Human Osteosarcoma U-2 OS Cells Is Carried
Out by Suppression of the Matrix
Metalloproteinase-2, ERK, and JNK Signaling
Pathways
Fu-Shin Chueh,1Ya-Yin Chen,2,3An-Cheng Huang,4Heng-Chien Ho,5Ching-Lung Liao,6 Jai-Sing Yang,7Chao-Lin Kuo,8* Jing-Gung Chung9,10*
1
Department of Health and Nutrition Biotechnology, Asia University, Taichung 413, Taiwan 2
Department of Chinese-Western Medicine Integration, Chung Shan Medical University Hospital, Taichung 402, Taiwan
3
School of Medicine, Chung Shan Medical University, Taichung 402, Taiwan 4
Department of Nursing, St. Mary’s Medicine Nursing and Management College, Yilan 266, Taiwan
5
Department of Biochemistry, China Medical University, Taichung 404, Taiwan 6
Graduate Institute of Chinese Medicine, China Medical University, Taichung 404, Taiwan 7
Department of Pharmacology, China Medical University, Taichung 404, Taiwan 8
School of Chinese Pharmaceutical Sciences and Chinese Medicine Resources, China Medical University, Taichung 404, Taiwan
9
Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan
10
Department of Biotechnology, Asia University, Taichung 413, Taiwan
Received 7 May 2011; revised 16 August 2011; accepted 18 August 2011
ABSTRACT: Bufalin has been shown to exhibit multiple pharmacological activities, including induction of apoptosis in many types of cancer cell lines. Osteosarcoma is a type of cancer which is difficult to treat and the purpose of this study was to investigate the effects of bufalin on the migration and invasion of human osteosarcoma U-2 OS cells. The wound healing assay and Boyden chamber transwell assay were used for examining the migration of U-2 OS cells. Western blotting and gelatin zymography assays were used for the expression and activities of metalloproteinase (MMP)-2, MMP-7 or MMP-9 levels. Western blotting analysis also was used for measuring the levels of growth factor receptor-bound protein 2 (GRB2), son of sevenless homolog 1 (SOS1), c-Jun N-terminal kinases 1/2 (JNK1/2), extracellular signal-regulated kinase
*These authors contributed equally to this work.
Correspondence to: Jing-Gung Chung; e-mail: [email protected] and Chao-Lin Kuo; e-mail: [email protected] Contract grant sponsor: China Medical University, Taichung, Taiwan; Contract grant number: CMU99-ASIA-22 Published online in Wiley Online Library (wileyonlinelibrary.com). DOI 10.1002/tox.20769
1/2 (ERK1/2), and p38 in bufalin-treated U-2 OS cells. Bufalin inhibited the cell migration and invasion of U-2 OS cells in vitro. Moreover, bufalin reduced MMP-2 and MMP-9 enzyme activities of U-2 OS cells. Bufa-lin also suppressed the protein level of MMP-2 and reduced the levels of mitogen-activated protein kinases (MAPKs) such as JNK1/2 and ERK1/2 signals in U-2 OS cells. Our results suggest that signaling pathways for bufalin-inhibited migration and invasion of U-2 OS cells might be mediated through blocking MAPK signaling and resulting in the inhibition of MMP-2. Bufalin could be a useful agent to develop as a novel antitumor agent by virtue of its ability to inhibit tumor cell migration and invasion.#2011 Wiley Periodicals, Inc. Environ Toxicol 00: 000–000, 2011.
Keywords: bufalin; migration and invasion; human osteosarcoma U-2 OS cells; matrix metalloproteinases; traditional Chinese medicine
INTRODUCTION
Cancer is a leading cause of death worldwide. On the basis of the 2009 report from the Department of Health R.O.C. (Taiwan), about 0.3 people per 100,000 die annually from bone cancer in Taiwan (Wu et al., 2011). Osteosarcoma is the most frequent malignant bone tumor in children and adolescents, and it has been characterized by formation of neoplastic bone tissue (Arndt and Crist, 1999; Thompson et al., 2002). The current treatments for bone cancer such as surgery, radiation, chemotherapy, or a combination of radiotherapy and chemotherapy have had mixed success (AbouEl Hassan et al., 2006). There has been growing in-terest in the use of natural products to inhibit the develop-ment of cancer in multiple animal models (Hong and Sporn, 1997) and these compounds also can be used as cancer ther-apy drugs (AbouEl Hassan et al., 2006).
An important strategy in cancer treatment is to prevent or reduce metastasis of cancer cells. Adhesion ability is associated with invasion and migration of tumor cells (Jung et al., 2008; Lee et al., 2009). The matrix metalloprotei-nases (MMPs) and urokinase plasminogen activator (u-PA) are over-expressed in cell metastasis, and MMPs contribute to the invasion and metastasis of various tumor cells (Gullu et al., 2000; Kilian et al., 2006; Mizutani et al., 2000). Hence, the inhibition of MMP expression or enzyme activ-ity (Guruvayoorappan and Kuttan, 2008; Okada et al., 2001; Waas et al., 2003) could be used as early targets for preventing cancer metastasis.
