BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS240, 51 – 56 (1997)
ARTICLE NO.RC977600
Characterization of
g
S-Crystallin Isoforms from Lip Shark
(Chiloscyllium colax): Evolutionary Comparison
between
g
S and
b
/
g
Crystallins
1
Fu-Ming Pan,* Ming-Hong Chuang,† and Shyh-Horng Chiou*
,†
,2Laboratory of Crystallin Research, †Institute of Biochemical Sciences, National Taiwan University,
and *Institute of Biological Chemistry, Academia, P.O. Box 23-106, Taipei, Taiwan
Received September 30, 1997
The abundant presence of various common and
spe-gS-Crystallin from shark eye lenses, formerly termed
cific classes of structurally conserved proteins in eye
bs crystallin in mammalian lenses, is structurally char-
lenses (crystallins) of different species of vertebrates
acterized in this study by cDNA cloning and sequencing.
constitutes a good model system to unravel the complex
To facilitate sequence characterization of gS-crystallin
process of evolution in structurally homologous
pro-possessing intermediate structural properties between
teins (1,2). Fish represents the oldest and most diverse
b- and g-crystallins, cDNA mixture was constructed
group of vertebrates. The modern fishes comprise two
from the poly(A)
/mRNA isolated from shark eye lenses,
major classes of piscine species: [1] Osteichthyes or
te-and amplification by polymerase chain reaction (PCR)
leostean (bony) fishes and [2] Chondrichthyes or
carti-was carried out to obtain nucleotide segments encoding
laginous fishes (sharks and skates).
multiple shark gS-crystallins. Sequencing several posi-
The study of lens crystallins from the lowest piscine
tive clones revealed that a multiplicity of isoforms exists
class is of special interest from the evolutionary point
in the gS-crystallin class of this cartilaginous fish,
simi-of view because they constitute the early protein forms
lar to authentic g-crystallin family characterized from
of vertebrates and are thought to have been ancestral
the same shark species. Comparison of protein
se-to those of land vertebrates. Sharks diverged from the
quences encoded by two representative shark gS1 and
Placodermi (one class of armored fishes) long before the
gS2 cDNAs with those published sequences of b-, g-, and
appearance of modern bony fishes and amphibians (3).
gS crystallins from bovine, human, bullfrog and carp
The characterization of shark crystallins would be
lenses indicated that there is about 35–64% sequence
deemed very important for the phylogenetic
compari-homology between shark gS crystallins and structurally
son in light of the recent elucidation of the complete
related crystallins from different evolutionary classes,
sequences of
g
-crystallins from several species of
tele-with a higher sequence similarity between shark gS and
ostean fishes in Osteichthyes (4 – 6).
mammalian g-crystallins than that of shark gS and carp
The present study was performed in the endeavor to
gS or bovine gS crystallins. A phylogenetic tree
con-have a structural characterization of one unique class
structed on the basis of the sequence divergence among
of lens crystallins with their primary structure lying
various b-, g-, and gS crystallins corroborates the closer
between the well-known
b
- and
g
-crystallins. This class
relatedness of shark gS to authentic g-crystallin than
of crystallin, formerly called
b
S and now renamed
g
S
to mammalian and teleostean gS crystallins. It further
crystallin (7,8), exists as a monomeric protein which is
strengthens the supposition that ancestral precursors
similar to the major authentic
g
-crystallins. However
of gS-crystallins were present in the shark lens long
before the appearance of present-day teleostean and
unlike
g
-crystallins which possess a free N-terminal
mammalian gS-crystallins.
q 1997 Academic Pressamino-acid residue,
g
S-crystallin has a blocked N
ter-minus as most members of
b
-crystallin family. In this
report we have for the first time cloned and sequenced
g
S crystallins from the shark eye lenses, which possess
some structural properties distinguishable from those
1The sequence data of cDNAs for sharkgS-crystallins have been
deposited in the EMBL Data Library under the Accession Numbers
of the existing
g
S-crystallins characterized from
mam-X79226 and X79227 forgS1 andgS2, respectively.malian species. The results indicate that
g
S crystallin
2Corresponding address: Institute of Biochemical Sciences,
Na-characterized from the lower class of cartilaginous fish
tional Taiwan University, P.O. Box 23-106, Taipei, Taiwan. Fax:Vol. 240, No. 1, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Nucleotide and deduced protein sequences of sharkgS-1 (A) and sharkgS-2 (B) crystallins. In (A) the nucleotide sequence of 522-base pairs is shown above the amino-acid sequence of 173 residues including translation initiation methionine; in (B) the nucleotide sequence comprises 524-base pairs encoding a protein sequence of 173 amino acids. Asterisks (*) are indicated in every 10-nucleotide segment for easy tracing of sequence contents. Amino acids are denoted by one-letter symbols. The 5* and 3* nucleotide segments used as primers for PCR reactions are underlined.
