台灣地區外來種緬甸小鼠的族群遺傳結構與生物地理起源
Genetic Patterns and Biogeographic Origins of the Exotic
Rodent (Rattus exulans) in Taiwan
儲瑞華
1曹又仁
2莊媛茹
2陳皇奇
3林曜松
1吳海音
3, *Jui-Hua Chu
1, Yu-Jen Tsao
2, Yuan-Ju Chuang
2, Huang-Chi Chen
3,
Yao-Sung Lin
1and Hai-Yin Wu
3,*1國立台灣大學生態與演化生物學研究所 台北市羅斯福路四段1 號 2天主教輔仁大學生命科學系 台北縣新莊市中正路510 號 3國立東華大學自然資源管理研究所 花蓮縣壽豐鄉志學村大學路二段1 號 1Institute of Evolutionary Biology and Ecology, National Taiwan University, Taipei, Taiwan
2Department of Life Science, Fu Jen Catholic University, Taipei, Taiwan 3Institute of Natural Resources, National Dong Hwa University, Hualien, Taiwan
* 通訊作者 * Corresponding author
摘
要
本研究利用SSCP (single-strand conformation polymorphism)技術,分析近年發現入侵台灣之 外來種緬甸小鼠(Rattus exulans)粒線體 DNA 單倍體基因型(haplotype)的多型性,探尋其可能來 源,並檢視各基因型在花蓮縣吉安鄉的分布狀況,以推論此外來種族群的擴散模式。在粒線體 DNA 控制區(mtDNA control region)中,以 SSCP 共判別出 4 種單倍體基因型(Type I-IV),其中 Type I 和 II 占大多數,且分布最廣;Type III 和 IV 則較少,皆僅見於單一捕捉點。經與 NCBI 資料庫 中的序列比對,發現Type I 與分布於泰國的序列相同,而 Type IV 則與蘇拉維西的序列相近,推 測入侵台灣的緬甸小鼠來自東南亞大陸及週邊島嶼區域,而由各基因型間的遺傳變異程度判斷, 緬甸小鼠可能至少有2 次以上的入侵史。
Abstract
Mitochondria DNA (mtDNA) haplotypes were identified with single-strand conformation polymorphism (SSCP) for the exotic rodent (Rattus exulans) found in Jian Township, Hualien County, Taiwan. The objectives of this study were to examine distribution patterns of the haplotypes and to determine biogeographical origins of this exotic rat. We found four distinct SSCP haplotypes, Types I, II, III and IV, in a segment of 372 base-pairs in the mtDNA control region. Types I and II were dominant and occurred widely in the sampling area, while Types III and IV were restricted, each to a single sampling location. When the sequences of the four haplotypes obtained in this study were compared with the sequences of 32 R. exulans haplotypes available in NCBI, Type I was identical to the sequence from Thailand, Type IV differed to one sequence from Sulawesi by only one base pair. The results indicated that this species of exotic rat in Hualien might be originated from the Southeast Asia mainland and Malay Archipelago. The presence of four haplotypes suggested that there were at least two invasion events. 關鍵詞:緬甸小鼠、SSCP、外來種、粒線體 DNA 控制區域
