中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/41258
129
0
0
全文
(2) i.
(3) 中文摘要 在動脈硬化發展過程中,血管平滑肌細胞移行到內層,並且在內 膜增生、堆積,對於動脈硬化早期的發生與血管再狹窄的形成非常重 要。基質金屬蛋白酶(Matrix Metalloproteinases;MMPs)的大量表 現,亦參與心血管疾病的致病過程。其中第Ⅳ型膠原酶或明膠蛋白酶 (MMP-2 和 MMP-9)對於動脈病變的發展經由調節血管平滑肌細胞 遷移和增殖是重要的。Inotilone 為桑黃(Phellinus linteus )的主要成 分之一,桑黃(Phellinus linteus)在中國,韓國,日本和其他亞洲國 家被用來作為一個傳統的藥用真菌;文獻證實桑黃具有抗菌,抗腫瘤, 抗纖維化,抗突變,抗氧化及抗發炎的功效,以及能刺激體液免疫和 細胞免疫。 本篇論文擬對探討桑黃乙酸乙酯層與 Inotilone 抑制腫瘤壞死因 子(TNF-α)誘導人類主動脈平滑肌細胞增生、移行與侵襲的表現。 在本研究中使用 MTT assay 測人類主動脈平滑肌細胞在桑黃乙酸乙 酯層與 Inotilone 作用下的存活率;由細胞移行(cell migration)及細 胞侵襲(cell invasion)試驗可看到桑黃乙酸乙酯層與 Inotilone 對於 TNF-α 所誘導的人類主動脈平滑肌細胞的移行及侵襲能力有抑制效 i.
(4) 果;為了解釋這個抑制效果,利用明膠蛋白酵素電泳法及西方點墨法 檢測桑黃乙酸乙酯層與 Inotilone 的蛋白表現作用;結果顯示利用 100 ng/mL TNF-α 處理人類主動脈平滑肌細胞後,會促使 MMP-2、 MMP-9 及磷酸化的 FAK、ERK、JNK、 p38 和 IκB 增加,但會降 低 TIMP-1 及 TIMP-2 的表現,而在桑黃乙酸乙酯層與 Inotilone 處 理後,則能減少 MMP-2、MMP-9 的表現及 FAK、ERK、JNK、p38 和 IκB 的磷酸化,增加 TIMP-1 及 TIMP-2 的表現。由以上結果可 知桑黃乙酸乙酯層與 Inotilone 有效地抑制 TNF-α 所誘導的人類主 動脈平滑肌細胞增生與移行的表現,具有預防血管疾病的潛力。. ii.
(5) Abstract The migration of vascular smooth muscle cells (VSMC) from the tunica media to the subendothelial region is a key event in the development of atherosclerosis and restenosis. The increased expression and activities of matrix metalloproteinases (MMPs) have been implicated in the pathogenesis of cardiovascular disease. Type Ⅳ collagenases or gelatinases (MMP-2 and MMP-9) are critical for the development of arterial lesions via its regulation of both VSMC migration and proliferation. Inotilone is a major component of Phellinus Linteus (PL) which has been used as a traditional medicinal mushroom in China, Korea, Japan and other Asian countries. Several studies demonstrated to exhibit. anti-bacterial,. anti-tumour,. anti-fibrotic,. antimutagenic,. anti-oxidant and anti-inflammatory functions, as well as stimulating humoral and cell-mediated immunity in several studies. In this study, we investigated the effects of the ethyl acetate layer from PL (PLEA) and inotilone, an active component of PL, on tumor necrosis factor-α (TNF-α)-induced cell proliferation and migration in iii.
(6) human aortic smooth muscle cells (HASMCs). The cytotoxicity of PLEA and inotilone on HASMCs was measured by the MTT assay method. The migration and Invasion assay showed that PLEA and Inotilone effectively inhibited TNF-α-induced migration and invasion of HASMCs as compared with the control group in a dose-dependent manner. To explain this inhibitory effect, PLEA and Inotilone were assayed by gelatin zymography and Western blot. In present study, HASMCs treated with 100 ng/mL TNF-α were found to increase expression of MMP-2, MMP-9, and. phosphorylation of FAK, ERK, JNK, p38, and IκB, but decreased. TIMP-1 and TIMP-2. Treatment of HASMCs with PLEA and inotilone attenuated TNF-α-induced the expression of MMP-2, MMP-9, and phosphorylation of FAK, ERK, JNK, p38, and IκB, but enhanced TIMP-1 and TIMP-2. These results suggest that PLEA and inotilone might effectively suppress TNF-α-induced HASMC migration and proliferation and represent potential agents for the prevention of vascular disorders.. iv.
(7) 致謝 時光飛逝,兩年的研究所生活即將結束,在這不算短的研究生活 中,受到許多人的指導與協助,讓我能夠順利的完成學業。 首先,誠摯的感謝指導教授黃冠中老師,在這兩年內悉心的教導, 使我得以一窺研究領域的深奧,培養學生獨立思考的能力,使研究路 途上能夠獨當一面,不時的討論並指點我正確的方向,並得以順利完 成論文;在課業上、生活上給我的指導及鼓勵,對我的包容、關心, 讓我在這兩年中獲益良多,由衷的感謝。 論文初成,承蒙中興大學胡淼琳教授及本校吳介信教授於百忙中 擔任我的口試委員,撥冗審閱批註,並對學生論文不厭其煩地詳加指 正,提供寶貴意見,才使本研究論文更臻完善,在此獻上衷心謝意。 研究的路途雖然艱辛但並不孤單,感謝香秀學姐、富擎學長及典 加學姐熱情大方的指導,因為有你們的傾囊相授,讓我減少許多實驗 中可能錯誤的嘗試,你們的衷心叮嚀與貼心關懷讓我倍感窩心;感謝 翔文、小ju及勳勳在我心情低落時,能幫我宣洩壓力,排解苦悶,這 段參雜喜怒哀樂的日子是最難忘的回憶;感謝老勇、小葳、思穎、阿 慶、拉其及阿嘉學長,有了你們的陪伴、鼓勵與關心,帶給實驗室許 v.
(8) 多歡樂的氣氛,讓我的研究生活變得炫麗多彩;此外,感謝老詹,因 為我們住在隔壁,不管遇到開不開心的事情,總是第一時間與妳分享; 同時我還要感謝一位好友創宇,你的支持與包容一直是我前進的原動 力。因為有你們讓我這兩年渡過快樂美好的研究生活。 最後我要感謝我最偉大的父母親,二十多年來辛勞教養及無怨無 悔的付出,在這兩年中,對我的鼓勵、安慰及支持,讓我有勇氣面對 所有的挑戰,做為我最有力的後盾,沒有你們就沒有今天的成就;謝 謝哥哥和姐姐,你們的體諒讓我每次回家就可以洗去一身的疲憊;謝 謝爺爺和奶奶,在學習的過程中給我最大的支持與鼓勵;因為有你們 的付出與用心才有今天的我,你們是最棒的家人! 感謝我的師長們、家人、朋友及所有曾經幫助過我的人,願與你 們分享我這份成就及喜悅。. 賴景揚 謹誌 於 中國醫藥大學 中國藥學暨中藥資源學系碩士班 中華民國一百年七月. vi.
(9) 目錄 中文摘要 .................................................................................................. i Abstract ................................................................................................. iii 致謝 ......................................................................................................... v 目錄 .......................................................................................................vii 圖目錄 .................................................................................................... xi 表目錄 ................................................................................................... xv 縮寫表 .................................................................................................. xvi 第一章. 研究背景及文獻回顧 ............................................................. 1. 第一節. 前言 ................................................................................. 1. 第二節. 動脈硬化之成因 .............................................................. 2. 第三節. 平滑肌細胞生理變化與動脈硬化的相關性 ................... 4. 第四節. TNF-α 與動脈硬化的相關性 .......................................... 5. 第五節. 基質金屬蛋白酶與動脈硬化的相關性 ........................... 7. 第六節. Mitogen-Activated Protein Kinases (MAPK)與動脈硬. 化之相關性 ................................................................................... 11 第七節. 基質金屬蛋白酶組織抑制劑......................................... 14 vii.
(10) 第八節. 桑黃與 Inotilone 之簡介 ................................................ 16. 第二章. 研究目的 .............................................................................. 18. 第三章. 研究方法 .............................................................................. 19. 第一節. 論文實驗架構 ................................................................ 19. 第二節. 研究材料 ........................................................................ 20. 一、 藥品與試劑 ................................................................ 20 二、 儀器設備與器材 ......................................................... 24 第三節. 實驗方法 ........................................................................ 25. 一、 細胞培養 .................................................................... 25 二、 細胞存活率試驗(Cell viability assay) ................... 28 三、 明膠蛋白酵素電泳法(Gelatin zymography) ......... 29 四、 細胞移行試驗(Cell migration assay) ..................... 32 五、 細胞侵襲試驗(Cell invasion assay) ....................... 32 六、 西方點墨法(Western blotting) ............................... 34 七、 統計分析(Statistic analysis):................................. 39 第四章. 實驗結果 .............................................................................. 40. 第一節. 桑黃乙酸乙酯層對人類主動脈平滑肌細胞毒性及存活. 率之影響 ....................................................................................... 40 viii.
(11) 第二節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞表現 MMP-9 活性之影響 ....................................................... 41 第三節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞移行能力的影響 ........................................................................ 42 第四節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞侵襲能力的影響 ........................................................................ 44 第七節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞分泌 FAK 蛋白質表現量之影響 ............................................ 50 第八節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞 MAPK 訊息傳遞路徑之影響 ................................................. 51 第九節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌細. 胞分泌 IκB 蛋白質表現量之影響 .............................................. 53 第十節. Inotilone 對人類主動脈平滑肌細胞存活率之影響 ..... 54. 第十一節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞表現 MMP-9 活性之影響 ..................................................................... 55 第十二節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞移行 能力的影響 ................................................................................... 56 第十三節 Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞侵襲能 ix.
(12) 力的影響 ....................................................................................... 58 第十四節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞分泌 MMP-2 及 TIMP-2 蛋白質表現量之影響 ................................. 60 第十五節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞分泌 MMP-9 及 TIMP-1 蛋白質表現量之影響 ................................. 61 第十六節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞分泌 FAK 蛋白質表現量之影響 .......................................................... 62 第十七節 ............ Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞 MAPK 訊息傳遞路徑之影響....................................................... 63 第十八節 ... Inotilone 對 TNF-α 刺激人類主動脈平滑肌細胞分泌 IκB 蛋白質表現量之影響 ............................................................ 64 第五章. 討論 ...................................................................................... 65. 圖表 ....................................................................................................... 73 參考文獻 ............................................................................................... 95 附錄 ..................................................................................................... 107. x.
