The role of Desmocollin-2 (DSC2) gene in human non-small-cell lung cancer
cell
DSC2 基因在人類非小細胞肺癌細胞功能之分析
Yu-Yi Chen (陳祐翊), Shih-Ting Huang (黃士庭), Po-Chun Ou (歐柏均), Yu-Chih Hu (許育
誌), Yi-Ting Lin (林怡廷), Tai-Lin Lee (李泰林), Meng-Feng Tsai (蔡孟峯)
大葉大學 (彰化縣大村鄉學府路 168 號)
[email protected]
摘要肺癌是這個世界上癌症最常見死亡的原因。非小細胞肺癌是肺癌主要類型,非小細胞肺癌患者預後 不良的原因是轉移,選擇重要和確定的分子標記來標記肺癌患者,特別是侵入和轉移。為了發現新的有 關轉移基因,我們使用了CL1-0、CL1-5 細胞株和微陣列來鑑別基因表現與侵入和轉移功能的相關性在這 項研究中,我們選擇一個差異表現的基因DSC2,進一步了解功能特性DSC2 是鈣依賴的跨膜醣蛋白,是橋 粒蛋白分布最廣的形式,再上皮細胞扮演重要的作用。最近,許多報告指出,在大腸和食道癌DSC2 表現 很低,在腫瘤中可能扮演重要的作用。然而,DSC2 在肺癌的機制還不清楚。在初步結果中,我們發現 DSC2 基因表現與肺癌細胞的侵入、增生和轉移作用呈現負相關。我們認為,DSC2 抑制之後能促進肺癌 細胞侵入、增生和轉移。我們的數據表明,抑制DSC2 確實能 促進肺癌細胞侵入、增生和轉移。我們也 將探討在肺癌中DSC2 的分子機制。 Keywords: 非小細胞肺癌、DSC2、侵入、增生、轉移。 1.前言 1.1 肺癌 肺癌是一種肺部組織的細胞增生失去控制為特徵的疾病。這些增生失去控制的細胞如果沒有及時治 療,將會侵入周圍的組織,甚至轉移至人體的其他部位,從肺部開始產生的癌變也稱為原發性肺癌,主 要是由上皮細胞病變所產生的肺部惡性上皮細胞腫瘤。肺癌是造成全球癌症死亡的重要原因,根據世界 衛生組織統計(http://www.who.int/en/),每年有一百四十萬人因肺癌導致死亡。根據癌細胞在顯微鏡下的 觀察以大小和外觀在組織學上將 肺癌主要分為小細胞肺癌(small-cell lung cancer : SCLC)和非小細胞肺癌 (non-small-cell lung cancer : NSCLC)兩種,而小細胞肺癌佔肺癌比例為 16.8%,非小細胞肺癌佔肺癌比例 為 80.4% (Travis et al., 1995)。小細胞肺癌又稱為燕麥細胞癌,比較少見,較常出現在呼吸道(主要或分 支氣管),相較於其他類型肺癌發展較為迅速,其增生速度也比其它類型肺癌快(Collins et al., 2007)。小細 胞肺癌裡有神經內分泌顆粒,是含有神經內分激素的囊泡,使小細胞肺癌有可能引發腫瘤內分泌的併發 症(Rosti et al., 2006)。在非小細胞肺癌中又可細分為鱗狀細胞癌、肺腺癌、大細胞肺癌,因為三者在預後 的治療方式相似,故一併歸類為非小細胞肺癌。和其他癌症一樣,癌細胞的轉移是主要的致命原因,而 原發性的肺癌經常轉移到腎上腺、肝、腦和骨頭(Sugarbaker et al., 2011)。在癌症的治療上基本上還是以 手術切除、化療、放射線治療為主要治療方式,在手術切除的治療上必須考量病灶是否已發生轉移,並 且必須評估患者的肺功能是否足以承受肺葉的切除,但即使術前經過縝密的評估依然有 4.4%的死亡風險
(Strand et al., 2007),而化療與放射線治療也都對身體有極大負擔。根據統計,經過治療後的肺癌病患存 活五年以上者只有 10.4% 且平均存活期僅為七個月(Grivaux et al., 2011)。因此,許多實驗性治療肺癌的 方法也在發展當中。
1.2 Desmosome
Desmosome 是 細 胞 與 細 胞 間 的 連 結 , 連 接 細 胞 與 細 胞 間 的 質 膜 和 中 間 絲 , 在 細 胞 膜 上 是 由
Desmogleins 和 Desmocollins 這兩個細胞膜上的鈣黏著蛋白家族所組成,在細胞質中 Desmogleins 和 Desmocollins 的尾端透過一連串蛋白的聯結與中間絲連接,這個聯結是由 Plakoglobin、Plakophilins、 Desmoplakin 所組成,首先由 Plakoglobin 和 Desmogleins 和 Desmocollins 的尾端連接,再由 Desmoplakin 連接 Plakoglobin,Plakophilins 會在 Desmoplakin 和 Desmoplakin 的 N 端互相連接,最後由 Desmoplakin 與中間絲連接(Kottke et al., 2006)。細胞與細胞的聯結中 Desmosome 是關鍵的角色,在必須面對機械性壓 力的組織中更是至關重要,例如心肌、膀胱、胃腸道黏膜、皮膚等(Getsios et al., 2004) (Holthofer et al., 2007),Desmosome 第一次被觀察到是在表皮的棘皮層被發現,在 1920 年時被命名 Desmosome,有緊固 的意思(Culkins et al., 2007)。Desmogleins(Dsgs)和 Desmocollins(Dscs)是兩種橋粒鈣粘素的類型(Nollet
et al., 2000),在人類中,Desmocollins 有三種異構體(DSC1-3)而 Desmogleins 四種異構體(DSG1-4)(Green et al.,2007),三個 Desmocollin 基因的異構體在轉錄時都會進行選擇性剪接,形成 DSC“A”和較短的
