丹參酸及厚朴酚對大鼠胸主動脈平滑肌細胞增生的抑制作用; The inhibitory effects of Salvianolic acid B and Magnolol on vascular smooth muscle cells proliferation
71
0
0
全文
(2) 圖. 次. 圖 1. Sources of reactive oxygen species produced endogenously by the vessel wall and key metabolic pathways for these species 圖 2. Redox regulation of signal transduction components 圖 3. Signal transduction pathways for thr mitogen-activated protein kinases(MAPK) 圖4. Model for mechanical stress initiated-signal transductions in VSMCs 圖5. Death receptor signaling 圖6. Salvianolic acid B 和 Magnolol 對血凊引起平滑肌細胞DNA合 成的影響 圖 7. Salvianolic acid B 及 Magnolol 對平滑肌細胞 PCNA 表達程度的 影響 圖 8. Salvianolic acid B 及 Magnolol 對平滑肌細胞 RIP 表達程度的影 響 圖 9. Salvianolic acid B及 Magnolol對平滑肌細胞 Caspase-3 表達程度 的影響 圖 10. Salvianolic acid B 對 NF-?B promoter 活性的影響 圖 11. Salvianolic acid B 及 Magnolol 在大鼠血清中之抗氧化能力. 2.
(3) 符號與縮寫 AP-1:activator protein-1 BHT:butylated hydroxytoluene BrdU:5-Bromo-2’-deoxyuridine cAMP:cyclic adenosine monophosphate DEPC:diethylpyrocarbonate DMEM:Dulbeco’s modified eagle medium DMSO:dimethy1sulfoxide DTT:Dithiothreitol EGF:epidermal growth factor ELISA:enzyme-linked immunosorbent assay ERK:Extracellular signal-regulated kinase FBS:fetal bovine serum FGF:fibroblast growth factor HPLC:high performance liquid chromatography ICAM-1:Intercellular adhesion molecule type 1 IGF:insulin-like growth factor MAPK:mitogen activated protein kinase MCP-1:monocyte chemoattractant protein-1. 3.
(4) MDA:malondialdehyde MOPS:4-morpholinepropanesulfonic acid NF-κB:nuclear factor -κB NO:nitric oxide PBS:phosphate buffer saline PCNA:proliferating cell nuclear antigen PDGF:platelet-derived growth factor PTCA:percutaneous transluminal coronary angioplasty RIP:Receptor Interacting Protein ROS:reactive oxygen species SAPK:stress-activated protein kinase SOD:superoxide dismutase TBA:2-thiobarbituric acid TCA:trichloroacetic acid TEMED:N, N, N', N'-Tetramethyl-ethylenediamine TGF-β1:transforming growth factorβ1 VCAM-1:vascular cell adhesion molecule-1. 4.
(5) 中文摘要 根據衛生署統計心臟疾病是國人十大死因之一,其中冠狀動脈阻 塞佔了大部份。目前對於冠狀動脈阻塞的治療大多施以血管擴張術 (angioplasty) 或血管繞道手術 (surgical bypass grafting) ,近年發展的 經皮穿腔冠狀動脈血管成形術 (percutaneous transluminal coronary angioplasty, PTCA) 及支架置入方法可以不經開心手術即可達到治療 的效果,對冠狀動脈治療助益頗多。在美國一年有約 40 萬件的血管 擴張術病例,而這些病例中約 30~50% 在六個月後會發生血管再阻 塞現象而必須再次接受血管擴張術,在國內比例則更高。這對病患及 其家屬造成很大的心理及經濟上的負擔。其中平滑肌細胞不正常的增 生是造成再阻塞現象的主因之一。 因此我們利用傳統中藥丹參萃取物 salvianolic acid B 及厚朴萃 取物 magnolol 對大鼠胸主動脈平滑肌細胞進行 in vitro 的實驗。在細 胞增生實驗的結果看來 0.05mg/ml 的 magnolol 對 PCNA 的表達及 DNA 合成有抑制的作用;而 salvianolic acid B 則無效果。不過二者對 RB 和 FAK 沒有影響。在探討藥物是否引起細胞凋亡的實驗中, western blot的結果顯示 salvianolic acid B和 magnolol明顯的引發 RIP 及 caspase-3 的表達。另外,0.1mg/ml salvianolic acid B 能抑制 NFκB promoter 的活性。在抗氧化實驗中,salvianolic acid B 和 magnolol. 5.
(6) 明顯的減少大鼠血清中 MDA 的產生。 根據這些結果,我們認為 salvianolic acid B 和 magnolol 對平滑 肌細胞的增生趨勢有抑制作用。salvianolic acid B 可能經由抑制 NFκB 引起細胞凋亡。Magnolol 則是抑制細胞進入細胞週期中的 S 期。. 6.
(7) 第一章 前言 第一節 經皮穿腔冠狀動脈血管成形術(Percutaneous Transluminal Cornary Angioplasty) 冠狀動脈是環繞在心臟上的營養血管,它提供心臟收縮或舒張時 所需要的氧氣及營養分,假如膽固醇等脂肪物質在冠狀動脈內壁積聚 成為粥狀斑,則血管會造成狹窄硬化或阻塞而引起心絞痛、呼吸急促 等心臟缺氧之症狀,如此即稱為冠狀動脈疾病 (coronary artery disease, CAD )又稱為冠心病。CAD 的臨床特徵就是,當人在大量運動時會產 生心肌缺氧、心絞痛、EKG 異常及心肌灌注掃描有缺影(defect)的現 象。心臟是個需要大量氧氣供應的器官,如果血流不順暢,心臟無法 正常運轉,心臟的肌肉就會很難正常工作,最後導致心絞痛、心肌梗 塞等心臟病症狀。急性心肌梗塞是冠狀動脈疾病的一種臨床表現,冠 狀動脈因為血栓栓塞,導致心肌細胞缺乏血液供應而缺氧、甚至壞 死。對於急性心肌梗塞的治療,最首要的目標是使梗塞的冠狀動脈能 儘速重新獲得血液灌注(reperfusion)。早期心肌梗塞,若未能及時搶 救,死亡率高達 30%以上,1980 年代使用血栓溶解劑用於急性心肌 梗塞後,死亡率可降至 10%左右。現今採用緊急冠狀動脈氣球擴張術 來治療急性心肌梗塞之病人,這個方法是在病患心肌梗塞發作 12 小 時以內,利用緊急心導管手術施行冠狀動脈氣球擴張術,立即打通梗. 7.
(8) 塞的血管,建立足夠的冠狀動脈血流。以氣球擴張術緊急施術搶通血 管,合併使用血管支架,死亡率更降到 5%。所以這項手術已超越以 往 80 年代之血栓溶解治療,成為一安全而有效之治療方式。因此緊 急冠狀動脈氣球擴張術及冠狀動脈支架置放術已成為治療急性心肌 梗塞病人的新趨勢。 根據衛生署統計心臟疾病是國人十大死因之一,其中冠狀動脈阻 塞佔了大部份。目前對於冠狀動脈阻塞的治療大多施以血管擴張術 (angioplasty) 或血管繞道手術 (surgical bypass grafting) ,近年發展的 經皮穿腔冠狀動脈血管成形術 (percutaneous transluminal coronary angioplasty, PTCA) 及支架 (stent) 置入方法可以不經開心手術即可 達到治療的效果,對冠狀動脈治療助益頗多。在美國一年有約 40 萬 件的血管擴張術病例,而這些病例中約 30~50% 在六個月後會發生 血管再阻塞現象而必須再次接受血管擴張術[1~3],在國內比例則更 高。這對病患及其家屬造成很大的心理及經濟上的負擔。過去二十年 許 多 藥 物 [ 例 如 : 血 小 板 抑 制 劑 (antiplatelet) 、 血 拴 抑 制 劑 (antithrombotic)、抗凝血劑 (anticoagulant)… ] 及技術上對治療血管再 阻塞的努力均無很好的效果。直到 1995 年經皮穿透血管擴張術後之 血管內支架置入 (endovascular stenting) 的方法才使動脈再阻塞的治 療有了進展,但其仍有 20% 的再阻塞機率。. 8.
(9) 這是由於當氣球擴張後給予血管內壁外力而造成血管內壁受 損 ,導致局部發炎或出血因而使血液基質、單核巨噬細胞 (mononuclear macrophage) 及淋巴球 (lymphocyte) 在此累積;另外, 血管逐漸產生組織重組 (remodeling) [4~7] ,細胞外間質(extracellular matrix)的形成[8,9]以及血小板附著與聚集在血管受傷的位置[10~12] 並且開始活化,導致多種對血管有不同作用的物質與調節因子的分泌 [13],進而活化平滑肌細胞和巨噬細胞表達出數種激素、生長因子, 或其他基因產物 等, 造成對管壁平滑肌細胞的刺激而使之遷移 ( migration)及增殖(proliferation),如此過度的反應使血管內膜 (intima)增厚因而造成血管再次狹窄。. 第二節. 血管再阻塞(restenosis) 的病理學. 血管再阻塞發生的早期 (數小時至數天) 與血栓形成及血管再 彈縮回(recoil)有關;大部分的病例中顯示血管再阻塞後期主要由 細胞外間質及炎性細胞的刺激下造成纖維組織細胞的增生[14]。在大 鼠的動物模式中,當頸動脈受氣球擴張破壞後 24 至 72 小時內可觀察 到血管中層的平滑肌細胞開始進行 DNA 合成[15,16],接著穿過內部 纖維層而構成新生的內膜,這些在內膜新生的細胞開始增生且在細胞 外間質處產生大量的膠原蛋白。仔細探究其形成之機轉可歸納成下面. 9.
(10) 幾點原因: 一、內膜損傷: 手術過程中造成的內膜損傷導致不同程度的內皮 剝落,使形成血栓的下內皮曝露,接著造成該處發炎[17~20]而使得 單核巨噬細胞及淋巴球浸滲(infiltration)到該處。同時由於血管壁 的破裂造成出血,使血小板釋出多種 cytokines,其中一種 cytokine 是血小板衍生的生長因子 (platelet-derived growth factor, PDGF),是一 種 很 強 的 化 學 性 趨 化 物 質 (chemoattractant) , 促 使 monocyte chemoattractant protein-1 (MCP-1)表現,使得更多的單核細胞及巨噬細 胞聚集,形成一個信號增幅的迴路,巨噬細胞 本身也會釋放 cytokines、metalloproteinases 和 growth factors,如此持續對血管壁平 滑肌細胞的激發與刺激而造成其遷移與增殖。如此在傷口該處便有血 液基質的聚集反應、血管組織的聚集反應,及彼此間的相互作用,而 造成血管肌內膜細胞增生 (myointima cellular hyperplasia),進而形成 血管再阻塞的發生。 因此,內皮受傷害的程度對於血管的修復來說是個很重要的決定 因子。舉例來說,受過損傷的動脈中部分剩餘的內皮細胞可緩和細胞 增生並減少新生內膜的增殖[20,21];Nugent et al. [22]在豬及牛的頸動 脈損傷模式中將內皮細胞移植至血管後發現可以減少新生內膜增 厚,顯示內皮細胞在血管生理中的鄰近調控(paracrine)調節中是個. 10.