Key words: Rattus exulans, SSCP, exotic species, mtDNA control region
收件日期:95 年 3 月 6 日 接受日期:95 年 7 月 27 日 Received: March 6, 2006 Accepted: July 27, 2006
緒
言
緬甸小鼠(Rattus exulans)是 1999 年 4、5 月間,始被發現入侵台灣的外來鼠種。是年, 行政院農業委員會農業藥物毒物試驗所協同花 蓮區農業改良場,於花蓮縣吉安鄉(121 34 E, 23 57 N)休耕甘藷田監測田間鼠類組成時,捕 獲此外來種(陳及高 2001),而後經 Motokawa et al. (2001)確認其種類。緬甸小鼠在分類上屬 於鼠科(Murinae)、家鼠屬(Rattus),其全球分 布範圍僅次於小家鼠(Mus musculus)、家鼠(R. rattus)與溝鼠(R. norvegicus),是分布第四廣泛的鼠種(Wodzicki and Taylor 1984),主要廣布
於東南亞及大洋洲上各島嶼之亞熱帶及熱帶地 區(Wirtz 1973; Corbet and Hill 1992),台灣及 琉球(Ryukyus)則是在近幾年才確認有緬甸小 鼠 的 蹤 跡(陳 及 高 2001; Motokawa et al. 2001)。
成熟緬甸小鼠的頭軀幹長約11.5- 15 cm, 尾長約與頭軀幹長相等,後腳長約23 mm,平 均體重為36-80g (Lekagul and McNeely 1988; Tobin 1994)。小鼠的生殖表現因地理區的不同 而有變化(Wirtz 1972; Tobin 1994),入侵台灣 花蓮地區之緬甸小鼠則在春季與夏季時處生殖 狀況之個體的比例較高(吳海音,未發表資料), 懷孕雌鼠的胚胎數在1-7 隻間,以 4-5 個胚胎
較為常見(吳等 2003)。緬甸小鼠偏好人類活動 造成與維持的多類環境,包括住家、穀倉、耕 地、灌 叢、林 地 邊 緣(Musser and Newcomb 1983; Wodzicki and Taylor 1984; Corbet and Hill 1992)。其於台灣主要棲息於植被高度 1m 以 下,且植被組成以非禾本科草本植物為主的休 廢耕草生地(吳等 2001)。 由田野調查中捕獲之緬甸小鼠數量及年齡 判析,此種鼠類侵入花蓮縣吉安鄉已有一段時 間,並已建立繁殖族群(盧及徐 2003)。後續 相關單位的調查結果顯示,緬甸小鼠並未出現 於台灣各主要港埠區的週邊,而其在花蓮的分 布則局限在吉安溪、台九線、木瓜溪與花蓮海 岸間,包括吉安鄉台九線以東的永興村、稻香 村、仁和村、仁安村與干城村域內(吳等 2001)。 2004 與 2005 年的監測調查則發現,緬甸小鼠 的分布已擴展至吉安鄉台九線以西與壽豐鄉木 瓜溪以南一帶(吳海音,未發表資料)。 在分子生態學的相關研究中,DNA 序列 分析是目前常用的技術。大多數生物粒線體 DNA 具有母系遺傳的特性,且單一細胞內的 分子套數為核DNA 的 500-1,000 倍。配合 PCR 技術的使用,粒線體DNA 序列成為研究族群 結構、類緣關係、生物地理、生態保育等領 域 最 適 用 的 遺 傳 標 幟 之 一(Avise and Walker 1998; Emerson 2002; Wan et al. 2004; Ballard and Rand 2005)。雖然目前取得 DNA 序列不 再那樣昂貴和耗時,但這仍是大部分研究中 的一個限制步驟。在採用DNA 序列分析法來 研究族群遺傳結構時,常需要較多的樣本數, 然而在這種研究尺度下,大部分的個體可能 攜帶相同的 DNA 序列,逐一定序的花費不 貲。此時即可應用DNA 單股構造多態性(single-strand conformation polymorphism, SSCP)此一 簡單、便宜、且可靈敏的偵測出不同序列的 技術,以有效減少需要定序的樣本數量(Sunnucks
et al. 2000)。
簡 單 來 說,SSCP 利 用 聚 丙 烯 醯 胺 凝 膠 (polyacrylamide gel, PAGE)電泳,區分因鹼基 序列不同而三級摺疊結構相異的單股 DNA, 使 之 在 膠 體 上 呈 現 不 同 的 片 段 模 式(Hayashi 1991)。將 SSCP 應用於檢測 PCR 產物的序列 變異,稱為PCR-SSCP 分析,能進一步提升檢 測的簡便性和靈敏性。PCR-SSCP 的敏感性與 分析片段的長度大小成反比(Girman 1996),雖 然過去研究結果顯示,SSCP 可區分出序列長 度達775 個鹼基之單股 DNA 小分子上的單一 鹼基變異(Orti et al. 1997),但一般多應用於區 分長度小於400 個鹼基的單股 DNA 小分子上。 配合一般族群層次之分子生態研究選擇的分子 標幟多為變異程度大的短 DNA 片段來看, PCR-SSCP 可說是極適當的應用技術。 