DSC“B”兩種形式的蛋白質,兩者的差別在羧基端域的長度不同(Collins et al., 1991) (Parker et al., 1991)。 Desmogleins 和 Desmocollins 在細胞外都包含四個重複的鈣粘著蛋白的區域和第五個稱為 extracellular anchor(EA)的區域,這些重複性鈣粘著蛋白,每一個約由 110 個胺基酸組成,這些鈣粘著蛋白是產生黏附 的主要原因(Pokutta et al., 2007),接著是跨膜區域 transmembrane domain(TM),在 transmembrane domain 之後是錨定在質膜的 intracellular anchor (IA) (Kowalczyk et al., 1999),再來則是 intracellular cadherin-like sequence (ICS),和 desmocollin 不同的是 Desmogleins 在 ICS 之後還有 intracellular proline-rich linker (IPL)、 repeat unit domain (RUD)、Desmoglein terminal domain (DTD)(Garrod et al., 2008)。相較於 Desmogleins, Desmocollin 的進化和結構更接近典型的鈣粘素(例如 E-cadherin 和 N-cadherin) (Kljuic et al., 2004)。Dsg2 和 Dsc2 廣泛的表現在所有含有 Desmosome 的組織中,而其他的橋粒鈣粘素主要是在分佈在上皮細胞。 在上皮組織,共有 7 種橋粒鈣粘素。Dsg2 和 Dsg3 主要分佈在整個上皮組織下層,而 Dsg1 主要在上皮組 織上層中表現,Dsg4 主要表現在毛囊和顆粒層,在 Desmocollins 方面 Dsc2 和 Dsc3 主要表現在底層和棘 層,而 Dsc1 表現在顆粒層中。由不同的橋粒鈣粘素出現不同的分佈情形來看,不同組織內的橋粒鈣粘素 蛋白的功能想必會有明顯不同。橋粒鈣粘素在不同組織中會有組織特異性的表現情形,而確切的作用還 不理解,但組織特異性的表現情形是維持組織穩定的關鍵。Desmocollins 在表皮結構的穩定和角質細胞的 增生和分化中發揮了重要作用。在缺乏 Dsc1 的小鼠中,在表皮組織顯示細胞與細胞間的粘附因此而喪失, 在顆粒層會過度增生,毛囊變性,在傷口處角蛋白表現會增加(Chidgey et al., 2001)。同樣,在缺乏 Dsc3 的小鼠中,出現表皮脆弱和脫髮的情形(Chen et al., 2008)。 1.3 Desmocollin-2 (DSC2) 基因和已知的功能與分子機制 Desmocollin 是形成細胞與細胞間附著的鈣粘著蛋白家族的成員之ㄧ,也是組成 Desmosome 的要素, 其結構與典型的鈣粘著蛋白類似(例如 E-cadherin 和 N-cadherin)。在人類中有三種 Desmocollin (Dsc1,Dsc2 和 Dsc3),Desmocollin-2 在轉錄時經過選擇 性剪接 後會有兩種異構體 ,分別 是 Desmocollin-2a 和
Desmocollin-2b,較長的 Desmocollin-2a 是由 901 個胺基酸所組成,較短的 Desmocollin-2b 是由 847 個胺 基酸所組成,兩者在結構上的差異是在位於細胞內 C 端的羧基有所不同,Desmocollin-2a 在 C 端的末端 是由能夠和plakoglobin 連接的intracellular cadherin-like sequence 所組成,Desmocollin-2b 的 C 端則無此 構造(Lorimer et al., 1994),兩種異構體的表現情形則因不同的組織而有所差異(De Bortoli et al., 2010)。 Desmocollin 通常表現在細胞膜上,在細胞膜內經由 plakoglobin 連接中間絲,在細胞膜外的 extracellular anchor 則會和鄰近細胞的 Desmogleins 連結,這個機制使 Desmocollin 對必須承受械性應力的組織產生穩 定的作用,例如心臟、表皮組織(Cheng et al., 2004)。在 Desmocollin-2 knock-down 的斑馬魚模式實驗中也 已經證實 Desmocollin-2 在心臟組織中的功能與重要性(Heuser et al., 2006)。有研究指出 Desmocollin-2a 會 與 connexin43 產生連結(Gehmlich et al., 2011),connexin43 是心室間隙的主要聯接蛋白,與心肌細胞的電 耦合有密切的關聯(Simon et al., 1998)。在許多的研究中發現某些右心室心律不整病患中的 Desmocollin-2 有突變的情形(Christensen et al., 2010)。在心臟組織中 Desmocollin-2b 的表現量遠高於 Desmocollin-2a,而 在右心室心律不整病患中發現 Desmocollin 的突變點 p.A897KfsX4 、p.A897fsX900 (Syrris et al., 2006), 這些突變的位置在羧基的末端,較短的 Desmocollin-2b 並不包含這些區域,所以這些突變只會影響到 Desmocollin-2a,這也使 Desmocollin 在右心室心律不整中的關鍵性目前仍有所爭議(De Bortoli et al., 2010)。
1.4 Desmocollin-2 (DSC2) 與癌症之間的研究
在 1998 年時,在口腔癌中的研究發現 Desmocollin 的表現量與口腔癌的轉移呈現負相關的情形 (Shinohara et al., 1998)。另外在成纖維母細胞中大量表現 Desmocollin-1 時,導致成纖維母細胞出現聚集 的現象,同時也降低成纖維母細胞的侵入基質的能力,因此 Desmocollin 被認為是扮演抑制轉移和侵入的 角色(Tselepis et al., 1998)。但 Desmocollin 減低的現象並不會出現在所有的癌症中,例如在更早之前在結 腸癌的分化與轉移研究中找不出與 Desmocollin 減低的關連性(Collins et al., 1990)。在之後的研究中進一 步的對 Desmocollin 的異構體做進一步的探討,後來發現 Desmocollin 的異構體在腫瘤中的表現情形並不 一致,例如在食道癌中 Desmocollin-2 和 Desmocollin-3 的表現量會下降,但是 Desmocollin-1 反而是增加 的,當時不確定 Desmocollin-1 是否扮演促使腫瘤形成的角色,或者 Desmocollin-1 只是腫瘤形成時的副 產物,有可能是 Desmocollin-2 和 Desmocollin-3 的減少導致 Desmocollin-1 增加(Chidgey et al.,2002)。在 結腸癌中的檢體分析研究中指出 Desmocollin-1 和 Desmocollin-3 會在腫瘤組織表現,而 Desmocollin-2 則 在正的常組織表現,在腫瘤組織中受到抑制(Khan et al., 2006)。進一步研究發現在 Desmocollin-2 的5′端 有兩個 CDX 轉錄因子的結合點,証實 Desmocollin-2 的表現會受到 CDX1、CDX2 兩個轉錄因子的調控, CDX1、CDX2 同時是調控腸道上皮細胞增生、凋亡、細胞附著性以及分化的轉錄因子,因此 Desmocollin-2 被推測可能是扮演抑制 EMT 的角色(Funakoshi et al.,2008)。