cDNA amplification by PCR, cloning and sequencing ofg
S-crys-mammalian class, consists of a multitude of isoforms,
tallin isoforms. Two oligonucleotide primers of sense and antisense
and are more related to
g
-crystallin than
g
S or
b
-crys-orientations, covering 5*- and 3*-nucleotide coding regions forN- and
tallins based on sequence alignment and phylogenetic
C-terminal 4-6 amino-acid segments of the previously determined
comparison.
cDNA sequence for one carp gS-crystallin (10), with the forwardsequence, 5*-CATGGGCAAG(A/G)TCA(T/C)CTT(C/T)-3* (19-mer) and the reverse sequence, 5*-C(A/G)TCACTCCA(T/C)(G/A)A(T/
MATERIALS AND METHODS
G)GCG-3* (17-mer) (with slant lines indicating use of degenerate codons in the primers) were synthesized. The conditions for PCR
Isolation of mRNA from shark lenses. The small shark
(Chiloscyl-reactions were similar to the previous reports for cDNA amplification
lium colax, brownbanded bambooshark or brown-spotted catshark
of teleostean lenses (5,6), i.e. subjecting to 40 cycles of heat denatur-as commonly called) of less than 1-year-old wdenatur-as obtained from a local
ation at 947C for 2.5 min, annealing the primers to the DNAs at 487C aquarium shop under a special contract for scientific research. Shark
for 1 min and 20 sec and running DNA chain extension with Taq lenses were removed and stored in liquid-nitrogen container
immedi-polymerase at 727C for 3 min, followed by a final extension at 72 7C ately after they were dissected and before the processing for mRNA
for 10 min. Products were treated with Klenow Fragment and T4 isolation. Two deep-frozen lenses from one shark were homogenized
polynucleotide kinase, and separated on a 1.2 % agarose gel and and RNA was extracted according to the standard cloning manual
electroeluted according to standard procedures. The DNA fragments of Maniatis et al. (9). To obtain a full-length crystallin cDNA,
were subcloned into pUC18 previously digested with SmaI/BAP, and poly(A)/RNA was purified using QuickPrep mRNA preparation kit
then transformed into E. coli strain JM 109. Plasmids purified from (Pharmacia, Uppsala, Sweden) and then subjected to the synthesis
positive clones were prepared for nucleotide sequencing by dideoxy-of cDNA mixture by cDNA Synthesis System/Plus kit (Amersham,
deter-Vol. 240, No. 1, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1—Continued
mined by conventional isotope-labeled manual method was double- align highly evolved gene families that have clear evolutionary rela-checked by automatic fluorescence-based sequencing of templates tionships.
amplified by PCR using model 373A DNA sequencing System (Ap-plied Biosystems Inc., CA, USA) with a Taq DyeDeoxy terminator cycle sequencing kit (Applied Biosystems Inc.).
RESULTS AND DISCUSSION
Sequence comparison of deduced sharkgS-crystallins and homol-ogy search. A commercially available software package (DNASTAR Inc., Madison, WI, U.S.A.) was used for the estimation of sequence
There were sharks in the oceans of earth long before
homology based on percent sequence identity (5).the first animals had begun to colonize the land
sur-Construction of a phylogenetic tree forb-,g- andgS-crystallins of
face. Their history stretches back for at least 700
mil-vertebrate species. A software package of LaserGene for the Apple
lion years, a vast period of time as compared with
Macintosh computer from DNASTAR, Inc. was used for theestima-shorter evolutionary history of other vertebrates (3).
tion of sequence homology based on percent similarity and divergenceamong different cDNA sequences ofb-,g- andgS-crystallins. Percent
That sharks were once considered to be the most
primi-divergence is calculated by comparing sequence pairs in relation totive are now thought to have been relatively specialized
the phylogenetic tree. On the other hand the percent similarity isregarding their complex biology to be ranked with birds
estimated by comparing sequences directly without accounting forand mammals as highly evolved creatures. In this
re-phylogenetic relationships. Phylogenetic tree was then constructedusing the algorithm of Hein (12) in the MegAlign programs of the
port the structural characterization of
g
S-crystallins
package. It is a multiple-sequence alignment program that buildsfrom shark lenses by PCR and its comparison with
trees as it aligns DNA or protein sequences using a combination ofthose of teleostean and mammalian lenses is of special
distance matrix and approximate parsimony methods. This methodimportant to unravel the complex evolutionary history
constructs multiple-alignment by imposing restrictions based onVol. 240, No. 1, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Multiple sequence alignment and comparison of seven crystallin sequences from species of different classes. The identical amino-acid residues among various sequences based on the first one (sharkgS-1) were expressed in white letters against black-background blocks. The gaps were introduced for optimal alignment and maximum homology for the sequences. Note that the middle region (residues# 70 – 130) shows a greater sequence variation than the N- and C-terminal regions among the compared sequences. Amino acid residues are denoted by one-letter symbols.
