行政院國家科學委員會專題研究計畫 成果報告
膽道閉鎖嬰兒及輪狀病毒感染誘發肝外膽管阻塞鼷鼠之基
因表現(3/3)
研究成果報告(完整版)
計 畫 類 別 : 個別型 計 畫 編 號 : NSC 95-2314-B-002-022- 執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 國立臺灣大學醫學院一般醫學科 計 畫 主 持 人 : 許宏遠 計畫參與人員: 碩士級-專任助理:江政倫 處 理 方 式 : 本計畫可公開查詢 中 華 民 國 96 年 10 月 29 日行政院國家科學委員會補助專題研究計畫
■ 成 果 報 告
□期中進度報告
膽道閉鎖嬰兒及輪狀病毒感染誘發肝外膽管阻塞鼷鼠之基因表現
Gene expression profiles in infants with biliary atresia and in mice with rotavirusinduced extrahepatic biliary obstruction
計畫類別:█ 個別型計畫 □ 整合型計畫 計畫編號:NSC 95-2314-B-002-022 執行期間: 2006 年 08 月 01 日至 2007 年 07 月 31 日 計畫主持人:許宏遠 共同主持人: 計畫參與人員: 江政倫 成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、列管計 畫及下列情形者外,得立即公開查詢 □涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位:台灣大學醫學院一般醫學科 中 華 民 國 96 年 10 月 30 日
膽道閉鎖嬰兒及輪狀病毒感染誘發肝外膽管阻塞鼷鼠之基因表現
Gene expression profiles in infants with biliary atresia and in mice with rotavirusinduced extrahepatic biliary obstruction
Hong-Yuan Hsu, M.D. Department of Primary Care Medicine, National Taiwan University College of Medicine, Taipei, Taiwan
Supported by grant 95-2314-B-002-022 from National Science Council of Taiwan
INTRODUCTION
Biliary atresia (BA) is a rare hepatobiliary disorder in early infancy characterized by a progressive, inflammatory process of extrahepatic as well as intrahepatic bile ducts leading to fibrosis and obliteration of the biliary tracts, resulting in biliary cirrhosis and finally requiring liver transplantation.1,2The incidence of BA is higher in Chinese than in Caucasians. The pathogenesis underlying this dynamic necro-inflammatory destructive cholangitis remains unknown. Several possible pathogenetic mechanisms for the development of BA have been proposed.2 Previous studies suggested that biliary epithelial injury of this disease possibly result from the interplay between genetic susceptibility and environmental exposure (viruses, toxic agents and metabolic insults) at perinatal period.1-4 Immunohistochemical studies demonstrated the infiltration of CD4+, CD8+ lymphocytes and natural killer (CD56+) cells predominated in the liver and extrahepatic bile ducts and an increase in CD68+ macrophage infiltration in portal tracts and biliary remnant tissue in BA patients with poor outcome after the portoenterostomy procedure.5-8 These findings suggested that inflammation involving several kinds of immune cells produce biliary epithelial injury such as pyknosis and necrosis of biliary epithelial cells and also promote hepatic fibrosis and cirrhosis in infants with BA. The mechanisms underlying recruitment of inflammatory cells, immunologic dysregulation, ductular apoptosis and progressive hepatic fibrosis in BA remains unclear. By using DNA microarrays in studying gene expression profile in infants with biliary atresia and neonatal hepatitis, we hope to delineate the complex gene regulation networks involved in the the genesis of BA. In addition, we compared gene expression in the liver in BA infants with postoperative good bile flow versus BA infants without good bile flow and tried to elucidate genes that are related to the pathogenesis of the specific clinical feature of BA.