2.研究動機
肺癌的轉移是導致病患死亡的主因,而肺癌的治療上又缺乏診斷轉移的生物指標,因此肺癌的轉移 一直都是大家努力研究的目標,本研究希望透過微陣列技術來分析肺癌轉移的模式細胞株,針對肺癌的 轉移找出容易應用在診斷上的生物指標。Desmocollin-2 從組織學上被發現後,對其在細胞膜上的結構已 經有一定程度的了解,但是在分子機制的探討上則只有在 Desmocollin-2 的缺失導致心臟發育異常上有較
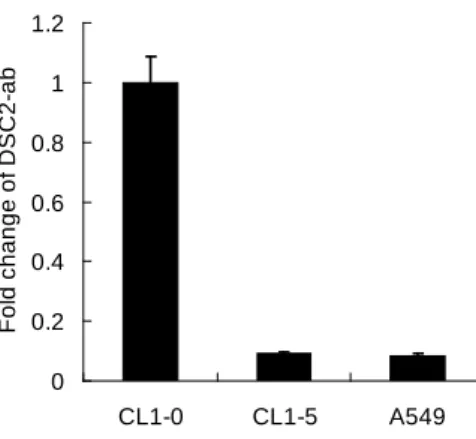
多的研究,至於在癌症方面的研究卻相當缺乏,從目前已經被發表的研究來看,大多數提及 Desmocollin-2 的癌症研究中都只有觀察到 Desmocollin-2 似乎與腫瘤的惡化呈現負相關的關係如:食道癌、胃癌等,但都 沒有針對 Desmocollin-2 在癌症中的分子機制詳加探討,目前只有在結腸癌中有針對 Desmocollin-2 進行 分子機制的研究,但為數也不多的,Desmocollin-2 在癌症中所扮演的角色與分子機制依然有許多尚未明 瞭的地方,Desmocollin-2 在肺癌上更是缺乏相關的研究,因此我們認為這是一個相當具有研究價值的題 目。 3.結果 3.1 藉由即時定量 PCR 的方式分析 CL1-5 以及 CL1-0 肺癌細胞中的基因表現情形 為了驗證 microarray 的結果,因此使用即時定量 PCR 的方式分析 CL1-5 以及 CL1-0 肺癌細胞中的基 因 表 現 情 形 , 我 們 選 擇 了 CDC42 effector protein 3 (CDC42EP3) 、 Forkhead box A2 (FOXA2) 、 Platelet/endothelial cell adhesion molecule (PECM1)、DnaJ (Hsp40) homolog (DNAJB6)、Desmocollin2 (DSC2)、BCL6 corepressor (BCOR6)等六個基因做檢測(圖 1),CDC42EP3 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 5 倍(圖 1A),FOXA2 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 13 倍(圖 1B),PECM1 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 1540 倍(圖 1C),DNAJB6 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 15 倍(圖 1D),DSC2 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 0.25 倍(圖 1E),DSC2 在 CL1-5 細胞中的表現量是在 CL1-0 細胞中的 0.3 倍(圖 1F),由即時定量 PCR 的結果可以確定 microarray 的趨勢是正確的。 3.2 以及時定量 PCR 的方式分析 Desmocollin-2-ab 在侵入能力不同的細胞株中 mRNA 表現量的差異 將不同的肺癌細胞株以即時定量 PCR 的分析,從分析的結果 Desmocollin-2-ab 在較惡性 CL1-5 和 A549 細胞株的中表現量都呈現較少的趨勢,相較於 CL1-0 細胞 Desmocollin-2-ab 在 CL1-5 和 A549 細胞株的中 表現量只有 10 分之 1(圖 2)。 3.3以即時定量PCR的方式分析 shDSC2 的抑制效果
將抑制Desmocollin-2 表現的shDSC2-497、shDSC2-499 、shDSC2-500 載體 Transient transfection至高 度 表 現 Desmocollin-2 的 CL1-0 細 胞 中 , pGIPZ 為 shRNA 的 空 載 體 。 以 即 時 定 量 PCR 的 方 式 分 析 shDSC2-497、shDSC2-499 、shDSC2-500 在CL1-0 細胞中的抑制效果。從 即時定量PCR的結果證 明 shDSC2-497 、 shDSC2-499 、 shDSC2-500 載 體 能 夠 有 效 抑 制 Desmocollin-2 的 mRNA 表 現 量 。 shDSC2-497、shDSC2-499、shDSC2-500 載體分別將CL1-0 細胞中的Desmocollin-2 抑制了 76%、72%、80% 的表現量(圖 3)。
將帶有 shDSC2 的載體轉染至 CL1-0 細胞後,以 puromycin 篩選出穩定表現 shDSC2 片段的細胞株, 並以即時定量 PCR 的方式檢測其 Desmocollin-2 基因的 mRNA 表現量,最後篩選出穩定抑制 Desmocollin-2 基因的細胞株(圖 4)。相較於轉染 pGIPZ 的 CL1-0 細胞 shDSC2-497-3 細胞株中 Desmocollin-2 基因的 mRNA 表現量只有 38%、相較於轉染 pGIPZ 的 CL1-0 細胞 shDSC2-497-4 中 Desmocollin-2 基因的 mRNA 表現量 只有 31%、相較於轉染 pGIPZ 的 CL1-0 細胞 shDSC2-500-6 Desmocollin-2 基因的 mRNA 表現量只有 55%。
