© CSIRO 2002 10.1071/MF01135 1323-1650/02/020411
Early life history of Acanthopagrus latus and A. schlegeli (Sparidae)
on the western coast of Taiwan: temporal and spatial partitioning
of recruitment
Chih-Wei Chang
A, Chih-Chieh Hsu
B, Yu-Tzu Wang
A, Wann-Nian Tzeng
ACA
Institute of Zoology, College of Science, National Taiwan University, Taipei, Taiwan 106, ROC
B
Department of Biology, College of Science, National Taiwan Normal University, Taipei, Taiwan 106, ROC
C
Corresponding author: [email protected]
MF01135 Early life history of Chih-Wei Chang
etal.C.-W.Chang, C.-C.HsuY.-T.Wang, W.-N.Tzeng
Abstract. Acanthopagrus latus and A. schlegeli are phylogenetically closely related. The seasonal occurrence and
distribution, age and length at recruitment, growth rate and hatching dates of their larvae were compared by collecting fortnightly specimens from four estuaries on the north-eastern and western coast of Taiwan between September 1997 and August 1998. Age and growth rate were determined from the daily growth increments in otoliths of the larvae. Occurrences of A. latus and A. schlegeli larvae on the western coast of Taiwan were temporally and spatially separated. Recruitment of the former progressed from north to south from November through March, whereas the latter progressed from south to north from December through May. Age and length of the larvae at estuarine arrival were greater in A. latus; these values increased southward for A. latus and northward for A. schlegeli. Age at recruitment was inversely correlated with growth rate which was positively correlated to water temperature. A. latus spawned mainly in autumn, whereas A. schlegeli spawned mainly in spring with a minor spawning peak in autumn. Coastal currents and the spawning behaviour of the adults may influence the geographic gradients of the seasonal occurrence and distribution of the larvae on the western coast of Taiwan.
Introduction
Acanthopagrus spp. are widely distributed in shallow coastal
waters, lagoons and estuaries of the Indo–West Pacific (Springer 1982). Adults spawn in coastal waters and the larvae move to estuaries (Hu 1983). Four species of
Acanthopagrus are recorded from Taiwan: Acanthopagrus australis, A. berda, A. latus and A. schlegeli (Jean et al.
1992). Yellow sea bream (A. latus) and black porgy (A. schlegeli) are comparatively abundant and economically important for recreation and aquaculture. In Taiwan, wild larvae and juveniles of these two species have been harvested in estuaries for restocking for many years. However, many aspects of their natural life history are poorly understood.
The otolith is a powerful proxy for describing the early life history of fish (Pannella 1971; Campana and Neilson 1985). Daily growth increments in otoliths are widely used to determine larval ages, to estimate their growth rates and hatching dates, as well as to separate different cohorts of a species with protracted spawning period (Tzeng 1990; Stevenson and Campana 1992; Tzeng et al. 1998; Wang and Tzeng 1999; Chang et al. 2000). Previous studies of the
Acanthopagrus spp. in Taiwan have focused mainly on
aquaculture (Tang and Twu 1979), artificial propagation (Hu 1983; Leu et al. 1991) and taxonomy (Lee 1983; Jean et al.
1992, 1995). Age and growth of A. schlegeli larvae have been studied for a particular estuary on the western coast of Taiwan (Huang and Chiu 1997b). Information on the seasonal occurrence, abundance and distribution of A. latus and A. schlegeli larvae on the western coast of Taiwan is fragmentary (Liu 1978; Huang et al. 1985; Hung and Chiu 1991; Tzeng and Wang 1992, 1993, 1997; Tzeng 1995; Huang and Chiu 1997a; Tzeng et al. 1997).
A. latus and A. schlegeli are similar phylogenetically,
morphologically and in their habitat use. This raises the question of whether members of these two species compete for habitat use and food resources. If they do not, how have they evolved differences in habitat use in time and space? To elucidate this, the early life history of these species on the western coast of Taiwan was studied, including seasonal occurrence and distribution, age and length at recruitment, growth rates and hatching dates and the linkage between larval dispersal and coastal currents induced by the monsoon.