(13) 圖目錄 Figure 1.. Inotilone 化學結構圖 .......................................................... 17. Figure 2.. 論文實驗架構圖 ................................................................. 19. Figure 3.. Cytotoxicity of ethyl acetate layer from Phellinus linteus. (PLEA) on human aortic smooth muscle cells (HASMCs). .................... 73 Figure 4.. Effect of ethyl acetate layer from Phellinus linteus (PLEA) on. the MMP-9 activity of TNF-α-induced human aortic smooth muscle cells (HASMCs). ............................................................................................ 74 Figure 5.. Effect of ethyl acetate layer from Phellinus linteus (PLEA) on. TNF-α-induced transwell migration of human aortic smooth muscle cells (HASMCs). ............................................................................................ 75 Figure 6.. Effect of ethyl acetate layer from Phellinus linteus (PLEA) on. TNF-α-induced Matrigel invasion of human aortic smooth muscle cells (HASMCs). ............................................................................................ 76 Figure 7.. Effect of ethyl acetate layer from Phellinus linteus (PLEA) on. TNF-α-induced protein expression of MMP-2 and TIMP-2. .................. 77 Figure 8.. Effect of ethyl acetate layer from Phellinus linteus (PLEA) on xi.
(14) TNF-α-induced protein expression of MMP-9 and TIMP-1. .................. 78 Figure 9.. Ethyl acetate layer from Phellinus linteus (PLEA) inhibits the. protein expression of FAK in human aortic smooth muscle cells (HASMCs). ............................................................................................ 79 Figure 10.. Ethyl acetate layer from Phellinus linteus (PLEA) inhibits. the protein expression of ERK in human aortic smooth muscle cells (HASMCs). ............................................................................................ 80 Figure 11.. Ethyl acetate layer from Phellinus linteus (PLEA) inhibits. the protein expression of JNK in human aortic smooth muscle cells (HASMCs). ............................................................................................ 81 Figure 12.. Ethyl acetate layer from Phellinus linteus (PLEA) inhibits. the protein expression of p38 in human aortic smooth muscle cells (HASMCs). ............................................................................................ 82 Figure 13.. Ethyl acetate layer from Phellinus linteus (PLEA) inhibits. the protein expression of IκB in human aortic smooth muscle cells (HASMCs). ............................................................................................ 83 Figure 14.. Cytotoxicity of inotilone on human aortic smooth muscle. cells (HASMCs)..................................................................................... 84 xii.
(15) Figure 15.. Effect of inotilone on the MMP-9 activity of TNF-α-induced. human aortic smooth muscle cells (HASMCs). ...................................... 85 Figure 16.. Effect of inotilone on TNF-α-induced transwell migration of. human aortic smooth muscle cells (HASMCs). ...................................... 86 Figure 17.. Effect of Inotilone on TNF-α-induced Matrigel invasion of. human aortic smooth muscle cells (HASMCs). ...................................... 87 Figure 18.. Effect of Inotilone on TNF-α-induced protein expression of. MMP-2 and TIMP-2. ............................................................................. 88 Figure 19.. Effect of Inotilone on TNF-α-induced protein expression of. MMP-9 and TIMP-1. ............................................................................. 89 Figure 20.. Inotilone inhibits the protein expression of FAK in human. aortic smooth muscle cells (HASMCs). ................................................. 90 Figure 21.. Inotilone inhibits the protein expression of ERK in human. aortic smooth muscle cells (HASMCs). ................................................. 91 Figure 22.. Inotilone inhibits the protein expression of JNK in human. aortic smooth muscle cells (HASMCs). ................................................. 92 Figure 23.. Inotilone inhibits the protein expression of p38 in human. aortic smooth muscle cells (HASMCs). ................................................. 93 xiii.
(16) Figure 24.. Inotilone inhibits the protein expression of IκB in human. aortic smooth muscle cells (HASMCs). ................................................. 94. xiv.
(17) 表目錄 Table 1.. 8 % SDS-PAGE 製備表 ........................................................ 30. Table 2.. 蛋白質檢量線配置表 ........................................................... 35. Table 3.. 10 % SDS-PAGE 製備表 ...................................................... 37. xv.
(18) 縮寫表 Ammonium persulfate:APS Bovine serum albumin:BSA Cyclooxygenase-2:COX-2 Dimethyl sulfoxide:DMSO Enchanced chemiluminescene:ECL Extracellular Matrix:ECM Ethyl acetate layer from Phellinus linteus:PLEA Extracellular signal-regulated kinase :ERK1/2 Fetal Bovine Seurm:FBS Focal adhesion kinase:FAK High-Density Lipoprotein:HDL Human aortic smooth muscle cell:HASMC Interleukin-1:IL-1 c-Jun NH2-terminal protein kinase: JNK Mitogen-actived protein kinase:MAPK Matrix Metalloproteinases:MMPs xvi.
(19) 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium. bromide:MTT. Matrix metalloproteinase-2:MMP-2 Matrix metalloproteinase-9:MMP-9 Nuclear factor kappa B:NF-κB Oxidized low density lipoprotein:OxLDL Penicillin/Streptomycin:PS Phosphate Buffered Saline:PBS Reactive oxygen species:ROS Sodium dodecyl sulfate:SDS Tissue inhibitors of matrix metalloproteinase-1:TIMP-1 Tissue inhibitors of matrix metalloproteinase-2:TIMP-2 Tumor necrosis factor-alpha:TNF-α. xvii.
(20) 第一章 研究背景及文獻回顧 第一節. 前言. 近年來,國人從農業社會逐步進入到高科技產業社會,生活型態 及飲食習慣皆有很大的改變,因此國人罹患心血管慢性疾病的比例日 益增加;粥狀動脈硬化(Atherosclerosis)是脂質與發炎細胞的聚積,. 伴隨著平滑肌細胞增生與細胞外間質液分泌所引起的細胞內膜纖 維變性(intimal fibrosis)的過程,最主要的特徵是動脈壁增厚或失 去彈性的病理變化,在許多已開發的國家中因為引起高度的心肌梗 塞、中風及周邊動脈疾病的致病率而成為主要的致死原因之一;根 據行政院衛生署公布民國九十八年十大死因資料顯示,腻血管與心. 臟疾病僅次於惡性腫瘤之後,分居於第二及第三名(附錄1);其 與生活型態、遺傳、慢性危險因子皆息息相關,動脈硬化也常伴隨腻 中風、心肌梗塞等症狀的產生,對現代人的健康造成相當大的威脅, 因此了解動脈硬化的形成與機轉及如何預防或治療,是一項未來急需 要研究及探討的重要課題之一。. 1.
(21) 第二節. 動脈硬化之成因. 動脈硬化是一種複雜的致病過程,其病理特徵大致上可分為五個 階段:1. 血管內膜增厚 (intimal thickening) ;2. 脂肪塊 (fatty streak); 3. 中度損傷(intermediate lesions) ;4. 纖維性斑塊(fibrous plaques) ; 5. 複雜性病灶(complicated lesions)[1]。高血壓、糖尿病、抽煙、 肥胖、高膽固醇血症、家族史、高油脂飲食等,是導致動脈硬化及冠 狀動脈硬化症等心臟疾病之主要危險因子,另外,血漿中膽固醇過高、 高密度脂蛋白(high-density lipoprotein; HDL)濃度低、低密度脂蛋 白(low-density lipoprotein; LDL)濃度高且被氧化修飾,皆會促使血 脂異常、血管內皮細胞功能損傷、發炎反應、血管平滑肌細胞增生移 行等現象。 當血管內皮細胞損傷或功能失調是形成動脈硬化的初期主因,會 使細胞產生急性和慢性發炎之代償反應做為保護,造成內皮細胞對脂 蛋白及血漿中的其他組成分子的通透性增加,使低密度脂蛋白容易移 入內皮細胞下間隙(subendothelial space)和內膜層(intima),在血 管內膜下空間堆積,並且會氧化產生氧化性低密度蛋白(oxidized low-density lipoprotein; oxLDL);此氧化低密度脂蛋白會刺激內皮細 胞或平滑肌細胞釋放出許多化學趨性物質,吸引血液中的單核球 (monocyte)進入內皮細胞下間隙,接著單核球活化分化成巨噬細胞 2.
(22) (macrophage) ,將氧化低密度脂蛋白吞噬而形成泡沫細胞(foam cell) 堆積在血管內層中,此作用會促使更多的單核球進入內皮層。泡沫細 胞會分泌許多促發炎細胞素(pro-inflammatory ctokines)、前列腺素 (prostaglandins; PGs )及生 長因子 , 如腫瘤壞 死因 子-α (tumor necrosis factor-α; TNF-α)、血小板衍生生長因子(platelet derived growth factor; PDGF)、介白素-1(interleukin-1; IL-1)和內毒素 (lipopolysaccharide; LPS)[2, 3]等,刺激血管中層的平滑肌細胞移行 (migration)至血管內皮層並增生(proliferation),這些堆積在血管 內層的分子交互作用及壞死,形成血管管壁纖維斑塊(plaque),使 血管肥厚,血管管徑變小,最後增大的斑塊破裂及血小板引發的血栓, 使得病灶更加惡化,造成血管管壁慢性的發炎反應或臨床急性心血管 疾病。. 3.
(23) 第三節. 平滑肌細胞生理變化與動脈硬化的相關性. 血管的組成可分為血管內層(intima):由單層扁平內皮細胞組 成、血管中層(media) :由平滑肌細胞組成、及血管外層(adventitia): 由纖維母細胞(fibroblast)和第一型膠原蛋白所組成。其中,血管中 層的平滑肌細胞為大型血管中的主要細胞;且平滑肌細胞的增生和移 行作用是造成血管內層增厚,使動脈硬化斑塊形成的主要原因之一。 平滑肌細胞具有收縮態(contractile)和合成態(synthetic)兩種 不同的表現型(phenotype)[4],血管在正常未受損狀態下,平滑肌 細胞保持在收縮態,細胞分裂表現非常低,且細胞內含有大量與收縮 相關的蛋白,如 smooth muscle α-actin等,其主要的功能在提供機械 力,維持血管壁彈性與強韌度、調控血壓;若受到血管收縮劑如 angiotention II 或 arginine vasopressin刺激會造成收縮[3]。 當血管損傷或病變時,平滑肌細胞轉變成活化的合成態,使動脈 管壁中膜層的平滑肌細胞移行至內膜層,並且大量增生,為動脈硬化 斑塊形成的重要原因之一;此合成態的平滑肌細胞內所含和收縮相關 的蛋白會減少,而細胞外基質蛋白(extracellular matrix protein)與生 長激素分泌均會增加,同時平滑肌細胞會促使細胞素(cytokine)及 黏附因子的表現[5],使平滑肌細胞呈現發炎狀態,造成病灶惡化。. 4.