METHODS
Gene expression study in infants with biliary atresia and neonatal hepatitis:
As shown in Table 1, liver tissue was taken by wedge biopsy from 9 infants (aged 1-3 months, patients A to I) undergoing Kasai operation for BA after diagnostic investigation for biliary atresia at The Department of Pediatrics, National Taiwan University Hospital. They were further divided into two groups according to the clinical features at the end of 12 months follow-up after Kaisai operation: group 1 patients (N=3, patients A, B and C) were jaundice free with good bile flow and group 2 patients (N=6, patients D to I, arbitrarily divided into 2a and 2b groups) had persistent and even aggravated jaundice and finally underwent liver transplantation. In addition, liver biopsy samples were obtained from group 3 infants (N=6, patients J to O, arbitrarily divided in to 3a and 3b groups) with advanced stages of biliary atresia at the time of transplantation (aged 7.5-14.6 months). Needle biopsy liver samples were also obtained from group 4 infants (N=5, age 1.5-2 months, patients P to T) with neonatal hepatitis but without identifiable specific metabolic or transport defects served as disease control. Liver samples from normal, age-matched infants were not obtained due to ethical consideration.
For DNA microarrays, total RNA was extracted using TRIzol Reagent (Invitrogen, Carlsbad, CA) according to themanufacturer’sprotocol.Total RNA was further purified using the RNeasy Mini Kit (Qiagen, Valencia, CA). The structural integrity of the isolated RNA was confirmed by electrophoresis of the RNA through a 1.2% gel. Purity of RNA was ascertained by inclusion of samples with an absorbance ratio 260/280 nm >1.8. 25 ug total RNA from each patient were pooled into five groups (group 1, 2a, 2b and 3a and 3b, respectively) according to the classification described above. Each pooled sample was biotin-labeled and hybridized to oligonucleotide based Genechip Human Genome U133 plus 2.0 Array containing 47400 gene products, representing 38500 well-characterized human genes. Specific hybridization and gene expression were monitored by image analysis of the chip with GCOS (GeneChip Operation System, Affymetrix). GeneSpring 7.0 (Silicon Genetics, Redwood, CA) was used to identify genes differentially expressed in liver of infants with biliary atresia and in liver of infants with neonatal hepatitis.
To verify the microarray results, we performed quantitative RT-PCR on selected genes using the individual samples contributing to RNA pools. We used High-Capacity cDNA Reverse Transcription kits (Applied Biosystems) to generate cDNA. PCR was carried out on a ABI PRISMR 7700 Sequence Detection System (Applied Biosystems) using the iTagTM SYBRR Green Supermix with ROX (BIO-RAD) as described in manufacturer’s instructions. Primer pairs for osteoopontin, osteonectin, LYZ and GAPDH
were chosen from Primer Assay list. Reactions were carried out under the following conditions: 95o C for 2 to 3 min, then 40 cycles of 95o C for 15 sec, 55o C for 30 sec. Melting curves analyses were performed to verify the amplification specificity. For quantification, gene expression of the target sequence was normalized in relation to the expressed housekeeping gene GADPH. For comparative purpose, the adjusted expression value of each target sequence in BA patients was further normalized in relation to an average of expression of the corresponding gene in all NH patients
Hepatobiliary pathology and gene expression study in RRV-infected mice.
The MMU18006 strain of rhesus rotavirus (RRV) was purchased from American Tissue Culture Collection (ATCC). Within the first 24 hours of life, the newborn Balb/c mice were inoculated through intraperitoneal route with a volume of 20 ul containing 2x106 PFU of RRV. The same volume of cell culture media was also administered to the subjects of the control group. Mice were examined daily for the development of icterus of non fur-covered skin and acholic stool, and sacrificed at 3, 7, 14 and 21 days after RRV or culture media injection. At the time of sacrifice, the extrahepatic bile ducts and gallbladder and liver were harvested. These specimens were used for histopathological studies (fixed in buffered 10% formalin solution, embedded in paraffin). Consecutive sections of specimens were subsequently stained with hematoxylene and eosin and subjected to RNA isolation. Groups of 2-3 bile ducts/gallbladders for each experimental and control group were obtained, used to generate biotinylated cRNA pools, and hybridized separately with the high-density, oligonucleotide-based Affymetrix MOE430 GeneCh, as described previously.