Materials and methods
Sampling design
Fish larvae and juveniles were collected while immigrating into four estuaries, Shuangchi Creek (SC), Gongshytyan Creek (GST), Tatu Creek (TT) and Tongkang River (TK) (Fig. 1). The west coast of
Taiwan is separated from China by the Taiwan Strait which has a wide and shallow continental shelf suitable as a spawning and nursery area for coastal fishes. In contrast, on the east coast the continental shelf is narrow except in the north-east and is influenced by the Kuroshio current. The direction of coastal currents on the western coast that transport larvae to the estuary changes with the seasonal monsoon (Chu 1963). Water temperatures on the western coast decrease when the NE monsoon prevails in autumn and winter and increase when the SW monsoon prevails in spring and summer.
An anchored net was set against the tidal current in the estuaries to collect fish larvae and juveniles during the night-time flood tide around the new and full moons between September 1997 and August 1998. The mesh size of the net ranged from 0.8 to 1.8 mm. A. latus and A. schlegeli larvae were found abundantly from October through May, which is the commercial fishing season for collecting the larvae for restocking. Larval abundance was estimated roughly by the number of fish caught per hour. Surface water temperature was also measured to 0.1°C on a microprocessor conductivity meter (Model LF196, Wissenschaftlich-Technische Werkstätten) during sampling.
Specimen preparation and measurement
Both Acanthopagrus species were sorted and separated from the other larvae collected by the net. They were identified in fresh samples according to the melanophore distribution patterns as described by Okiyama (1988). The larvae were then preserved in 95% alcohol for length measurement and otolith analysis. Larval total length was measured to the nearest 0.1 mm at a magnification of 10X by a profile projector. Larval shrinkage due to alcohol preservation was not adjusted for. The shrinkage of total length was estimated as ~4.7± 1.7% (n = 39).
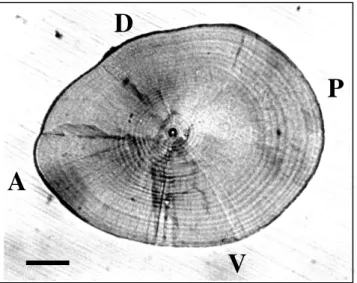
Sagittal otoliths of the larvae were extracted, air-dried, embedded in resin, ground, polished and etched. Preparation of otoliths for age determination followed Chang et al. (2000). Growth increments in the larval otoliths were clearly discernible by microscope under reflected light and no sub-daily increments were found (Fig. 2). The increments
in otoliths of A. schlegeli larvae have been validated as deposited daily (Huang and Chiu 1997b). The increments in otoliths of A. latus were assumed to be deposited daily because they are related species. Daily growth increments in the magnified (300–600X) photographs of otoliths were counted independently by two people. The daily age of the larvae was the average of the daily growth increments from these two independent counts if the difference in counts between readers was <5%; otherwise, the otolith was excluded from the age estimation. Larval hatching dates were back-calculated from their daily age and capture dates. The increment widths on the maximum otolith radius of the larvae were measured to evaluate the growth rate of an individual on the assumption that otolith and somatic growth are coupled. Increment widths were measured by digital caliper on the magnified (300–600×) photographs.
Data analysis
Significant differences in age and length between species and among estuaries and sampling dates were determined by one-way analysis of variance (ANOVA) and Tukey multiple-comparison test (Winer 1971).
