台大農藝系 遺傳學 601 20000 Chapter 7 slide 1
CHAPTER 8
Recombinant DNA
Technology
Peter J. Russell
edited by Yue-Wen Wang Ph. D. Dept. of Agronomy, NTU
DNA Cloning
1. The goal of molecular cloning is large amounts of pure
DNA that can be further manipulated and studied.
2. Summary of the procedure:
a. Isolate DNA from the organism.
b. Use restriction enzymes to cut the DNA, and ligate fragments
into a cloning vector.
c. Transform recombinant DNA into a host, which will replicate
the DNA (molecular cloning) and pass copies to all progeny.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 3
Restriction Enzymes
1. Restriction endonucleases (restriction enzymes) each recognize a specific DNA sequence (restriction site), and break a phosphodiester linkage between a 3’ carbon and phosphate within that sequence.
2. Restriction enzymes are used to create DNA fragments for cloning, and to analyze positions of restriction sites in cloned or genomic DNA.
3. Restriction enzymes are a bacterial defense against viral DNA.
Restriction sites in the bacterial chromosome are methylated, and thus protected.
4. A restriction enzyme has a three-letter name derived from the genus and species of the organism from which it was isolated; it is underlined or italicized. Roman numerals and sometimes letters designating a
particular bacterial strain may follow.
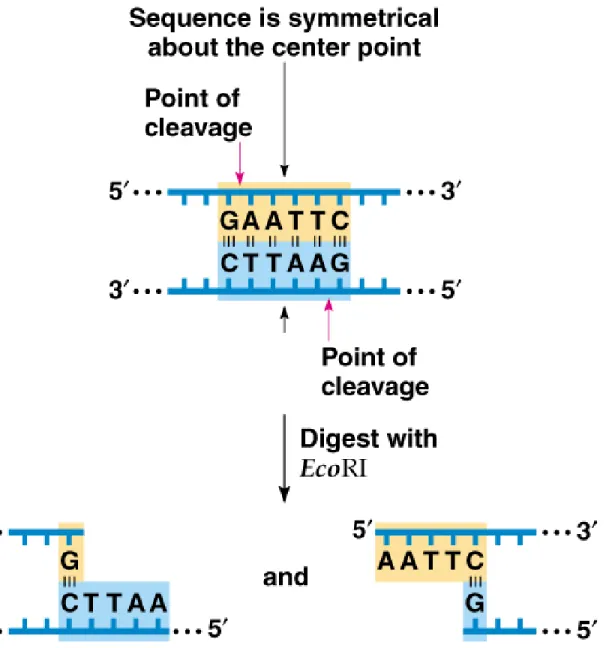
5. Many restriction sites are palindromes of 4-, 6- or 8-base pairs, but others are not completely symmetrical (Figure 8.1 and Table 8.1).
台大農藝系 遺傳學 601 20000 Chapter 7 slide 4
Fig. 8.1 Restriction site in DNA, showing symmetry of the sequence around the
6. All copies of a chromosome will contain the same restriction sites, and will be cut into identical fragments.
7. Based on probability, a specific short DNA sequence occurs more frequently than a longer one.
a. In a 50% G-C organism with random distribution of bases, the probability of a specific base at a given position is 1⁄4.
b. Therefore, the frequency of a particular restriction site is (1⁄4)n,
where n is the number of base pairs in the recognition sequence. 8. One major class of restriction enzymes recognizes and cuts DNA
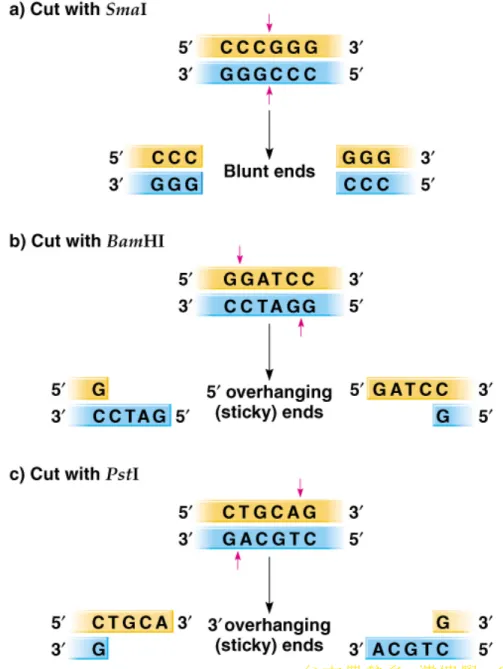
at specific sequences. Two types of DNA ends can be generated (Figure 8.2).
a. Some restriction enzymes produce blunt ends, where both DNA strands are cut between the same base pairs.
b. Others create sticky (staggered) ends.