Bufalin is one of the major digoxin-like components in Chan-Su extracts from the venom ofBufo bufo gargarizan, and it acts as a Na1-K1-ATPase inhibitor for increasing the intracellular Ca21 concentration (Bagrov et al., 1993; Krenn and Kopp, 1998). In Asian countries, bufalin has long been used as a treatment for heart failure in Chinese medicine (Datta and Dasgupta, 2000; Yu et al., 2008). Bufalin is also known as a topoisomerase II inhibitor (Hashimoto et al., 1997; Pastor et al., 2002), and it induced differentiation (Zhang et al., 1992) and apoptosis (Watabe et al., 1996) in leukemia cells as well as the inhibition of solid tumor growth in vivo (Han et al., 2007). Bufalin triggered endothelial cell proliferation and angiogenesis (Lee et al., 1997) which resulted in elevated Ca21 levels
and apoptotic death in prostate cancer cells (Yeh et al., 2003). Recently, Jiang et al. reportd that bufalin inhibited human lung cancer cell proliferation via VEGFR1/ VEGFR2/EGFR/c-Met–Akt/p44/42/p38-NF-jB signaling pathways (Jiang et al., 2010). In addition, bufalin induced apoptosis of hepatoma HepG2 cells through both Fas-and mitochondria-mediated signaling pathways (Qi et al., 2011).
To date, there is no information showing bufalin-inhibited migration and invasion of human bone cancer cells. In this study, we investigated the effects of bufalin on the migration and invasion of human osteosarcoma U-2 OS cells. This study demonstrated that bufalin suppressed migration and invasion of U-2 OS cells by down-regulating the levels of MMP-2, ERK and JNK signaling pathwaysin vitro.
MATERIALS AND METHODS
Materials and Reagents
Bufalin, dimethyl sulfoxide (DMSO), propidium iodide (PI), crystal violet, Tween-20, and formaldehyde were purchased from Sigma-Aldrich Corp. (St. Louis, MO). McCoy’s 5a medium, L-glutamine, fetal bovine serum
(FBS), penicillin-streptomycin, and trypsin-EDTA were obtained from Invitrogen Life Technologies (Carlsbad, CA). Primary antibodies such as MMP-2, MMP-7, GRB2, SOS1, JNK1/2, ERK1/2, p38, and b-actin, and second antibodies were purchased from Santa Cruz Bio-technology (Santa Cruz, CA) and diluted in PBS Tween-20 before use.
U-2 OS Cell Culture
The human osteosarcoma cell line (U-2 OS) was purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan). Cells were plated onto 75 cm2 tissue culture flasks with 90% McCoy’s 5a medium supple-mented 2 mM L-glutamine, 10% FBS, and 1%
penicillin-streptomycin (100 Units/mL penicillin and 100 lg/mL streptomycin) and grown at 378C under a humidified 5% CO2atmosphere (Huang et al., 2010).
Determination of Percentage of Viable Cells
U-2 OS cells (23 105cells/well) were maintained in 12-well plates for 24 h and then treated with 25, 50, 75, and 100 nM of bufalin for 24 and 48 h, respectively. Cells were harvested, stained with PI (5 lg/mL) (a PI exclusion method) and then analyzed by flow cytometry (BD Bio-sciences, FACSCalibur Instruction, San Jose, CA) as previ-ously described (Lu et al., 2010; Chiang et al., 2011).
Wound Healing Assay
U-2 OS cells (5 3 105cells/well) were allowed to form a confluent monolayer in 6-wells plates and then cells were wounded with a 200-lL pipette yellow tip. All cells in the plates were treated with bufalin at final concentrations of 25, 50, and 75 nM and then were incubated in new McCoy’s 5a medium with 1% FBS for 12 and 24 h. Photo-graphs were taken using a phase-contrast microscope. The cell-free area of each treatment in the well was measured as previously described (Ho et al., 2009). Cell migration was calculated as the percentage of the remaining cell-free area compared with the area of the initial wound.