RESULTS
Gene expression profile in BA infants and neonatal hepatitis infants
Because only a small amount of RNA sample was available for NH patients and some BA patients, pooled RNA for each group patients was used for microarray analysis. Thus we were unable to use data filtering and cluster analysis to identify those transcripts with significant differences in mean signal intensity between BA patients and neonatal hepatitis. Instead, we identified those genes that were differentially expressed in livers of infants with biliary atresia compared with that of neonatal hepatitis by selecting those genes (with high relevance to immune response, inflammation, cell proliferation or cell-matrix communication) with hepatic mRNA expression in both group 1 and group 2 BA infants with at least 4 fold change above or below expression when compared with that in infants with neonatal hepatitis. The reproducibility of the signal intensity was confirmed for these genes by RT-PCR. The relevant genes included genes encoding
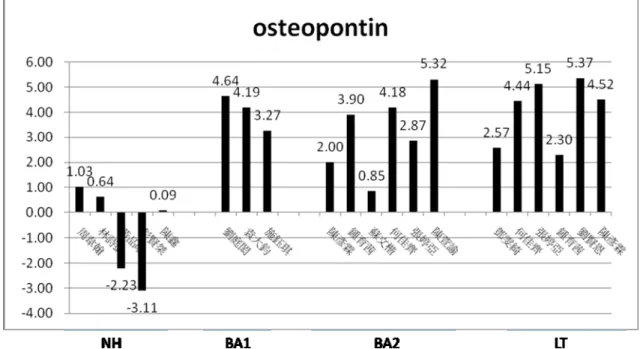
integrin alpha-2; integrin beta-3; T-cell receptor alpha locus; killer cell immunoglobulin-like receptor, two domains, long cytoplaslic tail; interleukin 23, alpha subunit P19; C-type lectin domain family 4 member C; Immunoglobulin heavy conctant delta; UL16 binding protein 2; leukotriene C4 synthase; interleukin 31 receptor A; secreted phosphoprotein 1 (osteopontin) and matrix metallopeptidase (MMP-7) (Table 2). On the other hand, several genes with known function displaying significantly lower signal intensity in group 2 BA infants than in diseased controls (neonatal hepatitis patients) are genes encoding immunoglobulin kappa constant; immunoglobulin light chain variable region complementarity determing region; interleukin 27 receptor, alpha; cytochrome P450, family 7, subfamily A, polypeptide 1 (Table 2). The reproducibility of the signal intensity was confirmed for osteopontin by RT-PCR (Figure 1-3).
Gene expression profile in BA infants with and without postoperative good bile flow We found several genes that differentially expressed in the liver of BA infants with and without postoperative bile flow. These genes with higher expression in the liver in BA without good bile flow encoded osteonectin/sparc, fibroblast growth factor binding protein 7, nitric oxide synthase 1; C-type lectin domain family 4, member C; interleukin 23, alpha subunit P19; arachidonate 15-lipoxygenase; CD80 molecule; integrin, alpha 2; Neurophilin 1; melanoma cell adhesion molecule; peroxisomal proliferation-activated receptor delta; sorbin and SH3 domain containing 1; jagg 1 (Allagille syndrome); multimerin 1; chemokine (C-C motif) ligand 14 or 15; MHC class II, DR beta 4. The genes with lower expression in BA infants without postoperative bile flow encoded genes encoding lysozyme (renal amyloidosis); hepatocellular carcinoma-associated gene (Table 3).
Histopathological findings of liver and extrahepatic bile ducts in RRV-infected mice.: An acute inflammatory process was noted in the liver and in the extrahepatic bile ducts (EHBD) in the first week after RRV infection. Histopathology of cross-sections of the extrahepatic bile duct after RRV challenge showed mild inflammatory cell infiltration at 3 days, and marked cholangitis and injury to duct epithelium at 7 days. In the first week after infection, focal necrosis in hepatic lobules and portal tracts and mononuclear cells and neutrophil infiltration in EHBDs were noted. Complete or interrupted occlusion of EHBD with mainly lymphocyte infiltration both in the liver and in the EHBDs was found 14-21 days after infection.