some conformational properties as revealed by circular
Characterization of cDNAs Encoding
g
S-Crystallins
dichroism (14,15). Shark
g
-crystallin showed a much
from Shark Lenses
more complex pattern in isoelectric focusing (data not
Previous studies have indicated the unusual struc-
shown), revealing that it consists of various charge
iso-tural characteristics of shark
g
-crystallin as compared
forms (13, 14). Due to the complexity of
g
-crystallin,
with those associated with the lenses of teleostean
we suspect that the same multiplicity of isoforms may
fishes such as carp (13 – 15). Especially noteworthy is
be also present for
g
S-crystallin, a lens protein with
the finding that the amino acid compositions of
g
-crys-
dual structural properties of
b
- and
g
-crystallins (7,8
tallins seem to lack the unique characteristic of high
and the references therein). We have hence resorted to
methionine content (ú 10%) as commonly observed for
the recent rapid method of cloning and sequencing by
that of teleostean fishes (5,6). In contrast it is closer to
means of PCR technique for the determination of cDNA
mammalian
g
-crystallins regarding both the amino
sequences of these multiple isoforms.
acid composition,
N-terminal partial sequence plusPCR amplification of total lens cDNA mixtures
pre-pared from two lenses of a single shark with the
de-signed primers based on partial DNA coding sequences
of carp
g
S-crystallin (10) achieved the isolation of one
PCR fragment corresponding to the complete reading
frame encoding at least two
g
S-crystallin isoforms from
this shark species. The size determination of
PCR-am-FIG. 3. Pair-wise comparison of amino-acid sequence homologyplified cDNA coding for
g
S crystallin was estimated to
between two sharkgS-crystallins and various b-, g- andgS-crys-be about 520 bp, in agreement with a protein of about
tallins from species of different classes. Analysis of sequence homol-170 – 180 amino-acid residues for mammalian
g
- and
ogy was carried out using the software package (DNASTAR Inc.,g
S-crystallins. The PCR-amplified DNA fragments
Madison, WI, USA) on the published sequences of carpgS (10),bo-were then subcloned into pUC18 previously digested
vine gS (7), bullfrogb2 (24), bovine b2 (25), bovine gII (26) andVol. 240, No. 1, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
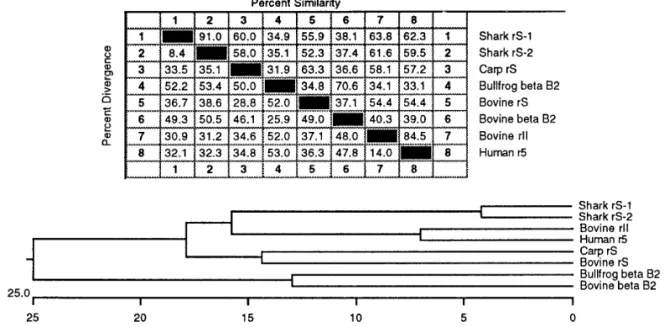
FIG. 4. Pair-wise comparison of protein sequence similarity and divergence (Top) and construction of phylogenetic tree (Bottom) of eight crystallin sequences from various species of three major classes of vertebrates. Analysis of sequence data was carried out in a software package of LaserGene for the Apple Macintosh computer (DNASTAR Inc., Madison, WI, U.S.A.). Percent divergence is calculated by comparing sequence pairs in relation to the relative positions in the phylogenetic tree. On the other hand the percent similarity is estimated by comparing percent sequence identity directly without accounting for phylogenetic relationships. A phylogenetic tree was then constructed based on the percent divergence between protein sequences using a combination of distance matrix and approximate parsimony methods in the phylogeny generation program of Hein (12). This algorithm carries out multiple-alignment by imposing restrictions based on evolutionary relatedness of the aligned sequence pairs. The tree was built using clustal method and weighted residue-weight table. The length of each pair of branches represents the sequence distance between aligned pairs. The scale beneath the tree measures the distance between sequences (in millions of years). Bovine and bullfrogbB2 crystallins are included to indicate the fact thatb- andg-crystallins form ab/g
superfamily.
strain JM 109. Plasmids purified from two positive
173 amino-acid residues including the initiating
methi-onine, which is close to carp
g
S (174 a.a.) and slightly
clones were then prepared for nucleotide sequencing.
lower than bovine
g
S (178 a.a.).