Cell Migration Assay
Analysis of migration in U-2 OS cells was assessed using an OrisTMUniversal Cell Migration Assembly kit (Platypus Technologies, Madison, WI) following the manufacturer’s instructions (Li et al., 2010; Oh and Santy, 2010). Briefly, U-2 OS cells were initially stained with 1lM of CellTrack-erTMGreen CMFDA (Invitrogen) for 45 min under growth conditions for long-term staining (Barhoumi et al., 1993). Afterward, cells were centrifuged and washed twice with cultured medium. Labeled U-2 OS cells at a density of 13 104/100lL were seeded into each test 96-well of the Oris plate with the well inserts (stoppers) and then incubated at 378C in a 5% CO2 humidified chamber to permit cell attachment. After a 12-h incubation, the stoppers of each well were removed and then U-2 OS cells were incubated with 0.5% DMSO (as a control) and bufalin (25, 50, and 75 nM), and migrated cells were determined and images were taken for 0, 12, 24, 36, and 48-h periods of time using a fluorescence microscope (Oh and Santy, 2010; Suh and Han, 2010). Migrated cells were assessed by area closure of the detection zone.
In Vitro Boyden Chamber Assay For Migration
Assay
U-2 OS cell migration was using the chemotactic direc-tional migration assay with a 24-well Transwell insert (Ho et al., 2009). Briefly, 8 lM pore filters (Millipore, Billerica, MA) were coated with 30 lg type I collagen (Millipore) for 1 h. Then, cells at a density of 13 104cells/
0.4 mL in 90% McCoy’s 5a medium were placed in the upper chamber and treated with 0.5% DMSO (as a control) and with bufalin (25 and 50 nM) before undergoing migra-tion for 24 and 48 h. Nonmigrated cells were removed in the upper chamber by using a cotton swab and the filter from each treatment was individually stained with 2% crys-tal violet in 2% ethanol after fixing with 3.8% formalde-hyde in PBS. Migrated cells adhering to the underside of the filter were examined, photographed, and counted under using a light microscope at 2003 magnification. Each treat-ment including the control condition was assayed twice and three independent experiments were performed as previ-ously described (Chen et al., 2010; Ho et al., 2009).
In Vitro Boyden Chamber Assay For Invasion
Assay
Invasion of U-2 OS cells was determined using Matrigel (BD Biosciences, Franklin Lakes, NJ)-coated transwell cell culture chambers (8lm pore size, Millipore) as previously described (Ho et al., 2009; Liu et al., 2011). Briefly, cells were cultured for 24 h in serum-free-McCoy’s 5a medium, collected and resuspended in serum-free medium. Isolated cells (1 3 104 cells/well) were then placed in the upper chamber of the transwell insert and treated with 0.5% DMSO (as a control) and bufalin (25 and 50 nM). McCoy’s 5a medium containing 10% FBS was placed in the lower chamber. All samples were incubated for 24 and 48 h at 378C in a humidified atmosphere with 95% air and 5% CO2. Noninvasive cells were then removed in the upper chamber using a cotton swab and the invasive cells were fixed with 3.8% formaldehyde in PBS and stained with 2% crystal violet in 2% ethanol. Invasive cells in the lower sur-face of the filter which penetrated through the Matrigel then were counted and photographed using a light microscope at
Fig. 1. Bufalin affected the percentage of viable U-2 OS cells in vitro. Cells were placed in 90% McCoy’s 5a medium 1 10% FBS and treated with 25, 50, 75, and 100 nM of bufalin for 24 and 48 h. The cells were collected and ana-lyzed for cell viability by flow cytometry as described in Materials and Methods. Each point is mean6 S.D. of three experiments. *p\ 0.05.
2003 magnification as previously described (Ho et al., 2009; Chen et al., 2010).
Gelatin Zymography For MMP-2 and MMP-9
Activity
U-2 OS Cells were plated in 12-well tissue culture plates at a density of 23 106cells/well and then were incubated in serum-free McCoy’s 5a medium in the presence of 0, 10, 20, 40, 60, and 80 nM of bufalin for 24 h. The conditioned medium was then collected and it was separated by electro-phoresis on 10% SDS-PAGE containing 0.18% gelatin (Sigma-Aldrich Corp.). After the electrophoresis, the gels
were soaked in 2.5% Triton X-100 in dH2O twice for a total of 60 min at 258C, and then were incubated in substrate buffer (50 mM Tris base, 0.2 M NaCl, 5 mM CaCl2, and 0.02% Brij 35 in distilled water, pH 8.0) at 378C for 24 h. Bands corre-sponding to activity of MMP 2 and 9 were visualized by nega-tive staining using 0.3% Coomassie blue (Sigma-Aldrich Corp.) in 50% methanol and 10% acetic acid as previously described (Chen et al., 2010; Liu et al., 2011).