DISCUSSION
Among those genes exhibiting higher expression in either in group 1 and group 2 BA infants than in NH infants, the increase in the signal intensity for osteopontin was noted. Osteopontin is an extracellular matrix cell adhesion protein which is also a novel
substrate for matrix metalloproteinase 3 and 7 (MMP-3 and MMP-7). MMP-cleaved osteopontin has increased activities in cell migration, cell survival, and most notably, cytokine-like functions such as a central regulator for the T-helper 1 (Th-1) commitment of lymphocytes in vivo.9 We found an increased expression of genes encoding osteopontin and MMP-7 at early stage of biliary atresia compared with that of age-matched neonatal hepatitis infants, suggesting that a potential role of an early T helper 1 (Th-1) lymphocyte activation play a significant role in the pathogenesis of biliary atresia. Furthermore, up-regulation of osteopontin gene continues in group 3 BA patients, suggesting that activation of Th1 pathway is also important throughout the progression to biliary cirrhosis in biliary atresia. In addition, livers of our group 1 and group 2 BA infants showed increased expression of genes encoding UL16 binding protein 2, T-cell receptor alpha locus, interleukin 23 receptor and C-type lectin domain which involves antigen processing and presentation, natural killer cell activation and T cell mediated immune response.
Of interest is that genes encoding leukotriene C4 synthase (involved in Notch signaling pathway and leukotriene biosynthetic process) and ectodysplasin A (involved in activation of NF-kappa B transcription factor) are also upregulated in BA patients, suggesting that other unrevealed mechanisms may participate the progression of hepatobiliary pathology in biliary atresia.
We could also detect a lower expression of genes encoding immunoglobulin kappa constant and immunoglobulin light chain variable region complementarity determing region, indicating a suppression of Th2 lymphocyte response at the early phase of biliary atresia. The significance of the lower expression of several immunity/inflammation genes, such as a decreased infiltration of B lymphocyte or a suppressed B lymphocyte functions within the liver in group 2 BA patients needs further studies.
Our findings are in agreement with a recent report in infants with biliary atresia reported by Bezerra et al.10 They analyzed a large-scale gene expression in the livers of infants with BA or neonatal intrahepatic cholestasis and found a predominant and coordinated activation of immunity/inflammation genes within the livers of infants with BA. Most of the genes showed differential lymphocyte function, with activation of T helper lymphocytes and suppression of immunoglobulin genes in early stages of disease. Direct comparison in gene expression of liver with age-matched healthy infants was not possible due to ethical reasons in obtaining liver biopsy samples from healthy, age-matched infants. It has been shown that in normal infants, T-helper phenotype in circulating mononuclear cells can be skewed toward a Th-2 phenotype at birth, but this functional commitment may depend directly on a timely exposure to antigens.11,12 Our results, therefore, add further support to the view that a dominant pro-inflammatory
differentiation of lymphocytes in affected livers may have a key role in the pathogenesis of BA.