Sequence Analysis of cDNA Encoding Shark
Sequence Alignment and Comparison of
b
-,
g
- and
g
S-Crystallins
g
S-Crystallins from Different Species
More than five positive clones have been identified,
with their 5
* and 3* nucleotide sequences being deter-
Fig. 2 aligns seven sequences encompassing
repre-sentative
b
-,
g
- and
g
S-crystallins from characterized
mined to be essentially identical to those of the
de-signed primers, indicative of the existence of multiple
crystallins of the major classes in vertebrates, which
have all been deduced from nucleotide sequences
cod-isoforms for shark
g
S-crystallin, which is in contrast
to bovine (7,8) and human
g
S-crystallins with only one
ing for these crystallins. It is noteworthy that there is
only about 35 – 64% sequence homology between shark
sequence being identified (16). In order to avoid
se-quencing errors introduced in the manual sese-quencing
g
S crystallins and structurally related
b
-,
g
- and
g
S-crystallins from different evolutionary classes.
How-of isotope-labeled cDNA, sequence accuracy was doubly
checked and confirmed by automatic fluorescence-
ever one salient feature is that some of the key residues
(such as Tyr-6, Glu-7, Phe-11, Gly-13 and Ser-34 based
based sequencing technique. The only uncertainty may
lie in the first and last few nucleotides present in the
on bovine
g
II sequence numbering) for the
mainte-nance of stability in
g
-crystallins (17 – 19) are mostly
5
* and 3* region of the PCR fragment even though we
have used some degenerate codons in the primers. Fur-
retained and conserved in all
b
-,
g
- and
g
S-sequences
even from species of distantly related classes, attesting
ther genomic analysis of shark
g
S-crystallins my help
solve the ambiguity in these short segments. The de-
to the conservative structural aspects of
b
/
g
superfam-ily. It is also of interest to find that N- and C-terminal
duced protein sequences together with their genetic
coding sequences of two clones, designated as shark
regions of these crystallins are more conserved than the
middle regions of the sequences (residues# 70 – 130).
g
S-1 and
g
S-2 are shown in Fig. 1A and 1B. The cDNA
sequences encoding shark
g
S-1 and
g
S-2 were both
In the pair-wise sequence homology comparison (Fig.
3) of various deduced amino-acid sequences from
spe-found to consist of 519 nucleotides excluding the stop
Vol. 240, No. 1, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
package (DNASTAR program), it is found that shark
REFERENCES
g
S-crystallins show 62 – 64% sequence identity to bo-
1. Chiou, S.-H. (1986) FEBS Lett. 201, 69 – 73.vine
g
II crystallins whereas only 52 – 56% sequence
2. de Jong, W. W., and Hendriks, W. (1986) J. Mol. Evol. 24, 121 –identity is found between shark and bovine
g
S-crys-
129.3. Keeton, W. T. (1972) in Biological Science, 2nd ed., W. W.
Nor-tallins, indicating that shark
g
S is evolutionarily more
ton & Company, New York.
related to authentic mammalian
g
- than
g
S-crystallins
4. Chang, T., Jiang, Y.-J., Chiou, S.-H., and Chang, W.-C. (1988)
and may represent one intermediate crystallin form
Biochim. Biophys. Acta 951, 226 – 229.
from the divergent evolution of
g
-crystallin gene
fam-5. Pan, F.-M., Chang, W.-C., Chao, Y.-K., and Chiou, S.-H. (1994)
ily. On the other hand, shark
g
S-crystallins show only
Biochem. Biophys. Res. Commun. 202, 527 – 534.58-60% sequence identity to carp
g
S, underlying the
6. Pan, F.-M., Chang, W.-C., Lin, C.-H., Hsu, A.-L., and Chiou, S.-H. (1995) Biochem. Mol. Biol. International 35, 725 – 732.distinct differences of
g
S-crystallins present in the
car-7. Quax-Jeuken, Y., Driessen, H. P. C., Leunissen, J., Quax, W., de
tilaginous and teleostean fishes.
Jong, W., and Bloemendal, H. (1985) EMBO J. 4, 2597 – 2602. 8. van Rens, G. L. M., Raats, J. M. H., Driessen, H. P. C.,
Old-enburg, M., Wijnen, J. T., Khan, P. M., de Jong, W. W., and
Construction of a Phylogenetic Tree Based on Protein
Bloemendal, H. (1989) Gene 78, 225 – 233.