Western Blotting Analysis
U-2 OS cells seeded in 6-well plates at density of 13 106 cells/well were incubated with bufalin (25 and 50 nM) for
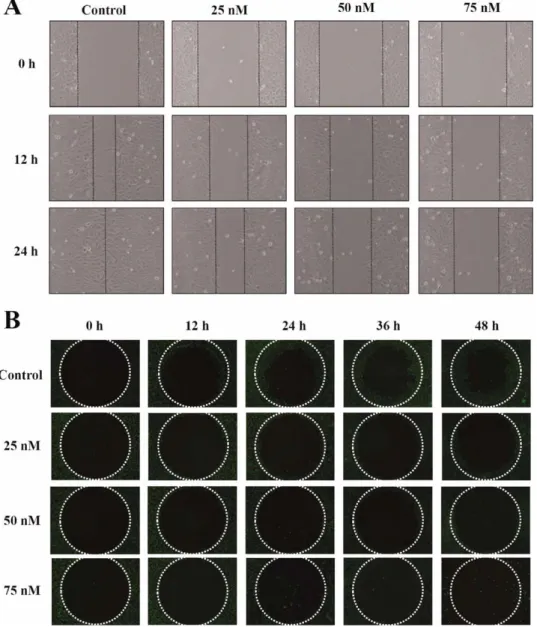
Fig. 2. Bufalin affected the motility and migration of U-2 OS cells. Cells were exposed to 0, 25, 50, and 75 nM of bufalin for various periods of time (12, 24, 36, or 48 h). Cell migra-tion was examined by wound healing assay (A) and OrisTMUniversal Cell Migration Assem-bly kit (B) as described in Materials and Methods. All results represented the average of three experiments and had similar effects.
Fig. 3. Bufalin influenced migration and invasion of U-2 OS cells in vitro. Cells were treated with 0, 25, and 50 nM of bufalin for 24 and 48 h. A: Cell migration was examined by Boyden chamber assay and type I collagen-coated transwell; B: cell invasion was examined by using Matrigel-coated transwell cell culture chambers. Abilities of migration (C) and inva-sion (D) in U-2 OS cells were quantified by counting the number of cells that migrated or invaded the underside of the porous polycarbonate membrane under a phase-contrast microscope and represented the average of three experiments. *p\ 0.05 was compared with the untreated control (Ctl). [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
24 and 48 h. Cells from each treatment were isolated and resuspended in ice-cold PRO-PREPTM protein extraction solution (iNtRON Biotechnology, Seongnam, Gyeonggi-Do, Korea) to extract the total protein fractions. Each sample was centrifuged at 13,0003 g for 10 min at 48C to remove cell debris and for collecting the supernatant. The total protein of each sample was determined using a Bio-Rad protein assay kit (Hercules, CA) using bovine serum albumin (BSA) as the standard (Ho et al., 2009). Protein abundance of JNK1/2, ERK1/2, p38, GRB2, SOS1, MMP-2, and MMP-7 was measured by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting as previously described (Lu et al., 2010; Chiang et al., 2011).
Statistical Analysis
All data are presented as means6 S.D. of three experiments. Statistical comparisons were performed using the Student’s t-test and differences considered significant at the *p less than 0.05 between bufalin-treated and control groups.
RESULTS
Bufalin Affected the Percentage of Viable U-2
OS Cells
In Vitro
U2 OS cells were treated with various concentrations (25, 50, 75, and 100 nM) of bufalin for 24 and 48 h. Cells from each treatment were isolated to determine viability. Figure 1 shows that there were fewer viable cells at bufalin concentrations of 75 and 100 nM (p\ 0.05) when compared with the control sample.
Bufalin Inhibited Cell Migration and Invasion
in U-2 OS Cells
Cell migration was measured using the wound healing assay and the OrisTM Universal Cell Migration Assembly
kit. These results are shown in Figure 2(A,B), which indi-cated that bufalin inhibited the migration of U-2 OS cells in a dose-dependent manner. These results were confirmed by examining effects of bufalin on migration and invasion of U-2 OS cells in vitro by using a 24-well Transwell insert and type I collagen and Matrigel-coated transwell cell cul-ture chambers, respectively. The migration assay showed that bufalin had a significant inhibitory effect on cell migra-tion at concentramigra-tions between 25 and 50 nM [Fig. 3(A,C)] and these results are in agreement with the wound healing data (Fig. 2). U-2 OS cells moved from the upper chamber to the lower chamber in the absence of bufalin, but the pen-etration of the Matrigel-coated filter by U-2 OS cells was inhibited in the presence of bufalin [Fig. 3(B,D)] when cells
Fig. 4. Bufalin altered MMP-2 and MMP-9 enzyme activities from U-2 OS cells. Cells (23 106cells/well) were incubated with 0, 10, 20, 40, 60, and 80 nM of bufalin for 24 and 48 h. The supernatant were harvested after U-2 OS cells treatment and separated by gelatin zymography as described in Materi-als and Methods. The ratio of MMP-2 and MMP-9 activities was quantitated by using ImageJ software.