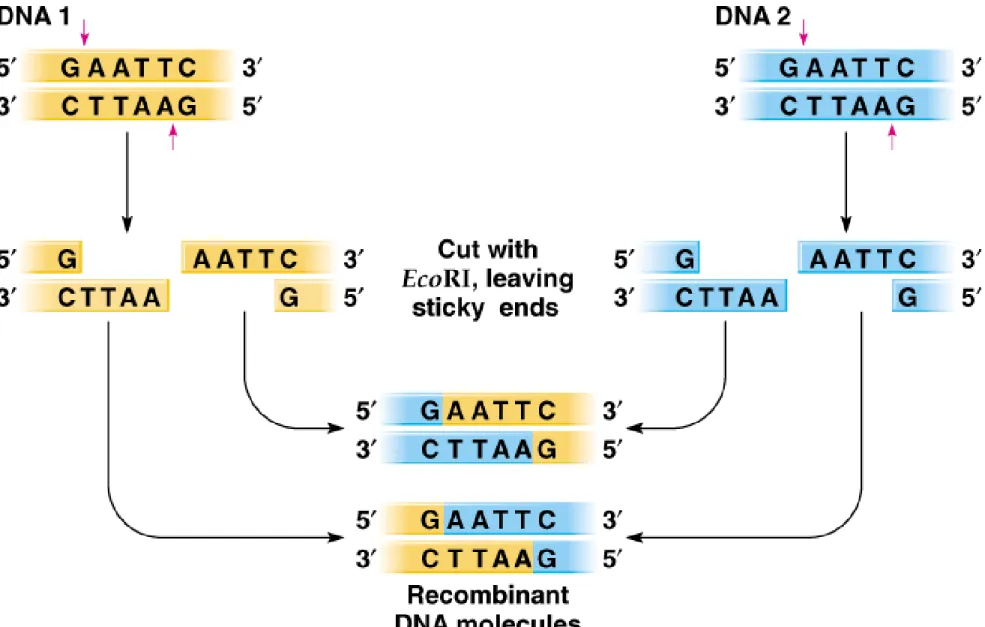
9. Sticky ends are useful in cloning, because complementary sequences hydrogen bond (anneal) and are held together so that DNA ligase can covalently link them (Figure 8.3).
台大農藝系 遺傳學 601 20000 Chapter 7 slide 7
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 8
台大農藝系 遺傳學 601 20000 Chapter 7 slide 9
Cloning Vectors and DNA Cloning
Animation: DNA Cloning in a Plasmid Vector
1. Several types of cloning vectors have been constructed, each with different molecular properties and cloning capacity.
2. Plasmid cloning vectors are derived from natural plasmids, circles of dsDNA that include origin sequences (ori) needed for replication in bacterial cells. An E. coli plasmid vector, for example, must contain these features:
a. An ori sequence for replication.
b. A selectable marker, such as antibiotic resistance.
c. Unique restriction sites, so that a particular restriction enzyme cuts only once in the plasmid. A fragment of insert DNA cut with the same enzyme is commonly inserted into the unique restriction site.
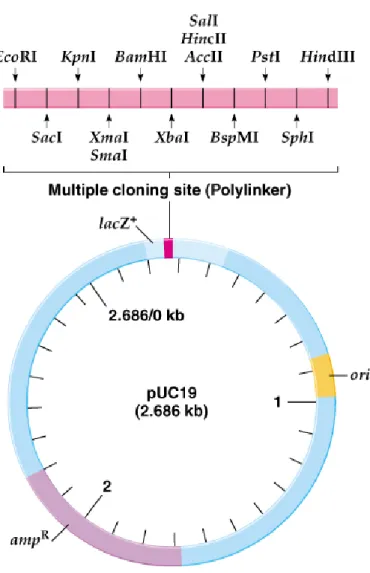
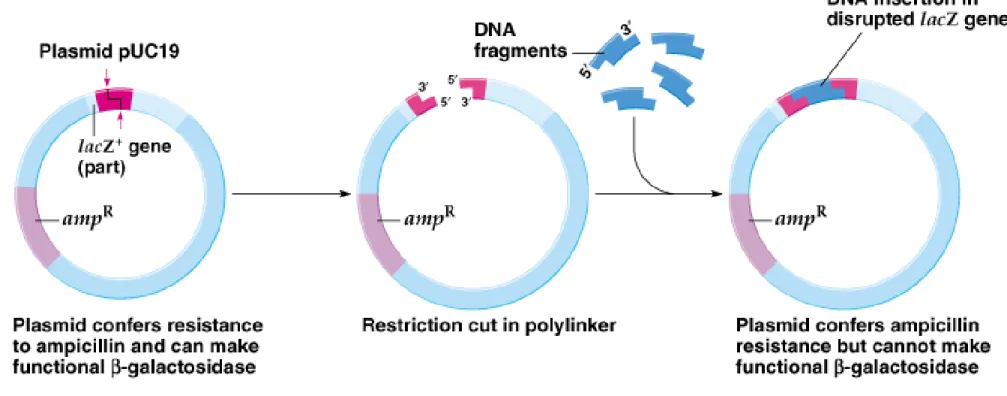
3. An example of a typical E. coli cloning vector is pUC19 (2,686-bp). The pUC19 plasmid features:
a. High copy number in E. coli, with nearly a hundred copies per cell, provides a good yield of cloned DNA. b. Its selectable marker is ampR.