吳 等(2001)曾 利 用 捕 獲 到 緬 甸 小 鼠 的 組 織,進行粒線體 DNA 控制區(control region) 276 個 鹼 基 序 列 分 析,並 與 NCBI (National Center for Biotechnology Information)核酸序列 資料庫中的資料比對,結果除了肯定早先其他 學者利用形態所做的分類鑑定(Motokawa et al. 2001),更在所定序的 35 隻個體中歸類出 7 種 基因型,且最接近NCBI 資料庫中採集自紐西 蘭地區樣本的序列。由於該研究中所定序的樣 本數量有限,且當時NCBI 資料庫與相關文獻 中可供比對的序列樣本多採樣於紐西蘭及大洋 洲各島嶼(Matisoo-Smith et al. 1998),缺乏東 南亞大陸及週邊島嶼區域的樣本,因此比較後 所得之結果有限。本研究延續之前的研究,使 用過往採集到的緬甸小鼠樣本,以PCR-SSCP 技術普查粒線體DNA 控制區域單倍基因型有 多少種片段模式,針對不同的片段模式進行定 序,並與NCBI 序列資料庫中晚近增添之序列 比對分析,以探尋台灣緬甸小鼠的可能來源。 此外,將進一步分析採樣區域粒線體DNA 基 因型的歧異度及分布狀況,藉以探討此外來族 群侵入台灣後的擴散模式。
材料與方法
一、研究材料 研究材料為2001 年及 2003 年間,在花蓮 縣吉安鄉進行緬甸小鼠捕捉監測時所捕獲的樣 本。捕捉採樣的方法是先將樣區劃分為 95 個 500m 500m 的方格區塊,2001 年時在每一個 方格內選擇一塊草生地設置捕捉樣站,扣除中 華紙槳廠與光榮砂石專業區等無法設置樣站的 方格外,共計 91 處草地樣站,另於樣區內選 擇10 處的樹林樣站;2003 年時在各方格內選 擇一樹林或草生地設置樣站,共計 92 處。捕 捉樣站的設置參考Bowman et al. (2001),在選 取的樣站中心,以十字形放置薛門氏鼠籠(Sher-man s Live Trap, 26 cm 8 cm 10 cm) 5 個,每 站進行1 次連續 3 晚的捕捉。2003 年中,在對 全樣區系統取樣捕捉後,另對捕獲量較高的兩 方格區塊及中華紙漿廠廠區分別進行 110 與 195 籠次的捕捉。兩年度的捕捉努力量分別為 1,515 與 1,685 籠次,兩年度的捕捉個體各為 100 及 80 隻,共計 180 隻。對捕獲的鼠隻樣 本,依吳等(2003)建立的成幼體判別標準,將 體重在25g 以上的雄鼠,22g 以上的雌鼠判為 成體。捕捉到的個體取肌肉組織樣本,以酒精 及STE 溶液保存,以備 DNA 萃取之用。 二、粒線體DNA 控制區域基因型分析 本研究將採集到的緬甸小鼠組織樣本均質 後萃取粗 DNA,並根據先前研究所定之粒線 體DNA 控制區域序列(吳等 2001)設計 PCR 引 子組,以LR-SSCP (5 -CCCCAAGCATATAAG-CATGT-3 ) 及 HR-SSCP (5 -CGTCCCTCTTA-AATAAG ACATCTCG-3 )進行 PCR,增幅出長 度為372 個鹼基對的 PCR 片段。PCR 產物經 鹼解後,以濃度8 %的 PAGE 進行 13-16 hr 之 單股 DNA 片段低溫直立式電泳。電泳後的 PAGE 膠體以銀染法呈現 SSCP 片段模式,並 將膠體乾燥保存,以供資料判別。將不同的 SSCP 片段模式判定為不同的粒線體 DNA 控制 區域單倍基因型,對出現個體數目較多的基因 型,隨機挑選15 - 20%的個體,而出現個體數 目較少的基因型,則選取全數個體,利用前述 的引子組對這些個體的基因型進行定序,以檢 視相同基因型的序列是否相同,藉此確認SSCP 結果的可信度。以MEGA 3.1 軟體(Kumar et al. 2003)計算 各基因型序列(長度為 372 個鹼基對)間的鹼基 差異數目及遺傳距離(Tamura-Nei Distance)。 為判別台灣外來種緬甸小鼠的可能來源,將本 研究所定序出的粒線體DNA 控制區域序列與 NCBI 資料庫中 Matisoo-Smith and Robins (2004) 發表的 32 種緬甸小鼠序列(R1-R25 及 R27-R33,AY604202-AY604233),經序列排列比對 (alignment)後,選取落在粒線體 DNA 控制區 域高變異區(hyper variable region I, HVR I)內 長度為174 個鹼基對的序列,利用 MEGA 3.1 軟體以鄰聚法(neighbor-joining method)建構關 係樹型圖,並以 Boostrap (1,000 次)檢驗分群 的可信度,以觀察台灣外來種緬甸小鼠與這些 序列間的相似程度,藉此推測其來源地。另 外,依照Matisoo-Smith and Robins (2004)對這 些基因型區分出的分群模式,包括東南亞群 (Southeast Asian Haplogroup)、東南亞及近大 洋洲群(Southeast Asian & Near Oceanic Haplo-group)、遠大洋洲群(Remote Oceanic Haplogroup) 及僅有一基因型的泰國分支,並將台灣的緬甸 小鼠視為一群,計算5 組資料的群內及群間平 均鹼基差異數目及遺傳距離,以比對目前台灣 外來緬甸小鼠的遺傳變異程度。 三、粒線體DNA 控制區域基因型的空間分布 為檢視緬甸小鼠不同粒線體DNA 控制區 域基因型在研究區內的空間分布,以捕捉採樣 之方格系統的方格為單位,將兩年間自同一方 格不同樣站的捕捉資料加以合併,計算各方格 內不同基因型的頻度。以吳逸華(2003)修改姜