3.5 以西方墨點法分析穩定抑制 Desmocollin-2 基因 mRNA 的細胞株
將穩定抑制 Desmocollin-2 基因 mRNA 的細胞株,用西方墨點法分析這些細胞株中的 Desmocollin-2 蛋白質表現情形(圖 5)。從西方墨點法分析結果證明穩定抑制 Desmocollin-2 基因 mRNA 的細胞株其 Desmocollin-2 基因的蛋白表現量也受到有效抑制。shDSC2-497-3、shDSC2-497-4、shDSC2-500-6 細胞株 中 Desmocollin-2 蛋白質表現量都明顯低於控制組 pGIPZ-1、pGIPZ-6 的 Desmocollin-2 蛋白質表現量。
3.6 以 transwell migration 分析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株
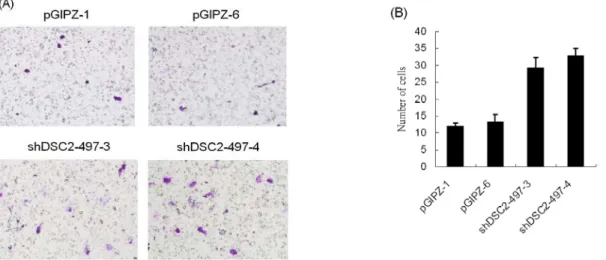
以轉染 pGIPZ 空載體的 CL1-0 細胞 pGIPZ-1、pGIPZ-6 做為控制組進行 transwell migration assay,分 析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株 shDSC2-497-3、shDSC2-497-4,穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株其 migration 能力明顯較高(圖 6 (A))。相較於轉染 pGIPZ 空載體的 CL1-0 細胞 pGIPZ-1、pGIPZ-6,穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株 shDSC2-497-3、shDSC2-497-4 穿過 transwell migration 的細胞數多了 2.5 倍(圖 6 (B))。
3.7 以 colonyformation assay 分析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株其細胞群落形成的能力
將穩定抑制Desmocollin-2 基因的CL1-0 細胞株shDSC2-497-3、shDSC2-497-4 以colonyformation assay 分析其細胞群落形成的能力(圖 7 (A))。與 Transient transfection shDSC2 的 分析結果呈現一致的趨勢。相 較於轉染pGIPZ空載體的CL1-0 細胞pGIPZ-1、pGIPZ-6,穩定抑制Desmocollin-2 基因的CL1-0 細胞株 shDSC2-497-3、shDSC2-497-4 形成colonies的數量多了 2.3 倍(圖 7 (B))。 3.8 以 MTT assay 分析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株增生的速度 將穩定抑制Desmocollin-2 基因的CL1-0 細胞株進行MTT assay去分析其增生的速度(圖 8)。可以從MTT 的結果證明當CL1-0 肺癌細胞中的Desmocollin-2 被抑制時會促進CL1-0 肺癌細胞的增生。以同樣 3000 顆 的細胞生長 4 天後穩定抑制Desmocollin-2 基因的CL1-0 細胞株shDSC2-497-3、shDSC2-497-4 增生速度與 控制組的 pGIPZ-1、pGIPZ-6的CL1-0 細胞增加了一 1 倍。 4.討論
癌症是一種在基因表現上具有高度變化的疾病,因此在癌症的研究上就必須對癌症中多樣且複雜的 基因表現情形去做深入的研究與探討,microarray 在分子研究上是一個十分強大的工具,是其他傳統分析 方式所無法比較的,這個技術讓癌症的研究可以同時檢查大量的基因表現情形(Russo et al., 2003),在過 去多年的研究中 microarray 已經成功的應用在癌症的研究中(Velculescu et al., 1995),microarray 的重要性 在於容易使用不再需要做大規模定序,並且能夠對單一樣進行大量基因的檢測,而基因的表現圖譜是癌 症研究中被廣泛使用的方式,雖然並非所有癌症基因表現情形都是透過轉錄來呈現,但是 microarray 幾 乎是對於找出癌症分子中最全面的方法,它清楚的顯示了細胞的生理狀態,這個十分有效的方法已經成 功的應用在許多的癌症研究中,包括乳癌、頭頸癌、肝癌、肺癌、卵巢癌、胰腺癌、前列腺癌和胃癌 (Bhattacharjee et al., 2001) (Dhanasekaran et al., 2001) (Garber et al.,2001) (Tonin et al., 20016) (Al Moustafa
et al., 2002) (Belbin et al., 2002) (Chen et al.,2002) (Han et al., 2002) (Hedenfalk et al., 2002) (Hippo et al.,,
2002) (Luo et al., 2002)。
肺癌的轉移一直是治療上最棘手的症狀,也是肺癌導致死亡最大的原因,因此我們利用 CL1-0 和 CL1-5 這兩個細胞株進行 microarray 分析,這兩個肺癌細胞株是從 CL1 肺癌細胞株所分離出來的,其分 離的方式是透過 transwell invasion chamber 來區別不同轉移能力的細胞,經過多次重覆的篩選後根據轉移 能力從弱到強由 CL1 細胞分離出 CL1-0、CL1-1 至 CL1-5 細胞株,藉由 microarray 的分析找出 CL1-0 和 CL1-5 這兩個細胞株中表現具有差異的基因,然後再從這些表現有差異的基因中從基因功能以及前人的 研究中推測可能影響細胞轉移的基因,而癌細胞的轉移與細胞附著之間有極大的關係,目前的研究普遍 認為細胞間附著力較弱的細胞其轉移的能力也相對較高,從 microarray 分析的結果來看我們發現 Desmocollin-2 基因相較於在 CL1-0 細胞中的表現量,在 CL1-5 中表現量明顯的較低,而 Desmocollin-2 基因在細胞間的附著中扮演著關鍵的角色(Getsios et al., 2004),Desmocollin-2 是一個跨膜蛋白,在細胞內 部透過數個蛋白的連結最後會與細胞內的中間絲做連結(Kottke et al., 2006),在細胞外部則是會與鄰近細 胞的 Desmoglein 相連結形成細胞間的附著力,而 Desmocollin-2 在具有較高轉移能力的 CL1-5 細胞中表 現量明顯較低於 CL1-0 細胞株,這讓人直覺聯想到可能是 CL1-5 中因為 Desmocollin-2 的低表現量導致 CL1-5 細胞間的附著力下降,最後促進了 CL1-5 細胞的轉移能力,因此從 microarray 的結果來看,我們 認為 Desmocollin-2 具有研究的價值。