c. It has a cluster of unique restriction sites, called the polylinker (multiple cloning site).
d. The polylinker is part of the lacZ (β-galactosidase) gene. The pUC19 plasmid will complement a lacZ- E. coli, allowing it to become lacZ+. When DNA is cloned into the polylinker, lacZ is disrupted, preventing
complementation from occurring.
e. X-gal, a chromogenic analog of lactose, turns blue whenβ-galactosidase is present, and remains white in its absence, so blue-white screening can indicate which colonies contain recombinant plasmids (Figure 8.4).
台大農藝系 遺傳學 601 20000 Chapter 7 slide 10
台大農藝系 遺傳學 601 20000 Chapter 7 slide 11
4. DNA can be inserted into a cloning vector by restriction digestion and then ligation (Figure 8.5).
a. Cut pUC19 (the vector plasmid) with a restriction enzyme that has a unique site in the polylinker.
b. Cut the DNA to be cloned (insert DNA) with the same enzyme.
c. Mix insert DNA with pUC19 DNA and allow random joining of fragments to occur.
d. Resulting plasmids are transformed into E. coli either through chemical treatment of the cells or by electroporation. The cells are grown on media plates containing ampicillin and X-gal. e. Ampicillin-resistant colonies result from pUC19 sequences. Blue colonies contain only the vector with its ends rejoined, while white colonies often contain pUC19 with its lacZ gene inactivated by insert DNA.
f. If the 5’-phosphates of vector DNA are removed by alkaline phosphatase, DNA ligase will not rejoin their ends, and fewer blue colonies will result.
5. If one enzyme is used in cloning, two orientations are possible for the insert. A two-enzyme strategy results in only one orientation.
6. Many plasmid cloning vectors are available, with features including different arrays of unique restriction sites in the polylinker, and phage promoters (e.g., T7, T3, SP6) that can be used to control transcription of the cloned DNA.
7. Plasmid cloning vectors are available for many prokaryotic and eukaryotic organisms. In some cases the plasmids are unable to replicate, but are maintained because they
integrate into the genome.
8. Size of the insert DNA is limited in plasmid cloning vectors, and plasmids carrying more than 5–10 kb are often unstable.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 12
台大農藝系 遺傳學 601 20000 Chapter 7 slide 13
Shuttle Vectors
1. A cloning vector capable of replicating in two or
more types of organism (e.g., E. coli and yeast) is
called a shuttle vector. Shuttle vectors may
replicate autonomously in both hosts, or integrate
into the host genome
Expression Vectors
1. Expression vectors have sequences to allow transcription
and translation of the cloned gene(s). Pharmaceuticals
produced by biotechnology are an example.
2. Plasmid cloning vectors are modified to include:
a. Promoter and transcription terminator if needed, suitable to the
host organism.
b. Any modifications needed to cross prokaryotic/eukaryotic
boundary (e.g., Shine-Delgarno sequence is added for translation
of eukaryotic sequences in E. coli).
台大農藝系 遺傳學 601 20000 Chapter 7 slide 15
Artificial Chromosomes
1. Cloning vectors that can accommodate very large
pieces of DNA produce molecules resembling
small chromosomes. Two examples are YACs
and BACs.