Results
Spatial distribution
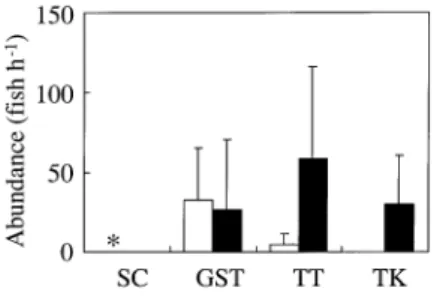
The A. latus larvae were most abundant at GST (32.2 ± 33.14
fish h–1), followed by TT (4.8 ± 6.3 fish h–1) and SC
(0.3 fish h–1). In contrast, A. schlegeli larvae were most
abundant at TT (58.1 ± 57.9 fish h–1
), followed by TK
(30.0± 30.5 fish h–1) and GST (26.8 ± 43.5 fish h–1). The
larvae of A. latus were not caught in TK and A. schlegeli not in SC (Fig. 3). The density distribution of the larvae indicates that A. latus may originate from the north and disperse southward. In contrast, A. schlegeli may originate from the south and disperse northward.
Geographic gradient in seasonal occurrence of larvae and water temperature
The geographic gradient in seasonal occurrence was opposite for these two species (Fig. 4). Larvae of A. latus
Fig. 1. Sampling sites of A. latus and A. schlegeli larvae in the estuaries of Shuangchi Creek (SC), Gongshytyan Creek (GST), Tatu Creek (TT) and Tongkang River (TK). Dashed line, 200 m depth contour.
P
A
D
V
Fig. 2. Daily growth increments in the sagittal otolith of a 12.3 mm TL A. schlegeli larva. A, anterior; P, posterior; D, dorsal; V, ventral. Scale bar, 100 µm.
occurred earlier in the north (GST) than in the central west (TT) in the period between November and February. In contrast, larvae of A. schlegeli occurred first in the south (TK), then in the central west (TT) and later in the north (GST) in the period from December through May.
The peak abundance of A. latus larvae occurred at water
temperatures of 20.5–20.9°C in GST and 18.3°C in TT,
whereas for A. schlegeli larvae, water temperatures were
24.0–30.0°C in TK, 24.0–25.6°C in TT and 26.3°C in GST
(Fig. 4). Thus, the water temperature at peak larval occurrence was higher for A. schlegeli than for A. latus, but the temperature at peak occurrence of each species was similar among areas and shifted with the season. That is, the occurrence of larvae in the estuaries followed the seasonal changes in water temperature. A. latus larvae occurred early
in the north and later in the south, following the seasonal decrease in temperatures during autumn. In contrast,
A. schlegeli larvae occurred early in the south and later in the
north, following the seasonal increase in temperatures during spring (February–April), although a minor group of
A. schlegeli larvae (TK in December and TT in January), like A. latus, occurred in autumn following the seasonal decrease
in temperatures.
Spatial and temporal variations in age and length of the larvae at recruitment
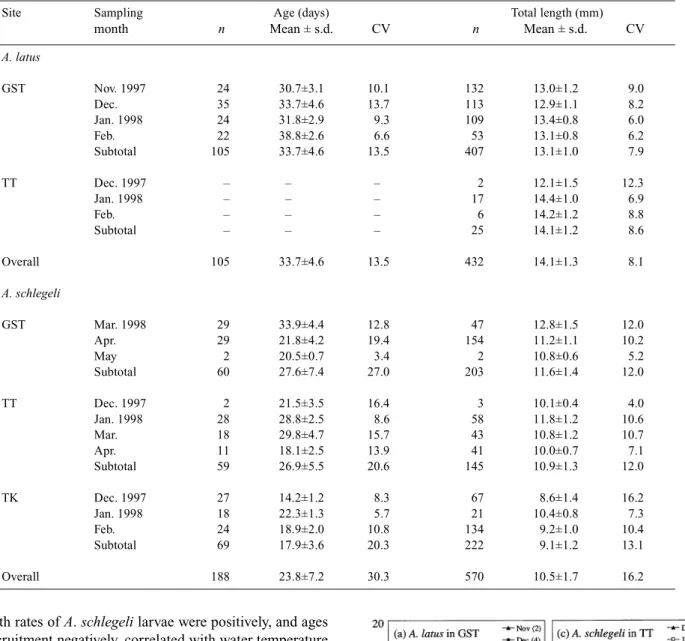
Mean ages and lengths of A. latus larvae (33.7 ± 4.6 days,
14.1 ± 1.3 mm TL) were greater than for A. schlegeli larvae
(23.8 ± 7.2 days, 10.5 ± 1.7 mm TL)(P <0.001). The mean
ages of A. schlegeli larvae did not differ significantly between GST (27.6 ± 7.4 days) and TT (26.9 ± 5.5 days), but were older in both those estuaries than in TK (17.9 ± 3.6 days) (P <0.001). A. latus larvae were larger in TT (14.1 ± 1.2 mm) than in GST (13.1 ± 1.0 mm) (P <0.001). Similarly, mean lengths of A. schlegeli larvae were greatest in GST (11.6 ± 1.4 mm), intermediate in TT (10.9 ± 1.3 mm) and smallest in TK (9.1 ± 1.2 mm) (P <0.001). The geographic gradients in mean ages and lengths of A. latus increased from north to south but A. schlegeli increased from south to north, again suggesting that the dispersal direction of the larvae differs between species.
