利用農藝性狀評估糯稻種原之遺傳歧異性
陳源俊
1、李長沛
2、黃惠娟
3、簡禎佑
1、林順福
1*
1國立臺灣大學農藝學系
2行政院農業委員會農業試驗所農藝組
3行政院農業委員會動植物防疫檢疫局
摘要
世界糯稻之研究甚少,且臺灣糯稻之育
種歷史或規模均遠不及
稻或秈稻,尤其在
糯稻種原之評估及利用方面尚待加強。為探
討糯稻種原之遺傳歧異性,本研究以蒐集自
10 個國家的 135 個糯稻(包括 64 個秈糯、
53 個 糯及 28 個陸糯)品種與 5 個非糯稻
(對照品種)為材料,於
2000 年一期及二
期作在臺中地區調查
27 個質的性狀及 7 個
數量性狀。資料分析結果發現,7 個數量性
狀在兩期作間之差異皆達極顯著水準,且部
分 性 狀 間 在 兩 期 作 之 相 關 顯 著 性 檢 定 結 果
不一致。以農藝性狀進行歧異性分群分析結
果,在兩期作均無明顯大分群出現,亦無法
區分
糯、秈糯及陸糯稻種原。但是來自相
同國家的種原則有集中於小群之趨勢。農藝
性狀之主成分分析結果顯示,所調查之數量
性 狀 較 質 的 性 狀 具 有 較 佳 之 種 原 區 分 效
果,其中株高與葉長兩性狀在第一期作共可
解 釋
87.5% 變 異 , 在 第 二 期 作 共 可 解 釋
90.7%變異,依此兩性狀可將 糯及秈糯種
原分群,而秈糯稻較
糯稻種原有較大之遺
傳歧異性,但是陸糯稻則分散於秈糯及
糯
稻種原群之中。
關鍵詞︰遺傳歧異性、糯稻、種原。
Genetic Diversity Evaluation for
Glutinous Rice Germplasm Based
on Agronomic Traits
Yuan-Jan Chen
1, Charng-Pei Li
2,
Huei-Jiuan Huang
3, Jen-You Jian
1and Shun-Fu Lin
1*
1
Department of Agronomy, National Taiwan
University, Taipei 106, Taiwan (ROC)
2
Agronomy Division, Taiwan Agricultural
Research Institue, Taichung Hsien 413, Taiwan
(ROC)
3
Bureau of Animal and Plant Health Inspection
and Quarantine, Council of Agriculture, Taipei
100, Taiwan (ROC)
ABSTRACT
There have been very few researches on
glutinous rices in all over the world. In contrast
to indica or japonica type, the breeding history
and activities of glutinous rices are far behind in
Taiwan. Studies specifically focus on the
evaluation and utilization of germplasm for
glutinous rices are necessary. To evaluate the
genetic diversity of glutinous rice germplasm, 135
glutinous rices (including 64 indica, 53 japonica
and 28 upland types) collected from 10 countries
and 5 non-glutinous varieties were used in this
study. Twenty-seven qualitative and 7
quantitative traits were investigated at Taichung,
Taiwan during the first and second cropping
* 通信作者, [email protected]投 稿 日 期:2003 年 10 月 1 日 接 受 日 期:2004 年 9 月 6 日
作 物 、 環境 與生 物 資 訊 2:11-30 (2005)
Crop, Environment & Bioinformatics 2:11-30 (2005) 189 Chung-Cheng Rd., Wufeng, Taichung Hsien 41301, Taiwan (ROC)
seasons of 2000. There were significant differences
between two cropping seasons in the investigated
quantitative traits. However, in corresponding
comparison with the significant test results of
correlation between traits in different years, some
inconsistent results were found. No distinct group
was identified from clustering analysis based on
agronomic traits, and 3 types of glutinous
germplasm were unable to be differentiated. The
germplasm originated from different countries
were separately gathered in a few small groups.
The quantitative traits had better categorization
effectiveness than qualitative traits in principal
component analysis (PCA). Different subspecies
of glutinous germplasm were separated by using
culm length and leaf length as decisive factors,
which accounted for 87.5% and 90.7% variation in
the first and second cropping seasons,
respectively. Compared to japonica type
germplasm, the indica type had wider genetic
diversity. And the upland glutinous germplasms
were distributed diversely in japonica or indica
group.
Key words: Genetic diversity, Glutinous rice,
Germplasm.