Yeast Artificial Chromosomes (YACs)
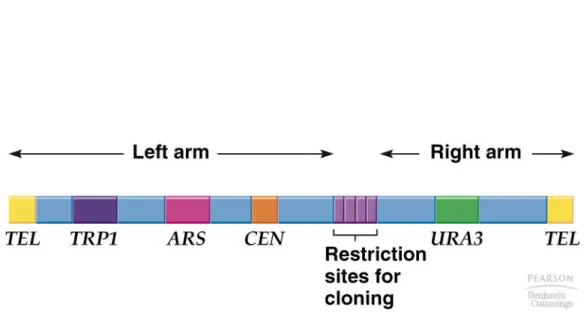
1. YAC vectors function as artificial chromosomes in yeast. Their features include (Figure 8.6)
a. Linear structure with a yeast telomere (TEL) at each end. b. A yeast centromere sequence (CEN).
c. A marker gene on each arm that is selectable in yeast. (e.g. TRP1 and URA3) d. A yeast origin of replication known as autonomous replicating sequence
(ARS).
e. Unique restriction sites for inserting foreign DNA.
2. Several hundred kb of insert DNA can be cloned in a YAC. However, frequent RNA rearrangements in the host make YACs unsuitable for genome sequencing. 3. YAC clones are made by:
a. Propagating the DNA in E. coli as a circular plasmid with telomeres end-to-end. b. Cut with restriction enzymes in multiple cloning site and another between the two
TELs, generating two arms.
c. Ligating long insert DNA fragment with the two arms. d. Transforming into yeast.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 17
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Bacterial Artificial Chromosomes (BACs)
1. BACs are used for cloning fragments up to about 200 kb
in E .coli. BAC vectors contain:
a. the ori of an E. coli plasmid called the F factor.
b. A multiple cloning sites.
c. A selectable marker.
d. Other features, “bells and whistles.”
2. BAC can be handles like regular bacterial plasmids, but
the F factor ori keeps copy number at one BAC molecule
per cell.
3. BACs do not undergo rearrangements in the host.
4. BACs are commonly used to study gene regulation in
vertebrates.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 19
Recombinant DNA Libraries
1. It is sometimes useful to have a genomic library, a
collection of clones containing at least one copy
of every DNA sequence in a genome. Genomic
libraries are available for many organisms.
2. Chromosome libraries are collections of cloned
fragments of individual chromosomes.
3. Complementary DNA (cDNA) libraries are
cloned collections of DNA copied from a cell’s
mRNA.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 21
Genomic Libraries
1. A genomic library is used to isolate and study a cloned DNA
containing a sequence of interest. Genomic libraries of eukaryotic DNA are constructed by digesting genomic DNA with a restriction enzyme, and ligating into a vector that can accommodate the genome in a manageable number of clones.
2. There are three general ways to produce genomic libraries:
a. Complete digestion with restriction enzyme, cleave at all relevant restriction sites. This has drawback:
1) Genes containning one or more sites for the restriction enzyme will be cloned into two or more pieces.
2) To screen the entire genome, a very large number of clones
would have to be examined, because insert DNA size is relative small.
b. Longer DNA fragments can be generated with mechanical sheering (e.g passage through a syringe needle) rather than restriction enzyme cutting. A disadvantage is the absence of uniform ends, require
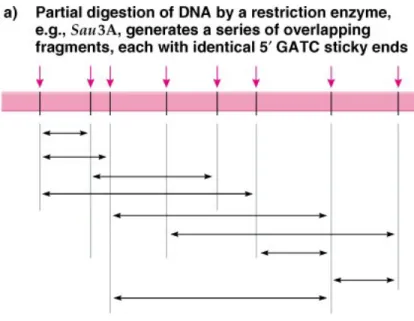
c. Partial digestion with a restriction enzyme is controlled so that it cuts only some of the available sites. Ideally, this results in cloning a
population of overlapping fragments representing the entire genome (Figure 8.7)
1) Partially digested DNA molecules in a certain size range are selected by density gradient centrifugation or agarose gel electrophoresis.
2) DNA fragments with sticky ends from restriction digestion can be cloned directly. Sometimes two enzymes with compatible sticky ends are used (e.g. BamHI and Sau3A), creating a hybrid recognition site.
3) Genomic sequences are not equally represented in the libraries, because:
a) Regions of DNA with relevant restriction sites very close together or very far apart are removed at the size selection.
b) Some regions of eukaryotic DNA prevent vector replication in E. coli, and so are eliminated from the library.
3. The number of clones needed for a complete library can be calculated based on the size of the genome and the average size of DNA
inserted into the vector. In practice, a library should contain many times more than the calculated minimum number of clones.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 23
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 8.7 Use of partial digestion with a restriction enzyme to produce DNA fragments
Chromosome libraries
1.