(11) 關鍵的角色。而且增生中或受損的內皮細胞所釋放出的 peptides 及 proteoglygans 能刺激平滑肌細胞生長[23~26]。相反地,靜止生長中的 內皮細胞藉由活化 transforming growth factorβ(TGF-β)及釋放具 抑制生長功能的 heparan sulate proteoglycans [27,28] ,提供一個防止 血栓形成的屏障來隔絕血清中致分裂原(mitogen)和化學性趨化物 質對平滑肌細胞產生刺激,進而抑制平滑肌細胞遷移和增生。 二、發炎反應:在 PTCA 後可以觀察到一些 adhesion recepters 會 大量表現,使得單核球和嗜中性白血球開始聚集的現象。有研究指出 對有冠狀動脈硬化斑的病患施行氣球擴張術後,嗜中性白血球的 Mac-1 (CD11b)和血小板上 the activated fibrinogen receptor (LIBS1)表 現增加 [29]。發炎反應及新生內膜形成的程度和血管損傷程度也十 分地有關係,因為單位受損面積上的單核球數目和動脈內膜的增長有 線性上的關係[30,31]。此外,藉由 Mac-1 (CD11b/CD18)-deficient 小 鼠以及給予兔子抗 Mac-1 單株抗體[32,33],皆能減少新生內膜增厚來 看,發炎反應在內膜受損後由白血球媒介的再阻塞中扮演著相當重要 的角色。 另一方面,也有學者在施行過 PTCA 的病人身上發現在急性期的 代表物質 c-reactive protein [34],這是一種當體內有急性炎症、細菌 感染、組織的破壞、惡性腫瘍時很快就出現而治癒時又很快就消失的. 11.
(12) 蛋白(急性期反應蛋白 acute phase reactant)。由這些發現得知,再 阻塞的病理進程中很明顯地受炎性細胞活化的影響。 三、氧化壓力:氧化壓力在血管受損後引發的反應中所扮演的調 節角色目前仍沒有很深入地探討,所以其機轉不是十分清楚。PTCA 的過程中當氣球撐開時會造成短暫的缺血,而氣球洩氣後血液就再灌 流回來,這樣的情形下一些與自由基( free radicals)侵害有關的局部 缺血性損害( ischemic damage)及氧化性傷害就有可能發生。以 reactive oxygen species (ROS) 為例,在一個早期針對施行氣球擴張術 後的研究中發現,在以氣球擴張破壞犬的冠狀動脈前給予 superoxide dismutase (SOD) 能完全避免早期的血管收縮及大塊狀血栓的形成 [35];Catalase, deferoxamine 和 dimethylthiourea 則沒有這樣的效用, 推測或許只有剛開始產生的過氧化物參與反應。Nunes et al. [36,37] 以氣球破壞豬的冠狀動脈 14 天後發現有 O2-產生,在手術前後給予抗 氧化劑 Vitamins C、E 可明顯地使 O2-下降,並改善血管管腔狹窄及血 管壁結構,證明在血管組織重組中還原過程的角色。文獻指出其他的 抗氧化劑如 Probucol, butylated hydroxytoluene (BHT) 在餵食高膽固 醇並施行氣球擴張術的兔子或豬的實驗中,均能減少新生內膜 (neointima)的增厚[38~42],其中 Probucol 更進一步的證實能降低 PDGF A chain、mitogen activated protein kinase (MAPK)及 Protein. 12.
(13) C-kinase 的基因,對於平滑肌細胞的增生亦有明顯的抑制作用 [41,42]。此外,在臨床試驗上,Probucol 對於減少 restenosis 的發生 的確有不錯的效果;相反地,multivitamins 則沒有作用[43~50]。 在血管內膜受傷害後所產生的氧自由基可視為第二訊息傳遞 者 : 譬 如 增 加 血 管 平 滑 肌 細 胞 DNA 合 成 以 及 原 致 癌 基 因 (pro-oncogene)c-myc 和 c-fos mRNA 的表達[51];給予抗氧化劑後 能降低人類內皮細胞上附著分子 vascular cell adhesion molecule-1 (VCAM-1)的表達[52]。 至於誘發早期過氧化物的產生,到底是機械性外力刺激下 (Arbault et,al. 以超微電極刺激纖維母細胞能活化 NADPH oxidase 產 生 H2O2 [53]。),或是和誘發 ROS 產生的生長因子相關的訊息傳遞分 子如 Extracellular signal-regulated kinase (ERKs)和 p38MAPK 所觸 發 [54~57],仍有待進一步的研究。 另一方面,因應氧化壓力產生的反應是多樣性的,以氧化還原進 程中的訊息傳遞分子來講,多數受到 thiol-disulfide 氧化還原化學反 應或易氧化還原的過渡金屬來調控。目前將焦點放在血管受損後氧化 還原啟動的訊息傳遞這方面研究並不多。Souza 等人指出,在以氣球 損傷的血管中早期被活化的轉錄因子 nuclear factor -κB (NF-κB) 能以 NADPH oxidase 抑制劑-dyphenylene iodonium 來降低其表達量. 13.
(14) [58];兔子的動脈損傷模式中,給予 Probucol 能抑制 MAP kinase 和 protein-C kinase 活化[42];Pollman 等人提出經氣球破壞的血管所引發 的 細 胞 凋 亡 ( apoptosis ) 現 象 和 氧 化 還 原 有 很 大 的 關 係 , 如 stress-activated protein kinase (SAPK)在血管受損後有數倍的表現 [59]。見圖 1,2,3。 四、動脈外膜損傷: 在血管中層受損後 3 天內,動脈外膜呈現細 胞過度增生(hyperplasia)的狀態,富含 vimentin 的纖維母細胞會逐 漸轉變成含 vimentin 及 a-SM actin 的肌纖維母細胞,在受損後 7~14 天達到最高峰[60]。而這樣的細胞轉變和一些生物活性有關,包括促 進膠原合成[61~63]及組織的收縮功能,參與疤痕的形成[64,65]。肌纖 維母細胞表現出收縮性細胞骨架蛋白(特別是 a-SM actin),被視為是 組織重組中膠原間質膠的特徵[66]。此外,動物實驗及血管內超音波 的結果中得知,包括動脈外膜損傷的緊縮性血管組織重組對於非支架 放入式冠狀動脈擴張處置後的血管管腔狹窄形成的相關性較新生內 膜形成來的大[4,7,67~69]。 五、血栓: 內皮細胞剝落後使得血管下表皮層的新生血栓成分曝 露出來,接著促進血小板附著、活化、聚集及血栓形成。首先,在施 行氣球擴張術後內皮細胞剝落處形成單層血小板;相反地,在動脈放 入支架造成深部傷害的支撐處則是形成厚且富含血小板的壁內血栓。. 14.
(15) 被活化的血小板一旦聚集會產生一些刺激血管平滑肌細胞的致 分裂原,包含PDGF、fibroblast growth factor (FGF)、epidermal growth factor (EGF)、insulin-like growth factor (IGF) 和 TGF-β1 [70,71]。IGF 和TGF-β1具調節細胞外間質的產生及組織重組的功能[72]。Bilder G et al,. 給 豬 餵 食 active PDGF-receptor tyrosine kinase 抑 制 劑 --RPR101511A,結果能減少PTCA後的內膜增生並維持較大的血管 管腔[73];此外,臨床上給病人cilostazol這種抗凝血劑,初步結果看 來能避免血管再狹窄的情形出現[74,75]。由此可推測,血小板聚集和 致分裂原參與平滑肌細胞遷移和增生以及細胞外間質的產生。 凝血酵素(thrombin)能活化血液循環中的血小板,趨化單核球細 胞並直接刺激血管平滑肌細胞增生[76~78]。在兔子施行氣球擴張術 後局部或全身給予低分子量的 heparin 可刺激並抑制平滑肌細胞增生 [79]。但是臨床試驗上全身給予 heparin 或凝血酵素抑制劑對於血管 再阻塞並無改善效果[80~84]。 六、新生內膜增生:由於血管平滑肌細胞不正常的增生和遷移以 及細胞外間質大量地產生,最終導致新生內膜增生[85]。血管修復期 中的細胞增生及再阻塞現象,主要是因應血管受到損傷之後引起的一 連串反應,如之前所述的發炎、血小板活化以及血栓。而且這段期間 可以觀察到中層血管平滑肌細胞遷移至內膜、細胞增生、細胞外間質. 15.
(16) 沉澱和血管組織重組[71]。一些複雜且多出的細胞內部訊息,如 cAMP, protein kinase C, protein kinase A 和 calcium 與能活化上述基因表現的 致分裂原和 cytokines 及促使血管平滑肌細胞進入增生狀態的細胞週 期(cell cycle)有關[86]。Synthetic phenotype 的血管平滑肌細胞和纖 維母細胞主要含有. chondroitin sulfate 和 dermatan sulfate. proteoglycans;但在組織重組的進程中,這些 proteoglycans 會轉變成 大型且纖維化的第一型膠原(typeⅠcollagen)及彈力蛋白(elastin) [71]。 血管平滑肌細胞的增生會因為在受損部位的內皮細胞再生長回 來、一氧化氮( NO)的抗增生作用以及 heparin 對這些已活化的平滑 肌細胞作用下而停止[87]。從動物模式中得知在氣球破壞後的第 21 至 56 天間內皮會產生新生現象( re-endothelialization) ;但在人體上, 根據一項對冠狀動脈置入支架後的解剖研究指出,內皮新生至少需要 3 個月[9,88]。 在兔子模式中以氣球破壞並去除內皮新生造成再狹窄的情況 下,局部給予 L-arginine 能促使血管平滑肌細胞產生一氧化氮合成? (nitric oxide synthase)來合成一氧化氮,而局部一氧化氮的增加使 得單核球的結合能力降低,也會促使巨噬細胞凋亡 [89]。這一點可 以解釋局部給予 L-arginine 為何能在施行氣球擴張術後抑制損傷的形. 16.
(17) 成。這也凸顯出前面所提的內皮剝落和正常內皮細胞鄰近調控功能喪 失二者對新生內膜增生及再阻塞形成的重要性。. 第三節. 血管平滑肌細胞受機械性壓力所引起的訊息傳遞. 機械性外力對不同組織中細胞的形態及功能的影響是不容忽視 的,尤其是心血管系統。血管平滑肌細胞的增生對於動脈粥狀硬化、 氣球擴張術後引起的再阻塞的形成均有關係,其中機械性外力是主要 的因素。 正常的成人動脈平滑肌細胞是能收縮的,並且不易被能刺激細胞 遷移和增生的生長因子或生長調節物質影響[70]。當機械性外力的刺 激出現時,原本存在於正常動脈壁中的收縮性蛋白質開始逐步地減 少 , 如 smooth mucle myosin heavy chain isoforms (α -actin 、 h-caldesmon、calponin);而本來主要存在於胎兒血管壁的 non-muscle myosin heavy chain 則在被活化的血管平滑肌細胞中表達[70,84,90], 結果產生顯著的結構重組。因此,這些細胞開始喪失收縮的功能,增 加蛋白質的分泌作用;另一方面,動脈中層平滑肌細胞受到機械性壓 力影響下引發自主調控(autocrine)及鄰近調控,使得對生長因子的反 應增加,進而刺激使動脈中層平滑肌細胞肥大(hypertrophy)或動脈壁 上的內膜增生(intimal hyperplasia) [70,91,92]。. 17.