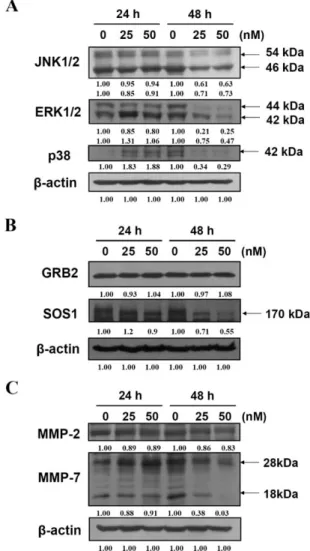
Fig. 5. Bufalin affected on the protein levels associated with migration and invasion in U-2 OS cells. Cells were treated with 0, 25 and 50 nM of bufalin for 24 and 48 h. The total proteins were collected and the protein levels of JNK1/ 2, ERK1/2, p38 (A), GRB2, SOS1 (B), MMP-2, and MMP-7 (C) were examined by SDS-PAGE and Western blotting as described in Materials and Methods. Direct reprobing with anti-b-actin antibody as an internal control.
were incubated with bufalin for 24 and 48-h exposure, respectively.
Bufalin Attenuated the Activities of MMP-2
and MMP-9 in U-2 OS Cells
It is well documented that MMP-2 and -9 play important roles in invasion of cancer cells (Gullu et al., 2000; Kilian et al., 2006). We determined if bufalin could inhibit the secretion of MMP-2 and -9 in U2 OS cells. Gelatin zymog-raphy was used to perform bufalin-affected MMP-2 and MMP-9 activities for 24 and 48-h treatment. Figure 4 indi-cates that bufalin inhibited MMP-2 and MMP-9 activities in U-2 OS cells.
Bufalin Altered Levels of Proteins Associated
With Migration and Invasion in U-2 OS Cells
Effects of bufalin on levels of proteins associated with migration and invasion in U-2 OS cells were examined. Results are shown in Figure 5, which indicated that bufalin decreased protein levels of JNK1/2, ERK1/2, p38 [Fig. 5(A)], GRB2, SOS1 [Fig. 5(B)], MMP-2, and MMP-9 [Fig. 5(C)] in U-2 OS cells. The extracellular signal-regu-lated kinase (ERK) signaling pathway has been reported to up-regulate the expression of MMPs (Chakraborti et al., 2003). Hence, we investigated the effects of bufalin on the ERK and JNK signaling pathways. Results indicated that bufalin inhibited the ERK1/2 and JNK1/2 [Fig. 5(A)] in
U-2 OS cells, suggesting that bufalin suppressed migration and invasion of U-2 OS cells by suppressing the ERK and JNK signaling pathways.
DISCUSSION
Cancer cells interact with their surrounding cells and the matrix proteins allowing cancer cells to replicate, gain nourishment, and migrate to other locations. The interaction of cancer cells with extracellular matrices (ECM) is essen-tial for metastasis which is the principle cause of death in cancer patients (Tsuji et al., 2002). Tumor metastasis (the movement of tumor cells from a primary site to a secondary site) is done via a series of steps such as cell attachment, invasion, migration, vessel formation and cell proliferation, and these steps are regulated by an extremely complex mechanism (Fidler 2002; Fidler et al., 2007).
Many investigators are focused on the development of new agents for blocking cancer cell metastasis (Chakraborti et al., 2003). However, the efficacy of agents is remains unsatisfactory. In normal bone tissue, matrix is constantly degraded and replaced by new matrix (Duffy et al., 2008), and the proteolytic enzymes (matrix metalloproteinases; MMPs) are required in this process (Everts et al., 1992; Ortega et al., 2003). MMPs play an important role in tumor angiogenesis, metastasis and stimulation of growth factor release from the ECM (Coussens and Werb, 1996). Other studies have shown that bufalin inhibited the growth of and induced apoptosis in human osteosarcoma U-2 OS cells (Yin et al., 2007). There is however, no available report on bufalin-inhibiting migration and invasion of U-2 OS cells. We investigated the antimetastatic functions of bufalin on the invasion/migration of human osteosarcoma U-2 OS cells. Our results indicated that bufalin can inhibitin vitro migration and invasion ability of U-2 OS cells [Fig. 3(A,B)]. Our results also showed that bufalin inhibited the MMP-2 and -9 activities (Fig. 4). Bufalin inhibited the pro-tein levels of GRB2, SOS1, ERK1/2, JNK1/2, p38, MMP-2, and -7 (Fig. 5) in U-2 OS cells.