聖 華(2000)的 研 究 區 道 路 圖 層 為 底 圖,利 用 ArcView 3.0 將各方格內不同基因型的頻度分 布圓形圖套疊其上。
結
果
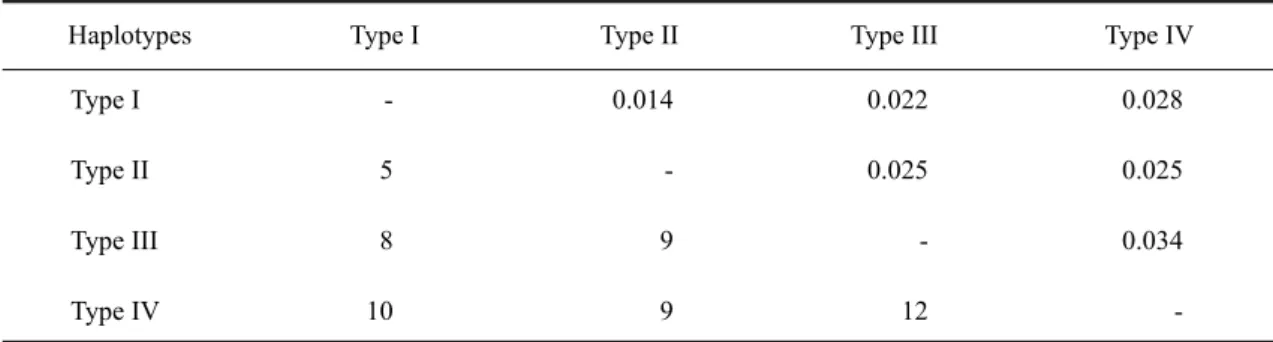
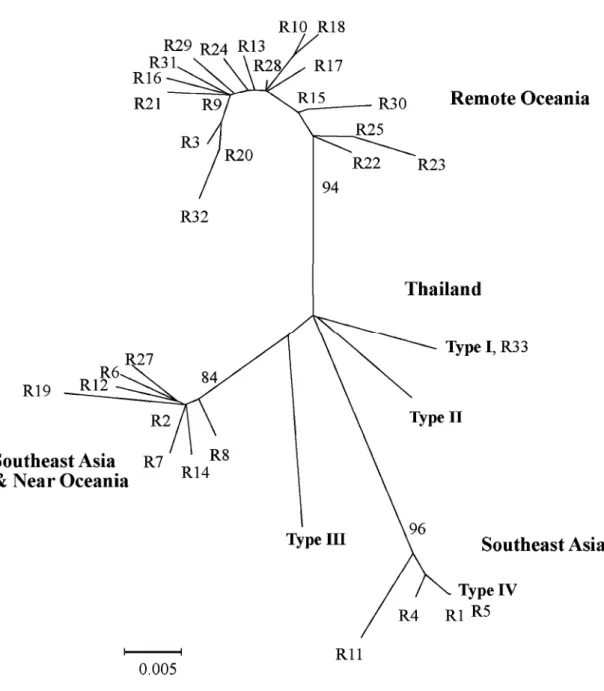
2001 年及 2003 年捕獲的 180 隻緬甸小鼠 中,包括雄鼠 102 隻,雌鼠 78 隻,雄鼠數量 略多於雌鼠,但性比未偏離1:1 (p = 0.074)。 其中雄性成鼠有99 隻,最大體重為 67g,平均 體重41.1g (SD = 10.1g),雌性成鼠有 68 隻, 最大體重為54g,平均體重 36g (SD = 6.8g), 成熟雄鼠的體重顯著大於成熟雌鼠(p < 0.01)。 以PCR-SSCP 分析 180 隻緬甸小鼠的粒線 體DNA 控制區域片段,共分辨出 4 種 SSCP 片 段模式,分別有123、53、2 及 2 隻樣本,取 其中的24、8、2 及 2 隻個體進行定序(共 36 隻 個體),經定序後確認為 4 種不同的粒線體 DNA 控制區域單倍基因型,且相同SSCP 片段模式 之樣本定序出的序列均相同。以發現的先後順 序將之命名為Type I、Type II、Type III 及 Type IV。在攜帶 Type I 基因型的 123 隻個體中有 74 隻雄鼠,49 隻雌鼠;攜帶 Type II 的 53 隻個體 中有25 隻雄鼠,28 隻雌鼠;攜帶 Type III 的 2 隻個體為 1 隻雄鼠及 1 隻雌鼠;攜帶 Type IV 的僅有2 隻雄鼠。由所定序的 372 個鹼基對來 看,各基因型間的鹼基差異數目介於5-12 個 鹼基對(表 1),其中 Type III 和 Type IV 間的差 異性最大(相差 12 個鹼基對),而 Type I 和 Type II 間的差異性最小(相差 5 個鹼基對)。4 種基 因型間以 Tamura-Nei 方法所計算出的遺傳距 離介於0.014-0.034 間(表 1)。 將本研究所發現的4 種基因型與 NCBI 資 料庫中的32 種緬甸小鼠粒線體 DNA 控制區域 單倍基因型重新建構樹型圖(圖 1)後發現,樹 型圖的主要分群結構分為東南亞群、東南亞及 近大洋洲群及遠大洋洲群3 個有效分群(Boot-strap=84-96%)與泰國分支。在台灣緬甸小鼠的 4 個基因型中,Type I 與採樣自泰國的緬甸小 鼠序列(R33)完全相同,Type IV 與採樣自蘇拉 維西的小鼠序列(R5)僅相差 1 個鹼基對,而 Type II 及 III 則各自成單獨分支,但與東南亞 區域的樣本序列較為接近。 表 1. 台灣地區緬甸小鼠粒線體 DNA 控制區域 4 種單倍基因型間的鹼基差異數目(左下)及以 Tamura -Nei 方法計算出來之遺傳距離(右上)Table 1. Nucleotide differences (below diagonal) and Tamura-Nei genetic distances (above diagonal)
among four mtDNA haplotypes (Types I-IV) of Rattus exulans in Taiwan
Haplotypes Type I Type II Type III Type IV Type I Type II Type III Type IV -5 8 10 0.014 -9 9 0.022 0.025 -12 0.028 0.025 0.034
-圖 1. 緬甸小鼠(Rattus exulans)粒線體 DNA 控制區域 4 種單倍基因型(Types I-IV),與 NCBI 核酸序 列資料庫中32 種緬甸小鼠基因型(R1-R25 與 R27-R33)間以鄰聚法(neighbor-joining method)建構之 親緣關係圖。
Fig. 1. Phylogenetic relationships expressed by a neighbor-joining network among four mtDNA haplotypes
(Types I-IV) of Rattus exulans from Hualien, Taiwan and 32 haplotypes of those reported from Southeast Asia, Southeast Asia and Near Oceania, and Remote Oceania by NCBI (R1-R25 and R27-R33).