In group 2 BA infants, persistent or aggravated jaundice was noted after portoenterostomy and liver transplantation was performed later in life. It was reasonable to hypothesize that injury to hepatobiliary system was more serious than in group 1 BA infants at the time of liver samples obtained. Among those genes overexpressed in group 2 when compared with group 1 BA infants, osteonectin/sparc was found to be related to the pathogenesis of hepatic fibrosis and sparc mRNA is weakly expressed in normal liver but upregulated in fbrotic liver and in activated stellate cells.13,14 Chemokine (C-C motif) ligand 14 can induce changes in intracellular calcium concentration and enzyme release in monocyte, chemokine (C-C motif) ligand 15 is chemotactic for T cells and monocytes and induces N-acetyl-beta-D-glucosaminidase release in monocytes. CD80 molecule involves T cell activation and positive regulation of T helper-1 cell differentiation. Ribosomal protein S6 kinase is involved in proteolysis and epidermal growth factor receptor signaling pathway. Neuroplilin 1 is involved in positive regulation of cell proliferation, angiogenesis and multicellular organismal development. Peroxisome proliferators-activated receptor delta is involved in cell proliferation and apoptosis. Fibroblast growth factor binding protein regulates progression through cell cycle, cell proliferation, epidermis development and response to wounding. Jagged 1 (Alagille syndrome) regulates cell proliferation, differentiation and migration, is involved in Notch signaling pathway and is expressed in ductal plate epithelia.15 Missense mutations of jagged 1 gene was recently identified in some biliary atresia patients with worse prognosis but without phenotypic features of Alagille syndrome.15
These genes up-regulated in group 2 BA patients may represent an adaptive change with more serious proliferation of intrahepatic bile duct cells and a prominent fibrogenesis in response to serious injury to hepatobiliary system in early stages of biliary atresia resulting in a rapid progression to biliary cirrhosis in these affected infants. However, it is possible that activation of specific cells in the seriously injuried liver directly induce a rapid progression of the disease. Based on functional classification, those genes overexpressed in group 2 BA infants involved more prominent immune response regarding T cell immune response, inflammation, cell proliferation, regulation of apoptosis, and fibrogenesis. The genes underexpressed in group 2 BA infants encode lysozyme (involved in defense response to bacteria); Kilirin, RhoGEF kinase (involved in regulation of Rho protein signal transduction); PRKC, apoptosis, WT1, regulator (involved in apoptosis and negative regulation of T cell proliferation) and intraflagellar transport 57 homolog (involved in regulation of apoptosis and caspase activation).
involved in lymphocyte differentiation leading to activation of T-helper 1 cell-mediated immunity and suppression of immunoglobulin genes in early stages of biliary atresia. A perpetuation of such T cell immune response perpetuated in concert with enhanced expression of genes regarding inflammation, cell proliferation, apoptosis and fibrogenesis at the time of diagnosis of biliary atresia points to specific clinical feature of BA, such as BA without good bile flow after portoenterostomy. Whether the expression level of these genes can serve as a marker that reflects the severity of liver histopathology of BA and predict a rapid worsening of disease course requires further study in the future.
Table 1. Characteristics of infants with biliary atresia(BA) or neonatal hepatitis(NH) Sex (m/f) Age (days) Diagnosis portoenterostomy at the time of liver sample obtained Group I A m 47 BA No B f 47 BA No C f 83 BA No Group IIa No D f 34 BA No E m 40 BA No F m 74 BA No Group IIb No G f 41 BA No H f 55 BA No I m 69 BA No Group IIIa J m 227 BA Yes K f 401 BA Yes L f 439 BA Yes Group IIIa M m 251 BA Yes N f 272 BA Yes O m 394 BA Yes Group IV P m 43 NH No Q f 46 NH No R m 48 NH No S m 60 NH No T m 61 NH No
Group I:BA infants with jaundice free and good bile flow after portoenterostomy.
Group II:BA infants with persistent jaundice after portoenterostomy and finally underwent liver transplantation.
Group III:BA infants with advanced stages of BA at the time of transplantation. Group IV:neonatal hepatitis infants without identifiable specific causes.
Table 2. Representive list of up-regulated or down-regulated gene (4 fold change above or below) in infants with biliary atresia in comparison with level in infants with neonatal hepatitis.