The mean ages and lengths of A. latus and A. schlegeli larvae at recruitment also differed significantly among sampling dates (P <0.005), perhaps because the growth history and recruitment process of the larvae differed among cohorts. At recruitment, the larvae were less variable in size (lower CV) than in age (Table 1).
Temporal changes in growth rate
Mean increment widths in otoliths of A. latus larvae collected from GST in November increased from ~2 µm to 11 µm at 29 days after hatching, whereas those collected in December and February only increased to ~6 µm 36 days after hatching (Fig. 5a). The age of A. latus larvae at recruitment was thus inversely correlated to a growth rate that was faster in November than in December and February. The monthly difference in growth rate and age at recruitment of A. latus larvae may be due to the higher water temperatures in November than in December and February (Fig. 4a).
A similar phenomenon was also found in A. schlegeli larvae collected in GST, TT and TK (Figs 5b–d). The larvae in GST grew more quickly and had a smaller age at recruitment in April and May than in March. In TT the larva collected in December had grown faster and was younger than the larva collected in January, and the one collected in April had grown faster and was younger than the one in March. Larvae in TK grew faster and were younger at recruitment in December than in January and February. The
Fig. 3. Abundance of (䊐) A. latus and (䊏) A. schlegeli larvae in four estuaries, Shuangchi Creek (SC), Gongshytyan Creek (GST), Tatu Creek (TT) and Tongkang River (TK). Vertical lines, s.d. *Small number of A. latus.
Fig. 4. Seasonal occurrence of (䊐) A. latus and (䊏) A. schlegeli larvae in (a) Gongshytyan Creek (GST), (b) Tatu Creek (TT) and (c) Tongkang River (TK) estuaries, November 1997 through May 1998. 䊊, water temperature.
growth rates of A. schlegeli larvae were positively, and ages at recruitment negatively, correlated with water temperature (Fig. 4). Growth rates were faster in A. schlegeli than in
A. latus. Hatching date
A. latus and A. schlegeli differed in hatching dates (Fig. 6). A. latus hatched between late September and early January
with a peak in October in GST (Fig. 6a). A. schlegeli hatched from late November to January, with peaks in early December and late January in TK (Fig. 6c), from late November to March with peaks in December and March in TT (Fig. 6b), and from February to April with peaks in February and March in GST (Fig. 6a). The hatching dates of
A. latus and A. schlegeli were separated either temporally, if
they occurred in the same estuary, or spatially if they occurred in the same season.