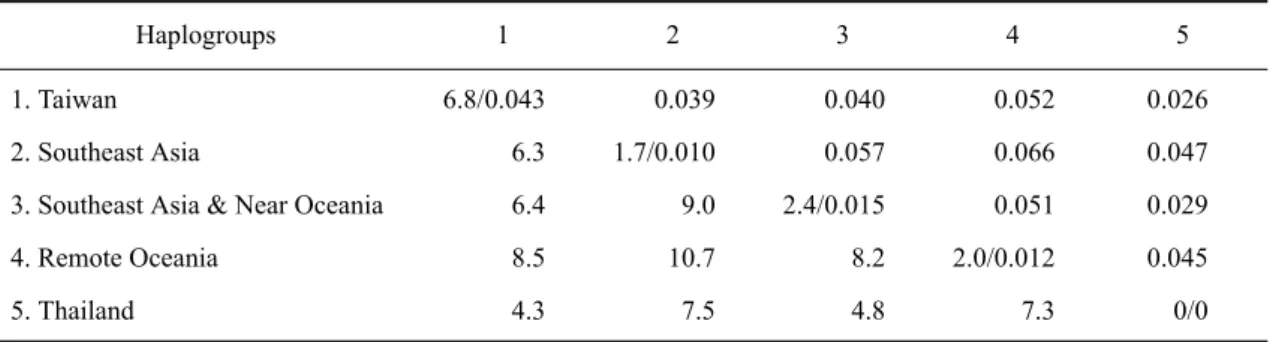
表 2. 台灣地區緬甸小鼠與 Matisoo-Smith and Robins (2004) 提出之緬甸小鼠粒線體 DNA 基因型分 群間,群內與群間的平均鹼基差異(左下) 及以 Tamura-Nei 方法計算出的遺傳距離 (右上)
Table 2. Mean nucleotide differences (below diagonal) and Tamura-Nei genetic distances (above diagonal
) of mtDNA haplogroup of Taiwan and those from Southeast Asia, Southeast Asia and Near Oceania, Remote Oceania, and Thailand proposed by Matisoo-Smith and Robins (2004)
Haplogroups 1 2 3 4 5
1. Taiwan 2. Southeast Asia
3. Southeast Asia & Near Oceania 4. Remote Oceania 5. Thailand 6.8/0.043 6.3 6.4 8.5 4.3 0.039 1.7/0.010 9.0 10.7 7.5 0.040 0.057 2.4/0.015 8.2 4.8 0.052 0.066 0.051 2.0/0.012 7.3 0.026 0.047 0.029 0.045 0/0
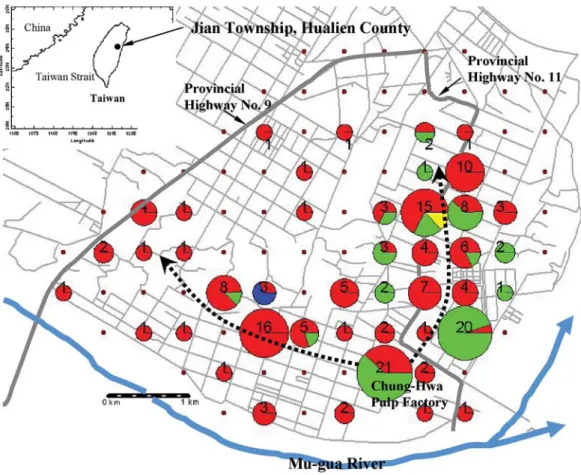
計算Matisoo-Smith and Robins (2004)的基 因型分群(東南亞群、東南亞與近大洋洲群、 遠大洋洲群及泰國),及台灣緬甸小鼠群 5 組 資料的群間及群內平均鹼基對差異數及遺傳距 離,結果顯示台灣緬甸小鼠4 個基因型的群內 平均鹼基差異數(6.8)與遺傳距離(0.043)遠大於 其他 3 群區域族群群內的差異(鹼基差異數, 1.7-2.4;遺傳距離,0.010-0.015),而與這 3 個 區域族群群間的差異(鹼基差異數,8.2-10.7; 遺傳距離,0.051-0.066)較相似(表 2)。 檢視小鼠各基因型在樣區的空間分布,可 發現數量最多之基因型Type I 和 Type II 的分 布也最廣(圖 2)。在總數 95 個採樣方格內,共 有44 個方格內捕捉到緬甸小鼠,而 Type I 及 Type II 分別出現於 40 及 14 個方格,且兩者的 分布具重疊性,同時出現在 10 個方格中。若 以中華紙漿廠為中心,則鄰近紙漿廠的採樣點 多為兩基因型分布重疊區,而距離紙漿廠較遠 的採樣點則多僅有Type I 的分布。數量最少的 Type III 和 Type IV 基因型,則各自局限在單 一方格,且與Type I 或 Type I、II 共存。