Probe set ID Gene Symbol Gene Title Fold change Function
Gr. I Gr. IIa Gr. IIb upregulated
227314_at ITGA2 integrin, alpha 2 26 30 24 cell-matrix adhesion
211579_at ITGB3 integrin, beta 3 32 14 8 cell-matrix adhesion
217394_at TRA@ T cell receptor alpha locus 62 16 31 immune response
208179_x_at KIR2DL3 killer cell Ig-like receptor, two domains, long cytoplasmic tail, 3
18 5 10 immune response
234865_at IL23A Interleukin 23, alpha subunit p19 33 15 8 Immunity/inflammation 1555687_a_at CLEC4C C-type lectin domain family 4,
member C
21 21 7 immune response
215621_s_at IGHD Ig heavy constant delta 22 8 19 immune response
238542_at ULBP2 UL16 binding protein 2 14 9 6 immune response
206480_at LTC4S leukotriene C4 synthase 6 15 19 Notch signaling pathway
leukotriene biosynthetic process
1555431_a_at IL31RA interleukin 31 receptor A 3 12 11 Immune response
209875_s_at SPP1 secreted phosphoprotein 1
(osteopontin)
7 5 4 Immune response
(early T cell activation)
218995_s_at EDN1 endothelin 1 5 7 8 cell proliferation
211131_s_at EDA ectodysplasin A 20 10 8 Immune response
(activation of NF-kappaB transcription factor) downregulated
211644_x_at IGKC Ig kappa constant 6 16 7 Immune response
214777_at --- Ig light chain variable region complementarity determining region (CDR3)
5 13 4
immune response
217702_at IL27RA interleukin 27 receptor, alpha 2 9 9 Immune response
207406_at CYP7A1 cytochrome P450, family 7, subfamily A, polypeptide 1
192 24 6 bile acid biosynthetic ;
lipid, steroid metabolic process
Group I:jaundice free with good bile flow.
Group II:persistent jaundice after portoenterostomy and finally underwent liver transplantation. Ig:immunoglobulin
Table 3. Representive list of up-regulated or down-regulated genes in biliary atresia infants with persistent jaundice after portoentrostony, in comparison with level in biliary atresia infants with good bile flow after portoentrostony
Probe set ID Gene Symbol Gene Title Fold change Function
Gr. IIa Gr. IIb upregulated
235342_at SPOCK3 sparc/osteonectin 57 15 fibrosis
205612_at MMRN1 multimerin 1 8 3 cell-matrix interaction
205392_s_at CCL14 CCL15
chemokine (C-C motif) ligand 14 chemokine (C-C motif) ligand 15
2 3 immune response
209728_at HLA-DRB4 major histocompatibility complex, class II, DR beta 4
1.14 2.41 immune response 239132_at NOS1 Nitric oxide synthase 1 (neuronal) 17 12 nitric oxide biosynthetic
process
cell-cell signaling 1552772_at CLEC4D C-type lectin domain family 4,
member D
4 4 immune response
217328_at IL23A Interleukin 23, alpha subunit p19 3 6 immune
response/inflammation 238453_at FGFBP3 fibroblast growth factor binding
protein 3
7 7
207328_at ALOX15 arachidonate 15-lipoxygenase 5 11 inflammation/
leukotriene biosynthetic
1555689_at CD80 CD80 molecule 9 9 immune response
1554819_a_at ITGA11 integrin, alpha 11 4 9 cell-matrix adhesion
integrin-mediated signaling pathway
1561365_at NRP1 Neuropilin 1 6 11 positive regulation of cell
proliferation
multicellular organismal development
209086_x_at MCAN melanoma cell adhesion molecule 5 9 cell adhesion anatomical structure morphogenesis 210636_at PPARD peroxisome proliferator-activated
receptor delta
5 8 cell proliferation
211705_s_at SORBS1 sorbin and SH3 domain containing 1
5 6 cell-matrix adhesion
209097_s_at JAG1 jagged 1 (Alagille syndrome) 4 4 regulation of cell proliferation multicellular organismal development
1554319_at RPS6KA5 ribosomal protein S6 kinase, 90kDa, polypeptide 5
8 4 epidermal growth factor
receptor signal pathway downregulated
1555745_a_at LYZ lysozyme (renal amyloidosis) 8 6 immune response
220437_at LOC55908 hepatocellular
carcinoma-associated gene TD26
4 3 unknown
222519_s_at IFT57 intraflagellar transport 57 homolog (Chlamydomonas)
14 6 apoptosis
caspase activation
214090_at PAWR PRKC, apoptosis, WT1, regulator 3 3 apoptosis
negative regulation of T cell proliferation
Both group IIa and group IIb are biliary atresia infants with persistent jaundice after portoentrostony and finally
Fig 1 RT-PCR shows increased expression for osteopontin in liver of infants with biliary atresia ( BA1,BA2 and BA3 ) when compared with that of infants with neonatal hepatitis(NH). The value (Q) was obtained from two steps of calculation including step1: Q=Target gene/GAPDH and then step2 Q sample/average of QNH.