(18) 血管平滑肌細胞到底如何感受到細胞外的機械性訊息並將此訊 息傳遞至細胞核中,最後造成基因表現上的變化,是一個很重要的課 題。近幾年的研究中證實,機械性壓力能很快的引起 PDGF receptor 的磷酸化[93] 以及 integrin a vß3 receptor [94,95]、 stretch-activated cation channels [100] 及 G proteins 的活化[96~99] ,最後都會造成 MAPK cascades 的活化[101~107]。因此,當細胞感受到機械性外力 時,protein kinase C 和 MAPKs 會被活化,導致 c-fos 和 c-jun gene [108]表現增加,同時促使轉錄因子 AP-1 對 DNA 的結合能力增加 [109]。見圖4。. 第四節 丹參的藥理作用 丹參,始載於《本經》 。為唇形科植物丹參 Salvia miltiorrhiza Bge. 的乾燥根及根莖。主產於安徽、山西、河北、四川、江蘇等地。別 名赤參、紫丹參。《本草綱目·卷十二·丹參》:“五參五色配五 臟,故人參入脾曰黃參,沙參入肺曰白參,玄參入腎曰黑參,牡蒙 入肝曰紫參,丹參入心曰赤參。”臨床處方時,則以丹參色紫紅而 常用紫丹參名。性味:苦,微寒。. 功效. 主. 治. 18.
(19) 活血化瘀. 月經不調;血滯經閉;產後腹痛;心腹痛;跌打損傷.. 涼血消癰. 溫病熱入營血;瘡癰腫毒. 養血安神. 心悸;失眠. 丹參活血化瘀作用較佳,神農本草經列為上品,古代廣泛用於 心血管、血液類之疾病。如《本草正義》所言:“丹參,專入血分, 其供在於活血行血,內之達臟腑而化瘀滯,……外之利關節而通脈 絡。”說明丹參活血化瘀的適應範圍很廣泛,能用於各科瘀血阻滯 病證。過去數十年,中國大陸對丹參做了許多現代化的研究,尤以 純化脂溶性的「丹參酮」,發現其明顯的抗血小板凝集作用,隨後 將丹參、紅花、川芎、赤芍和降香製成「冠心二號方」,廣泛用於 臨床上治療冠狀動脈心臟病,久為海內外心血管中藥研究者所熟 知。1980 年代,大陸學者從丹參根部分離純化水溶性多酚類萃取物 -丹參酸(Salvianolic acid),後經北京中國醫學科學院藥物研究所劉 耕陶教授等人,發現丹參酸在肝細胞有明顯的抗氧化作用。. 成分:丹參主含丹參酮( tanshinone) ? ? ? A、? B,異丹參酮 (isotanshinone)? ? ? , 隱丹參酮(dryptotanshinone)、異隱丹參 酮(isocryptotanshinone)、羥基丹參酮(hydroxytanshinone)? A、 丹參新酮( miltirone)、左旋二氫丹參酮(dihydrotanshinone)? ? 丹. 19.
(20) 參酚(salviol)等。此外,尚有原兒茶醛、原兒茶酸、乳酸、維生素 E 等。. 【現代藥理】 (1) 對心臟及冠狀血管的影響:丹參注射液能增加狗冠狀血流量 60 %,降低冠狀循環阻力。丹參素對離體豬冠狀動脈有擴張並增 加血流的作用,心肌收縮力先抑制後增強,心率有一定程度減 慢。但丹參酮ⅡA 磺酸鈉,原兒茶醛顯著收縮豬冠狀動脈。丹參 可減輕缺血再灌注的心肌損傷,並增加 SOD 及 GSH-Px 的活性 有效清除缺血再灌注產生的氧自由基。 (2)對心肌缺血及心肌梗塞的影響:丹參酮能明顯抑制心肌壞死導致 的嗜中性白血球溶小體釋放、吞噬和黏附,減少血清及心肌 MDA,升高心肌 SOD 的活性,抑制白血球向心肌缺血區的浸潤 及心肌中 PGE2 的合成。由此可知,丹參酮有防治心肌梗死的作 用。用丹參治療心肌梗塞的狗,可見到梗塞區壞死心肌清除較 快,巨噬細胞活躍,纖維母細胞分化和膠原纖維形成較明顯,肉 芽形成較成熟,說明丹參調節組織的修復與再生。 (3)對血液流變性的影響:丹參注射液有增加紅血球電泳率、降低血 小板黏附性、降低血球壓積、抑制血小板凝集、抑制血栓形成等 作用。降低冠心病患者的血漿粘滯性作用。丹參對老年大鼠能降 20.
(21) 低全血比粘度纖維蛋白源及增加紅血球電泳時間。 (4) 改善微循環作用:丹參有加快微循環血流速度,增加毛細血管網 而改善冠心病病人的外周微循環。 (5) 降血脂及抗動脈粥樣硬化,提高耐缺氧:對動脈粥狀硬化的家 兔,可降低血和肝中的三酸甘油酯。在細胞模型中丹參素具有降 低細胞內膽固醇合成及抗脂蛋白氧化作用,氧化脂蛋白中的 MDA 明顯減少,氧化脂蛋白對細胞的毒性作用明顯減弱。 (6) 抗菌作用:高濃度丹參煎劑的體外實驗對綠膿、大腸、傷寒、痢 疾桿菌均有抑菌作用。 (7) 鎮靜、鎮痛作用:丹參素靜脈注射對大小鼠、狗均產生明顯的鎮 靜作用。使狗腦電波型由低波轉變為高慢波。 (8)丹參注射液及其多種水溶成分(原兒茶醛、丹參素、Sod A 等), 顯著抑制肝臟脂質過氧化。其機制可能為透過自由基的清除作用 [110~112]。. 第五節. 厚朴的藥理作用. 厚朴,始載於《本經》 。為木蘭科(Magnoliaceae)落葉喬木植物厚 朴 Magnolia officinalis Rehd﹒et Wils﹒或凹葉厚朴 Magnolia officinalis Rehd﹒et Wils﹒var blioba Rehd﹒et Wils﹒ 的樹皮、根皮或枝皮。生. 21.
(22) 長於四川、湖北、湖南、浙江、陜西、貴州。以四川、湖北所產質量 最佳。其性狀為卷成單筒狀或雙筒狀,長約 35cm,厚約 2~5cm,表 面灰暗棕色,外表粗糙不平為鱗片狀,質堅硬,折斷面不平整,成纖 維狀,氣芳香,味苦澀。 《本草綱目·卷三十五·厚朴》云:“其木 質朴而皮厚,味辛烈而色紫赤,故有厚朴、烈、赤諸名。”依本草備 要解析所述,其性味為苦辛溫,具有厚腸胃,行結水,破宿血的功能。 【功效主治】 功效. 主治. 行氣、消積. 脾胃氣滯證;積滯. 行氣燥濕. 濕阻中焦證;泄瀉. 下氣、消痰平喘. 咳嗽氣喘. 厚朴味辛行散,疏利氣機,為行氣除脹要藥,凡脾胃氣滯,脕腹 脹滿者宜用之。 成分:含揮發油約 1%,油中主要成分為厚朴酚、異厚朴酚、四 氫厚朴酚。另有一種水溶性生物鹼,稱厚朴鹼。 【現代藥理】 (1) 本品煎劑在試管中,對金黃色葡萄球菌有很強的抑制作用。 (2) 厚朴鹼和厚朴酚有鬆弛骨骼肌作用。 (3) 厚朴酚對實驗性胃潰瘍有防治作用,並對組織胺所致十二指腸 22.
(23) 痙攣有一定的抑制作用。 (4) 厚朴鹼和厚朴花有降壓作用[113]。 在粥狀動脈的形成過程中,大量的脂肪尤其是氧化的脂肪酸,正 常時就會進入平滑肌層,有高膽固醇的人可能會因為平滑肌細胞受到 改變或傷害促使平滑肌增生,所有這些不正常的情形皆被認為與脂質 的過氧化有關,而氧化還原最主要的場所是在粒線體。有研究指出 magnolol 及 honokiol(厚朴成分中兩種最主要的抗氧化成分)能保護 大白鼠心臟的粒線體以防止脂質過氧化,抑制缺血再灌注引起的心室 心律不整並增加一氧化氮合成,對於 Mac-1 及 neutrophil 附著也有抑 制作用,這些都可能和 magnolol 的抗氧化作用有關[114~119]; magnolol 及 honokiol 尚具有防止血小板聚集的作用[120,121],就前面 提到血小板聚集會促進平滑肌增生。所以厚朴可能對因氣球擴張所造 成新生內膜增生的抑制作用及分子機轉,是本篇論文的研究重點之 一。. 23.
(24) 第二章. 材料. 1. 40%Acrylamide/Bis(MDBio) 2. Agarose(Sigma) 3. Ammonium persulfate (APS)(USB) 4. Ampicillin (sodium salt)(USB) 5. Anti-Human p27 kip 1(Upstate) 6. Anti-mouse IgG(horseradish peroxidase conjugated)(Pierce) 7. Anti-p21(NeoMarkers) 8. Anti-PCNA(Upstate) 9. Apoptosis sampler kit(BD) 10. Bradford 11. Bromophenol blue(USB) 12. BSA(Sigma) 13. Cell proliferation ELISA kit(Boehringer Mannheim) 14. Chloroform(Showa) 15. Chloralhydrate(Merck) 16. Cupric sulfate (anhydrous)(Sigma) 17. DEPC 18. Developer(Kodak) 19. DMEM(Biochrom). 24.
(25) 20. DMRIE-C Reagent(GIBCO) 21. DMSO(Sigma) 22. DTT 23. ECL(Amersham) 24. Ethidium bromide(USB) 25. 99.8% ethanol(NASA) 26. FBS(Biowest) 27. Fixer(Kodak) 28. Glycerol(USB) 29. Glycine(USB) 30. Haemocytometer(Boeco) 31. Heparin (ammonium salt)(Sigma) 32. Hydrogen peroxide(Showa) 33. Iron (II) sulfate heptahydrate(Showa) 34. Luciferase assay system(Promega) 35. Methanol(TEDIA) 36. Methyl-[3 H] thymidine(PerkinElmer) 37. MOPS(USB) 38. NFκB promoter 39. Paraformaldehyde(Merck) 25.