在即時定量 PCR 的分析中也證明了在高轉移能力的肺癌細胞株 CL1-5 和 A549 中 Desmocollin-2 的 mRNA 以及表現量都呈現較低的趨勢,從這個結果暗示 Desmocollin-2 的表現量與細胞株的轉移能力之間的關聯性假設是成立的。另外從前人的研究中得知 Desmocollin-2 有兩 種異構體分別為 Desmocollin-2a 和 Desmocollin-2b (Lorimer et al., 1994),為了進一步釐清在高度轉移能力 的細胞中 Desmocollin-2 基因表現量的下降與這這兩種 Desmocollin-2 的兩種異構體之間的關係,所以將 Desmocollin-2a 和 Desmocollin-2b 獨立出來進行即時定量 PCR 的分析,分析的結果顯示在高度轉移能力 的細胞中 Desmocollin-2 基因表現量的下降並不是由 Desmocollin-2 的單一種異構體得表現量下降所造 成 , 比 較 分 析 的 結 果 後 我 們 認 為 在 高 度 轉 移 能 力 的 細 胞 中 Desmocollin-2 基 因 表 現 量 的 下 降 與 Desmocollin-2 的兩種異構體之間並沒有獨立關係,最後在西方墨點法的分析中也明顯的證明在高度轉移 能力的細胞中 Desmocollin-2 的蛋白質表現量都低於轉移能力較低的細胞株,這個結果證明 Desmocollin-2 的表現與肺癌細胞的轉移能力呈現負相關的情形,而這個結果與結腸癌檢體中的分析結果相似,在結腸 檢體中的分析顯示在惡性腫瘤組織中的 Desmocollin-2 表現量都明顯較低(Khan et al., 2006),這表示假設 Desmocollin-2 在肺癌細胞中所扮演的角色與結腸癌相似的話,有可能影響肺癌細胞的增生與肺癌細胞的 惡化。在確定了 Desmocollin-2 在高度轉移能力的細胞中具有低表現量的特性後我們為了進一步探討
Desmocollin-2 基因在肺癌細胞中所扮演的角色,我們假設將高度表現 Desmocollin-2 基因的 CL1-0 細胞株 中以 shRNA 的方式將內源性的 Desmocollin-2 抑制掉後,預期此舉將會促進 CL1-0 細胞株的轉移能力, 首先將帶有抑制 Desmocollin-2 基因的 shRNA 載體 shDSC2-497、shDSC2-499、shDSC2-500 轉染至 CL1-0 細胞株中,以即時定量 PCR 的方式分析短暫性轉染 shDSC2-497、shDSC2-499、shDSC2-500 的 CL1-0 細 胞,從即時定量 PCR 的分析結果 shDSC2-497、shDSC2-499、shDSC2-500 這三個載體都能有效抑制 CL1-0 細胞中的 Desmocollin-2 基因的 mRNA 表現量,之後將短暫性轉染 shDSC2 的 CL1-0 細胞以 wound healing assay 來分析其 migration 的能力,分析的結果顯示當 CL1-0 細胞中 Desmocollin-2 基因的 mRNA 被抑制時 會促進 CL1-0 的 migration 能力,從 colonyformation assay 中的分析結果來看,當 CL1-0 細胞中 Desmocollin-2 基因的 mRNA 被抑制時也會促進 CL1-0 細胞株 colony 形成的能力,這些結果與我們當初預期相符,由此 能進一步的證明 Desmocollin-2 基因與細胞轉移能力的關係,另外也對短暫性轉染 shDSC2 的 CL1-0 細胞 進行了 MTT 的細胞增生速率分析,分析的結果顯示 Desmocollin-2 基因被抑制時會促進細胞的增生速率, 從短暫性轉染 shDSC2 的 CL1-0 細胞中所分析得到的初步結果都一再說明 Desmocollin-2 基因在 CL1-0 細 胞中扮演著抑癌基因的角色。進一步將轉染了 shDSC2-497、shDSC2-499、shDSC2-500 的 CL1-0 細胞以 puromycin 進行篩選,最後篩選出穩定表現 shDSC2 的單一細胞株,在以即時定量 PCR 的方式篩檢出穩定 抑制 Desmocollin-2 基因 mRNA 的細胞株,最後再以西方墨點法,檢測這些細胞株中 Desmocollin-2 的蛋 白表現量,最後篩選出了穩定抑制 Desmocollin-2 的 CL1-0 細胞株 shDSC2-497-3、shDSC2-497-4、 shDSC2-500-6,將這些細胞株以 transwell migration 分析這些細胞株 migration 的能力,從分析的結果證明 高度表現 Desmocollin-2 基因的 CL1-0 細胞中當 Desmocollin-2 被抑制時會促進細胞 migration 的能力,而 colonyformation assay 的分析結果與短暫性轉染 shDSC2 的 CL1-0 細胞的分析結果一致,當 CL1-0 細胞 Desmocollin-2 被抑制時促進了 CL1-0 細胞株形成 colony 的能力,這充分說明了 Desmocollin-2 基因在細 胞中扮演抑制轉移的角色,最後在 MTT 與細胞增生的分析中也與短暫性轉染 shDSC2 的 CL1-0 細胞的分 析結果相符,證明 Desmocollin-2 在細胞中除了扮演抑制轉移的角色之外,也參與細胞增生的調控。 