REFERENCES
1. Balistreri, WF, et al. Biliary atresia: current concepts and research directions. Summary of a symposium. Hepatology 1996;23:1682-1692
2. Sokol, RJ, Mack C, Narkewicz, MR, Karrer, FM. Pathogenesis and outcome of biliary atresia: current concepts. J Pediatr Gastroenterol Nutr 2003;37:4-21
3. Yoon PW, Bresee JS, Olney RS, James LM, Khoury MJ. Epidemiology of biliary atresia: a population-based study. Pediatrics 1997;99:376-82
4. Riepenhoff-Talty M, Gouvea V, Evans MJ, Svensson L, Hoffenberg E, Sokol RJ, Uhnoo I, et al. Detection of group C rotavirus in infants with extrahepatic biliary atresia. J Infect Dis 1996:174:8-15
5. Ohya T, Fujimoto T, Shimomura H, Miyano T. Degeneration of intrahepatic bile duct with lymphocyte infiltration into biliary epithelial cells in biliary atresia. J Peditr Surg 1995;30:515-518
6. Ahmed AF, Ohtani H, Nio M, Funaki N, Shimaoka S, Nagura H, Ohi R. CD8+ T cells infiltrating into bile ducts in biliary atresia do not appear function as cytotoxic T cells: a clinicopathological analysis. J Pathol 2001;193:383-389
7. Davenport M, Gonde C, Redkar R, Koukoulis G, Tredger M, Mieli-Vergani G, Portmann B, Howard ER. Immunohistochemistry of the liver and biliary tree in extrahepatic biliary atresia. J Pediatr Surg 2001;36:1017-1025
8. Mack CL, Tucker RM, Sokol RJ, Karrer FM, Kotzin BL, Whitington PF, Miller SD. Biliary atresia is associated with CD4+ Th1 cell-mediated portal tract inflammation. Pediatr Res 2004;56:79-87
9. Chabas D, Baranzini SE, Mitchell D, et al. The influence of the proinflammatory cytokine, osteopontin, on autoimmune demyelinating disease. Science 2001;294:1731-35
10.Bezerra JA, Tiao G, Ryckman FC, et al. Genetic induction of proinflammatory immunity in children with biliary atresia. Lancet 2002;360:1653-9
11.Prescott SL, Macaubes C, Yabuhara A, et al. Developing patterns of T cell memory to environmental allergens in the first two years of life. Int Arch Allergy Immunolol 1997;113:75-79
12.Marchant A, Aaby P, Newborns develop a Th1-type immune response to Mycobacterium bovis bacillus Calmette-Guerin vaccination. J Immunol 1999;163:2249-55
13.Frizell E, Liu SL, Abraham A, et al. Expression of SPARC in normal and fbrotic livers. Hepatology 1995;21(3):847-54
14.Blazejewski S, Le Bail B, Boussarie L, et al. Osteonectin (SPARC) expression in human liver and in cultured human liver myofbroblasts. Am J Pathol 1997;151(3):651-7 Riepenhoff-Talty M, Schaekel K, Clark HF, et al. Group A
rotaviruses produce extrahepatic biliary obstruction in orally inoculated newborn mice. Peditr Res 1993;33:394-99
15.Kohsaka T, Yuan ZR, Guo SX, et al. The significance of human jagged 1 mutations detected in severe cases of extrahepatic biliary atresia. Hepatology 2002;36:904-12


![TraditionalMLCalgorithmsmainlytacklethebatchMLCproblem,wheretheinputdataarepresentedinabatch[24,28].Nevertheless,inmanyMLCapplicationssuchase-mailcategorization[22],multi-labelexamplesarriveasastream.Onlineanalysisistherefore dimensionreducermotivatedbyma](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)