討
論
由台灣及NCBI 資料庫中緬甸小鼠序列比 對結果顯示,台灣外來種緬甸小鼠與東南亞大 陸以及鄰近大陸之島嶼區族群的類緣關係較為 接近(圖 1)。且台灣所發現的 4 種緬甸小鼠基 因型,其群內平均鹼基差異數與遺傳距離遠大 於其他3 群區域族群群內的差異,而與這 3 個 區域族群群間的差異較相似(表 2)。由以上結 果推測,目前在台灣所發現的緬甸小鼠,其來 源可能並不僅止於單一族群,而是來自數個區 域族群,或是來自一個擁有多重來源的混合族 群。由於台灣常自東南亞各國進口原木或造紙 用木料,因此緬甸小鼠可能是在運輸木材的過 程中經貨船挾帶進入台灣。 本研究發現攜帶基因型Type I 和 Type II 的小鼠隻數最多,且分布範圍相對來說較為廣 泛,因此我們推測攜帶Type I 和 Type II 之小 鼠可能為較早入侵台灣之族群,其入侵後能適 應台灣的棲地環境,因此逐步擴散範圍。由圖 2 可見攜帶基因型 Type II 小鼠的分布區域多與 Type I 重疊,顯示帶有兩種不同基因型的入侵 者在進入台灣後,具有類似的播遷模式。這同 時也顯示,攜帶這兩種基因型的小鼠可能侵入 台灣的時間點很接近,有可能Type I 先入侵, Type II 隨後入侵。但也不排除兩者同時入侵圖 2. 花蓮縣吉安鄉緬甸小鼠(Rattus exulans)粒線體 DNA 控制區域 4 種單倍基因型分布模式。圓形 圖為在每個方格捕獲緬甸小鼠樣本中不同基因型的頻度分布,紅色為基因型Type I、綠色為 Type II、深藍色為 Type III、黃色為 Type IV,數字代表各採樣區內的樣本數。未捕獲緬甸小鼠的方格 以棕色點表示。由基因型分布模式推論,緬甸小鼠可能由中華紙漿廠逐漸向北及向西擴散至紙漿 廠週邊(如虛線箭頭所示)。
Fig. 2. Distribution patterns of four mtDNA haplotypes (Types I-IV) of Rattus exulans in Jian Township,
Hualien County (pies, haplotypic frequencies of rats captured at each of the sampling locations; red, green, blue, and yellow, respectively, Types I, II, III and IV; dotted lines with arrow heads, hypothesized directions of range expansion of the rats from the Chung-Hwa Pulp Factory).