(26) 40. Phenol(Merck) 41. Pentobarbital(榮民製藥) 42. penicillin/streptomycin(GIBCO) 43. Ponceu S(Sigma) 44. Potassium dihydrogen phosphate(Merck) 45. PVDF membrane(Millipore) 46. Sodium acetate(USB) 47. Sodium chloride(Merck) 48. Sodium dihydrogen phosphate(Merck) 49. Sodium dodecyl sulfate (USB) 50. Sodium hydrogen carbonate(Merck) 51. Sodium hydroxide(島久) 52. Sodium Pyruvate(Biochrom) 53. TBA(Sigma) 54. TCA(Sigma) 55. TEMED(Pharmacia) 56. Tri Reagent(Molecular research center) 57. Tris-base(GERBU) 58. Trypan blue 59. Trypsin(GIBCO). 26.
(27) 60. Tween 20(USB) 61. Urethane(Sigma). 27.
(28) 第三章. 實驗方法. 1. 厚朴抽取物的製備 將木蘭科(Magnoliaceae)植物 Magnolia officinalis REHD. et WIL 之乾燥根皮溶於 methanol,經迴流萃取,過濾,濃縮,再以 chloroform 溶解,以 chloroform 可溶部分以水萃取,再取 chloroform 溶解以 n-hexane 萃取,將 n-hexane 層經濃縮,再結晶即可得厚朴之抽取物。 經由本方法所抽取的抽取物大部分為厚朴中之 magnolol (經 TLC 定性 的結果得知 90%為 magnolol)。. 2. 血管平滑肌細胞的培養 將含有 10% FBS 的 DMEM 培養基(含 3.7g/l NaHCO3, 4.5g/l D-Glucose, 1.028g/l N-Acetyl L-alanyl-L-glutamine, 1% Na-Pyruvate, 100 units/ml penicillin G 及 100 ug/ml streptomycin sulfate)在 37℃之 水浴恆溫槽中加溫。將儲存在液態氮中大鼠主動脈平滑肌細胞(購自 食品工業發展研究所,Cell no.:60127,代號 A10)取出後迅速放入 37℃之水浴恆溫槽中快速解凍(自液態氮容器中取出冷凍管,檢查蓋 子是否旋緊,由於熱脹冷縮過程,此時蓋子易鬆掉),以避免冰晶傷 害細胞(細胞活化後,需立刻培養,儘速放入培養基,需數日或繼代 一至二代,細胞生長及特性表現才會恢復)。以 70%酒精棉花擦拭冷 凍管外部, 移入無菌操作台內,以 1 ml pipette 將解凍之細胞懸浮液. 28.
(29) 吸出加入有培養基之離心管中(10 倍以上稀釋),混合均勻,離心 1,200 rpm, 5 分鐘,移去上清液,加入新鮮培養基,混合均勻,放入 10 公 分培養皿中,在 37℃的細胞培養箱 (95% O2, 5%CO2) 中培養,每 二天換一次培養基。當細胞長滿後,以 9 吋滴管接抽氣幫浦移除培養 皿中的培養基,用 10 ml pipette 吸取 10 ml 1X PBS 至培養皿中並稍 微搖晃後再以 9 吋滴管抽去,重複此步驟二次以維持培養皿中為中性 環境,接著以 1 ml pipette 吸取 1 ml trypsin (吸取的量以蓋過培養皿 表面為原則) 加到培養皿中,再放入細胞培養箱 5分鐘,之後加入 2 ml 培養基中和 trypsin 並以 1 ml pipette 來回沖刷培養皿上的細胞,盡量 使細胞呈單顆懸浮,將細胞懸浮液吸到 100ml 血清瓶後補加培養基 至 10 ml,以 haemocytometer 計算細胞數(細節將在下文敘述)。 注意事項: laminar flow 使用前先讓其抽氣運轉 10 分鐘,再以 70%酒 精擦拭桌面。所有進入 laminar flow 的物品均需先噴灑 70%酒精,有 蓋容器旋開瓶蓋前先過火。培養基若使用量小,則盡量置於室溫下回 溫,以避免 pH 值變化。. 3. 利用 haemocytometer 來計數細胞數目 在細胞要做實驗前,先取出 100 ul 細胞懸浮液至薄壁微量管,再 加入 100 ul trypan blue(0.06%K2HPO4, 0.8% NaCl, 0.4% trypan blue)充分混合後取出 10 ul 加到 haemocytometer,在顯微鏡放大 100 29.
(30) 倍下以計數器計算在 haemocytometer 九大格中的左上,左下,右上, 右下及中間格的細胞數總合,除以 5 (等於每一大格的平均細胞數) , 再乘以 104(haemocytometer 中的一大格體積是 1×10-4 ml),再乘以 2 (因為之前加入 trypan blue 使得細胞數為原先的二分之一) ,則可得 到原本細胞懸浮液中 1 ml 所含的細胞數。. 4. 利用[3H]thymidine incorporation assays來測定血管平滑肌細胞的 生長週期 將96 well培養盤的每一個well種 5x103細胞,放入細胞培養箱中 培養24小時,換上200ul 0.5% FBS的DMEM培養基繼續培養24小時 作同步化(Synchornization)的動作。之後分別在第0,24,30,36小時換 成200ul 15% FBS的DMEM (n=8) 培養48小時,第44小時加入2.5 uCi/ml methyl-[3H] thymidine 繼續培養4小時,接著利用細胞收集器將 細胞沖碎並轉至尼龍膜上並將膜置於抽氣中的laminar flow中隔夜,最 後將膜部分撕下放入含4ml catel溶液的計數小管中,最後把計數小管 放入機器判讀。. 5. 西方墨漬法 ( Western blot ) 將 3.5×105 的平滑肌細胞種至 6 well 培養盤的每一個 well,再以 培養基補至最終體積為 2 ml,放入細胞培養箱培養 24 小時後取出,. 30.
(31) 抽去培養基且每一個 well 以 2 ml PBS 洗一次,換上 0.5% FBS 的 DMEM 培養基繼續培養 27 小時作同步化。接著開始給藥,positive control 為 15% FBS 之 DMEM, negative control 為 0.5%FBS 之 DMEM, 丹參抽取物 Salvianolic acid B 給予劑量為 0.001mg/ml, 0.01mg/ml, 0.1 mg/ml,厚朴抽取物 Magnolol 給予劑量為 0.001mg/ml, 0.01 mg/ml, 0.05 mg/ml (n=3);接著在細胞培養箱中培養 24 小時。最 後,每一個 well 以 2 ml 1×PBS 洗一次並加入 100ul SDS sample buffer (2%SDS, 50mM DTT, 62.5mM Tris-HCl, pH6.8)萃取蛋白質,接著 將培養盤放在冰上以刮勺將細胞刮下吸至薄壁微量管,把薄壁微量管 置入乾浴器中以 95℃加熱 5 分鐘(此時原本黏稠的細胞萃取液會變的 不黏稠)。測量蛋白質濃度的步驟如下: 先將分光光度計電源打開暖機 30 分 鐘 , 設 定 波 長 為 540nm 。 取 6 個 薄 壁 微 量 管 分 別 標 示 0,5,10,15,20,25 並個別加入二次水, Bradford, Bovine serum albumin (BSA),如下表所示: 二次水 0 5 10 15 20 25. 800 ul 750 ul 700 ul 650 ul 600 ul 550 ul. Bradford. BSA. 200 ul 200 ul 200 ul 200 ul 200 ul 200 ul. 0 ul 50 ul 100 ul 150 ul 200 ul 250 ul. 待反應 5 分鐘後,依 0~25 的次序分別放入分光光度計中測吸光值, 31.
(32) 以製作 standard curve,其 R-squared 值在 0.980~0.999 之間。再取薄 壁微量管製作待測樣本:10ul sample+200ul Bradford+790ul 二次水, 同樣地等反應 5 分鐘後放入分光光度計中測吸光值,再以 standard curve 所求出的方程式換算出樣本蛋白質濃度。 接著製作 SDS-PAGE,配方如下:. ddH2O 1.5M Tris, pH8.8. 下層膠. 上層膠. 9.5 ml. 6.08 ml. 5.0 ml. 0.5M Tris, pH6.8 10% SDS 40%. 2.5 ml 200 ul. 100 ul. 5.0 ml. 1.22 ml. 100 ul. 100 ul. 20 ul. 12 ul. Acrylamide/bis 10% APS TEMED. 待膠體完全硬後,靜置 30 分鐘,之後以 150V 預跑 30~60 分鐘,再 來將在 95℃中加熱 5 分鐘的變性樣本蛋白質及蛋白質分子量標誌劑 注入至電泳槽中,以 100V 進行電泳。然後將電泳膠體上的蛋白質以 90~100V 的電壓且在 4℃的環境下轉印一小時至 PVDF 轉漬膜上 (PVDF 轉漬膜使用前先用甲醇浸泡 1 分鐘,再泡入 transfer buffer) ; 接著將轉漬膜泡入 5%低脂牛奶(5g 牛奶及 0.1ml Tween-20 溶於 100ml PBS)室溫下搖一小時;用 0.1% PBST 緩衝液( 1ml Tween-20 32.
(33) 溶於 1L PBS)每 5 分鐘洗一次,共洗三次;將轉漬膜放入有一級抗 體的封口袋中在 4℃搖一夜(此實驗中所用的抗體有 PCNA 1:1000, RIP 1:1000, Caspase-3 1:1000, Bcl-2 1:500, Bad 1:500, PARP 1:500, Fas-ligand 1:1000, TRADD 1:250, p27 1:1000, p21 1:1000, p53 1:1000) ;同樣再以 0.1% PBST 緩衝液每 5 分鐘洗一次,共洗三次,將轉漬 膜放入有二級抗體(peroxidase-conjugated anti-mouse IgG)的封口袋 中在室溫下搖一小時,以 0.1% PBST 緩衝液每 5 分鐘洗一次,共洗 三次;在暗房中將轉漬膜和 ECL 呈色劑反應一分鐘,用感光底片紀 錄訊號;最後用柯達的分析墨漬軟體分析。. 6. 評估丹參抽取物對 NFκB promoter轉殖至血管平滑肌細胞的表現 採用 Gibco 的 DMRIE-C 試劑(屬於微脂體) 。將 3.5×105 的平滑 肌細胞種至 6 well 培養盤的每一個 well,培養 15 小時。準備 12 管 薄壁微量管,6 管包含 4ul DMRIE-C+500ul 無血清培養基,另外 6 管包含 1ug NF-κB promoter-Luciferase Construc t+500ul 無血清培養 基,將不同的二管混合作用 20~30 分鐘成 lipid-DNA 複合物。移去培 養盤中的培養基,加入剛才的 lipid-DNA 複合物培養 6 小時後移去, 再換成 10% FBS 培養基繼續培養至隔天。接著給予丹參抽取物 Salvianolic acid B,給予劑量為 0.001mg/ml, 0.01mg/ml, 0.1 mg/ml, postive control 為 15% FBS 的培養基,培養 24 小時。萃取細胞的蛋. 33.