MMP-2 and -9 are involved in the invasive metastatic potential of tumor cells (Zhang et al., 2004) and both enzymes (MMP-2 and -9) were present in human osteosar-coma cells (Meikle et al., 1992). Thus, the inhibition of MMPs expression or enzyme activity can be early targets for preventing cancer metastasis (Coussens and Werb, 1996; Waas et al., 2003; Guruvayoorappan and Kuttan, 2008). It was reported that ERK1/2 is the key molecule of the ERK signaling pathway (Mendes et al., 2007), which promotes tumor invasion and metastasis (Peng et al., 2009). We have used an inhibitor of JNK (SP600125) to pretreat U-2 OS cells which were then exposed to bufalin causing a reduction in MMP-2 and MMP-9 levels (data not shown).
Taken together, our findings suggest that bufalin has multiple anti-metastatic activities in U-2 OS cells. The
Fig. 6. The proposed model for a schematic presentation of the mechanisms in bufalin-suppressed migration and invasion in human osteosarcoma U-2 OS cells. [Color figure can be viewed in the online issue, which is available at wileyonlinelibrary.com.]
possible signaling pathways for bufalin-inhibited migration and invasion in U-2 OS cells may occur by blocking MAPKs (JNK and ERK) resulting in inhibition of MMP-2. Future in vivo efficacy studies with bufalin should be done in animal models.
REFERENCES
AbouEl Hassan MA, Braam SR, Kruyt FA. 2006. Paclitaxel and vincristine potentiate adenoviral oncolysis that is associated with cell cycle and apoptosis modulation, whereas they differ-entially affect the viral life cycle in non-small-cell lung cancer cells. Cancer Gene Ther 13:1105–1114.
Arndt CA, Crist WM. 1999. Common musculoskeletal tumors of childhood and adolescence. N Engl J Med 341:342–352. Bagrov AY, Roukoyatkina NI, Fedorova OV, Pinaev AG,
Ukha-nova MV. 1993. Digitalis-like and vasoconstrictor effects of en-dogenous digoxin-like factor(s) from the venom of Bufo mari-nus toad. Eur J Pharmacol 234(2-3):165–172.
Barhoumi R, Bowen JA, Stein LS, Echols J, Burghardt RC. 1993. Concurrent analysis of intracellular glutathione content and gap junctional intercellular communication. Cytometry 14:747–756. Chakraborti S, Mandal M, Das S, Mandal A, Chakraborti T. 2003. Regulation of matrix metalloproteinases: an overview. Mol Cell Biochem 253(1-2):269–285.
Chen YY, Chiang SY, Lin JG, Ma YS, Liao CL, Weng SW, Lai TY, Chung JG. 2010. Emodin, aloe-emodin and rhein inhibit migration and invasion in human tongue cancer SCC-4 cells through the inhibition of gene expression of matrix metallopro-teinase-9. Int J Oncol 36:1113–1120.
Chiang JH, Yang JS, Ma CY, Yang MD, Huang HY, Hsia TC, Kuo HM, Wu PP, Lee TH, Chung JG. 2011. Danthron, an anthraquinone derivative, induces DNA damage and caspase cascades-mediated apoptosis in SNU-1 human gastric cancer cells through mitochondrial permeability transition pores and Bax-triggered pathways. Chem Res Toxicol 24:20–29.
Coussens LM, Werb Z. 1996. Matrix metalloproteinases and the development of cancer. Chem Biol 3:895–904.
Datta P, Dasgupta A. 2000. Interactions between drugs and Asian medicine: displacement of digitoxin from protein binding site by bufalin, the constituent of Chinese medicines Chan Su and Lu-Shen-Wan. Ther Drug Monit 22:155–159.
Duffy MJ, McGowan PM, Gallagher WM. 2008. Cancer invasion and metastasis: changing views. J Pathol 214:283–293. Everts V, Delaisse JM, Korper W, Niehof A, Vaes G, Beertsen W.
1992. Degradation of collagen in the bone-resorbing compart-ment underlying the osteoclast involves both cysteine-protein-ases and matrix metalloproteincysteine-protein-ases. J Cell Physiol 150:221–231. Fidler IJ. 2002. The organ microenvironment and cancer
metasta-sis. Differentiation 70(9-10):498–505.
Fidler IJ, Kim SJ, Langley RR. 2007. The role of the organ micro-environment in the biology and therapy of cancer metastasis. J Cell Biochem 101:927–936.
Gullu IH, Kurdoglu M, Akalin I. 2000. The relation of gelatinase (MMP-2 and -9) expression with distant site metastasis and tumour aggressiveness in colorectal cancer. Br J Cancer 82:249.
Guruvayoorappan C, Kuttan G. 2008. Amentoflavone inhibits experimental tumor metastasis through a regulatory mechanism involving MMP-2, MMP-9, prolyl hydroxylase, lysyl oxidase, VEGF, ERK-1, ERK-2, STAT-1, NM23 and cytokines in lung tissues of C57BL/6 mice. Immunopharmacol Immunotoxicol 30:711–727.