(24) 第四節. TNF-α 與動脈硬化的相關性. 細胞激素和粥狀動脈硬化的慢性發展似乎具有協同作用,可以促 使粥狀動脈硬化斑塊及血栓的形成,而其內部中可發現大量的促發炎 激素(pro-inflammatory cytokine)堆積在其中,進而造成細胞增生、 移動及泡沫形成,促進血栓或纖維斑塊的形成,而產生動脈硬化惡化 的情形。 細胞激素具有兩種主要不同的方法去影響粥狀動脈硬化,一為促 進其形成:促發炎激素,另一則為抑制其形成:抗發炎激素 (anti-inflammatory cytokine),粥狀動脈硬化的形成受兩種激素間的 反應而影響。 IL-1 及 TNF-α 為促發炎激素[6];IL-1 會被單核球、巨噬細胞 和內皮細胞在不同刺激時所分泌,而雖然巨噬細胞的活化是 TNF-α 主要的來源,但也會經由各種不同的細胞產生,例如:纖維母細胞 (fibroblasts)、星狀細胞(astrocytes)、肝巨噬細胞(Kupffer cell)、 平滑肌細胞(smooth muscle cell)、角質細胞(keratinocyte)及各式 各樣的腫瘤細胞[7]。在細胞中發現 IL-1α、IL-1β 及 TNF-α皆可促進 內皮細胞及平滑肌細胞分泌黏著因子 [8] ,促進活性氧(reactive oxygen species;ROS)增加所帶來的自由基傷害;透過基因轉錄的方 式 TNF-α 可影響巨噬細胞清除者受體的表現[9]、促血管平滑肌細胞 5.
(25) (vascular smooth muscle cells;VSMCs)的生長[10],而 IL-1 則可 影響 LDL 的代謝、刺激平滑肌細胞增生及白血球吸附至內皮層上 [11],因此 TNF-α 及 IL-1 被認為是主要促使動脈硬化的發炎因子 之一。 發炎細胞產生的細胞激素經由不同的傳導路徑來影響作用細胞, 與細胞內氧化還原的狀態息息相關,其中活化核轉錄因子-Kappa B (nuclear factor-κB, NF-κB)是最易受到各種刺激誘發,進而調節細 胞增生、分化或者死亡。同時,也受到各種上游的激素調控,例如 NF-κB 的次單位與 IκB 結合,在細胞未受刺激時在細胞質中保持不 活化的狀態;但細胞受發炎因子刺激後,則促使 IκB 被磷酸化, NF-κB 游離出來進入細胞核內與 DNA 作用(附錄2),產生一連串 的發炎反應[7, 12]。. 6.
(26) 第五節. 基質金屬蛋白酶與動脈硬化的相關性. 動脈粥狀硬化的病理過程中,細胞外基質(extracellular matrix; ECM)的 分解扮 演重 要角色 [13, 14],細胞 外基質 包含膠 原蛋 白 (collagens) 、彈性蛋白(elastin)及多醣蛋白(proteoglycans)[15], 不同組織器官,各種組成成分比例不同;然而蛋白質過度的分解,造 成血管平滑肌細胞增生與移行及腫瘤細胞的入侵與轉移。因此血管平 滑肌細胞增生與移行時,會發現大量的蛋白水解酶伴隨產生,進而分 解細胞外基質,這些蛋白水解酶如:serine protease (plasmin, urokinase plasminogen activator; u-PA )、 cysteine protease ( cathepsin, matrix metalloproteinases; MMPs) 等。 在上述的蛋白水解酶中,又以基質金屬蛋白酶最為受到關注。基 質金屬蛋白酶家族為一群結構類似且需要鋅和鈣離子的蛋白分解酶, 參與各種正常生理過程,幾乎可分解所有細胞外基質,無論在正常或 非正常細胞中,皆扮演關鍵角色。適量的基質金屬蛋白酶可維持正常 的生理功能,如:傷口癒合、重建組織、骨骼生長、血管新生 (angiogenesis)等;但大量基質金屬蛋白酶的分泌也與許多疾病有 關,如:發炎現象、動脈硬化、腫瘤細胞的生長、侵襲及轉移、心血 管疾病等,在動脈硬化病理過程的不同階段會受到基質金屬蛋白酶的 調控[16-18]。 7.
(27) MMPs 家族酵素結構具有兩大特徵:(1) cysteine switch motif 及 (2) zinc-binding motif [19];但 MMP-23 為例外,因缺乏 cysteine motif, 其催化部位胺基酸序列與 MMP-1 相關,所以仍歸類為 MMPs 家族成 員之一。 MMPs 目前可分為六類(附錄 3): 1. Collagenases : 包 括 interstital collagenase ( MMP-1 )、 neutrophil collagenase (MMP-8) 、collagenase 3 (MMP-13) 及 collagenase 4 (MMP-18)為此類。這類具有在特殊位置 切斷 interstitial collagen I、II、III 的能力,形成氨基端 -3/4 與 羧基端 -1/4 的氨基酸片段,然後在生理的狀態下就會自然 分 解 [20] , 或 是 由 其 他 的 MMP 進 行 進 一 步 的 分 解 , collagenases 也可分解一些其他 ECM 以及非 ECM 分子。 2. Gelatinases : 包括 gelatinase-A(MMP-2)及 gelatinase B (MMP-9)。主要可分解 gelatin、collagen、laminin 等。 MMP-2 會分解第一型的膠原蛋白,還會活化 pro-MMP-9 [21];而 gelatin 是細胞外基質中最主要的組成物質之一,因 此 gelatinases 被視為是和動脈硬化最相關的基質金屬蛋白 酶[22]。 3. Stromelysins:包括 stromelysin-1、 procollagen(MMP-3) 、 8.
(28) stromelysin-2(MMP-10)及 stromelysin-3(MMP-11) 。其中 MMP-3 分解效率較高,除了分解細胞外基質之外,MMP-3 可以活化一些 pro-MMP,尤其對 MMP-1 活化非常重要。 4. Matrilysins:MMP-7 和 MMP-26 屬於此類。其特徵在於結構 缺乏 hemopexin。除了分解細胞外基質之外,MMP-7 還可以 分解一些細胞膜表面分子。 5. Membrane-Type MMPs:此類 MMP 位於細胞膜上,包括 type 1 transmembrane protein (MMP-14、MMP-15、MMP-16 和 MMP-24)以及 glycosyl phophatidyl inosited anchored protein (MMP-17 和 MMP-25) , MMP-14 具有分解 Type I、II、III collagen 的能力,且於血管新生作用扮演重要角色[23]。 6. Other MMPs : 包 括 一 些 尚 未 被 歸 類 或 是 最 近 才 發 現 : enamelysin (MMP-20)、XMMP(xenopus)(MMP-21)及 CMMP(chicken)(MMP-22)及 MMP-23。 MMPs 是由結締組織細胞所分泌,細胞初期所製造與分泌的 MMPs 皆為不具活性的型態(latent pro-form) ,必頇經過位於活性中 心的 Zn2+/Ca2+ 來活化,使 MMPs 具有分解細胞外基質的能力,而 MMPs 會以 zymogen. (inactive form)的方式被分泌出來,之後. zymogen 需要一些外來的蛋白水解酵素或是 organomercurials 來活 9.
(29) 化,活化後的 MMPs 其分子量大約會減少 10 kDa,因為 N 端的 propeptide domain 被切掉,去除其與活性中心之間的交互關係,達成 活化的目的;然而 MMPs 的活性可以被內生性的抑制因子 TIMPs (tissue inhibitor of metalloproteinases)所抑制。 MMPs 中又以 MMP-2 及 MMP-9 能夠分解血管基底膜 type IV collagen,與細胞的增生與移行最為相關。MMP-2 是一種在體內具有 持續性表現的基質金屬蛋白酶,能調控細胞的增生、移行及黏附作用, MMP-2 的活性會隨著年齡的老化而增加[24]。另外,MMP-9 是一種 誘發性的基質金屬蛋白酶,當 MMP-9 表現過量,會促使血管的平滑 肌細胞移行,並且在血管受損後,MMP-9 會使血管中層的平滑肌細 胞移行至內層,造成動脈硬化[25, 26]。. 10.
(30) 第六節. Mitogen-Activated Protein Kinases (MAPK). 與動脈硬化之相關性 當血管細胞受到損傷時,不論是活化的巨噬細胞、內皮細胞或血 管平滑肌細胞都會釋放各種生長因子,這些生長因子對於平滑肌細胞 的增生與移行都有某些程度的調控,而造成動脈硬化複雜的病理機制; 生長因子及細胞外間質蛋白等皆會藉由活化細胞膜上的受器,使細胞 外的訊息傳遞至細胞內,透過細胞內. kinases 及 磷 酸 化. (phosphorylation)作用活化訊息傳遞因子,進而產生反應,使細胞 存活下來或是走向死亡,其中 MAPKs(mitogen activated protein kinase pathway)就是一種訊息傳遞分子[27]。 MAPKs 路徑為傳遞外界訊息進入細胞內的重要途徑,此路徑在 真核細胞的訊息傳遞上扮演重要角色[28],同時參與細胞的生長、分 化、凋亡與轉移等現象,也能調節轉錄因子及基因的表現[29]。在細 胞 中 最 常 被 探 討 的 三 個 MAPK family 分 為 : extracellular signal-regulated. kinases ( ERK1/2 ) ; c-Jun. NH2-terminal. kinases/stress-activated protein kinase (JNK/SAPK) 和 p38 MAPK (p38 MAP kinase) 。MAPK cascades 的活化,頇經由 MAPK kinase kinase (MAPKKK) 、MAPK kinase (MAPKK) 、MAP kinase (MAPK) 11.
(31) 三個蛋白質激酶進行一連串的磷酸化作用[30]。 MAPK 家族分類: 1. ERK(extracellular signal–regulated kinases)pathway ERK 有兩種異構體:ERK1 (42 kDa) 和 ERK2 (44 kDa) ,可經由生長因子(growth factors) 、生長激素(cytokines) 、 病毒感染、G protein–coupled receptors 或致癌因子等外來刺 激的影響使 ERK 被磷酸化而活化[31]。ERK1/2 是經由三個 激酶的磷酸化調節以達到訊息的傳遞,促進轉錄因子 AP-1 活化以調控細胞素的表現,在細胞生理中,扮演調控細胞週 期、生長及分化等角色[32]。 2. JNK/SAPK(c-Jun NH2-terminal kinases/stress-activated protein kinase) pathway 此類包含 JNK1(46 kDa) 、JNK2(55 kDa)和 JNK3(57 kDa)三種異構體,是因抑制蛋白質合成,而形成此類因壓力 而活化的蛋白質激酶[33];在各種組織和細胞中 JNK1/2 廣泛 表現,而在心臟、腻及睪丸則主要表現 JNK3 [34]。當熱休克、 滲透休克、細胞激素、生長激素、蛋白質合成抑制劑、抗氧 化劑及過氧化物、UV 和 DNA 損害劑出現時,則可活化 JNK/SAPK 路徑,與DNA結合蛋白 c-Jun 結合並活化,同時 12.