Table 1. Mean age and length of A. latus and A. schlegeli larvae at recruitment in the Gongshytyan Creek (GST), Tatu Creek (TT) and Tongkang River (TK) estuaries
n, sample size. –, problems with specimen preservation
Site Sampling Age (days) Total length (mm)
month n Mean ± s.d. CV n Mean ± s.d. CV
A. latus GST Nov. 1997 24 30.7±3.1 10.1 132 13.0±1.2 9.0 Dec. 35 33.7±4.6 13.7 113 12.9±1.1 8.2 Jan. 1998 24 31.8±2.9 9.3 109 13.4±0.8 6.0 Feb. 22 38.8±2.6 6.6 53 13.1±0.8 6.2 Subtotal 105 33.7±4.6 13.5 407 13.1±1.0 7.9 TT Dec. 1997 – – – 2 12.1±1.5 12.3 Jan. 1998 – – – 17 14.4±1.0 6.9 Feb. – – – 6 14.2±1.2 8.8 Subtotal – – – 25 14.1±1.2 8.6 Overall 105 33.7±4.6 13.5 432 14.1±1.3 8.1 A. schlegeli GST Mar. 1998 29 33.9±4.4 12.8 47 12.8±1.5 12.0 Apr. 29 21.8±4.2 19.4 154 11.2±1.1 10.2 May 2 20.5±0.7 3.4 2 10.8±0.6 5.2 Subtotal 60 27.6±7.4 27.0 203 11.6±1.4 12.0 TT Dec. 1997 2 21.5±3.5 16.4 3 10.1±0.4 4.0 Jan. 1998 28 28.8±2.5 8.6 58 11.8±1.2 10.6 Mar. 18 29.8±4.7 15.7 43 10.8±1.2 10.7 Apr. 11 18.1±2.5 13.9 41 10.0±0.7 7.1 Subtotal 59 26.9±5.5 20.6 145 10.9±1.3 12.0 TK Dec. 1997 27 14.2±1.2 8.3 67 8.6±1.4 16.2 Jan. 1998 18 22.3±1.3 5.7 21 10.4±0.8 7.3 Feb. 24 18.9±2.0 10.8 134 9.2±1.0 10.4 Subtotal 69 17.9±3.6 20.3 222 9.1±1.2 13.1 Overall 188 23.8±7.2 30.3 570 10.5±1.7 16.2
Fig. 5. Increment widths in otoliths of (a) A. latus and (b–d) A. schlegeli larvae. In parenthesis, sample size. Vertical lines, s.d.
Discussion
Effect of physical processes on the larval distribution
Along the west coast of Taiwan there were opposite gradients in the seasonal occurrence and distribution of A. latus and
A. schlegeli larvae. A. latus larvae occurred progressively
from north to south between late autumn and late spring, whereas A. schlegeli larvae occurred progressively from south to north between winter and spring.
The opposite distribution patterns of the larvae of these two species coincides with the coastal currents on the western coast of Taiwan. In autumn and winter, the north-eastern monsoon prevails and induces a southward flow of the cold China Coastal Current from the East China Sea into Taiwan Strait, causing a decline in water temperatures on the western coast of Taiwan (Chu 1963). A. latus spawns in autumn and thus the larvae drift southward with the coastal current toward the western coast of Taiwan. However, a branch of the warm Kuroshio Current stops the cold China Coastal Current in the middle of Taiwan Strait (Fan 1982), which limits the southward transport of A. latus larvae. In contrast, in spring and summer the south-western monsoon prevails and induces a warm coastal current flow northward into the Taiwan Strait from the South China Sea, causing water temperatures to increase from south to north along the western coast (Chu 1963). Thus, A. schlegeli larvae are dispersed northward with the current to the northern Taiwan Strait. In contrast with the China Coastal Current, the South China Sea monsoon current can reach the northern part of
Taiwan Strait. This allows A. schlegeli to disperse to the north-western coast whereas A. latus can only disperse to the central western coast of Taiwan.
Variability of age and length of the larvae at recruitment
The recruitment of larvae to the estuaries may depend on size rather than age (Houde 1987). Our study supports this conclusion because the coefficient of variation of mean ages was larger than those of mean lengths for both A. latus and
A. schlegeli larvae. Age at recruitment was inversely
correlated to larval growth rate, as was found for other fishes (Tzeng 1990; Rutherford and Houde 1995; Wang and Tzeng 1999). Thus, the faster growing larvae arrived at the estuary at a younger age. In addition, the geographic gradient in the mean age of A. schlegeli larvae at recruitment, which was younger in the southern estuary (TK) than in the northern estuary (GST), corresponds to the geographic gradient in temperature which is higher in the south than in the north. Water temperature evidently plays an important role in determining the age and length of the larvae at recruitment.