(34) 白質並以分光光度計定量出濃度(方法與西方墨漬法相同)。取 5ml 透明小管先加入 30ug 的蛋白質,再以 PBS 補體積至 20ul(置於冰 上),上冷光分析儀前才加入 50ul luciferase reagent(每一管加入 luciferase reagent 與上機的間隔時間盡量一致)來測定 luciferase 表現 量。. 7. 利用 cell proliferation ELISA(BrdU)kit 來分析丹參及厚朴抽取 物對血管平滑肌細胞 DNA 合成的影響 本實驗是採用 Boehringer Mannheim 的 cell proliferation ELISA kit。首先,在 96 well 培養盤上的每一個 well 種 1×104 的平滑肌細胞 並培養 15 小時。移去培養基後用 1× PBS 洗一遍,加入 0.5% FBS 的 培養基,繼續培養 30 小時。移去培養基後開始給藥:postive control 為 15% FBS 的培養基,negative control 為 0.5% FBS 的培養基,丹 參抽取物 Salvianolic acid B 給予劑量為 0.001mg/ml, 0.01mg/ml, 0.1 mg/ml,厚朴抽取物 Magnolol給予劑量為 0.001mg/ml, 0.01 mg/ml, 0.05 mg/ml(每一個 well 均含有 100ul 培養基),培養 20 小時後,接著加 入 10ul/well 的 BrdU(10uM)再培養 4 小時。移去培養基後加入 200ul/well FixDenat,在室溫下靜置 30 分鐘。移去 FixDenat 後加入 100 ul/well anti-BrdU-POD , 在 室 溫 下 靜 置 90 分 鐘 。 移 去. 34.
(35) anti-BrdU-POD 後用 200 ul/well 的洗滌液洗 3 次。最後加入 100 ul/well substrate 後每隔 3 秒加入 25ul/well 1M H2SO4。將培養盤放在酵素免 疫分析測讀儀上以 450nm 的波長判讀。. 8. 利用測量 Malondialdehyde(MDA)來評估丹參及厚朴抽取物對 血管大鼠血清的抗氧化能力 用 23 號針從大鼠尾動脈抽取約 3ml 血液,以 13000rpm 離心 15 分鐘將血清分離出。控制組取 15ul 的二次水+10ul 血清+10ul 10 mM CuSO4(總體積為 50ul,「Cu2+」=2~3 mM);實驗組則加 10ul 的丹 參抽取物 Salvianolic acid B 最終濃度為 0.001mg/ml, 0.01mg/ml, 0.1 mg/ml,厚朴抽取物 Magnolol最終濃度為 0.001mg/ml, 0.01 mg/ml, 0.05 mg/ml,再加入 5ul 二次水和 10ul 血清以及 10ul 10 mM CuSO4。將樣 本充分混合後放入 37℃的水浴恆溫槽中 4 小時;加入 350ul 20% TCA,混合;接著加入 350ul 0.67% TBA(溶於 0.05N NaOH) ,混合; 將樣本放入 69℃的水浴恆溫槽中 30 分鐘,最後以 10000 rpm 離心 2.5 分鐘。每個樣本取 200ul 上清液加到 96 well 培養盤中,將培養盤放 在酵素免疫分析測讀儀上以 540nm 的波長判讀。. 9. 氣球擴張破壞大白鼠總頸動脈血管的實驗模式. 35.
(36) 本實驗模式是用 350~450g 品系為 SD 的大白鼠(購自國科會) 來做氣球擴張術。先由腹腔注射 3.6% chloral hydrate(1ml/100g)麻 醉大白鼠,將大白鼠腹面朝上並用麻繩綁住四肢將大白鼠固定在解剖 板上,頭部套上裝有乙醚的 50c.c.離心管維持麻醉,擦拭少許優碘在 大白鼠頸部,用大剪刀沿大白鼠頸部中間將其胸部到下顎的皮剪開, 用鑷子將大白鼠頸部左側皮膜及一些結締組織剝離乾淨即可見左總 頸動脈,將總頸動脈及內頸動脈和外頸動脈所在位置的結締組織剝離 乾淨之後,將外頸動脈用縫線綁死結(距總頸動脈及內、外頸動脈交 會處往頭部方向約 1.5 公分) ,將內頸動脈用縫線綁活結(距總頸動 脈及內、外頸動脈交會處約 0.5 公分),用縫線繞過總頸動脈並用止 血鉗夾住縫線以斷絕血流(距總頸動脈及內、外頸動脈交會處往尾部 方向約 5 公分),用眼剪將外頸動脈剪個小洞(距綁死結處約 0.2 公 分)並稍微放血以確定有剪破血管,將 Forgaty 2F 導管自上述的小洞 插入並往總頸動脈處平推約 3 公分,打入約 1.3~1.5 大氣壓的氣體使 氣球撐大,然後來回三次破壞總頸動脈的血管壁,最後將氣球導管洩 氣並抽出,用 1ml 注射筒前接細軟管打入 heparin,在小洞與三叉處 中間綁死結,鬆開總頸動脈與內頸動脈的縫線,I.M 給予 Ampicillin, 將頸部剪開部分以縫線縫合回去,滴少許優碘後將大白鼠放回籠中待 其甦醒即完成。. 36.
(37) 10. 統計方法 數據表示採用 mean±S.E,數據統計分析採用 student-t test,當 P<0.05 時,該實驗數據有統計學上的差異。. 37.
(38) 第四章. 實驗結果. 第一節 Salvianolic acid B 和 Magnolol 對血凊刺激下平滑肌細胞 DNA 合成的影響 平滑肌細胞在經過 30 小時同步化後,在 15%血清的刺激下給 予 salvianolic acid B 和 magnolol 作用 24 小時,並在收細胞前 4 小時 加入 BrdU。結果如圖 6 所示,salvianolic acid B 對 DNA 合成的抑制 沒有影響;而 0.01 mg/ml magnolol 有些許抑制作用(P<0.05),0.05 mg/ml magnolol 則是十分顯著地對平滑肌細胞 DNA 合成有抑制作用 (P<0.01)。. 第二節 Salvianolic acid B 和 Magnolol 對血凊刺激下平滑肌細胞 PCNA 表達的影響 平滑肌細胞在經過 27 小時同步化後,在 15%血清的刺激下給予 salvianolic acid B 和 magnolol 作用 24 小時,從我們 western blot 的結 果看來,如圖 7 所示,只有 0.05 mg/ml magnolol (P<0.01)能抑制 PCNA 的表達,salvianolic acid B完全沒有影響,這一點和圖 6 的結 果類似。. 第三節 Salvianolic acid B 和 Magnolol 對血凊刺激下平滑肌細胞 RIP 表達的影響. 38.
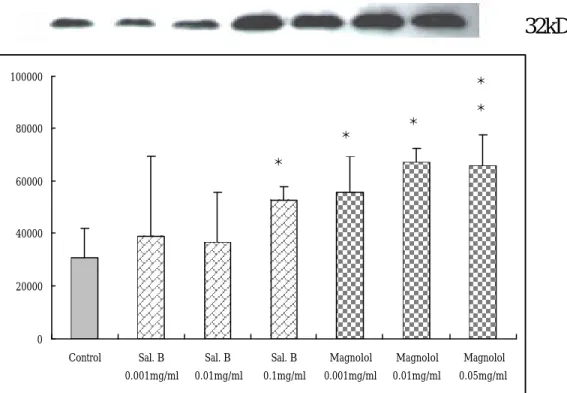
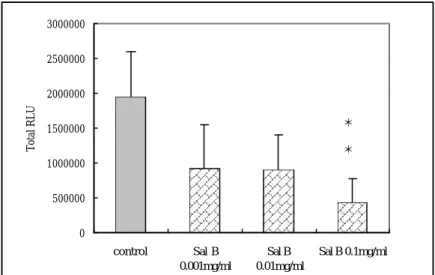
(39) 平滑肌細胞在經過 27 小時同步化後,在 15%血清的刺激下給予 salvianolic acid B 和 magnolol 作用 24 小時,從我們 western blot 的結 果看來,如圖 8 所示,salvianolic acid B 和 magnolol 能引起 RIP 的 表達,而且均呈劑量依附關係,不過仍以 magnolol 中 0.01 mg/ml (P<0.05)及 0.05 mg/ml (P<0.01)這兩種劑量最明顯 。. 第四節 Salvianolic acid B 和 Magnolol 對血凊刺激下平滑肌細胞 Caspase-3 表達的影響 平滑肌細胞在經過 27 小時同步化後,在 15%血清的刺激下給予 salvianolic acid B 和 magnolol 作用 24 小時,從我們 western blot 的結 果看來,如圖 9 所示,0.01mg/ml salvianolic acid B (P<0.05)和 magnolol 的三種劑量都能引起 Caspase-3 的表達,不過仍以 magnolol 中 0.01 mg/ml(P<0.05)及 0.05 mg/ml (P<0.01)這兩種劑量最明顯 。. 第五節 Salvianolic acid B 對轉殖至血凊刺激下平滑肌細胞的 NF-κ B promoter 的活性影響 以 liposome 的方式將 NF-κB promoter-Luciferase Construc t 轉殖 至平滑肌細胞中,在 15%血清的刺激下給予 salvianolic acid B 作用 24 小時後萃取蛋白質,測定 luciferase 表現量。結果如圖 10 所示,以 0.01mg/ml salvianolic acid B(P<0.01)對 NF-?B promoter 活性具明顯. 39.
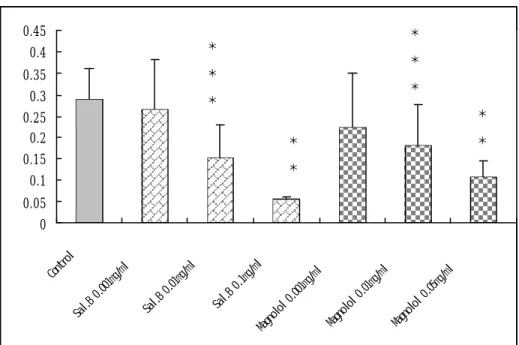
(40) 的抑制作用。. 第六節 Salvianolic acid B 和 Magnolol 在大鼠血清中的抗氧化能 力 在從血液中分離出的大鼠血清中加入 2mM CuSO4 及 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 或 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,作用 4 小時後再加入 TBA 和 TCA 產生 MDA。 從酵素免疫分析測讀儀測出 0.01mg/ml salvianolic acid B(P<0.01)及 0.05mg/ml magnolol( P<0.01)均能顯著的減少大鼠血清中 MDA 的產 生,而且有劑量依附關係,不過以 0.01mg/ml salvianolic acid B 效果 最好。. 40.
(41) 圖 1.. 41.
(42) 圖 2.. 42.
(43) 圖 3.. 43.
(44) 圖4. Model for mechanical stress initiated-signal transductions in VSMCs.. 44.
(45) 圖 5.. 45.