的可能,只是入侵的起始族群攜帶兩種基因型 的個體數量或入侵後的生殖成功率有差異,使 得 Type I 母系群族群成長較快而分布範圍較 Type II 母系群廣。此外,由兩種基因型間的 差異來看(兩基因型差 5 個鹼基對,遺傳距離 為0.014,表 1),攜帶這兩種基因型的小鼠有 可能皆來自東南亞鄰近泰國的區域。相對於 Type I 和 Type II 而言,攜帶基因型 Type III 和 Type IV 的小鼠個體數甚少(Type III & IV/Type I & II = 4 / 176),僅於 2001 年採集到,且兩
種基因型分布均局限於單一採樣點。行政院農 業委員會動植物防疫檢疫局為防治此外來種的 入侵,曾於 2002 年提撥經費在花蓮縣吉安鄉 進行滅鼠活動。經過此滅鼠活動後,2003 年 並未再採集到這兩種基因型,因此推測帶有這 兩種基因型的個體可能為較晚近才入侵的小族 群,族群尚未開始擴散即因滅鼠活動而降至極 少數甚或滅絕。 從基因型分布的模式來看,中華紙漿廠週 邊的住宅區或農地等採樣點,多為Type I 及 II 分布的重疊區,而在距離紙漿廠較遠之區域採 樣到的小鼠數量較少,基因型也減少為單一 型,且多為Type I (圖 2)。因此,我們推測中 華紙漿廠可能為攜帶這兩種基因型之緬甸小鼠 族群向外擴散的源頭。由於緬甸小鼠可能是在 運送木材的過程中被挾帶入台灣,推測紙漿廠 為小鼠族群擴散的源頭實屬合理。自基因型分 布的模式也可發現,小鼠族群可能自紙漿廠分 別向北及向西往紙漿廠週邊逐漸擴張,這兩個 擴散方向的棲地具有較高比例的雜林及長期休 耕的荒地,而雜林及荒地是緬甸小鼠較偏好的 棲地型態(吳 2003)。至於攜帶 Type III 及 Type IV 的緬甸小鼠由於個體數量太少,難以判定 兩種基因型的擴散來源是否即為其採樣點。 本研究採用PCR-SSCP 技術檢測台灣外來 種緬甸小鼠的遺傳結構,這個方法雖然簡易且 便宜,但亦有不足之處,如只能作為一種變異 檢測方法,要確定變異的位置和類型,還需進 一步定序,且電泳條件要求較嚴格,及當某些 位置的點突變對單鏈DNA 分子架構的改變不 起作用或作用很小時,有可能使PAGE 無法偵 測出基因型有差異。儘管如此,和其他方法相 比,此方法在分子生態研究的應用領域上仍具 優勢,能以低實驗耗費來普查大量樣本,並能 偵測出族群中大部分的變異(通常>90%) (Girman 1996; Miterski et al. 2000; Sunnucks et al. 2000)。 偵測序列變異的敏感性能因為修改實驗上的一 些細節而改善,必要時,可藉由改變溫度、膠
體組成、電泳緩衝溶液成分和跑膠狀態來改善 實驗結果(Hayashi 1991; Hayashi and Yandell 1993; Liu et al. 1999; Nataraj et al. 1999; Yip et
al. 1999; Sunnucks et al. 2000)。當實驗結果的
敏感度提高後,SSCP 甚至可應用來重複檢驗 定序結果的正確性。吳等(2001)依核酸定序後 所發表的7 種緬甸小鼠粒線體基因型(Type-1, Type-1 ,Type-1 ,Type-2,Type-3,Type-3 及 Type-4),經 SSCP 重複檢驗後發現這 7 種基因 型僅具有3 種 SSCP 片段模式。經重複定序當 時被判定為攜帶這些基因型的緬甸小鼠樣本 後,發現有4 種為電腦判別定序鹼基錯誤而 誤 判 的 基 因 型(Type-1 ,Type-1 ,Type-3 , Type-4),其中 Type-1 及 Type-1 與 Type-1 相 同,Type-3 與 Type-3 相同,而 Type-4 與 Type-2 相同。由上述情況可知,核酸定序未經多次的 重複確認,結果不必然正確。但是敏感度高的 SSCP 技術,在判別大量樣本中具有幾種不同 基因型上,錯誤率極低,例如在本研究中並未 發現相同的SSCP 片段模式具有不同的鹼基序 列。因此在族群尺度的分子生態研究上,PCR-SSCP 可算是初步性普查的最佳技術,值得未 來研究推廣使用。
謝
誌
本研究承蒙農業委員會動植物防疫檢疫局 提供田野調查的研究經費,農業藥物毒物試驗 所、花蓮區農業改良廠及花蓮港疾病管制局等 單 位,提 供 協 助 與 指 導,及 國 科 會 計 畫 (NSC-94-2313-B-002-06、92-2621-B-002-01) 提供分子實驗之經費補助,在此一併致謝。引用文獻
吳海音、吳逸華、儲瑞華、林曜松。2001。緬 甸 小鼠 在 台灣 之 發 生與 防 治。植 保會 刊 43: 205-214。吳海音、盧高宏、徐保雄、吳逸華。2003。緬 甸小鼠之防治。入侵種生物管理研討會論 文集。267-274 頁。 吳逸華。2003。外來種緬甸小鼠於兩種尺度中 的棲地利用。國立東華大學自然資源管理 研究所碩士論文。 姜聖 華。2000。花蓮地區都市擴張行為之研 究。國立東華大學自然資源管理研究所碩 士論文。 陳 保 良、高 清 文。2001。台灣 地 區 野鼠 之 防 治。農政與農情105: 60-62。 盧 高 宏、徐 保 雄。2003。緬 甸 小 鼠(Rattus exulans Peale)之棲群分布。植保會刊 45: 163-167。
Avise, J. C. and D. Walker. 1998. Pleistocene phy-logeographic effects on avian populations and the speciation process. Proceedings of the Royal Society of London Series B-Biological Sciences 265: 457-463.