(32) 促進且調節轉錄因子 activator protein-1 (AP-1) 的表現[35]。 當內皮細胞受到. TNF-α 刺激後,使細胞透過 JNK 會大量. 表現細胞黏附因子[36]。 3. p38 MAPK pathway p38 有四種異構體:p38α、p38β、p38γ 和 p38δ,均會 受到 MKK3 和 MAPK kinase MKK6 的磷酸化。當血管內皮 細 胞 或 平 滑 肌 細 胞 受 到 LPS ( lipopolysaccharide ) 及 pro-inflammatory cytokines [ 如 : tumor nucrosis factor-α (TNF-α)、interleukin-1(IL-1)] 的刺激時,p38 MAPK 的 路徑就會活化,使細胞產生發炎反應[37]。因此,p38 MAPK 在細胞訊息傳導中,主要是調控發炎、細胞凋亡等相關訊息。. 13.
(33) 第七節. 基質金屬蛋白酶組織抑制劑. 細胞的基質金屬蛋白酶含量及活性表現均受到嚴密的調節與控 制。細胞初期所製造和分泌的基質金屬蛋白酶皆為不具活性的型態 (latent pro-from),經過酵素活化後,使基質金屬蛋白酶具有分解細 胞外基質的能力;但基質金屬蛋白酶的活化與否,也和內生性組織抑 制劑(tissue inhibitor of metalloproteinases;TIMPs)有關。 基質金屬蛋白酶組織抑制劑是 MMPs 的內生性抑制劑,目前已 知有四種 TIMPs,分別為 TIMP-1,-2,-3,-4。TIMPs 會和活化態的 MMPs 以1:1比例形成非共價鍵結的複合物,並遮蔽 MMPs 活性中 心的鋅離子,使 MMPs 失去分解細胞外基質的功能[38]。在正常生 理狀態下,TIMPs 與 MMPs 的平衡是很重要的,一旦失去平衡,就 可能造成許多疾病,如:動脈硬化和癌細胞轉移等。 在 TIMPs 中,TIMP-1 和 TIMP-2 具有扮演細胞外基質組織和 分 解 平 衡 的 重 要 性 [39] 。 TIMP-1 對 活 化 的 MMP-1 、 MMP-3 和 MMP-9 具有較高抑制率[40],並可與 pro-MMP-9 形成複合物[41, 42];TIMP-2 可抑制 MMP-2 及 Pro-MMP-2,以非共價鍵形式與活 化態 MMP-2 結合形成複合物,有效地抑制 MMP-2 的細胞外基質 的分解活性,主要存於肺及肝中;TIMP-3 廣泛分布於體內,主要存 在於細胞外基質中[43],可促進細胞從細胞外基質分離與細胞型態的 14.
(34) 轉變[44],也可調節細胞分化及細胞週期;TIMP-4 可由腻、心和卵 巢等組織進行包外分泌,在抑制腫瘤生長和發展上扮演重要角色。 在許多組織和體液均存在 TIMPs,據目前研究顯示,TIMP-1 和 TIMP-2 是可溶性的(soluble form),但 TIMP-3 為不可溶性。四種 TIMPs 是由一個類似的 21 kDa 分子基質及兩個以雙硫鍵結穩定的 Domain 組成,其中 N-terminal domain 是 MMPs 主要的抑制部位, 同時亦 可提高 其專 一與 結合效 率, 而 C-terminal domain 則可 與 progelatinase 鍵結,可促進某些細胞生長。TIMPs 在官能基上各有不 同的胺基酸排列,因此對 MMPs 各有不同的專一性和親和力。. 15.
(35) 第八節. 桑黃與 Inotilone 之簡介. 桑黃泛指木層孔菌屬的一些種類;依其寄生的樹種不同,形狀、 顏色和所含的成分也會不同;而生長於桑樹上的桑黃子實體入藥最佳, 桑枝在民間藥用主要治療血崩、血淋、脫肛瀉血、帶下、閉經、脾虛 洩瀉等。目前國內外關於桑黃之生物活性的研究中,主要針對 Phellinus igniarius 及 Phellinus linteus 其子實體的藥理作用。而本研 究主要是以桑黃(Phellinus linteus)為主要基原,在其在分類上桑黃 屬 真 菌 界 ( Eumycota )、 擔 子 菌 門 ( Basidiomycotina )、 層 菌 綱 ( Hymeno-mycetidae )、 非 褶 菌 目 ( Aphyllophorales )、 刺 革 菌 (Hymenochaetaceae)、木層孔菌屬(Phellinus)、桑黃種(Phellinus linteus) 。 桑黃於《本草綱目》記載:性寒、味微苦、能利五臟、宣腸氣、 排毒氣、壓丹石、人熱發及止血等。主治女子漏下赤白,血病腹內結 塊,腫痛,陰陽寒熱,不孕。《藥性論》記載:性甘平、無毒,治血 崩、血淋、脫肛瀉血、帶下、閉經。在現代藥理學活性則是抑制腫瘤、 動脈粥狀硬化、降低血脂等作用[45]。桑黃在中國、韓國、日本及其 它亞洲國家,被使用做為一種傳統醫藥菌菇,用來治療各種疾病,桑 黃含有一種黃色的氧化酵素(Xanthine oxidase)具有抗氧化及抗血栓 的功能,在韓國已經被當作為傳統草藥使用多年[46, 47]。 16.
(36) 桑黃屬藥用真菌類植物,在亞洲地區是一傳統蕈菇類藥材,被用 來治療各種疾病;桑黃具有抗發炎[45]、抗病毒及抗癌等作用[48-50], 此外具有抗肝纖維化作用,改善肝功能,表現出清除自由基的作用 [49],也具有增強自體免疫能力之作用[48],研究顯示桑黃可以誘發 γ干擾素(IFN-γ)的能力、刺激巨噬細胞活化、刺激巨噬細胞產生 interleukin-10(IL-10) 、腫瘤壞死因子-α(tumor necrosis factor-α) 、誘 導殺手細胞活化進而調節自體免疫能力,抑制腫瘤細胞的增殖作用 [51] 。 inotilone. 又稱. (2E)-2-[(3,4-Dihydroxyphenyl)methylene]-5-. methyl-3(2H)-furanon , 為 黃 色 狀 粉 末 的 酚 類 衍 生 物 , 分 子 式 為 C12H10O4,分子量為 218.21,主要來自於 Phellinus linteus,研究結果 顯示,桑黃乙酸乙酯層與 inotilone 具有一些治療效果,如抗發炎[52], 抗高血糖及抗脂質過氧化[53],防癌及抗癌活性[54, 55],可降低細胞 內及細胞外 superoxide anion radical[56]。. Figure 1.. Inotilone 化學結構圖[56]. 17.
(37) 第二章 研究目的 動脈硬化是現代人的文明病,在過程中,許多細胞素、生長因子 及細胞表面黏附因子等,促使平滑肌增生與移行至內皮細胞下間隙, 會導致血管內膜增厚、動脈硬化斑塊、血栓與血管管腔狹窄,其致病 機轉及成因複雜,目前仍無法有效的治療或加以預防。 中草藥所含的天然植物成分,與化學合成之藥物相比,其副作用 較少,且經多年研究證實,許多天然植物多半具有抗氧化、改善免疫 功能及抗癌等效果,故近年來,中草藥的開發利用為一趨勢。 inotilone 為桑黃成分之一,具有很強的抗發炎抑制效果,但目前對於桑黃乙酸 乙酯層與 inotilone 抑制人類主動脈平滑肌細胞增生、移行與侵襲並 無相關文獻。當血管受到刺激而造成發炎因子 TNF-α 釋放會促使動 脈硬化疾病的發生,因此本研究擬探討桑黃乙酸乙酯層與 inotilone 對於人類主動脈平滑肌細胞在 TNF-α 誘導下的增生、移行與侵襲的 表現。. 18.
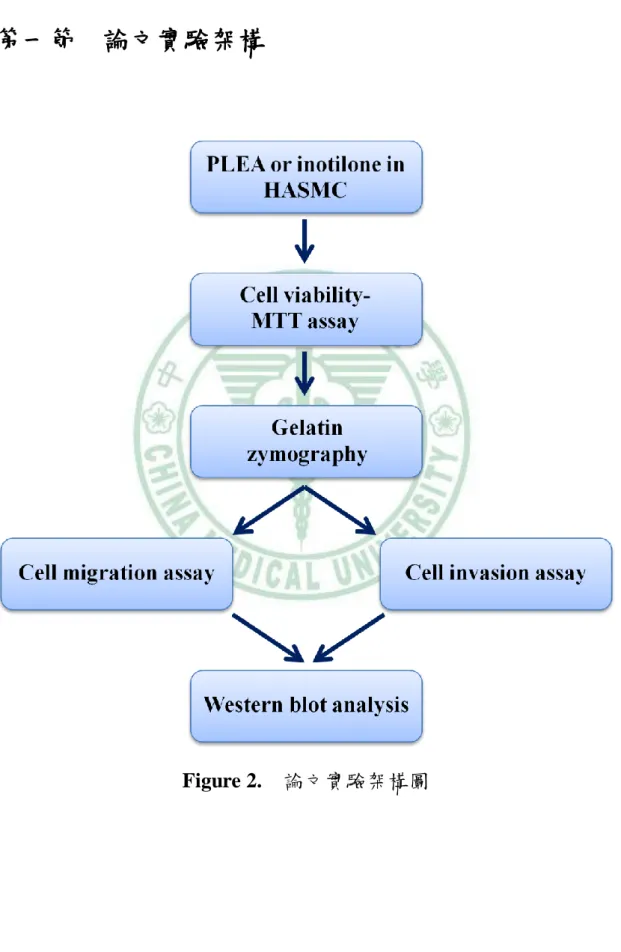
(38) 第三章 研究方法 第一節. 論文實驗架構. Figure 2.. 論文實驗架構圖. 19.
(39) 第二節. 研究材料. 一、 藥品與試劑 1. ABTS(購自 Sigma-Aldrich, USA) 2. 40% Acrylamide(購自 BioNovas) 3. Human apo-transferrin(購自 Sigma-Aldrich, USA) 4. Ammonium persulfate(APS)(購自 Sigma-Aldrich, USA) 5. Bovine serum albumin (BSA) (購自 Sigma-Aldrich, USA) 6. Butylatedhydroxytoluene(BHT)(購自 Sigma-Aldrich, USA) 7. Calcium dichloride(CaCl2)(購自 JT.Baker, USA) 8. Casein(購自 Wako, Japan) 9. Coomassie Blue R-250(購自 Sigma-Aldrich, USA) 10. Dimethyl sulfoxide(DMSO)(購自 Sigma-Aldrich, USA) 11. DPPH(購自 Sigma-Aldrich, USA) 12. Endothelial cell growth supplement(ECGS) (購自 Millipore) 13. Enchanced chemiluminescene(ECL)(購自 PerkinElmer) 14. Ethanol(購自東華化工) 15. Fetal bovine serum(FBS)(購自 GIBCO/BRL, USA) 16. Gelatin(購自 Sigma-Aldrich, USA) 17. Giemsa stain(購自 Sigma-Aldrich, USA) 20.