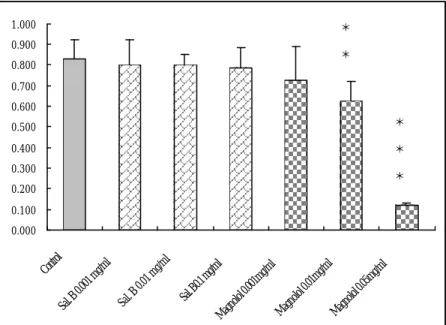
(46) 1.000. *. 0.900. *. 0.800 0.700 0.600 0.500. *. 0.400. *. 0.300. *. 0.200 0.100. Sa l.B 0.1 mg /m M l agn olo l 0. 00 1m g/m l M agn olo l0 .01 mg /m l M agn olo l 0. 05 mg /m l. 0.0 1m g/m l Sa l. B. 0.0 01 mg /m l. Sa l. B. Co ntro l. 0.000. 圖 6. Salvianolic acid B 和 Magnolol 對血凊引起平滑肌細胞 DNA 合 成的影響。所有平滑肌細胞都培養在含 15%血清的 DMEM 中,給藥 組 加 入 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 及 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,藥物作用時間 30 小時。 與 15% 血清 control group 比較起來只有 0.05mg/ml Magnolol對 BrdU incorporation 有明顯的抑制作用。 **. P < 0.01,. ***. P < 0.001 versus control group. 46.
(47) ß36kD. 1.40. sum of intensity. 1.20. *. 1.00. *. 0.80 0.60 0.40 0.20. M ag no lol 0.0 1m g/m l M agn olo l 0. 05 mg /m l. 0.0 01 mg /m l. M agn olo l. 0.1 mg /m l Sa l. B. 0.0 1m g/m l Sa l. B. 0.0 01 mg /m l. Sa l. B. Co ntr ol. 0.00. 圖 7. Salvianolic acid B 及 Magnolol 對平滑肌細胞 PCNA 表達程度的 影響。所有平滑肌細胞都培養在含 15%血清的 DMEM 中,給藥組加 入 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 及 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,藥物作用時間 24 小時。與 15% 血 清 control group 比較起來只有 0.05mg/ml Magnolol 有降低 PCNA 表 達的趨勢。 ** P < 0.01 versus control group. 47.
(48) ß74kD *. 60000. *. sum of intensity. 50000. *. *. *. 40000 30000 20000 10000. 0.1 mg /m M l agn olo l 0. 00 1m g/m l M agn olo l 0. 01 mg /m l M ag no lol 0.0 5m g/m l. Sa l. B. 0.0 1m g/m l. Sa l. B. 0.0 01 mg /m l. Sa l. B. Co ntro l. 0. 圖 8. Salvianolic acid B 及 Magnolol 對平滑肌細胞 RIP 表達程度的影 響。所有平滑肌細胞都培養在含 15%血清的 DMEM 中,給藥組加入 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 及 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,藥物作用時間 24 小時。Salvianolic acid B及 Magnolol 與 15% serum control 比較起來 RIP 的表達均有增 加,不過以 0.05mg/ml Magnolol 增加最多。 *. P < 0.05,. **. P < 0.01 versus control group. 48.
(49) ß32kD 100000. * * *. 80000. * *. 60000. 40000. 20000. 0 Control. Sal. B. Sal. B. Sal. B. Magnolol. Magnolol. Magnolol. 0.001mg/ml. 0.01mg/ml. 0.1mg/ml. 0.001mg/ml. 0.01mg/ml. 0.05mg/ml. 圖 9. Salvianolic acid B 及 Magnolol對平滑肌細胞 Caspase-3 表達程度 的影響。所有平滑肌細胞都培養在含 15%血清的 DMEM 中,給藥組 加 入 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 及 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,藥物作用時間 24 小時。 Salvianolic acid B 及 Magnolol 與 15% serum control 比 較 起 來 Caspase-3 的表達均有增加,不過以 0.05mg/ml Magnolol 增加最多。 *. P < 0.05,. **. P < 0.01 versus control group. 49.
(50) 3000000 2500000. Total RLU. 2000000 *. 1500000. *. 1000000 500000 0 control. Sal B 0.001mg/ml. Sal B 0.01mg/ml. Sal B 0.1mg/ml. 圖 10. Salvianolic acid B 對 NF-?B promoter 活性的影響。在平滑肌細 胞中加入 NF-?B promoter-Luciferase Construct 和 liposome 結合物作 transcient transfection 後,經培養24小時再加入 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B,藥物作用時間 24 小時後萃取蛋白質。將 30ug 蛋白質和 luciferase substrate 混合後測定 luciferase 表現量。結果 以 0.1mg/ml Salvianolic acid B 對 NF-?B promoter 活性具明顯的抑制 作用。 **. P < 0.01 versus control group. 50.
(51) 0.45. * *. 0.4. *. 0.35. *. 0.3. *. * *. 0.25 0.2. *. 0.15. *. *. 0.1 0.05. 0. 05 mg /m l. Ma gn ol ol. 0. 01 mg /m l. Ma gn ol ol. 0. 00 1m g/ ml. Ma gn ol ol. 0. 1m g/ ml Sa l. B. 0. 01 mg /m l Sa l. B. 0. 00 1m g/ ml. Sa l. B. Co nt ro l. 0. 圖 11. Salvianolic acid B 及 Magnolol 在大鼠血清中之抗氧化能力。在 大鼠血清中加入 2mM CuSO4 及 0.001mg/ml, 0.01mg/ml, 0.1mg/ml Salvianolic acid B 或 0.001mg/ml, 0.01mg/ml, 0.05mg/ml Magnolol,作 用 4 小時後再加入 TBA 和 TCA 產生 MDA。 Salvianolic acid B 及 Magnolol 均能顯著的減少大鼠血清中 MDA 的產生,而且有劑量依附 關係。 P < 0.01,. **. ***. P < 0.001 versus control group. 51.
(52) 第五章. 討論. 在前言中提到對於急性心肌梗塞的冠狀動脈疾病,在施行經皮穿 腔冠狀動脈血管成形術(PTCA)的介入性療法後,能對病患的症狀、 運動量以及生活品質作大幅度的改善。近年來台灣從事 PTCA 的醫院 數,心臟科專家數,以及病例數都快速增加,但是 PTCA 在動脈再狹 窄(約 30~50%)以及病人術後需進一步處置的機會仍然無法下降到 令人滿意的程度。因此尋找能降低內膜增生的治療方法是全球醫學界 共同努力的目標。 我們選用丹參萃取物 salvianolic acid B 以及厚朴萃取物 magnolol 來作為研究的藥物,主要是由於丹參本身活血化瘀的效用在中國傳統 醫學中對於冠心病的治療有相當重要的地位,現代的藥理研究也證實 其效果(參照前言第四節) ;而 salvianolic acid B 具有抗脂質氧化並 清除自由基的能力[122,123],此外能降低人的主動脈內皮細胞上的 VCAM-1 和 ICAM-1 的表達[124],所以選用來探討對氣球擴張後動 脈再狹窄的影響。厚朴則是有文獻指出 magnolol 藉其抗氧化的能力 減少大白鼠粒線體的脂質過氧化[116],對於 neutrophil 在細胞外間質 的附著也具有抑制效果,此外還能抑制心肌經缺血再灌流後所造成的 心室心律不整來保護心肌[116,117,119];而從本實驗室蔡百榮學長的 研究結果得知厚朴的確能減少內膜增生的幅度,因此我們繼續探討. 52.
(53) magnolol 對於減少內膜增生的分子機轉及途徑。 血管平滑肌細胞不正常的過度增生現象是氣球擴張術後產生 restenosis 的主要形成原因之一。首先就 salvianolic acid B 及 magnolol 在 in vitro 中對血清刺激下的平滑肌細胞增生現象是否會產生抑制作 用來探討。我們利用 PCNA 這個核蛋白來作為觀察指標:因為 PCNA 是 DNA polymerase d 的輔助因子,幫助 DNA polymerase d 和 DNA 結 合並促進 DNA 複製時的 DNA 合成;PCNA 的表達量在細胞週期中 的 S 期達到最高峰,而且在其他時期不易偵測[125~128];另一方面, 我們使用 Cell proliferation ELISA kit,偵測和細胞 DNA 合成時所需 Thymidine 結構相近的 5-Bromo-2’-deoxyuridine(BrdU)的含量,其 目的和 PCNA 一樣都是用來觀察藥物對 DNA 合成的影響。最近的研 究中指出 magnolol能抑制 COLO-205和 Hep-G2這兩種癌細胞的 DNA 合成[129]。從我們的結果中得知高劑量的 magnolol(0.05 mg/ml)對 於抑制血清刺激下的平滑肌細胞 DNA 合成同樣有明顯的效果,而 salvianolic acid B 則完全沒有影響。 前面提到由於血管平滑肌細胞不正常的增生是氣球擴張術後產 生 restenosis 的主要原因之一,所選擇的治療藥物除了直接抑制細胞 的 DNA 合成之外,還可以從誘引細胞進行 apoptosis 使 restenosis 的 進程中細胞數目維持恆定的方向著手。從一些研究指出 magnolol 在. 53.
(54) COLO-205和 Hep-G2這兩株癌細胞中會增加 p21, caspase-3, caspase-8, caspase-9 這些蛋白質的表達並抑制 bcl-2,促使 cytochrome c 從粒線 體轉移到細胞質液中,增加細胞質液中游離態 Ca2+,表示 magnolol 會引起細胞進行 apoptosis[130]。而 salvianolic acid B 同樣有研究指出 在餵食膽固醇混合水溶性丹參萃取物的兔子施行氣球擴張術後進行 apoptosis 的細胞數目比正常組和餵食膽固醇組施行氣球擴張術增加 許多並伴隨著 p53 的表達,內膜新生增厚面積也明顯減少[131]。由 我們從平滑肌細胞作有關 apoptosis 的相關蛋白質表達上得知,在血 清刺激下的平滑肌細胞給予 salvianolic acid B 和 magnolol比起單純只 有血清刺激下的平滑肌細胞均能明顯地增加 receptor-interacting protein(RIP)及 caspase-3 的表達(見圖 5) ,但這只是在蛋白質層面 上,仍需要從別的 apoptosis 特徵來求証。 NF-κB 是一種與 DNA 結合的轉錄因子,分為 p50 和 p65 這兩 種次單元之蛋白質複合物;在調節免疫及發炎反應、胚胎及細胞分化 之發育連繫、細胞凋亡、細胞週期及腫瘤生成(oncogenesis)上扮演 重要的角色。已有研究指出 NF-κB 在施行氣球擴張術後會開始活 化,而且當以抑制劑減弱 NF-κB/Rel 的活性及抑制 NF-κB promoter transactivation 可有效減少血管平滑肌細胞增生並且預防新生內膜形 成[132,133];在由 TNF-α引起的血管平滑肌細胞遷移的實驗中,去. 54.