Ballard, J. W. O. and D. M. Rand. 2005. The population biology of mitochondrial DNA and its phylogenetic implications. Annual Review of Ecology Evolution and Systematics 36: 621-642.
Bowman, J., C. V. Corkum and G. J. Forbes. 2001. Spatial scales of trapping in small-mammal research. Canadian Field-Naturalist 115: 472-475. Corbet, G. B. and J. E. Hill. 1992. The mammals of the Indomalayan region. Oxford University Press, New York.
Emerson, B. C. 2002. Evolution on oceanic islands: Molecular phylogenetic approaches to under-standing pattern and process. Molecular Ecology 11: 951-966.
Girman, D. 1996. The use of PCR-based single-stranded conformation polymorphism analysis (SSCP-PCR) in conservation genetics. pp. 167-182. In: Smith, T. B. and R. K. Wayne
(eds.). Molecular Genetic Approaches in Conservation. Oxford University Press, Oxford. Hayashi, K. 1991. PCR-SSCP: A simple and sensitive method for detection of mutation in the genomic DNA. PCR Methods and Applications 1: 34-38.
Hayashi, K. and D. W. Yandell. 1993. How sensitive is PCR-SSCP?. Human Mutation 2: 338-346. Kumar, S., K. Tamura and M. Nei. 2003. MEGA3: An integrated software for molecular evolutionary genetic analysis and sequence alignment. Integrative and Comparative Biology 43: 947-947.
Lekagul, B. and J. A. McNeely. 1988. Mammals of Thailand. Darnsutha Press, Bangkok. Liu, Q., J. Feng, C. Buzin, C. Wen, G. Nozari, A.
Mengos, V. Nguyen, J. Liu, L. Crawford, F. K. Fujimura and S. S. Sommer. 1999. Detection of virtually all mutations-SSCP (DOVAM-S): A rapid method for mutation scanning with virtually 100% sensitivity. Biotechniques 26: 932.
Matisoo-Smith, E., R. M. Roberts, G. J. Irwin, J. S. Allen, D. Penny and D. M. Lambert. 1998. Patterns of prehistoric human mobility in Polynesia indicated by mtDNA from the Pacific rat. Proceedings of the National Academy of Sciences of the United States of America 95: 15145-15150.
Matisoo-Smith, E. and J. H. Robins. 2004. Origins and dispersals of Pacific peoples: Evidence from mtDNA phylogenies of the Pacific rat. Proceedings of the National Academy of Sciences of the United States of America 101: 9167-9172.
Miterski, B., R. Kruger, P. Wintermeyer and J. T. Epplen. 2000. PCR/SSCP detects reliably and efficiently DNA sequence variations in
large scale screening projects. Combinatorial Chemistry & High Throughput Screening 3: 211-218.
Motokawa, M., K. H. Lu, M. Harada and L. K. Lin. 2001. New records of the Polynesian rat
Rattus exulans (Mammalia : Rodentia) from
Taiwan and the Ryukyus. Zoological Studies 40: 299-304.
Musser, G. G. and C. Newcomb. 1983. Malaysian murids and the giant rat of Sumatra. Bulletin of the American Museum of Natural History 174: 327-598.
Nataraj, A. J., I. Olivos-Glander, N. Kusukawa and W. E. Highsmith. 1999. Single-strand conformation polymorphism and heteroduplex analysis for gel-based mutation detection. Electrophoresis 20: 1177-1185.
Orti, G., M. P. Hare and J. C. Avise. 1997. Detection and isolation of nuclear haplotypes by PCR-SSCP. Molecular Ecology 6: 575-580. Sunnucks, P., A. C. C. Wilson, L. B. Beheregaray,
K. Zenger, J. French and A. C. Taylor. 2000. SSCP is not so difficult: The application and utility of single-stranded conformation poly-morphism in evolutionary biology and molecular ecology. Molecular Ecology 9: 1699-1710.
Tobin, M. E. 1994. Polynesian rats. pp.121-124.
In: Hygnstrom, S. E., R. M. Timm and G. E.
Larson (eds.). Prevention and control of wildlife damage. University of Nebraska, Lincoln.
Wan, Q. H., H. Wu, T. Fujihara and S. G. Fang. 2004. Which genetic marker for which con-servation genetics issue?. Electrophoresis 25: 2165-2176.
Wirtz, W. O. I. 1972. Population ecology of the Polynesian rat, Rattus exulans, on Kure Atoll,
Hawaii. Pacific Science 26: 433-464. Wirtz, W. O. I. 1973. Growth and development
of Rattus exulans. Journal of Mammalogy 54: 189-202.
Wodzicki, K. and R. H. Taylor. 1984. Distribution and status of the Polynesian rat Rattus exulans. Acta Zoolgica Fennica 172: 99-101. Yip, S. P., D. A. Hopkinson and D. B. Whitehouse.
1999. Improvement of SSCP analysis by use of denaturants. Biotechniques 27: 20-24.