(40) 18. Glycine(購自 Bionovas) 19. GSH(購自 Sigma-Aldrich, USA) 20. Human TNF-α (購自 Peprotech) 21. Hydrochloric acid(購自 Sigma-Aldrich, USA) 22. Insulin(購自 Sigma-Aldrich, USA) 23. L-ascorbia caid(購自 Sigma-Aldrich, USA) 24. Methanol(購自 Merck) 25. Matrigel(購自 BD Biosciences) 26. 3-(4,5-cimethylthiazol-2-yl)-2,5-diphenyl Tetrazolium Bromide(MTT) (購自 Bionovas) 27. Non-fat instant milk powder(購自 Fluka) 28. N,N,N’,N’-tetra-methyl-ethylenediamine (TEMED) (購自 Bionovas) 29. Nutrient mixture F-12 Ham(購自 Sigma-Aldrich, USA) 30. Pageruler TM. prestained protein ladder(購自 Fermentas). 31. Penicillin/ streptomycin (PS)(購自 GIBCO/BRL, USA) 32. 10 X Phosphate buffered saline(PBS)(購自 Onestar) 33. Phosphoric acid(購自 Sigma-Aldrich, USA) 34. Protease inhibitor cocktail(購自 Bionovas) 35. Protein assay-dye reagent concentrate(購自 Bio-Rad, USA) 21.
(41) 36. PVDF transfer membrane(購自 Pall) 37. Potassium peroxodisufate(購自 Sigma-Aldrich, USA) 38. Quercetin(購自 Sigma-Aldrich, USA) 39. RIPA buffer(購自 Bionavas) 40. 5x Sample buffer(購自 Bionovas) 41. Sodium azide(NaN3)(購自 Sigma-Aldrich, USA) 42. Sodium dodecyl sulfate(SDS)(購自 Bioman) 43. Sodium bicarbonate(購自 Sigma-Aldrich, USA) 44. Sodium selenite(購自 Sigma-Aldrich, USA) 45. TES (購自 Sigma-Aldrich, USA) 46. Trolox(購自 Sigma-Aldrich, USA) 47. Tris- base(購自 Bionovas) 48. Triton X-100 (購自 Sigma-Aldrich, USA) 49. Trypan blue(購自 Sigma-Aldrich, USA) 50. Trypsin-EDTA(購自 GIBCO/BRL, USA) 51. Tween-20(購自 Sigma-Aldrich, USA) 52. 一級抗體: (1) anti-β-actin(購自 Abcam) (2) anti- MMP-2(購自 Abcam) 22.
(42) (3) anti- MMP-9(購自 Abcam) (4) anti-TIMP-1(購自 Abcam) (5) anti-TIMP-2(購自 Abcam) (6) anti-FAK(購自 Millipore) (7) anti-ERK1/2(購自 Invitrogen) (8) anti-JNK1/2(購自 Invitrogen) (9) anti-p38(購自 Invitrogen) (10) anti-FAK(購自 Millipore) (11) anti-IκB (購自 Abcam) (12) anti-Phospho-ERK(購自 Invitrogen) (13) anti-Phospho-JNK(購自 Invitrogen) (14) anti-Phospho-p38(購自 Abcam) (15) anti- Phospho- IκB(購自 Abcam) 53. 二級抗體: (1) Rabbit polyclonal to mouse IgG-HRP(購自 Abcam) (2) Goat Anti-rabbit IgG horseradish peroxidase conjugated affinity purified antibody(購自 Chemicon) 54. inotilone:BJYM Pharm. & Chem. Co. Ltd. (Beijing, China). 23.
(43) 二、儀器設備與器材 1. 微量天平(Denver instrument) 2. 垂直循環式無菌操作台(SCITECH,VCM-620) 3. 二氧化碳恆溫培養箱(SAYNO,mco-17AC) 4. 高壓蒸氣滅菌鍋(TOMIN) 5. 低溫冷凍櫃(JOUAN,VXE-57) 6. 細胞計數器(Haemocytometer) 7. 高速離心機(Hermle) 8. 低溫冷凍超高速離心機 Z300(Hermle,Z-300K) 9. 倒立式顯微鏡(NIKON) 10. pH meter 11. 酵素免疫分析儀(VersaMax) 12. Transwell(Millipore) 13. 乾浴器(Basic Life) 14. 恆溫水浴鍋(TKS) 15. SDS-PAGE 電泳槽套組(Bio-Rad, USA) 16. 冷光分析系統 LAS4000(Fujufilm). 24.
(44) 第三節. 實驗方法. 一、細胞培養 1. 實驗細胞株(Cell line): 本實驗所採用的細胞株為人類主動脈平滑肌細胞(Human Aortic Smooth Muscle Cell/HASMC),購自於生物資源保存及研究中心 (Bioresource Collection and Research Center, BCRC)。. 2. 細胞培養條件 培養人類主動脈平滑肌細胞(HASMC)所使用的培養液為 nutrient mixture F-12 Ham(Ham’s F12K),此外仍頇額外添加 1.5 g/L sodium bicarbonate、10 mM HEPES、10 mM TES、0.05 mg/ml ascorbic acid、 0.01 mg/ml insulin、0.01 mg/ml tranferrin,10 ng/ml sodium Selenite、 0.03 mg/ml ECGS(endothelial cell growth supplement)及10 % 胎牛血 清(fetal bovine serum, FBS)。並將細胞置於恆溫 37 ℃、含 5 % CO2 的細胞培養箱(incubator)中,每三天更換一次培養液,並依細胞滿 度繼代,細胞培養至三、四代時即可進行實驗,實驗進行前以不含血 清蛋白培養液(serum free medium)培養至少 24 小時(starvation)。. 25.
(45) 3. 細胞解凍 將儲存於液態氮桶的人類主動脈平滑肌細胞(HASMCs)取出, 迅速放入 37 ℃ 水浴鍋快速解凍,將解凍後的細胞液吸取至含有 10 mL Ham’s F12K medium 的培養盤中,置於 37 ℃,5% CO2 ,95 % Air 的培養箱培養一天。一天後吸出原本的培養液,以 PBS(phosphate buffered saline) 清洗後,加入新的 Ham’s F12K medium,放置培養 箱培養。. 4. 細胞繼代 細胞培養至八至九分滿時,將培養液吸出,以 PBS 清洗兩次, 加入 1 mL 之 0.1 % Trypsin-EDTA 進行反應,置入培養箱內,待 2 分鐘後取出,輕輕拍打培養盤周圍,使細胞成懸浮液狀態,當細胞打 下後,加入等量 medium 中和 Trypsin-EDTA 活性,並用 1 ml 的 pipette 以緩慢來回抽吸方式將細胞打散,將細胞培養液吸取至 15 mL 離心管以 1,000 rpm,離心五分鐘。離心後將吸出上層廢液,輕 拍散底部細胞後,加入 medium 稀釋,取適量細胞液放入含有 10 mL Ham’s F12K medium 培養盤中,放入培養箱培養。. 26.
(46) 5. 細胞計數 將細胞的培養液吸出,以 PBS 清洗兩次,加入 1 mL 之 0.1 % Trypsin-EDTA 進行反應,當細胞打下後,加入等量 Medium 中和 Trypsin-EDTA 作用,將細胞培養液離心。吸出上層廢液,加入適量 Medium 回溶,取 20 μL 細胞液置於 Eppendorf 與等體積 Trypan Blue 混合均勻,取 10 μL 加至血球計數板上,在倒立式顯微鏡下計 算活細胞數目。. 6. 細胞加藥處理 分別將萃取及購買來的桑黃乙酸乙酯層與 inotilone(分子量為 218.12)以 DMSO 為溶媒,配製桑黃乙酸乙酯層 100 mg/mL 及 inotilone 1 M 之庫存液(stock solution),配置完成後避光儲放於 -20 ℃ 冰箱中,待後續實驗用。實驗進行時,以新鮮培養基將 stock solution 稀釋成實驗所需之濃度。實驗過程中,DMSO 所佔比例不得 超過 0.1 %。. 27.
(47) 二、 細胞存活率試驗(Cell viability assay) 1. 原理: 利用細胞粒腺體中所含琥珀酸去氫酶的作用將. MTT. (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide)的黃 色 tetrazolium salt 會 被 粒 線 體 中 succinate-tetrazolium reductase system 之 dehydrogenase 分解為藍紫色的 formazan,MTT 轉變僅 在活細胞中進行,並堆積在細胞內,且 formazan 形成量與細胞數目 呈正比,以 MTT 為受質,測量存活細胞粒線體中 dehydrogenase 之 活性,以檢定細胞存活率。. 2. 步驟: 將細胞培養於 96 well plate 中,濃度為每格 8 ×103 個細胞,待 細胞貼附之後,每 well 以 PBS 清洗,starvation. 24 小時之後,依. 實 驗 設 計 分 別 更 換 含 不 同 濃 度 桑 黃 乙 酸 乙 酯 層 與 inotilone 的 medium(100 μl/per well),培養 24 小時。移除 medium 後,加入 MTT(0.5 mg/ml,MTT:medium = 1:9),於培養箱中培養 4 小 時,再加入 DMSO 後,以 ELISA reader 測定,吸光值為 570 nm。 以未加藥組之 OD 值當為 control 組,實驗組之 OD 值除以 control 組之 OD. 值為細胞之相對存活百分率(% of control)。 28.
(48) 三、明膠蛋白酵素電泳法(Gelatin zymography) 1. 原理: 有些細胞會分泌 matrix metalloproteinase(MMP),因此細胞培 養液中會含有 MMP,其中 MMP-1、MMP-2、MMP-3、MMP-7、 MMP-9 具有分解 gelatin 的功能;又以 MMP-2 及 MMP-9 能力最 強 。 利 用 加 入 gelatin 作 為 proteinase 基 質 的 SDS-PAGE gel (sodium dodecyl sufate/polyacrylamide gel)將細胞的上清液進行蛋白 質電泳後,蛋白質中具有 proteinase 活性的區域會將膠中的基質分解, 之後再以 coomassie blue 染色。具 gelatinase 活性的區域無法被染色, 再依據區域大小及深淺度來判斷實驗樣品蛋白質中 proteinase 活性 的高低。. 2. 樣品的製備: 將細胞培養於 96 well plate 中,濃度為每格 8 ×103 個細胞,待 細胞貼附之後,每個 well 以 PBS 清洗,starvation 24 小時之後,依 實驗設計分別加入含 TNF-α(100ng/ml) 之不同濃度桑黃乙酸乙酯 層 與 inotilone 的 medium 培 養 24 小 時 ( control 組 不 加 藥 或 TNF-α),收取上清液以備用。. 29.