(55) 除 NF-κB 的活性可減弱 TNF-α引起的血管平滑肌細胞遷移作用 [134];另外一項研究指出,由 NO 引起的細胞凋亡與 protein kinase C signaling 及 NF-κB 結合活性的減少有關[135]。從這些研究中得知 NF-κB 對於 restenosis 的形成相當重要。由我們 Luciferase assay 的結 果看來,高劑量的 salvianolic acid B(0.1 mg/ml)對於 NF-κB promoter 的活性有十分顯著的抑制作用,由此可推測 salvianolic acid B 可能經 由減少 NF-κB 的表達促使細胞進行 apoptosis,見圖 5。 氧化低密度脂蛋白(Oxidatively modified LDL,ox-LDL)是動脈硬 化形成和氣球擴張術後血管再狹窄的主要致病因素之一。最重要的是 它被巨噬細胞的吸入是無控制性的,它的細胞毒性,和它的趨化性。 細 胞 或 無 細 胞 系 統 的 LDL 氧 化 主 要 是 經 由 一 個 自 由 基 過 程 [136~139],在此,LDL 的 polyunsaturated fatty acid(PUFAs)會被脂 質過氧化作用(lipid peroxidation)轉換成脂氫過氧化物,然後再被分解 成 不 同 的 產 物 , 包 括 活 躍 的 乙 醛 如 malondialdehyde( MDA), 4-hydroxynonenal (4-HNE),hexanal 和很多其它的。多方面的研究提 議出這些 lipid peroxidation 在 LDL 粒子內所產生的產物會和 Apo-B 氨基酸基作共價鍵的結合;例如離氨基酸(lysine)基的 e-氨基 groups, 藉以產生使 Apo-B 對巨噬細胞清道夫接受器(macrophage scavenger receptors)有高度親和力的修飾 [138,139]接 而形成泡沫細胞,同時. 55.
(56) oxLDL 和其產物也會吸引血液中的單核球和血管中層的平滑肌細胞進 入血管內皮下空間 。 Ox-LDL 的細胞毒性也有可能由乙醛 lipid peroxidation 的產物傳達到某一些程度[138]。HNE,2-alkenal,和 2,4 alkadienals 在 mmol 的濃度下對內皮細胞有細胞毒性[140]。因此從血 管內膜的 ox-LDL 擴散出來的乙醛會導至血管系統內皮細胞之破壞, 而加速動脈硬化病變或血管再阻塞病變的形成。這些發現,和 ox-LDL, 以及 MDA 和 HNE 修飾 LDL 出現在粥狀動脈硬化病灶的發現 [141~143],指出 lipid peroxidation 和乙醛性 lipid peroxidation 的產物 在致粥瘤性啟始作用過程中(包括刺激平滑肌細胞增生)扮演著一個 重要的角色[144];血清中的 MDA-LDL 對於氧化壓力來說可當作是 測量指標,而且在一項臨床研究中,MDA 在病患施行 PTCA 將氣球 洩氣後 60 秒會明顯增加[145]。另一方面,ox-lipoprotein-A 也被証實 能引起內皮細胞中 PDGF-B 的表達[146]。在前言中提到在血管內膜 受傷害後所產生的氧自由基可視為第二訊息傳遞者:譬如增加血管平 滑肌細胞 DNA 合成以及原致癌基因 c-myc 和 c-fos mRNA 的表達。 基於上面的理由,我們加入和細胞實驗中相同濃度的 magnolol 和 salvianolic acid B 至含 2mM CuSO4 的大鼠血清中觀察是否對血清中 MDA 的產生發揮抑制作用,結果發現二者都能顯著地降低 MDA 的 形成,而且呈 dose-dependent manner。. 56.
(57) 第六章. 參考文獻. [1] Faxon DP.(1993) Restenosis after angioplasty: what have we learned from clinical trail? Cardiol Rev 1:209-217. [2] Kuntz R.E., Baim D.S.(1993) Defining coronary restenosis: newer clinical and angiographic paradigms. Circulation 88:1310-1323. [3] Serruys P.W., Foley D.P., (1993) Kirkeeide R.L.,King S.B. Restenosis revisited: insights provided by quantitative coronary angioplasty. Am Heart J 126:1243-1267. Editorial. [4] MJ Post, C Borst, and RE Kuntz. (1994) The relative importance of arterial remodeling compared with intimal hyperplasia in lumen renarrowing after balloon angioplasty. A study in the normal rabbit and the hypercholesterolemic Yucatan micropig. Circulation 89:2816-2821. [5] Antoine Lafont, Luis A. Guzman, Patrick L. Whitlow, Marlene Goormastic, J. Fredrick Cornhill, and Guy M. Chisolm. (1995) Restenosis After Experimental Angioplasty : Intimal, Medial, and Adventitial Changes Associated With Constrictive Remodeling. Circ Res 76:996-1002. [6] Yi Shi, Marc Pieniek, Ali Fard, James O'Brien, John D. Mannion, and Andrew Zalewski. (1996) Adventitial Remodeling After Coronary Arterial Injury. Circulation 93:340-348. [7] Henning Rud Andersen, Michael Mæng, Martin Thorwest, and Erling Falk. (1996) Remodeling Rather Than Neointimal Formation Explains Luminal Narrowing After Deep Vessel Wall Injury : Insights From a Porcine Coronary (Re)stenosis Model. Circulation 93:1716-724. [8] Schwartz RS. Edwards WD. Huber KC. Antoniades LC. Bailey KR. Camrud AR. Jorgenson MA. Holmes DR Jr. (1993) Coronary restenosis: prospects for solution and new perspectives from a porcine model.Mayo Clinic Proceedings. 68:54-62. [9] Carter AJ. Laird JR. Farb A. Kufs W. Wortham DC. Virmani R. (1994) Morphologic characteristics of lesion formation and time course of smooth muscle cell proliferation in a porcine proliferative restenosis model. J Am Coll Cardiol 24:1398-405. [10] Lam JY. Chesebro JH. Steele PM. Dewanjee MK. Badimon L. Fuster V. (1986) Deep arterial injury during experimental angioplasty: relation to a positive indium-111-labeled platelet scintigram, quantitative platelet deposition and mural thrombosis. J Am Coll 57.
(58) Cardiol 8:1380-6. [11] Ip JH. Fuster V. Israel D. Badimon L. Badimon J. Chesebro JH. The role of platelets, thrombin and hyperplasia in restenosis after coronary angioplasty. J Am Coll Cardiol 1991;17(6 Suppl B):77B-88B [12] Meyer BJ. Fernandez-Ortiz A. Mailhac A. Falk E. Badimon L. Michael AD. Chesebro JH. Fuster V. Badimon JJ. Local delivery of r-hirudin by a double-balloon perfusion catheter prevents mural thrombosis and minimizes platelet deposition after angioplasty. Circulation 1994;90:2474-80. [13] Crowley ST. Ray CJ. Nawaz D. Majack RA. Horwitz LD. Multiple growth factors are released from mechanically injured vascular smooth muscle cells. Am J Physiol 1995;269(5 Pt 2):H1641-7, [14] Ward Casscells, MD Migration of smooth muscle and endothelial cells. Critical events in restenosis Circulation 86;3:723-9 [15] Schwartz SM, Heimark RL, Majesky MW. Developmental mechanisms underlying pathology of arteries. Physiol Rev 1990;70:177-209. [16] Clowes AW, Reidy MA J Vasc Surg 1991;13:885-891. [17] Austin GE, Ratliff NB, Hollman J, Tabei S, Phillips DF. Intimal proliferation of smooth muscle cells as an explanation for recurrent coronary artery stenosis after percutaneous transluminal coronary angioplasty. J Am Coll Cardiol 1985; 6:369 –375. [18] Uchida Y, Hasegawa K, Kawamura K, Shibuya I. Angioscopic observation of the coronary luminal changes induced by percutaneous transluminal coronary angioplasty. Am Heart J 1989;117:769 –776. [19] Block PC. Restenosis after percutaneous transluminal coronary angioplasty— anatomic and pathophysiological mechanisms: strategies for prevention. Circulation 1990;81:IV2–IV4. [20] Farb A, Sangiorgi G, Carter AJ, Walley VM, Edwards WD, Schwartz RS, Virmani R. Pathology of acute and chronic coronary stenting in humans. Circulation 1999;99:44 –52. [21] Rogers C, Parikh S, Seifert P, Edelman ER. Endogenous cell seeding: remnant endothelium after stenting enhances vascular repair. Circulation 1996; 94:2909 –2914. [22] Nugent HM, Rogers C, Edelman ER. Endothelial implants inhibit intimal hyperplasia after porcine angioplasty. Circ Res 58.
(59) 1999;84:384 –391. [23] Gajdusek C, Carbon S, Ross R, Nawroth P, Stern D. Activation of coagulation releases endothelial cell mitogens. J Cell Biol 1986;103:419-428. [24] Koo EWY, Gotlieb AI Lab Invest 1991;64:743-750. [25] Wight TN Arteriosclerosis 1989;9:1-20. [26] Dzau VJ, Gibbons GH Hypertension 1991;18:15-21. [27] Antonelli-Orlidge A, Saunders KB, Smith SR, D’Amore PA. An activated form of transforming growth factor ß is produced by cocultures of endothelial cells and pericytes. Proc Natl Acad Sci USA 1989;86:4544-4548. [28] Herman IM, Castellot JJ Jr Arteriosclerosis 1987;7:463-469. [29] Neumann FJ, Ott I, Gawaz M, Puchner G, Schomig A. Neutrophil and platelet activation at balloon-injured coronary artery plaque in patients undergoing an-gioplasty. J Am Coll Cardiol 1996;27:819 –824. [30] Kornowski R, Hong MK, Tio FO, Bramwell O, Wu H, Leon MB. In-stent restenosis: contributions of inflammatory responses and arterial injury to neoin-timal hyperplasia. J Am Coll Cardiol 1998;31:224 –230. [31] Rogers C, Welt FG, Karnovsky MJ, Edelman ER. Monocyte recruitment and neointimal hyperplasia in rabbits: coupled inhibitory effects of heparin. Arterioscl Thromb Vasc Biol 1996;16:1312–1318. [32] Rogers C, Edelman ER, Simon DI. A mAb to the beta2-leukocyte integrin Mac-1 (CD11b/CD18) reduces intimal thickening after angioplasty or stent implantation in rabbits. Proc Natl Acad Sci USA 1998;95:10134 –10139. [33] Simon DI, Chen Z, Seifert P, Edelman E, Ballantyne CM, Rogers C. Markedly reduced neointimal thickening in Mac-1 (CD11b/CD18)-deficient mice after carotid artery dilation and endothelial denudation (abstract). Circulation 1998;98:I-238. [34] Buffon A, Liuzzo G, Biasucci LM, Pasqualetti P, Ramazzotti V, Rebuzzi AG, Crea F, Maseri A. Preprocedural serum levels of C-reactive protein predict early complications and late restenosis after coronary angioplasty. J Am Coll Cardiol 1999;34:1512–1521. [35] Laurindo FRM, da Luz PL, Uint L et al. Evidence for superoxide radical-dependent coronary vasospasm after angioplasty in intact 59.