Screening the genomic library of an organism
with a large genome is laborious. Screening time
can be reduced if a gene has been localized to a
chromosome, by examining a library made from
only that chromosome. Human, for example,
have 24 different chromosome libraries (22
autosomes, X and Y).
2.
Separating chromosome so they may be
individually cloned is accomplished with
techniques such as flow cytometry.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 25
cDNA Libraries
1. A cell’s mRNA molecules can be copied to make complementary DNA strands (cDNA) and the cDNA can then be cloned, creating a library representing only the genes being expressed in the cells at that time. 2. The cDNA derives only from mature mRNA. Introns are not present. 3. The poly(A) tail at the 3’ end of the mRNA is useful for:
a. Isolating mRNA from cell lysates by passage over an oligo(dT) column.
i. The mRNA’s poly(A) tail sticks to the poly(T) attached to the column substrate.
ii. Other molecules pass through the column, but mRNAs are retained.
iii. mRNAs are eluted with decreasing ionic strength buffer, resulting in significant purification.
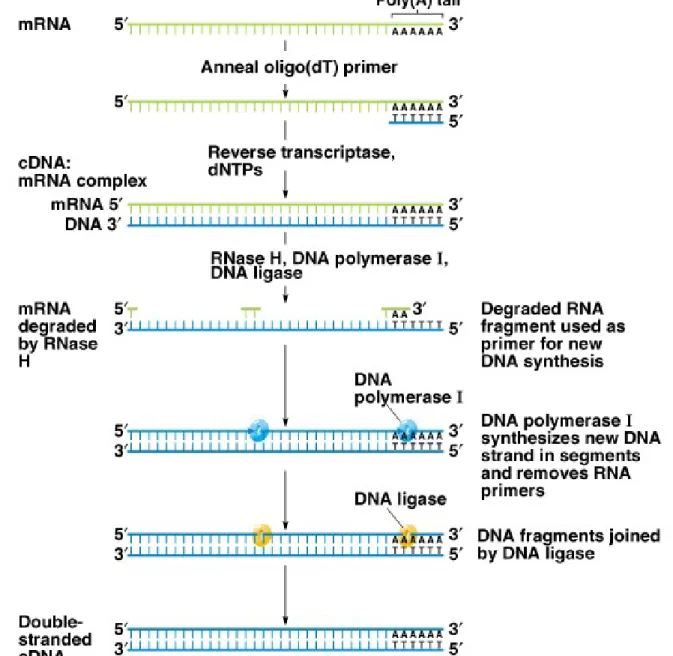
4. Synthesis of cDNA involves these steps (Figure 8.8):
a. A short oligo(dT) primer is used. It anneals to the mRNA’s
poly(A) tail, allowing reverse transcriptase to synthesize cDNA.
This creates a DNA-mRNA hybrid.
b. RNase H degrades the mRNA strand, creating small RNA
fragments that serve as primers.
c. DNA polymerase I makes new DNA fragments, and DNA ligase
connects the new DNA fragments to make a complete chain.
d. The resulting cDNA is a double-stranded copy of the starting
mRNA.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 27
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 8.8 The synthesis of complementary DNA (cDNA) from a polyadenylated mRNA
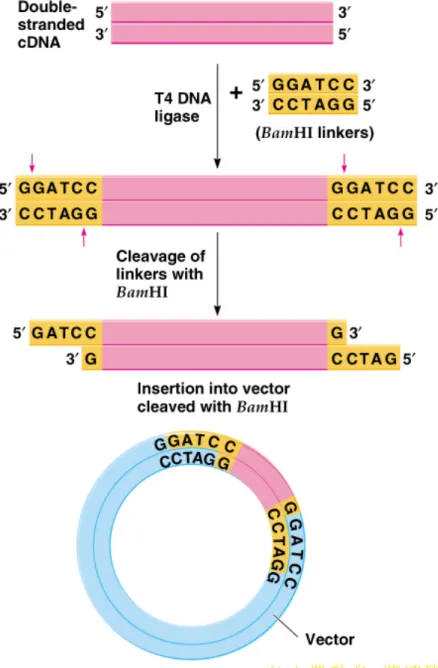
5. A method for cloning cDNA involves (Figure 8.9):
a. Introducing restriction site linkers to the ends of the cDNA by
blunt end ligation.
b. Digestion with the cognate restriction enzyme to create sticky
ends.
c. Mixing cDNA with vector DNA cut with the same restriction
enzyme in the presence of DNA ligase.
d. Transforming into an E. coli host for cloning.