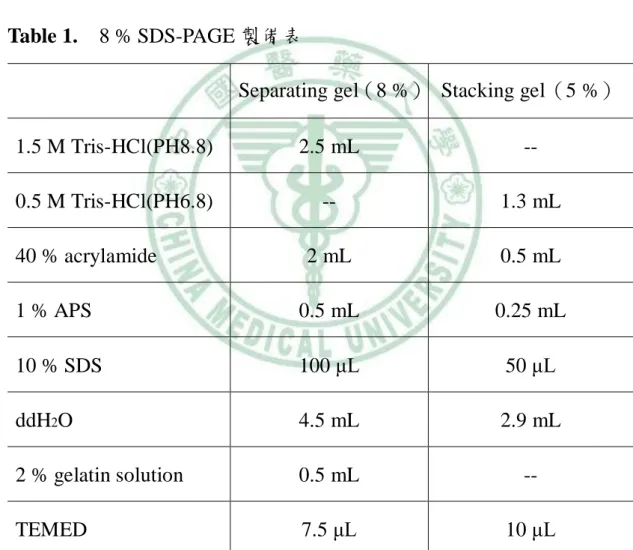
(49) 3. 膠體、緩衝液與溶液之配置: (1) 8 % SDS-PAGE 製備 首先把 glass plate sandwiches 裝置好,配置 0.75 mm 厚度 的 8 % SDS-PAGE,膠分上下兩層,依照下面表格(Table 1)依 序配製 8 % 下層膠(separation gel) 和 4 % 上層膠(stacking gel): Table 1.. 8 % SDS-PAGE 製備表 Separating gel(8 %) Stacking gel(5 %). 1.5 M Tris-HCl(PH8.8). 2.5 mL. --. 0.5 M Tris-HCl(PH6.8). --. 1.3 mL. 2 mL. 0.5 mL. 1 % APS. 0.5 mL. 0.25 mL. 10 % SDS. 100 μL. 50 μL. ddH2O. 4.5 mL. 2.9 mL. 2 % gelatin solution. 0.5 mL. --. TEMED. 7.5 μL. 10 μL. 40 % acrylamide. (2) Destaining solution 製備 70 % methanol、20 % ddH2O 和 10 % acetic acid。 30.
(50) (3) 10X Running buffer 製備 30 g Tris-base、147 g glycine 和 100 mL 10 % SDS 加 ddH2O 定量 至 1 L 即可。 (4) Reaction buffer 製備 40 mM Tris-HCl(pH8.0) 、10 mM CaCl2 和 0.01 % NaN3。 (5) Staining bolution 製備 0.125 % coomassie blue R-250、50 % methanol 和 10 % acetic acid 混合均勻即可。 (6) Washing buffer 製備 2.5 % Triton X-100 in ddH2O。. 4. 步驟: 將 0.1 % gelatin-8 % SDS-PAGE 電泳膠片置於含電泳緩衝液的 電泳槽中。將上清液與 5 X dye 均勻混合注入膠片,以 80 V 進行電 泳分離。結束後,以 washing buffer 在室溫下沖洗 30 分鐘 2 次, 再加入 reaction buffer,於 37 ℃ 恆溫箱中,搖晃反應 16 小時以上, 之後以 staining solution 染色 2 小時,再以 destaining solution 退染, 封膠後觀看結果。. 31.
(51) 四、 細胞移行試驗(Cell migration assay) 參考 Repesh 等人之方法並做些許修改後,以 transwell chamber 進行細胞移行試驗。將人類主動脈平滑肌細胞(HASMCs)以 5 x 105 cell/mL 細胞懸浮於低血清的 Ham’s F12K medium 中,將 transwell chamber 懸掛在 24 well 上,在每個 well 上加入 600 μL 低血清的 Ham’s F12K medium 與 100 ng/mL TNF-α 後,在 transwell chamber 上加入 5 x 104 cell/100 μL 的 HASMCs,再分別加入不同濃度的桑黃 乙酸乙酯層與 inotilone,於 37 ℃,5 % CO2 培養箱培養 8 小時。 將上層的細胞以棉花棒去除,下層細胞以甲醇固定 10 分鐘,晾乾後 以 Giemsa stain 染色,置於顯微鏡下拍照計數。每次試驗隨機挑選 5 個視野進行細胞數計算,並將其平均。相同試驗進行三重複。. 五、 細胞侵襲試驗(Cell invasion assay) Matrigel(BD Biosciences)是由小鼠(mice) Engelbreth Holm Swarm (EHS)腫瘤中所抽出的基底膜,其主要組成為 laminin、 collagen IV,以及 entactin 等。matrigel 在 4 ℃ 可保持於液狀,但 在室溫下則會快速凝固,形成膠狀的基底膜,因此 matrigel 便可以 用來模擬人體動脈血管內彈性膜(membrane elastica interna)的結構, 以便觀察細胞進行侵襲以及轉移的程度。 32.
(52) 同樣參考 Repesh 等人之方法並做些許修改後,cell invasion assay 步驟大致與 cell migration assay 相似。先將 transwell chamber 懸掛於 24 well plate 上,再取 matrigel 覆蓋於 transwell chamber 上 層薄膜,並放置於培養箱中一小時,待 matrilgel 凝固之後,接著將 培養盤中的人類主動脈平滑肌細胞(HASMCs)以 5 × 105 cell/mL 的 細胞懸浮於低血清的 Ham’s F12K medium 中,接著將 transwell chamber. 懸掛於 24 well plate 上,並分別在 24 well plate 的每個. well 中放入 600 μL 低血清的 Ham’s F12K medium 與 100 ng/ml TNF-α 後,於 transwell chamber 之上層再分別加入 5 × 104 cell/100 μL 的 HASMCs , 再 分 別 加 入 不 同 濃 度 的 桑 黃 乙 酸 乙 酯 層 與 Inotilone 後,於 37 ℃、5 % CO2 培養 24 小時。 接著將貼附於 transwell chamber 之下層細胞以甲醇固定十分鐘, 晾乾,以 PBS 清洗濾膜後,將 transwell chamber 上層濾膜細胞以及 matrilgel 以棉花棒完全去除,之後以 Giemsa stain 進行染色一小時, 然後再用蒸餾水清洗濾膜,隔天再將其置於載玻片上,並以倒立顯微 鏡下拍照並計數,每次試驗隨機挑選十個視野進行細胞計數,並將其 平均。. 33.
(53) 六、 西方點墨法(Western blotting) 1. 樣品的配置 將 HASMCs 種於 10 cm2 Dish,在 37 ℃,5 % CO2 培養箱培養 24 小時。待細胞貼附後,給予不同濃度的桑黃乙酸乙酯層、 inotilone 與 100 ng/ml TNF-α 培養 24 小時。待培養時間一到,取出 dish, 依照不同濃度組別,經 Trypsin-EDTA 處理的細胞懸浮液移到 15 mL 離心管,以 1,500 rpm,離心 5 分鐘。倒掉上清液,將細胞打散 後再加入 2. mL 的 PBS 離心(1,250 rpm,5 分鐘)。倒掉上清液,. 將細胞 Pellet 打散後,加入 150 μL 的 RIPA buffer 和 15 μL 的 proteinase inhibitor,置於冰上作用 30 分鐘,以超高速離心機(4 ℃, 15000 rpm,15 分鐘)離心,離心後取出上清液收集,此上清液即為 細胞蛋白質。. 2. 蛋白質標準品配製 根據 Bradford 建立之方法測量蛋白質溶液的濃度,在波長 595 nm 測得的吸光值,換算樣品蛋白質濃度,以一系列已知濃度的 BSA (bovine serum albumin)做成之 standard curve 換算蛋白質濃度。. 34.
(54) 依照下列 Table 2 製作蛋白質標準品檢量線: Table 2.. 蛋白質檢量線配置表 μg/mL. 組成 DDW(μL). 0.1mg/mL BSA(μL). 0. 2. 5. 8. 10. 12. 800. 780. 750. 720. 700. 680. 0. 20. 50. 80. 100. 120. Protein dye (μL). 200. 總體積(μL). 1000. 將配製好的蛋白質標準品混勻反應 5 分鐘後,不同濃度三重複, 加入 96 well 培養盤中,利用酵素免疫分析儀(ELISA reader)在 595 nm 測量各濃度蛋白質標準品之吸光值,求得平均值後,以吸光值和 蛋白濃度求出蛋白質標準品的標準品檢量線(standard curve)的趨勢 線方程式和 R2 值。. 3. 樣品蛋白質定量: 取 10 μL 蛋白質樣品與 790 μL 的 ddH2O 混合,再加入 200 μL protein dye(Bradford),混合均勻反應 5 分鐘後,分別將不同濃 度的樣品以三重複加入 96 well 培養盤中,以酵素免疫分析儀測得 595 nm 的吸光值,求出各濃度樣品的平均值,並代入標準品檢量線 35.
(55) 的趨勢線方程式,即可求得實際各濃度樣品的蛋白質濃度。求出濃度 後,依照所需的體積,將樣品與 5X protein loading dye 混合,以 95 ℃ 水浴加熱變性 5 分鐘,置於冰上冷卻 10 分鐘後離心,即為蛋白 質萃取樣品。. 4. SDS-PAGE 製作及電泳: (1) 原理: SDS 為一種介面活性劑且本身帶有負電荷,其可與蛋白質結 合使得蛋白質變性並且由負極朝向正極來移動;分子量較大的蛋 白質因移動較慢的關係,相對較靠近負極的位置,而分子量較小 的蛋白質則移動較快,因此較為靠近正極的位置,利用蛋白分子 量的不同移動速率不同而將蛋白質分離。 (2) 10 % SDS-PAGE 製備 首先把 glass plate sandwiches 裝置好,配置 1.5 mm 厚度的 10 % SDS-PAGE,膠分上下兩層,依照下面表格(Table 3)依序 配製 10 % 下層膠 (separation gel)和 4 % 上層膠(stacking gel):. 36.
(56) Table 3.. 10 % SDS-PAGE 製備表. 組成. Separating gel(10 %) Stacking gel(5 %). 1.5M Tris-HCl(pH 8.8). 3 mL. -. 0.5M Tris-HCl(pH 6.8). -. 1.3 mL. 40% Acrylamide. 4.2 mL. 0.7 mL. 1% APS. 0.6 mL. 0.25 mL. 10% SDS. 129 μL. 5 μL. ddH2O. 3 mL. 2.7 mL. TEMED. 10 μL. 10 μL. (3) Running buffer(10 X)製備 30 g Tris-base、144 g glycine、4 g SDS 加 ddH2O 至 1 L。使用時, 取 100 mL 10 X Running buffer,加入 900 mL ddH2O 即可。 (4) Transfer buffer(10 X)製備 30 g Tris-base、144 g glycine 加水至 1 L 使用時,取 100 mL 10 X Transfer buffer , 加 入 200 mL methanol , 再 加 入 700 mL 的 ddH2O。 (5) PBST 1000 mL PBS 加入 1 mL Tween-20。 37.