Spawning and hatching date
A. latus and A. schlegeli hatched respectively at ~60 h and
41–50 h after fertilization at temperatures of 17.5–20°C (Lin and Yen 1980; Liu and Hu 1980; W. Y. Tzeng 1982; Leu
et al. 1991), i.e. the larvae hatch about 2–3 days after
spawning. The back-calculated hatching dates of A. latus larvae in GST lasted from late September to early January and were consistent with the October–December spawning period that was estimated from the gonadosomatic index (GSI) of mature fish in the Pescadores Islands of central western Taiwan (Liu and Hu 1980). Similarly, the hatching dates of A. schlegeli (from late November to January in TK, late November to March in TT, and February to April in GST) were also consistent with the January–March spawning period that was estimated from GSI of mature fish in central western Taiwan (Lin and Yen 1980).
Effect of spawning behaviour on larval distribution
The hatching date distributions, and thus the spawning dates, of both A. latus and A. schlegeli were extended and did not overlap among estuaries but shifted with the season. The spawning behaviour of adult fish is the basis of the geographical gradients in seasonal occurrence and distribution of the larvae. Temperature is one of the principal factors determining the seasonal maturation and spawning of fish (Crecco and Savoy 1985; Rutherford and Houde 1995). The onset of spawning of sparids depends on the occurrence of optimal temperature (Crossland 1980; Kojima 1981; Hu 1983; Scott and Pankhurst 1992; Francis 1994). Water temperatures increase from south to north on the western coast of Taiwan in spring causing the timing of spawning and larval hatching of A. schlegeli to shift with season and
Fig. 6. Distribution of hatching dates of (䊐) A. latus and (䊏) A. schlegeli larvae in (a) Gongshytyan Creek (GST), (b) Tatu Creek (TT) and (c) Tongkang River (TK) estuaries.
latitude. This phenomenon was also found in white croaker,
Argyrosomus argentatus in the Taiwan Strait (Tzeng and Liu
1972). On the other hand, A. latus spawned in autumn (October) and a minor spawning period occurred in late autumn (November–December) for A. schlegeli, following the seasonal decrease in water temperature. The seasonal occurrence of A. latus larvae was consequently shifted from north to south, while A. schlegeli larvae were shifted from south to north. The seasonal occurrence and geographic distribution of larvae was highly correlated with adult spawning behaviour.
In conclusion, A. latus and A. schlegeli larvae on the western coast of Taiwan were opposite in terms of the geographic gradient in seasonal occurrence and distribution, mean age and length at recruitment, growth rate and hatching date. The monsoon-induced coastal current, water temperature, spawning behaviour and larval growth rate all have the potential to influence the variability of the geographic gradient in seasonal occurrence and distribution of larvae of these two species.
Acknowledgments
The study was financially supported by the Council of Agriculture (87-AST–1.4-FID–05(09), 88-AST–1.4-FID– 05(18) and 89-AST–1.2-FID–02(06)) and National Science Council (NSC89–2611-M–002–006 and NSC89–2611-M– 002–039) of the Republic of China. The authors are grateful to Messrs C. S. Jean, C. S. Lu, C. S. Chen and C. L. Chen for collection of specimens, to Mrs H. Y. Teng for specimen sorting, to Mr S. Y. Chang and Ms Y. Y. Liu for specimen preparation, and to Mr B. M. Jessop for helpful comments.
References
Campana, S. E., and Neilson, J. D. (1985). Microstructure of fish otolith. Canadian Journal of Fisheries and Aquatic Sciences 42, 1014–32.
Chang, C. W., Tzeng, W. N., and Lee, Y. C. (2000). Recruitment and hatching dates of grey mullet (Mugil cephalus L.) juveniles in the Tanshui Estuary of northwest Taiwan. Zoological Studies 39, 99–106.