在細胞增生的分析中我們發現穩定抑制Desmocollin-2 的CL1-0 細胞株中在生長的過程中,其細胞生 長形態異於控制組的CL1-0 細胞,穩定抑制Desmocollin-2 的CL1-0 細胞株其生長形態較為鬆散,而控制 組的CL1-0 細胞則較為集中,這個現象極可能是EMT的結果,從抑制Desmocollin-2 的表現後導致細胞轉 移能力的提昇,並改變了細胞的生長形態,在過去的研究報告提到在結腸癌中Desmocollin-2 會受到CDX1 與CDX2 這兩個轉錄因子的調控,可能因此進而促進結腸癌的EMT,這些證據都讓人聯想到Desmocollin-2 的被抑制而導致細胞發生EMT的現象(Funakoshi et al., 2008),我們認為這會是針對Desmocollin-2 缺失後 造成細胞轉移的一個研究方向,另外在前人的研究中提到在惡性的直腸癌中除了Desmocollin-2 也呈現較 低的表現量之外(Khan et al.,2006),EGFR的表現量也相對較高,這或許可能是Desmocollin-2 缺失後導致 細胞增生速率增加的原因,而導致EGFR表現量增加的機制與路徑目前還完全不知道,這必須進行更多的 試驗再進一步確認,再最近的研究中指出在結腸癌中Desmocollin-2 的缺失會促進 β-catenin的活性進而活 化Akt的路徑 (Kolegraff et al.,2011),但是這些現象在肺癌中都尚未被研究證實,但是從目前本研究的結果 來看Desmocollin-2 在肺癌中所扮演的角色似乎與結腸癌中相似,都是扮演一個腫瘤抑制基因的角色。
圖 1. 藉由即時定量 PCR 的方式分析 CL1-5 以及 CL1-0 肺癌細胞中的基因表現情形。DSC2 在 CL1-5 細 胞中的表現量是在 CL1-0 細胞中的 0.3 倍(圖 2F),由即時定量 PCR 的結果可以確定 microarray 的趨 勢是正確的。 0 0.2 0.4 0.6 0.8 1 1.2 CL1-0 CL1-5 A549 F ol d c hange of D S C 2-ab 圖 2. 以及時定量 PCR 的方式分析 Desmocollin-2-ab 在侵入能力不同的細胞株中 mRNA 表現量的差異。 相較於 CL1-0 細胞 Desmocollin-2-ab 在 CL1-5 和 A549 細胞株的中表現量只有 10 分之 1。
0 0. 2 0. 4 0. 6 0. 8 1 1. 2 pGI PZ shD SC 2-497 shD SC 2-499 shD SC 2-500 R el at iv e D S C 2 ex pr es s ion l ev el . ' ; 圖 3. 以即時定量PCR的方式分析shDSC2 的抑制效果 。shDSC2-497、shDSC2-499、shDSC2-500 載體分 別將CL1-0 細胞中的Desmocollin-2 抑制了 76%、72%、80%的表現量。 0 0.3 0.6 0.9 1.2 1.5 pGI PZ-1 pGI PZ-6 shD SC 2-49 7-3 shD SC 2-49 7-4 shD SC 2-50 0-6 F lod c hange of D S C 2 圖 4. 以即時定量 PCR 的方式檢測轉染 shDSC2 的 CL1-0 細胞株。相較於轉染 pGIPZ 的 CL1-0 細胞 shDSC2-497-3、shDSC2-497-4、shDSC2-500-6 Desmocollin-2 基因的 mRNA 表現量只有 38%、31%、 55%。 圖 5. 以西方墨點法分析穩定抑制 Desmocollin-2 基因 mRNA 的細胞株。shDSC2-497-3、shDSC2-497-4、 shDSC2-500-6 細胞株中 Desmocollin-2 蛋白質表現量都明顯低於控制組 pGIPZ-1、pGIPZ-6 的 Desmocollin-2 蛋白質表現量。
圖 6. 以 transwell migration 分析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株。 0 20 40 60 80 100 120 140 160 180 200 pGI PZ-1 pGI PZ-6 shD SC 2-49 7-3 shD SC 2-49 7-4 N um ber of c ol oni es
圖 7. 以 colonyformation assay 分析穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株其細胞群落形成的能力。 穩定抑制 Desmocollin-2 基因的 CL1-0 細胞株 shDSC2-497-3、shDSC2-497-4 形成 colonies 的數量多 了 2.3 倍。 0 5000 10000 15000 20000 25000 30000 0 1 2 3 4 Day N um ber of c el ls pGIPZ-1 pGIPZ-6 sh497-3 sh497-4 圖 8. 以MTT assay分析穩定抑制Desmocollin-2 基因的CL1-0 細胞株增生的速度。穩定抑制Desmocollin-2 基因的CL1-0 細胞株shDSC2-497-3、shDSC2-497-4 增生速度與 控制組的 pGIPZ-1、pGIPZ-6的CL1-0 細胞增加了 1 倍。
shDSC2 497-4
shDSC2 497-3
(A) (B)5.參考文獻
Al Moustafa AE, Alaoui-Jamali MA, Batist G, Hernandez-Perez M, Serruya C, Alpert L, Black MJ, Sladek R, Foulkes WD: Identification of genes associated with head and neck carcinogenesis by cDNA microarray comparison between matched primary normal epithelial and squamous carcinoma cells.