Sequence Comparison
9. Maniatis, T., Fritsch, E. F., and Sambrook, J. (1989) in MolecularCloning: A Laboratory Manual, Cold Spring Harbor Press, Cold
We have previously constructed a phylogenetic tree
Spring Harbor, N.Y.10. Chang, T., and Chang, W.-C. (1987) Biochim. Biophys. Acta 910,
of crystallins based on protein or DNA sequence
com-89 – 92.
parison with similar results (20 – 23). In Fig. 4 a
phylo-11. Sanger, F., Nicklen, S., and Coulson, A. R. (1977) Proc. Natl.
genetic tree is constructed based on multiple sequence
Acad. Sci. USA 74, 5463 – 5467.
alignment of eight protein sequences using a combina-
12. Hein, J. (1990) Methods Enzymol. 183, 626 – 645.tion of distance matrix and approximate parsimony
13. Siezen, R. J., Hom, C., Kaplan, E. D., Thomson, J. A., andmethods (12). It has been shown to be useful to align
Benedek, G. B. (1988) Exp. Eye Res. 46, 81 – 93.highly evolved gene families and their corresponding
14. Chiou, S.-H. (1989) FEBS Lett. 250, 25 – 29.15. Chiou, S.-H., Chen, S.-W., Itoh, T., Kaji, H., and Samejima, T.
protein sequences that have clear evolutionary
rela-(1990) FEBS Lett. 275, 111 – 113.
tionships as in the cases of various crystallin family.
16. Smith, J. B., Yang, Z., Lin, P., Zaidi, Z., Abbasi, A., and Russell,
It is noteworthy that the phylogenetic tree based on
P. (1995) Biochem. J. 307, 407 – 410.
the sequence divergence among these protein se-
17. Blundell, T., Lindley, P., Miller, L., Moss, D., Slingsby, C., Tickle,quences indeed exemplifies the close relatedness be-
I., Turnell, B., and Wistow, G. (1981) Nature 289, 771 – 777.tween shark
g
S-crystallins and
g
-crystallins from bo-
18. Wistow, G., Turnell, B., Summers, L., Slingsby, C., Moss, D.,Miller, L., Lindley, P., and Blundell, T. (1983) J. Mol. Biol. 170,
vine and human lenses. On the other hand, carp
g
S-175 – 202.
crystallin was grouped with bovine
g
S-crystallin, in
19. Liaw, Y.-C., Chiou, S.-H., Lin, C.-W., Chang, T., and Chang,
agreement with the percent homology shown in Fig. 3.
W.-C. (1992) J. Biochem. 112, 341 – 345.Especially interesting is the observation that two
b
2-
20. Chiou, S.-H., Yu, C.-W., Lin, C.-W., Pan, F.-M., Lu, S.-F., Lee,crystallin sequences from bullfrog and bovine indeed
H.-J., and Chang, G.-G. (1995) Biochem. J. 309, 793 – 800.21. Lu, S.-F., Pan, F.-M., and Chiou, S.-H. (1995) Biochem. Biophys.
locate at different branching points of the tree from
Res. Commun. 216, 881 – 891.
those of
g
- and
g
S-crystallins, indicative of two distinct
22. Pan, F.-M., Chang, W.-C., Lu, S.-F., Hsu, A.-L., and Chiou,
evolutionary pathways leading to
b
- and
g
/
g
S
crys-S.-H. (1995) Biochem. Biophys. Res. Commun. 217, 940 – 949.
tallins from the ancestral
b
/
g
protein.
23. Lu, S.-F., Pan, F.-M., and Chiou, S.-H. (1996) J. Protein Chem.15, 103 – 113.
24. Pan, F.-M., Chang, W.-C., Lu, S.-F., Hsu, A.-L., and Chiou, S.-H. (1995) Biochem. Biophys. Res. Commun. 217, 940 – 949.
ACKNOWLEDGMENTS
25. Hogg, D., Gorin, M. B., Heinzmann, C., Zollman, S., Mohandas, T. K., Klisak, I. J., Sparkes, R. S., Breitman, M. L., Tsui, L.-C., and Horwitz, J. (1987) Curr. Eye Res. 6, 1335 – 1341.
This work was supported by Academia Sinica and the National
26. Bhat, S. P., and Spector, A. (1984) DNA 3, 287 – 295. Science Council (NSC Grants 83-0203-B-001-086,
83-0418-B-001-020BA, 84-2311-B-001-050-BA & 86-2311-B-002-031-B15), Taipei, 27. Meakin, S. O., Du, R. P., Tsui, L.-C., and Breitman, M. L. (1987)
Mol. Cell. Biol. 7, 2671 – 2679.