Chu, T. Y. (1963). The oceanography of the surrounding waters of Taiwan. Report of the Institute of Fishery Biology of Ministry of Economic Affairs and National Taiwan University 1, 29–44. Crecco, V. A., and Savoy, T. F. (1985). Effects of biotic and abiotic
factors on growth and relative survival of young American shad, Alosa sapidissima, in the Connecticut River. Canadian Journal of Fisheries and Aquatic Sciences 42, 1640–8.
Crossland, J. (1980). The number of snapper, Chrysophrys auratus (Forster), in the Hauraki Gulf, New Zealand, based on egg surveys in 1974–75 and 1975–76. New Zealand Ministry of Agriculture and Fisheries, Fisheries Research Bulletin No. 22, 38 pp.
Fan, K. L. (1982). A study of water masses in Taiwan Strait. Acta Oceanographica Taiwanica 13, 140–53.
Francis, M. P. (1994). Duration of larval and spawning periods in Pagrus auratus (Sparidae) determined from otolith daily increments. Environmental Biology of Fishes 39, 137–52. Houde, E. D. (1987). Fish early life dynamics and recruitment
variability. American Fisheries Society Symposium 2, 17–29.
Hu, S. H. (1983). Artificial breeding of Sparidae in Penghu – with emphasis on red seabream, Chrysophrys major and black porgy, Acanthopagrus schlegeli. Annual Collected Report of Penghu Branch Taiwan Fisheries Research Institute 3, 1–48.
Huang, C. C., Tzeng, W. N., and Lee, S. C. (1985). Preliminary survey on larval fishes of Yen-Liao Bay, northeastern Taiwan. Bulletin of the Institute of Zoology, Academia Sinica 24, 147–54.
Huang, W. B., and Chiu, T. S. (1997a). Environmental factors associated with the occurrence and abundance of larval porgies, Acanthopagrus latus and Acanthopagrus schlegeli, in the coastal waters of western Taiwan. Acta Zoologica Taiwanica 8, 19–32. Huang, W. B., and Chiu, T. S. (1997b). Daily increments in otoliths and
growth equation of black porgy, Acanthopagrus schlegeli, larvae. Acta Zoologica Taiwanica 8, 121–31.
Hung, J. B., and Chiu, T. S. (1991). Eco-geographic difference of larval fish assemblage in the coastal waters of the western central Taiwan. Journal of the Fisheries Society of Taiwan 18, 241–56.
Jean, C. T., Lee, S. C., and Chen, C. T. (1992). Morphometric studies on the fishes of subfamily Sparinae (Perciformes: Sparidae) from the coastal waters of Taiwan. Journal of the Fisheries Society of Taiwan 19, 231–8.
Jean, C. T., Hui, C. F., Lee, S. C., and Chen, C. T. (1995). Variation in mitochondrial DNA and phylogenetic relationships of fishes of the subfamily Sparinae (Perciformes: Sparidae) in the coastal waters of Taiwan. Zoological Studies 34, 270–80.
Kojima, K. (1981). Spawning of the red bream (Pagrus major) in the waters around Iki Is. and Mishima Is. in the western part of the Japan Sea. Bulletin of the Seikai Region, Fisheries Research Laboratory 56, 71–87.
Lee, S. C. (1983). The family Sparidae (Pisces: Perciformes) of Taiwan. Journal of Taiwan Museum 36, 47–55.
Leu, M. Y., Chou, Y. H., Wu, C. H., and Lin, I. C. (1991). Induced spawning and mass production of the seedling of yellow-finned black porgy, Acanthopagrus latus (Houttuyn). Bulletin of Taiwan Fisheries Research Institute 50, 129–39.
Lin, K. J., and Yen, J. L. (1980). Artificial propagation of black porgy, Acanthopagrus schlegeli. Bulletin of Taiwan Fisheries Research Institute 32, 701–9.
Liu, C. H. (1978). Experiment on Acanthopagrus schlegeli (Bleeker) in Taiwan waters. China Fisheries Monthly 311, 3–6.