6. If the cDNA has sites for the same restriction enzyme
used in the polylinker, the cDNA will be cloned in pieces.
The problem is avoided by using polylinkers engineered
with appropriate ssDNA overhangs (sticky ends) so that
restriction digestion is unnecessary.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 29
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Finding a Specific Clone in a Library
1. A number of techniques have been developed for
identifying the clone of interest in a cDNA
台大農藝系 遺傳學 601 20000 Chapter 7 slide 31
Screening a cDNA library
1. One way to select a cDNA clone from the library is to detect a protein product it is producing (Figure 8.10).
2. For protein to be produced, an expression vector is needed, in which the cloned cDNA is inserted between a promoter and a transcription
terminator.
3. Labeled antibodies are used to detect the specific protein in a host colony. An array of colonies is transferred to a membrane fiter, cells are lysed and their proteins bind to the filter, which is incubated with the relevant antibody.
4. Radioactively labeled antibody bound to colonies is detected by an
autoradiogram, in which the dry fiter is placed on X ray film in the dark for a number of hours. Colonies with antibody bound will be visible as dark spots on the film.
5. Once identified, the cDNA can be used to:
a. Analyze the genome of the same or another organism for homologous sequences.
b. Isolate the nuclear gene for the mRNA from a genomic library. c. Quantify mRNA synthesized from the gene.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 32
Fig. 8.10 Screening for specific cDNA plasmids in a cDNA library by using an antibody
台大農藝系 遺傳學 601 20000 Chapter 7 slide 33
Screening a Genomic Library
1. Finding a specific clone in a plasmid or cosmid genomic library is similar to screening a cDNA library (Figure 8.11).
a. E. coli cells are transformed with the genomic library, and plated on
selective medium.
b. Colonies produced are replica plated onto a membrane filter on a plate of selective medium, and cells grow on the filter.
c. Cells are lysed, DNA is denatured and bound tightly to the filter.
d. Filter is incubated with the labeled single-stranded cDNA probe, which forms hybrids with complementary DNA molecules bound to the filter. e. Filter is washed and the label is detected by autoradiography for a
radioactive probe, or chemiluminescent or colorimetric assay for nonradioactive labeling.
f. Clones detected by the probe are then further characterized.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 34
Fig. 8.11 Using DNA probes to screen plasmid or cosmid genomic libraries for specific
台大農藝系 遺傳學 601 20000 Chapter 7 slide 35
Identifying Genes in Libraries by
Complementation of Mutations
1. Well-defined mutants may be used to clone genes by complementation, in which cloned genes overcome a defect in the mutant.
2. To clone a yeast gene by complementation:
a. A genomic library is made from the wild-type yeast strain in a yeast-E.
coli shuttle vector.
b. The library is transformed into a yeast strain with two mutations, one to allow selection of transformants, and the other a mutation in the gene for which the wild-type genes sought The ARGJ gene is an example (Figure 8.12).
c. Transformed yeast with the wild-type phenotype restored for the gene sought are selected and their plasmid DNA further characterized.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 36
Fig. 8.12 Example of cloning a gene by complementation of mutations: the cloning of
台大農藝系 遺傳學 601 20000 Chapter 7 slide 37
Identifying Specific DNA Sequences in Libraries
Using Heterologous Probes
1. It is possible to identify specific genes in a
genomic library using cloned equivalent genes
(heterologous probes) from other organisms,
especially if the gene is highly conserved or the
species are closely related.
Identifying Genes or cDNAs in Libraries
Using Oligonucleotide Probes
1. Synthetic oligonucleotides are useful in probing libraries, sequence data are available for part of the gene of interest. Knowledge of substitutions produced by mutation also aids probe selection. Sequences for many genes are available in GenBank.
2. Using the universal genetic code, the amino acid sequence is used to design a DNA oligonucleotide probe. Degeneracy in the genetic code means that a mixture of oligonucleotides must be prepared, each of which encodes the target protein.
3. The library is probed with the oligonucleotide mixture to detect the gene of interest. This is a powerful (but not perfect) technique for isolating specific clones.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 39
Molecular Analysis of Cloned DNA
1. Cloned DNA is used in many types of
experiments. Three examples are:
a. Restriction mapping
b. Southern blotting
c. Northern blotting
Restriction Mapping
Animation: Restriction Mapping
1. Cloned DNA can be cut with restriction enzymes and electrophoresed on agarose gels and visualized with ethidium bromide, in order to map its restriction sites (Figure 8.13).