Han KQ, Huang G, Gu W, Su YH, Huang XQ, Ling CQ. 2007. Anti-tumor activities and apoptosis-regulated mechanisms of bufalin on the orthotopic transplantation tumor model of human hepatocellular carcinoma in nude mice. World J Gastroenterol 13:3374–3379.
Hashimoto S, Jing Y, Kawazoe N, Masuda Y, Nakajo S, Yoshida T, Kuroiwa Y, Nakaya K. 1997. Bufalin reduces the level of to-poisomerase II in human leukemia cells and affects the cytotox-icity of anticancer drugs. Leuk Res 21:875–883.
Ho YT, Yang JS, Li TC, Lin JJ, Lin JG, Lai KC, Ma CY, Wood WG, Chung JG. 2009. Berberine suppresses in vitro migration and invasion of human SCC-4 tongue squamous cancer cells through the inhibitions of FAK, IKK, NF-kappaB, u-PA and MMP-2 and -9. Cancer Lett 279:155–162.
Hong WK, Sporn MB. 1997. Recent advances in chemoprevention of cancer. Science 278:1073–1077.
Huang WW, Chiu YJ, Fan MJ, Lu HF, Yeh HF, Li KH, Chen PY, Chung JG, Yang JS. 2010. Kaempferol induced apoptosis via endoplasmic reticulum stress and mitochondria-dependent path-way in human osteosarcoma U-2 OS cells. Mol Nutr Food Res 54:1585–1595.
Jiang Y, Zhang Y, Luan J, Duan H, Zhang F, Yagasaki K, Zhang G. 2010. Effects of bufalin on the proliferation of human lung cancer cells and its molecular mechanisms of action. Cytotech-nology 62:573–583.
Jung H, Lee KP, Park SJ, Park JH, Jang YS, Choi SY, Jung JG, Jo K, Park DY, Yoon JH, Lim DS, Hong GR, Choi C, Park YK, Lee JW, Hong HJ, Kim S, Park YW. 2008. TMPRSS4 promotes invasion, migration and metastasis of human tumor cells by facilitating an epithelial-mesenchymal transition. Oncogene 27:2635–2647.
Kilian M, Gregor JI, Heukamp I, Hanel M, Ahlgrimm M, Schimke I, Kristiansen G, Ommer A, Walz MK, Jacobi CA, Wenger FA. 2006. Matrix metalloproteinase inhibitor RO 28-2653 decreases liver metastasis by reduction of MMP-2 and MMP-9 concentration in BOP-induced ductal pancreatic can-cer in Syrian Hamsters: Inhibition of matrix metalloproteinases in pancreatic cancer. Prostaglandins Leukot Essent Fatty Acids 75:429–434.
Krenn L, Kopp B. 1998. Bufadienolides from animal and plant sources. Phytochemistry 48:1–29.
Lee DY, Yasuda M, Yamamoto T, Yoshida T, Kuroiwa Y. 1997. Bufalin inhibits endothelial cell proliferation and angiogenesis in vitro. Life Sci 60:127–134.
Lee JK, Bae JA, Sun EG, Kim HD, Yoon TM, Kim K, Lee JH, Lim SC, Kim KK. 2009. KITENIN increases invasion and migration of mouse squamous cancer cells and promotes pul-monary metastasis in a mouse squamous tumor model. FEBS Lett 583:711–717.
Li G, Luna C, Qiu J, Epstein DL, Gonzalez P. 2010. Targeting of integrin beta1 and kinesin 2alpha by microRNA 183. J Biol Chem 285:5461–5471.
Liu KC, Huang AC, Wu PP, Lin HY, Chueh FS, Yang JS, Lu CC, Chiang JH, Meng M, Chung JG. 2011. Gallic acid suppresses the migration and invasion of PC-3 human prostate cancer cells via inhibition of matrix metalloproteinase-2 and -9 signaling pathways. Oncol Rep 26:177–184.
Lu CC, Yang JS, Huang AC, Hsia TC, Chou ST, Kuo CL, Lu HF, Lee TH, Wood WG, Chung JG. 2010. Chrysophanol induces necrosis through the production of ROS and alteration of ATP levels in J5 human liver cancer cells. Mol Nutr Food Res 54:967–976.
Meikle MC, Bord S, Hembry RM, Compston J, Croucher PI, Reynolds JJ. 1992. Human osteoblasts in culture synthesize col-lagenase and other matrix metalloproteinases in response to osteotropic hormones and cytokines. J Cell Sci 103(Part 4):1093–1099.
Mendes O, Kim HT, Lungu G, Stoica G. 2007. MMP2 role in breast cancer brain metastasis development and its regulation by TIMP2 and ERK1/2. Clin Exp Metastasis 24:341–351. Mizutani K, Kofuji K, Shirouzu K. 2000. The significance of
MMP-1 and MMP-2 in peritoneal disseminated metastasis of gastric cancer. Surg Today 30:614–621.