Liu, F. G., and Hu, S. H. (1980). Preliminary report on the artificial fertilization and incubation of yellowfin porgy, Acanthopagrus latus. Bulletin of Taiwan Fisheries Research Institute 32, 673–8. Okiyama, M. (Ed). (1988). ‘An Atlas of the Early-stage Fishes in
Japan.’ (Tokai University Press: Tokyo.)
Pannella, G. (1971). Fish otoliths: daily growth layers and periodical patterns. Science 173, 1124–7.
Rutherford, E. S., and Houde, E. D. (1995). The influence of temperature on cohort-specific growth, survival, and recruitment of striped bass, Morone saxatilis, larvae in Chesapeake Bay. Fisheries Bulletin US 93, 315–32.
Scott, S. G., and Pankhurst, N. W. (1992). Interannual variation in the reproductive cycle of the New Zealand snapper Pagrus auratus (Bloch & Schneider) (Sparidae). Journal of Fish Biology 41, 685–96.
Springer, V. G. (1982). ‘Pacific Plate Biogeography, with Special Reference to Shorefishes.’ (Smithsonian Institution Press: Washington.)
Stevenson, D. K., and Campana, S. E. (1992). Otolith microstructure examination and analysis. Canadian Special Publication of Fisheries and Aquatic Sciences No. 117, 126 pp.
Tang, H. C., and Twu, J. Y. (1979). Experiments on the culture of black porgy, Acanthopagrus schlegeli. China Fisheries Monthly 319, 3–8.
http://www.publish.csiro.au/journals/mfr Tzeng, W. N. (1990). Relationship between growth rate and age at
recruitment of Anguilla japonica elvers in a Taiwan estuary as inferred from otolith growth increments. Marine Biology 107, 75–81.
Tzeng, W. N. (1995). Recruitment of larval and juvenile fishes to the Gong-Shy-Tyan River estuary of Taiwan: relative abundance, species composition. In ‘Condition of the World’s Aquatic Habitats’. (Eds N. B. Armantrout and R. J. Wolotira Jr.) pp. 360–85. (Oxford and IBH Publication: New Delhi.)
Tzeng, W. N., and Liu, H. C. (1972). Maturity and fecundity of white croaker, Argyrosomus argentatus (Houttuyn), in the East China Sea and the Taiwan Strait. Journal of the Fisheries Society of Taiwan 1, 20–30.
Tzeng, W. N., and Wang, Y. T. (1992). Structure, composition and seasonal dynamics of the larval and juvenile fish community in the mangrove estuary of Tanshui River, Taiwan. Marine Biology 113, 481–90.
Tzeng, W. N., and Wang, Y. T. (1993). Hydrography and distribution dynamics of larval and juvenile fishes in the coastal waters of the Tanshui River estuary, Taiwan, with reference to estuarine larval transport. Marine Biology 116, 205–17.
Tzeng, W. N., and Wang, Y. T. (1997). Movement of fish larvae with tidal flux in the Tanshui River Estuary, northern Taiwan. Zoological Studies 36, 178–85.
Tzeng, W. N., Wang, Y. T., and Chern, Y. T. (1997). Species composition and distribution of fish larvae in Yenliao Bay, northeastern Taiwan. Zoological Studies 36, 146–58.
Tzeng, W. N., Wu, C. E., and Wang, Y. T. (1998). Age of Pacific tarpon, Megalops cyprinoides, at estuarine arrival and growth during metamorphosis. Zoological Studies 37, 177–83.
Tzeng, W. Y. (1982). On the induced breeding and larval rearing of the black porgy, Mylio macrocephalus (Basilewsky) in Hong Kong. Journal of Taiwan Museum 35, 155–65.
Wang, Y. T., and Tzeng, W. N. (1999). Differences in growth rates among cohorts of Encrasicholina punctifer and Engraulis japonicus larvae in the coastal waters off Tanshui River Estuary, Taiwan, as indicated by otolith microstructure analysis. Journal of Fish Biology 54, 1002–16.
Winer, B. (1971). ‘Statistical Principles in Experimental Design.’ (McGraw-Hill: New York.)