2. Examples of uses for restriction maps:
a. Subcloning sections of gene or cDNA.
b. Confirming results of a cloning experiment. c. Comparing cDNA with its gene.
d. Constructing physical maps of chromosomes.
3. The DNA is cut with several different enzymes, and each cut is loaded in a lane of an agarose gel. Electrical current drives the negatively
charged DNA fragments through the gel. Small molecules move more quickly than large ones, so the fragments are separated by size.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 41
Peter J. Russell, iGenetics: Copyright © Pearson Education, Inc., publishing as Benjamin Cummings.
Fig. 8.13 Constructing a restriction map for EcoRI and BamHI in a DNA fragment
台大農藝系 遺傳學 601 20000 Chapter 7 slide 42
4. DNA is stained with ethidium bromide, which fluoresces
under UV light when complexed with DNA. The gel is
photographed, and the distance migrated by each band of
identical DNA molecules is measured and compared with
a calibration curve to determine the size of each fragment.
5. An example of restriction site mapping is shown in Figure
16.14. A real restriction map is more complex to
generate, involving more restriction sites, and more sites
for each enzyme.
6. Restriction mapping may be done with a circular plasmid,
a cloned sequence, or a fragment of plasmid prepared by
gel cutting.
7. To confirm that the orientation of a cloned DNA insert is
correct, restriction enzymes are selected that will give
台大農藝系 遺傳學 601 20000 Chapter 7 slide 43
Southern Blot Analysis of Sequences in the
Genome
1. Knowing the location of restriction sites in a genome region of interest may be useful for analyzing intron organization or cloning parts of a gene into a vector. 2. The restriction site map can be determined using cDNA or the same gene from a
related species as probe. The process is as follows (Figure 8.16) :
a. Genomic DNA samples are cut with different restriction enzymes.
b. Each sample is electrophoresed in an agarose gel, and stained with ethidium
bromide. Genomic DNA produces a large variety of fragments, which appear as a smear on the gel.
c. DNA is denatured to single strands, and Southern blotting transfers the DNA to a membrane filter. The DNA fragments on the filter are arranged just as they were in the gel.
d. Labeled probe is added to the filter, where it will hybridize with any complementary DNA fragments that were on the original gel. Bound DNA is visualized as
appropriate for the label type.
e. The bands showing a hybridization signal are compared with marker bands to determine their size, and construction of a map is begun. Additional data from individual and combined restriction digests may be needed to complete the map.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 44
台大農藝系 遺傳學 601 20000 Chapter 7 slide 45
Northern Blot Analysis of RNA
1. Northern blotting analyzes RNA much the same way that Southern blotting does DNA:
a. RNA is extracted from the cell, undergoes gel electrophoresis and is bound to a filter.
b. Hybridization between bound cellular RNA and a labeled probe occurs. The sizes of the RNA fragments detected by the probe can be
determined.
2. Northern blot analysis is used for determining:
a. The size(s) of mRNA encoded by a gene. Northern blots have shown that different mRNA species arise from the same region of DNA, suggesting differential use of promoters and terminators, and/or alternative mRNA processing.
b. Whether a specific mRNA is present in a cell type, and if so, at what levels. Gene activity is measured in this way, and RNA sampling is
widely used to study development, tissue specialization, or the response of cells to various physiological stimuli.
DNA Sequencing
Animation: Dideoxy DNA Sequencing
1. Once cloned, DNA fragments may be sequenced,
allowing identification of gene and regulatory
sequences, and comparison with homologous
genes from different organisms
.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 47
2. Dideoxy sequencing (Sanger, 1970s) is based on DNA
polymerase extending short primers, using either linear or
circular DNA as a template. In dideoxy sequencing
(Figure 8.17):
a. DNA is heat denatured, and a short oligonucleotide primer
(designed so 3’ end is near DNA sequence of interest) anneals to
one strand and serves as primer.
b. A reaction mix is set up with label on either the primer or
dNTP(s). The mix includes:
i. ssDNA template to be sequenced.
ii. Primer (anneals to template).
iii. DNA polymerase.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 48
台大農藝系 遺傳學 601 20000 Chapter 7 slide 49
c. The reaction mix is divided into four tubes, and to each is added
a small amount of a different dideoxynucleotide, so that one tube
receives ddCTP, another ddTTP, etc. (Figure 8.18).
d. Dideoxynucleotides lacks 3’-OH, having only 3’-H, and so are
unable to bond with the 5’-phosphate of another nucleotide. A
phosphodiester linkage cannot form, and the chain terminates at
the site of insertion of the ddNTP.
e. The specific ddNTP in a reaction competes with its
corresponding NTP for incorporation into the growing DNA
chain.
f. The four reaction mixes, one for each ddNTP, are typically run in
adjacent lanes on a polyacrylamide gel. The label on the DNA
bands reveals their location, and therefore their sizes.
g. DNA sequence is determined by reading the sequencing ladder
from bottom to top to give the sequence of the newly synthesized
strand from 5’3’ (Figure 8.19).
