGURE CAPTIONS
Figure 1. Ginger and zingerone. (A) Morphology of whole parts and cross sections of dried ginger. (B) Chemical structure of zingerone.
Figure 2. Effects of ginger and zingerone on LPS-induced NF-κB activities in cells. HepG2/NF-κB cells were treated with 100 ng/ml LPS and/or various amounts of ginger extract and zingerone. MG-132 (5 µM) was used as a positive control. Twenty-four hours later, κB activity was measured by luciferase assay. Results are expressed as relative NF-κB activity, which is presented as the comparison with RLU relative to solvent-treated cells. Values are mean ± standard error (n=6). ###p < 0.001, compared with mock. **p < 0.01, ***p < 0.001, compared with LPS.
Figure 3. NF-κB-driven luminescence in living mice. (A) In vivo image. Transgenic mice were administered with 1 mg/kg LPS and then treated with 100 mg/kg ginger or zingerone. Four hours later, mice were injected intraperitoneally with D-luciferin and imaged for 1 min. The color overlay on the image represents the photon/sec emitted from mice, as indicated by the color scale. Photos are representative images (n=5/group). (B) Quantification of photon emission from the whole body. Values are mean ± standard error. ###p < 0.001, compared with mock. *p < 0.05, ***p < 0.001, compared with LPS.
437 438 439 440 441 442 443 444 445 446 447 448 449 450 451 452 453 454 455 456
Figure 4. Effects of ginger and zingerone on LPS-induced IL-1β and TNF-α production in sera. Transgenic mice were challenged with 1 mg/kg LPS and then given with 100 mg/kg ginger or zingerone. Four hours later, mice were sacrificed and the amount of IL-1β (A) and TNF-α (B) in sera was quantified by ELISA. Values are mean ± standard error (n=5/group). ##p < 0.01, compared with mock. *p < 0.05, **p < 0.01, ***p < 0.001, compared with LPS.
Figure 5. NF-κB-dependent luminescence in individual organs. (A) Ex vivo imaging. Transgenic mice were administered with 1 mg/kg LPS and then treated with 100 mg/kg ginger or zingerone. Four hours later, mice were injected intraperitoneally with D-luciferin. Five minutes later, mice were sacrificed, and organs were excised rapidly and subjected to image (n=5/group). (B) Quantification of photon emission from organs. Values are mean ± standard error. ##p < 0.01, ###p < 0.001, compared with mock. *p < 0.05, **p < 0.01, ***p < 0.001, compared with LPS.
Figure 6. Anti-inflammatory effects of ginger and zingerone in various segments of small intestine. Transgenic mice were challenged with 1 mg/kg LPS and then given with 100 mg/kg ginger or zingerone. Four hours later, mice were injected intraperitoneally with D-luciferin. Five minutes later, mice were sacrificed, and organs were excised rapidly and subjected to image. Small intestine was divided equally into 35 segments and the photon emission from 457 458 459 460 461 462 463 464 465 466 467 468 469 470 471 472 473 474 475
each segment was quantitated. Results are expressed as inhibition (%). Heatmap of small intestine is shown on the bottom. The color overlay on the heatmap represents the photon/sec emitted from each segment of small intestine, as indicated by the color scale.
Figure 7. IHC staining analysis of p65, IL-1β, and CD11b in jejunum. Transgenic mice were challenged with 1 mg/kg LPS and then given with 100 mg/kg ginger or zingerone. Four hours later, mice were sacrificed. Section of mid-jejunum was stained with antibodies against p65, IL-1β, and CD11b (400× magnification). Photos are representative images (n=5/group) 476 477 478 479 480 481 482 483
Figure 1 (A) (B)
484 485 486 487 488 489 490 491 492 493
Figure 2 Mock LPS MG-132 0.5 1 2.5 5 10 50 100 0 1 2 3 4 5 Ginger Zingerone Concentration (μg/ml) R el at iv e N F -κ B a ct iv it y 494 495 496 497 498 499 500 501
Figure 3
(A)
Mock LPS LPS/Ginger LPS/Zingerone
(B)
Mock LPS LPS/Ginger LPS/Zingerone
0 50 100 150 200 250 300 350 400 T ot al fl ux (× 10 6 ph ot on /s ec ) photon/sec 502 503 504 505 506 507 508 509 510
Figure 4
(A)
Mock LPS LPS/Ginger LPS/Zingerone
0 100 200 300 400 500 600 700 IL -1 β co nc et ra ti on (p g/ m l) (B)
Mock LPS LPS/Ginger LPS/Zingerone
0 200 400 600 800 1,000 1,200 1,400 T N F -α c on ce tr at io n (p g/ m l) 511 512 513 514 515 516 517 518 519
Brain Heart Lung Liver Spleen Stomach Kidney Intestine Mock LPS LPS/Ginger LPS/Zingerone Figure 5
(A)
(B)
Brain Heart Lung Liver Spleen Stomach Kidney Intestine 0 2000 4000 6000 8000 10000 12000 14000 16000 18000 Mock LPS LPS/Ginger LPS/Zingerone L um in es ce nt in te ns it y (× 10 3 ph ot on /s ec /c m 2/ sr )
m
oc
k
br
ai
n
hr
ea
t
lu
ng
liv
er
spl
een
sto
mac
h
kid
ney
intest
ines
L
PS
Gin
ger
Zinge
rone
520 521 522 523 524 525 526Figure 6 R O I 1 R O I 2 R O I 3 R O I 4 R O I 5 R O I 6 R O I 7 R O I 8 R O I 9 R O I 1 0 R O I 1 1 R O I 1 2 R O I 1 3 R O I 1 4 R O I 1 5 R O I 1 6 R O I 1 7 R O I 1 8 R O I 1 9 R O I 2 0 R O I 2 1 R O I 2 2 R O I 2 3 R O I 2 4 R O I 2 5 R O I 2 6 R O I 2 7 R O I 2 8 R O I 2 9 R O I 3 0 R O I 3 1 R O I 3 2 R O I 3 3 R O I 3 4 R O I 3 5 0 20 40 60 80 Ginger Zingerone In hi bi ti on (o /o ) Mock LPS LPS/Ginger LPS/Zingerone photon/sec 527 528 529 530 531 532
533 534
Figure 7
Mock LPS LPS/Ginger LPS/Zingerone
p65 IL-1β CD11b 535 536 537 538 539