(57) 依上述 10 % SDS-PAGE 配方造膠,將造好的膠放入含 running buffer 的電泳設備裡,將標示分子量的 marker 和各個濃度的 sample 注入膠體孔槽,先以 65 V 進行電泳,待 loading dye 通過 stacking gel, 將電壓調成 100 V 繼續進行電泳。. 5. 蛋白質轉印: 將轉漬夾黑色面向下放,依序將海綿墊、濾紙、膠片、PVDF 膜 (polyvinylidene fluoride) 、濾紙、海棉墊放入轉漬夾,避免氣泡產生, 轉漬夾裝置完成後,放入裝有 transfer buffer 之轉漬裝置中,轉漬裝 置外以冰枕保持系統維持低溫狀態,以 100 V,300 mA 轉漬 1 小時。 轉漬結束後取出 PVDF 膜浸泡於 5 %. 脫脂牛奶中進行 blocking,. 於室溫震盪 1 小時,以阻斷非特異性結合。之後,以 PBST 緩衝液 清洗 PVDF 膜 3 次,每次 5 分鐘。. 6. 一級抗體(primary antibody)與二級抗體(secondary antibody) 加入稀釋之一級抗體置於 4 ℃ 隔夜反應後,再以 PBST 清洗 PVDF 膜 3 次,每次 5 分鐘。接著加入稀釋過的二級抗體反應 2 小 時,以 PBST 緩衝液清洗 3 次,每次 5 分鐘。隨後加入 enhanced chemiluminescence(ECL)進行呈色反應,以 LAS4000 進行拍照。 38.
(58) 七、 統計分析(Statistic analysis): 本實驗所得之數據,以 One-way ANOVA 分析其變異數,再以 Dunnett's test 檢定其間差異之顯著性,凡 p 值小於 0.05 以下時,則 認為有統計意義。. 39.
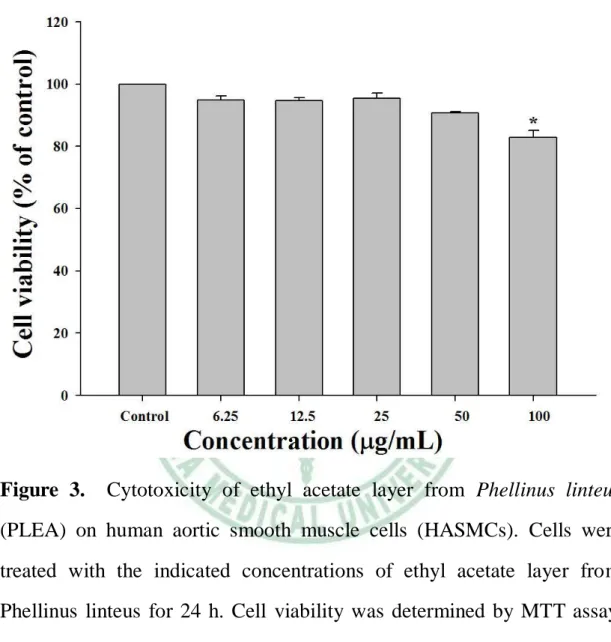
(59) 第四章 第一節. 實驗結果. 桑黃乙酸乙酯層對人類主動脈平滑肌細胞毒. 性及存活率之影響 首先利用 MTT assay 測試桑黃乙酸乙酯層(PLEA)對人類主 動脈平滑肌細胞存活率是否有影響。由於 MTT 可被細胞粒線體中的 琥珀酸去氫酶(Succinate dehydrogenase)還原形成藍紫色的 formazan, 經波長 570 nm 測得的吸光值與活細胞數呈正比,因此可用來偵測細 胞的存活率。本實驗對人類主動脈平滑肌細胞 (HASMCs)給予不 同濃度的 PLEA 培養 24 小時進行細胞存活率實驗,結果由 Figure 3 顯示。在 24 小時給予低劑量 PLEA (6.25 μg/mL),細胞存活 率與控制組無顯著差異(94.9 ± 1.2 %,p>0.05) ,若給予高劑量 PLEA (100 μg/ mL),細胞存活率則為 82.9 ± 2.2 %(p<0.05)。可得知 HASMCs 給予 PLEA 後,細胞的存活率會隨藥物濃度增加而減少, 故 PLEA 對 於 HASMC 的 生長 具 有 抑制 作 用。 藥 物劑 量 在 50 μg/mL 以下,並未對細胞產生明顯毒殺作用(存活率 > 85 %),因 此後續實驗就以 50 μg/mL 以下的濃度(0、 6.25、12.5、25、和 50 μg/mL)進行人類主動脈平滑肌細胞增生與移行的探討。. 40.
(60) 第二節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞表現 MMP-9 活性之影響 當血管平滑肌細胞移行時,會伴隨著分泌大量的蛋白質水解酶來 分解細胞外基質(Extracellular matrix, ECM),這些蛋白質水解酶其 中又以 MMPs 為代表,故本實驗利用 Gelatin zymography 來觀察不 同濃度(0、 6.25、12.5、25、和 50 μg/mL)的桑黃乙酸乙酯層(PLEA) 對於人類主動脈平滑肌細胞(HASMCs)在 TNF-α 刺激下所表現 MMP-9 活性的影響。由 Figure 4 的實驗結果顯示,HASMCs 在 100 ng/mL 的 TNF-α 誘導下,MMP-9 活性為控制組的 109.5 ± 2.0 % (p>0.05);給予低劑量 PLEA(6.25 μg/mL)不會顯著影響 MMP-9 之分泌(為控制組的 101.0 ± 4.8 %,p>0.05),而給予高劑量 PLEA (50 μg/ mL) 則有 82.9 ± 2.2 %(p<0.001)的抑制效果。由以上結果 發現隨著 PLEA 的濃度增加,對於 HASMCs 在 TNF-α 誘導下具有 抑制效果,呈現劑量依存性的關係。至於在 MMP-2 方面,我們發 現酵素活性很弱,不易定量,並且與對照組相比較,皆無顯著差異。. 41.
(61) 第三節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞移行能力的影響 利用 transwell 在離體情況下觀察不同濃度的桑黃乙酸乙酯層 (PLEA)是否會抑制人類主動脈平滑肌細胞(HASMC)在 TNF-α 誘導下穿越 8 μm 孔洞來進行遷移(migration)。將含有 100 ng/mL 的 TNF-α 的 細 胞 培 養 液 放 置 於 transwell 下 層 作 為 化 學 趨 化 物 (Chemo-attractant) ,並將 HASMCs 與不同濃度的 PLEA(0、1.56、 3.13、6.25、和 12.5 μg/mL)放置於 transwell 上層,而兩層之間則 為具有 8 μm 孔洞的濾膜。隨著時間增加,上層細胞受到下層化學趨 化物之吸引,會穿過 8 μm 孔洞濾膜,最後黏附於濾膜下層,藉此可 以觀察 HASMCs 於不同 PLEA 濃度下,移行至 transwell 下層的能 力。經過 8 小時後,將黏附於 8 μm 孔洞濾膜下層之細胞染色,並 於顯微鏡下計算移行細胞的比例以求得量化值。 由 Figure 5 的 實 驗 結 果 顯 示 , HASMCs 在 100 ng/mL 的 TNF-α 誘導下,移行細胞的比例增為控制組的 128.7 ± 7.9 % (p< 0.001);PLEA 的濃度為 1.56 μg/mL 時,移行細胞的比例下降為控 制組的 92.7 ± 5.7 %(p<0.001),而當濃度增加至 12.5 μg/mL 時, 移行細胞的比例下降為 11.66 ± 0.10 % (p<0.001),同樣在 50 42.
(62) μg/mL劑 量下 ,具有 顯著 差異 。由以 上結果 發現 ,PLEA 可抑 制 HASMCs 在 TNF-α 誘導下細胞移行的能力,並呈現劑量依存性的 關係。. 43.
(63) 第四節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞侵襲能力的影響 接下來同樣利用 transwell 在離體情況下觀察不同濃度的桑黃 乙酸乙酯層(PLEA)是否會抑制人類主動脈平滑肌細胞(HASMCs) 在 TNF-α 誘導下穿越 matrigel 及 8 μm 孔洞來進行侵襲(invasion)。 將含有 100 ng/mL 的 TNF-α 的細胞培養液放置於 transwell 下層 作為化學趨化物(chemo-attractant),並將 HASMC 與不同濃度的 PLEA(0、1.56、3.13、6.25、12.5、25、和 50 μg/mL)放置於 transwell 上層,而與遷移實驗不同的地方為兩層之間除了具有 8 μm 孔洞的濾 膜外,尚有一層模擬微血管外基底膜的 matrigel 存在。隨著時間增 加,上層細胞受到下層化學趨化物之吸引,會穿過 8 μm 孔洞濾膜, 最後黏附於濾膜下層,藉此可以觀察 HASMCs 於不同 PLEA 濃度 下,侵襲後並移行至 transwell 下層的能力。經過 24 小時的培養後, 將黏附於 8 μm 孔洞濾膜下層之細胞染色,並於顯微鏡下計算遷移細 胞的比例以求得量化值。 由 Figure 6 的 實 驗 結 果 顯 示 , HASMCs 在 100 ng/mL 的 TNF-α 誘導下,侵襲細胞的比例增為控制組的 116.0 ± 1.1 % (p< 0.05);在 PLEA 的濃度為 1.56 μg/mL 時,侵襲細胞的比例下降為 44.
(64) 控制組的 92.9 ± 4.4 %(p<0.01),而當濃度增加至 50 μg/mL 時, 侵襲細胞的比例下降為 61.3 ± 1.3 % (p<0.001) 。由以上結果可知, PLEA 對於 HASMCs 在 TNF-α 誘導下侵襲的能力具有劑量相關性 的抑制效果。. 45.