台大農藝系 遺傳學 601 20000 Chapter 7 slide 50
台大農藝系 遺傳學 601 20000 Chapter 7 slide 51
3. Automation based on the dideoxy method enables
rapid DNA sequencing. In the automated process
(Figure 8.20):
a. Only one reaction mix is needed, containing all four
dideoxynucleotides, each tagged with a different color.
b. DNA fragments generated are separated by
electrophoresis in a single lane.
c. The gel is scanned by a laser device that determines
which fluorescent label is present at each nucleotide
position in the sequence, and sends the information to
a computer.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 53
Fig. 8.20 Results of automated DNA sequence analysis using fluorescent dyes
4. Computer programs can analyze DNA sequences using
new data along with databases of previously reported
sequences. Computer analysis is used to:
a. Look for restriction sites.
b. Compare a variety of sequences from the same or different
species.
c. Locate homologous regions.
d. Find transcription regulatory sequences.
e. Detect open reading frames and predict the amino acid sequence
encoded, including protein structure and function.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 55
Polymerase Chain Reaction (PCR)
1. PCR starts with a mixture of DNA molecules and produces many copies of one specific DNA sequence (amplimers). Mullis developed the technique in the 1980s.
2. PCR begins with DNA containing the sequence to be amplified, and a pair of synthetic primers that flank the sequence. Steps in the process include (Figure 8.21):
a. Heat denature DNA (94–95°C).
b. Cool (37–65°C) and anneal primers to complementary sequences with their 3’ ends facing each other, flanking the target DNA.
c. Extend the primers (70–75°C) with heat-resistant DNA polymerase from the thermophilic bacterium, Thermus aquaticus.
d. Repeat the cycle of denaturation and primer binding.
e. Extend again with Taq DNA polymerase. Products the length of the target sequence begin to be produced.
f. Repeat the process, doubling the amount of target DNA with each round of extension.
3. In 20 PCR cycles, million-fold amplification of the target sequence occurs. The temperature changes are automated in a thermal cycler.
台大農藝系 遺傳學 601 20000 Chapter 7 slide 56
Fig. 8.21 The polymerase chain reaction (PCR) for selective amplification of DNA
台大農藝系 遺傳學 601 20000 Chapter 7 slide 57
Advantages and Limitations of PCR
1. PCR is more sensitive and faster than cloning, but there are limitations:
a. Specific primers require that sequence information be known.
b. Taq polymerase does not proofread, meaning that mismatches go uncorrected. Alternative polymerases such as Vent polymerase do proofread, decreasing errors. c. The sensitivity can result in amplification of contaminating sequences, a special
hazard in forensic applications.
2. Applications of PCR include:
a. Amplifying DNA for cloning.
b. Amplifying DNA from genomic DNA for sequencing without cloning. c. Mapping DNA segments.
d. Disease diagnosis.
e. Subcloning segments of cloned DNA (e.g., the yeast ARG1 gene)
i. Individual genes may be amplified from a cloned multigene DNA fragment. ii. Complementation is used to determine functions of each gene.
f. Forensics (the analysis of legal evidence) in samples including hair, blood, or semen. g. The study of molecular evolution.
RT-PCR and mRNA Quantification
1. Reverse transcriptase allows amplification of RNA by PCR in two steps:
a. cDNA is synthesized from RNA using oligo(dT) as primer and reverse transcriptase as polymerase.
b. The cDNA is amplified by PCR.
2. RT-PCR can:
a. Test for presence of an RNA (e.g., detect RNA virus genomes). b. Quantify an mRNA to determine the amount of gene expression.
i. DNA product is analyzed by agarose gel electrophoresis with ethidium bromide.
ii. Amount of product is visualized by intensity of fluorescence with UV. iii. Results are only semiquantitative. Northern blots are more accurate for quantifying mRNAs.
3. Real time RT-PCR is also more accurate to quantify mRNA levels.
a. Reverse transcriptase is used for the first step, as in RT-PCR.
b. DNA amplification done in the presence of SYBR green, a dye that stains dsDNA. c. Laser detector in the thermal cycler detects SYBR green fluorescence, quantifying