Oncogene 2002, 21(17):2634-2640.
Bhattacharjee RN, Banks GC, Trotter KW, Lee HL, Archer TK: Histone H1 phosphorylation by Cdk2 selectively modulates mouse mammary tumor virus transcription through chromatin remodeling. Mol
Cell Biol 2001, 21(16):5417-5425.
Chen J, Den Z, Koch PJ: Loss of desmocollin 3 in mice leads to epidermal blistering. J Cell Sci 2008, 121(Pt 17):2844-2849.
Chen X, Cheung ST, So S, Fan ST, Barry C, Higgins J, Lai KM, Ji J, Dudoit S, Ng IO et al: Gene expression patterns in human liver cancers. Mol Biol Cell 2002, 13(6):1929-1939.
Cheng X, Koch PJ: In vivo function of desmosomes. J Dermatol 2004, 31(3):171-187. Chidgey M: Desmosomes and disease: an update. Histol Histopathol 2002, 17(4):1179-1192.
Christensen AH, Benn M, Bundgaard H, Tybjaerg-Hansen A, Haunso S, Svendsen JH: Wide spectrum of desmosomal mutations in Danish patients with arrhythmogenic right ventricular cardiomyopathy. J
Med Genet 2010, 47(11):736-744.
Collins JE, Legan PK, Kenny TP, MacGarvie J, Holton JL, Garrod DR: Cloning and sequence analysis of desmosomal glycoproteins 2 and 3 (desmocollins): cadherin-like desmosomal adhesion molecules with heterogeneous cytoplasmic domains. J Cell Biol 1991, 113(2):381-391.
Collins JE, Taylor I, Garrod DR: A study of desmosomes in colorectal carcinoma. Br J Cancer 1990, 62(5):796-805.
Collins LG, Haines C, Perkel R, Enck RE: Lung cancer: diagnosis and management. Am Fam Physician 2007, 75(1):56-63.
Culkins CC, Setzer SV: Spotting Desmosomes: The First 100 Years. J Invest Dermatol 2007, 127(E1):E2-E3.
De Bortoli M, Beffagna G, Bauce B, Lorenzon A, Smaniotto G, Rigato I, Calore M, Li Mura IE, Basso C, Thiene G et al: The p.A897KfsX4 frameshift variation in desmocollin-2 is not a causative mutation in arrhythmogenic right ventricular cardiomyopathy. Eur J Hum Genet 2010, 18(7):776-782.
Dhanasekaran SM, Barrette TR, Ghosh D, Shah R, Varambally S, Kurachi K, Pienta KJ, Rubin MA, Chinnaiyan AM: Delineation of prognostic biomarkers in prostate cancer. Nature 2001, 412(6849):822-826.
Folkman J: Angiogenesis and its inhibitors. Important Adv Oncol 1985:42-62.
Funakoshi S, Ezaki T, Kong J, Guo RJ, Lynch JP: Repression of the desmocollin 2 gene expression in human colon cancer cells is relieved by the homeodomain transcription factors Cdx1 and Cdx2. Mol
Cancer Res 2008, 6(9):1478-1490.
M, Rosen GD, Perou CM, Whyte RI et al: Diversity of gene expression in adenocarcinoma of the lung.
Proc Natl Acad Sci U S A 2001, 98(24):13784-13789.
Garrod D, Chidgey M: Desmosome structure, composition and function. Biochim Biophys Acta 2008, 1778(3):572-587.
Gehmlich K, Lambiase PD, Asimaki A, Ciaccio EJ, Ehler E, Syrris P, Saffitz JE, McKenna WJ: A novel desmocollin-2 mutation reveals insights into the molecular link between desmosomes and gap junctions. Heart Rhythm 2011, 8(5):711-718.
Getsios S, Huen AC, Green KJ: Working out the strength and flexibility of desmosomes. Nat Rev Mol
Cell Biol 2004, 5(4):271-281.
Green KJ, Simpson CL: Desmosomes: new perspectives on a classic. J Invest Dermatol 2007, 127(11):2499-2515.