Oh SJ, Santy LC. 2010. Differential effects of cytohesins 2 and 3 on beta1 integrin recycling. J Biol Chem 285:14610–14616. Okada N, Ishida H, Murata N, Hashimoto D, Seyama Y, Kubota
S. 2001. Matrix metalloproteinase-2 and -9 in bile as a marker of liver metastasis in colorectal cancer. Biochem Biophys Res Commun 288:212–216.
Ortega N, Behonick D, Stickens D, Werb Z. 2003. How proteases regulate bone morphogenesis. Ann NY Acad Sci 995:109–116. Pastor N, Dominguez I, Mateos S, Cortes F. 2002. A comparative
study of genotoxic effects of anti-topoisomerase II drugs ICRF-193 and bufalin in Chinese hamster ovary cells. Mutat Res 515(1-2):171–180.
Peng L, Xing X, Li W, Qu L, Meng L, Lian S, Jiang B, Wu J, Shou C. 2009. PRL-3 promotes the motility, invasion, and me-tastasis of LoVo colon cancer cells through PRL-3-integrin beta1-ERK1/2 and-MMP2 signaling. Mol Cancer 8:110. Qi F, Inagaki Y, Gao B, Cui X, Xu H, Kokudo N, Li A, Tang W.
2011. Bufalin and cinobufagin induce apoptosis of human hepa-tocellular carcinoma cells via Fas- and mitochondria-mediated pathways. Cancer Sci 102:951–958.
Suh HN, Han HJ. 2010. Laminin regulates mouse embryonic stem cell migration: involvement of Epac1/Rap1 and Rac1/cdc42. Am J Physiol Cell Physiol 298:C1159–C1169.
Thompson RC Jr, Cheng EY, Clohisy DR, Perentesis J, Manivel C, Le CT. 2002. Results of treatment for metastatic osteosar-coma with neoadjuvant chemotherapy and surgery. Clin Orthop Relat Res 397:240–247.
Tsuji T, Kawada Y, Kai-Murozono M, Komatsu S, Han SA, Take-uchi K, Mizushima H, Miyazaki K, Irimura T. 2002. Regulation of melanoma cell migration and invasion by laminin-5 and alpha3beta1 integrin (VLA-3). Clin Exp Metastasis 19:127– 134.
Waas ET, Wobbes T, Lomme RM, DeGroot J, Ruers T, Hendriks T. 2003. Matrix metalloproteinase 2 and 9 activity in patients with colorectal cancer liver metastasis. Br J Surg 90:1556– 1564.
Watabe M, Masuda Y, Nakajo S, Yoshida T, Kuroiwa Y, Nakaya K. 1996. The cooperative interaction of two different signaling pathways in response to bufalin induces apoptosis in human leu-kemia U937 cells. J Biol Chem 271:14067–14072.
Wu CL, Huang AC, Yang JS, Liao CL, Lu HF, Chou ST, Ma CY, Hsia TC, Ko YC, Chung JG. 2011. Benzyl isothiocyanate (BITC) and phenethyl isothiocyanate (PEITC)-mediated gener-ation of reactive oxygen species causes cell cycle arrest and induces apoptosis via activation of caspase-3, mitochondria dysfunction and nitric oxide (NO) in human osteogenic sarcoma U-2 OS cells. J Orthop Res 29:1199–1209.
Yeh JY, Huang WJ, Kan SF, Wang PS. 2003. Effects of bufalin and cinobufagin on the proliferation of androgen dependent and independent prostate cancer cells. Prostate 54:112–124. Yin JQ, Shen JN, Su WW, Wang J, Huang G, Jin S, Guo QC, Zou
CY, Li HM, Li FB. 2007. Bufalin induces apoptosis in human osteosarcoma U-2OS and U-2OS methotrexate300-resistant cell lines. Acta Pharmacol Sin 28:712–720.
Yu CH, Kan SF, Pu HF, Jea Chien E, Wang PS. 2008. Apoptotic signaling in bufalin- and cinobufagin-treated androgen-depend-ent and -independandrogen-depend-ent human prostate cancer cells. Cancer Sci 99:2467–2476.
Zhang L, Nakaya K, Yoshida T, Kuroiwa Y. 1992. Induction by bufalin of differentiation of human leukemia cells HL60, U937, and ML1 toward macrophage/monocyte-like cells and its potent synergistic effect on the differentiation of human leuke-mia cells in combination with other inducers. Cancer Res 52:4634–4641.
Zhang L, Shi J, Feng J, Klocker H, Lee C, Zhang J. 2004. Type IV collagenase (matrix metalloproteinase-2 and -9) in prostate can-cer. Prostate Cancer Prostatic Dis 7:327–332.