(60) dogs. Circulation 1991;83:1705–1715. [36] Nunes GL, Sgoutas DS, Redden RA et al. Combinations of vitamins C and E alters the response to coronary balloon injury in the pig. Arterioscler Thromb Vasc Biol 1995;15:156–165. [37] Nunes GL, Robinson K, Kalynych A et al. Vitamins C and E inhibit O2-production in pig coronary arteries. Circulation 1997;89:3593- 3601. [38] Ferns GA, Forster L, Stewart-Lee A et al. Probucol inhibits neointimal thickening and macrophage accumulation after balloon injury in the cholesterol-fed rabbit. Proc Natl Acad Sci USA 1992; 89:11312–11316. [39] Freyschuss A, Stiko RA, Swedenborg J et al. Antioxidant treatment inhibits the development of intimal thickening after balloon injury of the aorta in hypercholesterolemic rabbits. J Clin Invest 1993;91:1282–1288. [40] Schneider JE, Berk BC, Gravanis MB et al. Probucol decreases neointimal formation in a swine model of coronary artery balloon injury. A possible role for antioxidants in restenosis. Circulation 1993;88:627–628. [41] Miyauchi K, Aikawa M, Tani T et al. Effect of probucol on smooth muscle cell proliferation and dedifferentiation after vascular injury in rabbits: possible role of PDGF. Cardiovasc Drugs Ther 1998;12:251–260. [42] Tanaka K, Hayashi K, Shingu T et al. Probucol inhibits neointimal formation in carotid arteries of normocholesterolemic rabbits and the proliferation of cultured rabbit vascular smooth muscle cells. Cardiovasc Drugs Ther 1998;12:19–28. [43] Tardif JC, Cote G, Lesperance J et al. Probucol and multivitamins in the prevention of reestenosis after coronary angioplasty. N Engl J Med 1997;337:365–372. [44] Lee YJ, Daida H, Yokoi H et al. Effectiveness of probucol in preventing restenosis after percutaneous transluminal coronary angioplasty. Jpn Heart J 1996;37:327–332. [45] Setsuda M, Inden M, Hiraoka N et al. Probucol therapy in the prevention of restenosis after successful percutaneous transluminal coronary angioplasty. Clin Ther 1993;15:374–382. [46] Yokoi H, Daida H, Kuwabara Y et al. Effectiveness of an 60.
(61) anti-oxidant in preventing restenosis after percutaneous transluminal coronary angioplasty: the Probucol Angioplasty Restenosis Trial. J Am Coll Cardiol 1997;30:855–862. [47] Watanabe K, Sekiya M, Ikeda S, Miyagawa M, Hashida H. Preventive effects of probucol on restenosis after percutaneous transluminal coronary angioplasty. Am Heart J 1996;132:23–29. [48] O’Keefe Jr. JH, Stone GW, McCallister Jr. BD et al. Lovastatin plus probucol for prevention of restenosis after percutaneous transluminal coronary angioplasty. Am J Cardiol 1996;77:649–652. [49] Sekiya M, Funada J, Watanabe K, Miyagawa M, Akutsu H. Effects of probucol and cilostazol alone and in combination on frequency of post-stenting restenosis. Am J Cardiol 1998;82:144–147. [50] Rodes J, Cote G, Lesperance J et al. Prevention of restenosis after angioplasty in small coronary arteries with probucol. Circulation 1998;97:429–436. [51] Rao, G. N.; Berk, B. C. Active oxygen species stimulate vascular smooth muscle cell growth and proto-oncogene expression. Circ. Res. 1992;70:593–599. [52] Marui, N.; Offermann, N. K.; Swerlick, R.; Kunsch, C.; Rosen, A.; Ahmad, M.; Alexander, R. W.; Medford, R. M. Vascular cell adhesion molecule-1 (VCAM-1) gene transcription and expres-sion are regulated through an antioxidant-sensitive mechanism in human vascular endothelial cells. J. Clin. Invest. 1993;92:1866–1874. [53] Arbault S, Pantano P, Sojic N et al. Activation of the NADPH oxidase in human fibroblasts by intrusion of a single cell with an ultramicroelectrode. Carcinogenesis 1997;18:569–574. [54] Abe JI, Berk BC. Reactive oxygen species as mediators of signal transduction in cardiovascular disease. Trends Cardiovasc Med 1998;8:59–63. [55] Susuki YJ, Forman HJ, Sevanian A. Oxidants as stimulators of signal transduction. Free Radic Biol Med 1997;22:269–285. [56] Sen CK, Packer L. Antioxidant and redox regulation of gene expression. FASEB J 1996;10:709–720. [57] Kunsch C, Medford RM. Oxidative stress as a regulator of gene expression in the vasculature. Circ Res 1999;85:753–766. [58] Souza HP, Souza LC, Anastacio VM, et al. Vascular oxidant stress early after balloon injury. Evidence for increased NAD(P)H oxidoreductase activity. Free Radic Biol Med 2000;in press. 61.
(62) [59] Pollman MJ, Hall JL, Gibbons GH. Determinants of vascular smooth muscle cell apoptosis after balloon angioplasty injury. Influence of redox state and cell phenotype. Circ Res 1999;84:113–121. [60] Yi Shi, Marc Pieniek, Ali Fard, James O'Brien, John D. Mannion, and Andrew Zalewski Adventitial Remodeling After Coronary Arterial Injury. Circulation 1996;93: 340-348 [61] Hogeman B, Gillessen A, Bocker W, Rauterberg J, Domschke W. Myofibroblast-like cells produce mRNA for type I and III procollagens in chronic active hepatitis. Scand J Gastroenterol.. 1993;28:591-594. [62] Zhang K, Rekhter MD, Gordon D, Phan SH. Myofibroblasts and their role in lung collagen gene expression during pulmonary fibrosis: a combined immunohistochemical and in situ hybridization study. Am J Pathol.. 1994;145:114-125. [63] Kirk TZ, Mark ME, Chua CC, Chua BH, Mayes MD. Myofibroblasts from scleroderma skin synthesize elevated levels of collagen and tissue inhibitor of metalloproteinase (TIMP-1) with two forms of TIMP-1. J Biol Chem.. 1995;270:3423-3428. [64] Darby I, Skalli O, Gabbiani G. a-Smooth muscle actin is transiently expressed by myofibroblasts during experimental wound healing. Lab Invest.. 1990;63:21-29. [65] Sappino AP, Schrürch W, Gabbiani G. Differentiation repertoire of fibroblastic cells: expression of cytoskeletal proteins as marker of phenotypic modulation. Lab Invest.. 1990;63:144-161. [66] Arora PD, McCulloch CAG. Dependence of collagen remodelling on a-smooth muscle actin expression by fibroblasts. J Cell Physiol.. 1994;159:161-175. [67] Lafont A, Libby P. The smooth muscle cell: sinner or saint in restenosis and the acute coronary syndromes? J Am Coll Cardiol 1998;32:283–285. [68] Kakuta T, Currier JW, Haudenschild CC, Ryan TJ, Faxon DP. Differences in compensatory vessel enlargement, not intimal formation, account for restenosis after angioplasty in the hypercholesterolemic rabbit model. Circulation 1994;89: 2809–2815. [69] Mintz GS, Kent KM, Pichard AD, Satler LF, Popma JJ, Leon MB. Contribution of inadequate arterial remodeling to the development of focal coronary artery stenoses: an intravascular ultrasound study. 62.
(63) Circulation 1997;95:1791–1798. [70] Ross R. The pathogenesis of atherosclerosis: A perspective for the 1990s. Nature 1993;362:801-809. [71] Forrester JS, Fishbein M, Helfant R, Fagin J. A paradigm for restenosis based on cell biology: clues for the development of new preventive therapies. J Am Coll Cardiol 1991;17:758 –769. [72] Chen JK, Hoshi H, McKeehan WL. Transforming growth factor type beta specifically stimulates synthesis of proteoglycan in human adult arterial smooth muscle cells. Proc Natl Acad Sci USA 1987;84:5287–5291. [73] Bilder G, Wentz T, Leadley R, Amin D, Byan L, O’Conner B, Needle S, Galczenski H, Bostwick J, Kasiewski C, et al. Restenosis following angioplasty in the swine coronary artery is inhibited by an orally active PDGF-recepto tyrosine kinase inhibitor, RPR101511A. Circulation 1999;99:3292–3299. [74] Yamasaki M, Hara K, Ikari Y, Kobayashi N, Kozuma K, Ohmoto Y, Oh-Hashi Y, Ako J, Nakajima H, Chiku N, et al. Effects of cilostazol on late lumen loss after Palmaz-Schatz stent implantation. Cathet Cardiovasc Diagn 1998;44:387–391. [75] Tsuchikane E, Fukuhara A, Kobayashi T, Kirino M, Yamasaki K, Kobayashi T, Izumi M, Otsuji S, Tateyama H, Sakurai M, Awata N. Impact of cilostazol on restenosis after percutaneous coronary balloon angioplasty. Circulation 1999; 100:21–26. [76] Bar-Shavit R, Kahn A, Wilner GD, Fenton JW II. Monocyte chemotaxis: stimulation by specific exosite region in thrombin. Science 1983;220:728 –731. [77] McNamara CA, Sarembock IJ, Gimple LW, Fenton JW II, Coughlin SR, Owens GK. Thrombin stimulates proliferation of cultured rat aortic smoothmuscle cells by a proteolytically activated receptor. J Clin Invest 1993;91:94 –98. [78] Bar-Shavit R, Benezra M, Eldor A, Hy-Am E, Fenton JW II, Wilner GD, Vlodavsky I. Thrombin immobilized to extracellular matrix is a potent mitogen for vascular smooth muscle cells: nonenzymatic mode of action. Cell Regul 1990;1:453– 463. [79] Oberhoff M, Novak S, Herdeg C, Baumbach A, Kranzhofer A, Bohnet A, Horch B, Hanke H, Haase KK, Karsch KR. Local and systemic delivery of low molecular weight heparin stimulates the reendothelialization after balloon angioplasty. Cardiovasc Res 63.
數據


+3
相關文件
The buffer cell sees very different optical influence from different neighboring cells. Center features almost remain
Write three nuclear equations to represent the nuclear decay sequence that begins with the alpha decay of U-235 followed by a beta decay of the daughter nuclide and then another
H&E 10x view showing vascular stroma with proliferation of spindle cells intermixed with extravasated RBCsb. H&E 40x view showing anastomosing vascular channels lined by
stained negative for a-smooth muscle actin because the main body of the tumor consisted of epithelioid or epithe- lial cells that contained few positive smooth muscle cells.. We
Stain (-) because the main body of the tumor consisted of epithelioid or epithelial cells that contained few positive smooth muscle cells. No clear ductal structures or
Toxic water from cars and factories causes acid rain.. Serious research on acid rain only started around 50
2.滴加蒸餾水或亞甲藍液製成水埋標本 3.觀察 石細胞 (厚壁細胞)及
Parry,“ Effect of temperature on growth, chemical composition and fatty acid composition of tropical Australian microalgae grown in batch cultures, ” Aquaculture, Vol.211, No.1-4,