Grivaux M, Zureik M, Marsal L, Asselain B, Peureux M, Chavaillon JM, Prudhomme A, Carbonnelle M, Goarant E, Maury B et al: Five-year survival for lung cancer patients managed in general hospitals.
Rev Mal Respir 2011, 28(7):e31-38.
Hedenfalk IA, Ringner M, Trent JM, Borg A: Gene expression in inherited breast cancer. Adv Cancer
Res 2002, 84:1-34.
Heuser A, Plovie ER, Ellinor PT, Grossmann KS, Shin JT, Wichter T, Basson CT, Lerman BB, Sasse-Klaassen S, Thierfelder L et al: Mutant desmocollin-2 causes arrhythmogenic right ventricular cardiomyopathy. Am J Hum Genet 2006, 79(6):1081-1088.
Hippo Y, Taniguchi H, Tsutsumi S, Machida N, Chong JM, Fukayama M, Kodama T, Aburatani H: Global gene expression analysis of gastric cancer by oligonucleotide microarrays. Cancer Res 2002, 62(1):233-240.
Holthofer B, Windoffer R, Troyanovsky S, Leube RE: Structure and function of desmosomes. Int Rev
Cytol 2007, 264:65-163.
Khan K, Hardy R, Haq A, Ogunbiyi O, Morton D, Chidgey M: Desmocollin switching in colorectal cancer. Br J Cancer 2006, 95(10):1367-1370.
Kljuic A, Bauer RC, Christiano AM: Genomic organization of mouse desmocollin genes reveals evolutionary conservation. DNA Seq 2004, 15(2):148-152.
Kolegraff K, Nava P, Helms MN, Parkos CA, Nusrat A: Loss of desmocollin-2 confers a tumorigenic phenotype to colonic epithelial cells through activation of Akt/{beta}-catenin signaling. Mol Biol Cell 2011, 22(8):1121-1134.
Kottke MD, Delva E, Kowalczyk AP: The desmosome: cell science lessons from human diseases. J
Cell Sci 2006, 119(Pt 5):797-806.
Kowalczyk AP, Bornslaeger EA, Norvell SM, Palka HL, Green KJ: Desmosomes: intercellular adhesive junctions specialized for attachment of intermediate filaments. Int Rev Cytol 1999, 185:237-302.
Lorimer JE, Hall LS, Clarke JP, Collins JE, Fleming TP, Garrod DR: Cloning, sequence analysis and expression pattern of mouse desmocollin 2 (DSC2), a cadherin-like adhesion molecule. Mol Membr
Luo J, Dunn T, Ewing C, Sauvageot J, Chen Y, Trent J, Isaacs W: Gene expression signature of benign prostatic hyperplasia revealed by cDNA microarray analysis. Prostate 2002, 51(3):189-200.
Nollet F, Kools P, van Roy F: Phylogenetic analysis of the cadherin superfamily allows identification of six major subfamilies besides several solitary members. J Mol Biol 2000, 299(3):551-572.
Parker AE, Wheeler GN, Arnemann J, Pidsley SC, Ataliotis P, Thomas CL, Rees DA, Magee AI, Buxton RS: Desmosomal glycoproteins II and III. Cadherin-like junctional molecules generated by alternative splicing. J Biol Chem 1991, 266(16):10438-10445.
Pokutta S, Weis WI: Structure and mechanism of cadherins and catenins in cell-cell contacts. Annu Rev
Cell Dev Biol 2007, 23:237-261.
Rosti G, Bevilacqua G, Bidoli P, Portalone L, Santo A, Genestreti G: Small cell lung cancer. Ann Oncol 2006, 17 Suppl 2:ii5-10.
Russo G, Zegar C, Giordano A: Advantages and limitations of microarray technology in human cancer.
Oncogene 2003, 22(42):6497-6507.
Shinohara M, Hiraki A, Ikebe T, Nakamura S, Kurahara S, Shirasuna K, Garrod DR: Immunohistochemical study of desmosomes in oral squamous cell carcinoma: correlation with cytokeratin and E-cadherin staining, and with tumour behaviour. J Pathol 1998, 184(4):369-381.
Simon AM, Goodenough DA: Diverse functions of vertebrate gap junctions. Trends Cell Biol 1998, 8(12):477-483.
Strand TE, Rostad H, Damhuis RA, Norstein J: Risk factors for 30-day mortality after resection of lung cancer and prediction of their magnitude. Thorax 2007, 62(11):991-997.
Subramanian J, Govindan R: Lung cancer in never smokers: a review. J Clin Oncol 2007, 25(5):561-570.
Sugarbaker DJ, Dasilva MC: Diagnostic workup of lung cancer. Surg Oncol Clin N Am 2011, 20(4):667-679.
Syrris P, Ward D, Evans A, Asimaki A, Gandjbakhch E, Sen-Chowdhry S, McKenna WJ: Arrhythmogenic right ventricular dysplasia/cardiomyopathy associated with mutations in the desmosomal gene desmocollin-2. Am J Hum Genet 2006, 79(5):978-984.
Tonin PN, Hudson TJ, Rodier F, Bossolasco M, Lee PD, Novak J, Manderson EN, Provencher D, Mes-Masson AM: Microarray analysis of gene expression mirrors the biology of an ovarian cancer model. Oncogene 2001, 20(45):6617-6626.
Travis WD, Travis LB, Devesa SS: Lung cancer. Cancer 1995, 75(1 Suppl):191-202.
Tselepis C, Chidgey M, North A, Garrod D: Desmosomal adhesion inhibits invasive behavior. Proc
Natl Acad Sci U S A 1998, 95(14):8064-8069.
Velculescu VE, Zhang L, Vogelstein B, Kinzler KW: Serial analysis of gene expression. Science 1995, 270(5235):484-487.