(65) 第五節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞分泌 MMP-2 及 TIMP-2 蛋白質表現量 之影響 血管平滑肌細胞增生與移行時,會伴隨著分泌大量的蛋白質水解 酶來分解細胞外基質(extracellular matrix, ECM) ,這些蛋白質水解酶 其中又以 MMPs 為代表;由於 TIMPs 為 MMPs 的內生性抑制劑, 當 TIMP-2 的蛋白質表現量增加時,會抑制 MMP-2 的分泌,使得 MMP-2 減少而無法分解細胞外基質。 故本實驗主要探討桑黃乙酸乙酯層(PLEA)對 TNF-α 刺激人 類主動脈平滑肌細胞(HASMCs)分泌 MMP-2 及 TIMP-2 蛋白質 表現量的情形。以不同濃度(0、 6.25、12.5、25、和 50 μg/mL)的 PLEA 及 100 ng/mL 的 TNF-α 刺激 HASMCs 24 小時後,以西方 點墨分析法觀察 MMP-2 及 TIMP-2 的表現情形。由 Figure 7 的實 驗結果顯示,HASMCs 在 TNF-α 誘導下,MMP-2 的表現增為控制 組的 135.5 ± 12.3 % (p<0.01) ,而 TIMP-2 下降為 84. 6 ± 6.2 % (p <0.05);在 PLEA 的濃度為 6.25 μg/mL 時,MMP-2 的表現下降 為控制組的 100.6 ± 0.6 %(p<0.05) ,而 TIMP-2 則無顯著影響 (為 控制組的 89.6 ± 6.3 %,p>0.05);當濃度增加至 50 μg/mL 時, 46.
(66) MMP-2 比例便降低至 79.5 ± 8.4 %(p<0.01),而 TIMP-2 增為 104.2 ± 3.1 %(p<0.05)。由以上結果可知,HASMCs 在 TNF-α 誘 導下,隨著 PLEA 的濃度增加,TIMP-2 的蛋白表現也有增加的趨 勢;同時也發現 PLEA 的濃度增加,亦可抑制 MMP-2 的蛋白表現 量,具有劑量依存性的關係。. 47.
(67) 第六節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞分泌 MMP-9 及 TIMP-1 蛋白質表現量 之影響 TIMPs 為 MMPs 的內生性抑制劑,當 TIMP-1 的蛋白質表現 量增加時,會抑制 MMP-9 的分泌,使得 MMP-9 減少而無法分解 細胞外基質(extracellular matrix, ECM) 。本實驗主要探討桑黃乙酸乙 酯層(PLEA)對 TNF-α 刺激人類主動脈平滑肌細胞(HASMCs) 分泌 MMP-9 及 TIMP-1 蛋白質表現量的情形。以不同濃度(0、 6.25、 12.5、25、和 50 μg/mL)的桑黃乙酸乙酯層(PLEA)及 100 ng/mL 的 TNF-α 刺 激 HASMCs 24 小 時 後 , 以 西 方 點 墨 分 析 法 觀 察 MMP-9 的表現情形。 由 Figure 8 的實驗結果顯示,HASMCs 在 TNF-α 誘導下, MMP-9 的表現增加為控制組的 117.2 ± 5.6 % (p<0.05),而 TIMP-1 下降為 84.5 ± 1.1 %(p<0.01);在 PLEA 的濃度為 6.25 μg/mL 時,MMP-9 和 TIMP-1 的表現皆無顯著影響(分別為控制組 的 119.7 ± 7.3 %,p>0.05 和 87.7± 8.1 %,p>0.05);當濃度增加 至 50 μg/mL 時,MMP-9 比例便降低至 86.9 ± 2.2 % (p<0.001), 而 TIMP-1 上升為 185.4 ± 4.6 %(p<0.001)。由以上結果可知, 48.
(68) HASMCs 在 TNF-α 誘導下,隨著 PLEA 的濃度增加,TIMP-1 的 蛋白表現量也有增加的趨勢;同時也發現 PLEA 的濃度增加,亦可 抑制 MMP-9 的蛋白表現量,具有劑量依存性的關係。. 49.
(69) 第七節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞分泌 FAK 蛋白質表現量之影響 FAK 為 MAPK pathway 的上游,本實驗主要探討桑黃乙酸乙酯 層 對 TNF-α 刺 激 人 類 主 動 脈 平 滑 肌 細 胞 ( HASMCs ) 表 現 phospho-FAK(分子量 125 kDa) 蛋白質表現量的情形。以不同濃度 (0、 6.25、12.5、25、和 50 μg/mL)的桑黃乙酸乙酯層(PLEA) 及 100 ng/mL的 TNF-α 刺激 HASMCs 24 小時後,利用西方墨點法 來偵測 phospho-FAK 蛋白質表現量的變化。 由 Figure 9 的實驗結果顯示,HASMCs 在 TNF-α 誘導下, phospho-FAK 的表現增為控制組的 108.3 ± 1.2 %(p>0.05);而 PLEA 的濃度為 6.25 μg/mL 時,phospho-FAK 的表現無顯著影響 (為控制組的 100.8 ± 3.0 %,p>0.05) ;當濃度增加至 50 μg/mL 時, phospho-FAK 比例降低至 66.0 ± 0.7 % (p<0.001)。由結果可知, HASMCs 在 TNF-α 誘導下,對於原態的 FAK 無顯著影響,但對於 phospho-FAK 則是隨著 PLEA 的濃度增加,有顯著抑制的效果,具 有劑量依存性的關係。. 50.
(70) 第八節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞 MAPK 訊息傳遞路徑之影響 MAPK pathway 是重要的訊息傳導路徑,同時參與細胞的生長、 分化、轉型與轉移等現象。MAPK 家族分為三類: ERK1/2(分子 量 42/44 kDa) ; JNK(分子量 46/52 kDa)和 p38(分子量 38 kDa)。 本實驗主要探討桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈平滑肌 細胞(HASMCs) 表現 MAPK 訊息傳遞路徑的情形。以不同濃度(0、 6.25、12.5、25、和 50 μg/mL)的桑黃乙酸乙酯層(PLEA) 及 100 ng/mL的 TNF-α 刺激 HASMCs 24 小時後,利用西方墨點法來偵測 蛋白質表現量的變化。 由 Figure 10 至 Figure 12 的 實 驗 結 果 顯 示 , HASMCs 在 TNF-α 誘導下 phospho-ERK、phospho-JNK 及 phospho-p38 的表現 分別增為控制組的 109.6 ± 3.6 % (p<0.01)、112.4 ± 1.4 % (p< 0.05) 及 115.1 ± 7.3 % (p<0.05);在 PLEA 的濃度為 6.25 μg/mL 時,phospho-ERK、phospho-JNK 及 phospho-p38 的表現皆無顯著影 響(分別為控制組的 106.9 ± 0.3 % ,p>0.05、105.0 ± 1.7 %,p>0.05 及 103.0 ± 0.6 %,p>0.05),而當濃度增加至 50 μg/mL 時,比例 便分別降低為 99.2 ± 0.4 % (p<0.01)、 90.5 ± 0.4 % (p<0.01) 51.
(71) 及 85.75 ± 0.45% (p<0.01)。由結果可知 HASMCs 在 TNF-α 誘 導 下 , 對 於 原 態 的 ERK 、 JNK 及 p38 無 顯 著 影 響 , 但 對 phospho-ERK、phospho-JNK 及 phospho-p38 則有顯著抑制的效果, 具有劑量依存性的關係。. 52.
(72) 第九節. 桑黃乙酸乙酯層對 TNF-α 刺激人類主動脈. 平滑肌細胞分泌 IκB 蛋白質表現量之影響 IκB 為 MAPK pathway 的上游,本實驗主要探討桑黃乙酸乙酯 層 對 TNF-α 刺 激 人 類 主 動 脈 平 滑 肌 細 胞 ( HASMCs ) 表 現 phospho-IκB 蛋白質表現量的情形。以不同濃度(0、 6.25、12.5、 25、和 50 μg/mL) 的桑黃乙酸乙酯層(PLEA)及 100 ng/mL 的 TNF-α 刺激 HASMCs 24 小時後,利用西方墨點法來偵測蛋白質表 現量的變化。 由 Figure 13 的實驗結果顯示,HASMCs 在 TNF-α 誘導下, phospho-IκB 的表現增為控制組的 123.8 ± 3.6 %(p<0.001);而 PLEA 的濃度為 6.25 μg/mL 時,phospho-IκB 的表現為控制組的 117.0 ± 3.3 %(p<0.01);當濃度增加至 50 μg/mL 時,phospho-IκB 比例便降低至 89.4 ± 1.6 %(p<0.001)。由結果可知,HASMCs 在 TNF-α 誘導下,對於原態的 IκB 無顯著影響,但對於 phospho- IκB 則是隨著 PLEA 的濃度增加,有顯著抑制的效果,具有劑量依存性 的關係。. 53.
(73) 第十節. Inotilone 對人類主動脈平滑肌細胞存活率. 之影響 首先利用 MTT assay 測試 Inotilone 對人類人類主動脈平滑肌 細胞存活率是否有影響。由於 MTT 可被細胞粒線體中的琥珀酸去氫 酶(Succinate dehydrogenase)還原形成藍紫色的 formazan,經波長 570 nm 測得的吸光值與活細胞數呈正比,因此可用來偵測細胞的存 活率。本實驗對人類主動脈平滑肌細胞(HASMC)給予不同濃度(0、 6.25、12.5、25、50、和 100 μM)的 Inotilone 培養 24 小時進行細 胞存活率實驗,結果由 Figure 14 顯示。在 24 小時給予低劑量 Inotilone (6.25 μM),細胞存活率為控制組的 93.8 ± 3.0 % (p< 0.05),若給予高劑量 Inotilone (100 μM),細胞存活率則下降至 81.9 ± 2.7 %(p<0.001)。可清楚得知 HASMCs 給予 Inotilone 後,細 胞的存活率會隨藥物濃度增加而減少,故 Inotilone 對於 HASMCs 的生長具有抑制作用。藥物劑量在 50 μM 以下,並未對細胞產生明 顯毒殺作用(存活率 > 85 %),因此後續實驗以 50 μM 以下的濃度 (0、 6.25、12.5、25、和 50 μM)進行人類主動脈平滑肌細胞增生 與移行的探討。. 54.
數據




+7
![Figure 1. Inotilone 化學結構圖[56]](https://thumb-ap.123doks.com/thumbv2/9libinfo/8952573.275066/36.892.195.703.834.1044/Figure1Inotilone化學結構圖56.webp)


相關文件
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
Stain (-) because the main body of the tumor consisted of epithelioid or epithelial cells that contained few positive smooth muscle cells. No clear ductal structures or
The chairman/representative director, and at least 2/3 of the board of directors of a limited company or a company limited by shares providing the above services shall be the Republic
[r]
Animal or vegetable fats and oils and their fractiors, boiled, oxidised, dehydrated, sulphurised, blown, polymerised by heat in vacuum or in inert gas or otherwise chemically
Buttermilk, curdled milk and cream, Yogurt, kephir and other fermented or acidified milk and cream, whether or not concentrated or containing added sugar or other sweetening matter
Milk and cream, in powder, granule or other solid form, of a fat content, by weight, exceeding 1.5%, not containing added sugar or other sweetening matter.
Vibrant expansion in the external trade of Mainland China stimulated leaps in merchandise imports which increased from 0.5% in the second quarter to 6.4% and exports which also