前言
稻原為熱帶原始作物,後經由長期的栽
培及人為的選拔,逐漸演變能適應各種不同
氣候品種,所以現今稻不僅栽培於熱帶,溫
帶 北 部 亦 有 適 應 的 品 種 。 目 前 栽 培 稻 包 括
Oryza sativa 及 Oryza glaberrima 兩個物
種,亞洲地區栽培種屬於
Oryza sativa,可
分 為 秈 稻
(indica)、 稻(japonica) 及爪哇
稻
(Javanica)三類群(Chang 1976)。稻米的
胚 乳 中 澱 粉 由 支 鏈 澱 粉
(amylopectin)及直
鏈澱粉
(amylose)組成,而非糯性的米煮熟
後 之 米 飯 較 糯 性 的 米 飯 乾 鬆 。 糯 稻
(glutinous rice; waxy rice)依穀粒形狀區
分,包括圓糯
( 糯)及長糯(秈糯),其胚乳
中澱粉大多由支鏈澱粉組成,其直鏈澱粉含
量一般在
1~12%(COA et al. 1987)。根據稻
作 改 良 年 報
(2000)以 及 臺 灣 地 區 糧 食 生 產
情形的資料顯示,
2000 年臺灣地區的水稻
栽培面積約
32.9 萬公頃,以一般用途來說,
米 為 一 般 食 米 , 秈 米 以 製 作 蘿 蔔 糕 、 米
粉、炒飯為主。糯米中, 糯米以釀酒、米
糕為用,秈糯米以八寶粥、粽子為主要用途
(COA 1996)。
就種原利用而言,育種家所關心的是如
何有效率地篩選出所需的育種材料;而對於
種原管理者而言,所關心的則是如何利用最
經濟的資源來保存這些種原,並提升種原收
集和保存的效率
(Chen et al. 1997)。種原的
保存即在維持變異,所以維持物種的多樣性
將可提供育種家對於育種材料上選擇。研究
種原變異的方法很多,其中以外表型態特性
的觀察為最方便之方法,最早被應用於種原
變異的調查。
Holcomb et al. (1977)由種原
庫中逢機選取
1,407 個 稻及 488 個秈稻種
原,並且調查
14 個數量性狀及 27 個質量性
狀,然後進行多變值分析,發現秈稻種原較
稻 種 原 具 有 較 大 之 遺 傳 歧 異 。
Mackill
and Lei (1997)調查 117 個栽培稻之生育
期、幼穗活力、分蘗數及落葉性等性狀,進
行 遺 傳 歧 異 性 分 析 , 結 果 呈 現 連 續 性 之 分
布,其中熱帶之
型稻品種分布介於溫帶之
型稻品種與秈型稻品種之間,而不同群間
則不易細分。
近年來由於
DNA 分子標誌廣泛地應用
於各種生物遺傳研究及育種,也應用在水稻
之 演 化 與 遺 傳 資 源 評 估
(Second 1991) 。
Zhang et al. (1992) 使 用 95 個 RFLP
(restriction fragment length polymor-
phism)分子標誌分析 12 個 indica 及 14 個
japonica 品種,發現 indica 型內之品種變
異 大 於
japonica 型 內 , 此 一 結 果 與
Holcomb et al. (1977)分析主要形態及生理
性狀之結果相符合。另外以聚合酵素連鎖反
應
(polymerase chain reaction, PCR)為基
礎之
DNA 分子標誌,發展初期以 RAPD
(random amplified polymorphic DNA)應
用最廣,
Mackill (1995)利用 21 個 RAPD 引
子可區分熱帶型
japonica (14 個品種)與溫
帶型
japonica (24 個品種)兩群之品種,但
仍無法區分同一分群內之品種。但是另一學
者則以
18 個 RAPD 引子所產生之 45 個分
子標誌區分
2 個 japonica 型及 10 個 indica
型品種,且結果與同功酵素分析一致
(Virk
et al. 1995) 。另外 Zhu et al. (1998)使用 179
個
AFLP (amplified fragment length poly-
morphism)分子標誌分析 57 個水稻種原,
分群結果與先前之同功酵素分析結果一致。
Davierwala et al. (2000) 利 用 gene
bank 裡的資料設計出一組引子,分析 6 個
水稻栽培種及
5 個水稻野生種,其所得到的
多 型 性 片 段 長 度 之 差 異 在 於 重 複 序 列 次 數
之 不 同 。
Yang et al. (1994)則利用 10 組
Intra-SSR 引子所得到的 93 個對偶基因分
析
238 個水稻種原之遺傳歧異性,發現秈稻
型種原較
稻型種原具有較高
(大於 14%)之
多型性,且在地方種較栽培種更為明顯,兩
不同亞種間之變異約佔總變異之
10%。另一
群學者使用
32 個 Inter-SSR 引子分析 59 個
具有代表性之水稻種原,亦可將秈稻型及
稻型兩群種原區分
(Blair et al. 1999)。
二 十 世 紀 以 來 國 際 間 之 引 種 利 用 頻
繁,不同種或亞種間相互雜交利用,使得稻
之遺傳變異更為複雜,因而增加種原收集、
評估及實際育種利用之困難。為了有效地管
理與利用種原,有必要進行稻種原之遺傳歧
異性分析。經由遺傳歧異分析除了可以建立
種原鑑定之資料,亦可避免種原重複收集,
篩檢混雜之樣品,防止種原流失,建立核心
種原庫,及提供優良品系或具有經濟價值特
性種原供育種利用。
世界糯稻之研究甚少,且臺灣糯稻之育
種歷史或規模亦遠不及
稻或秈稻,糯稻又
可區分為
糯、秈糯及陸糯稻,不同種類間
之農藝性狀及澱粉組成之差異,與遺傳歧異
性均未有系統性之研究。本研究將藉由糯稻
種原之農藝性狀變異分析,再利用外表性狀
進 行 糯 稻 種 原 之 群 聚 分 析
(cluster
analysis) 、 及 主 成 分 分 析 (principal
component analysis),來探討臺灣及世界
糯稻種原的遺傳歧異性,希望所得訊息將可
提供糯稻種原蒐集管理、遺傳研究與育種利
用之參考。
材料與方法
一、試驗材料
本 試 驗 所 用 的 材 料 係 由 行 政 院 農 業 委
會 農 業 試 驗 所 農 藝 組 稻 作 研 究 室 所 提 供 之
水稻種原,包含臺灣地區
糯、秈糯、陸糯;
大陸地區
糯、秈糯、陸糯;日本地區
糯、
陸糯;韓國、泰國、印度、印尼、菲律賓、
美國、寮國、義大利等地區糯稻各有
2-10
個品種
(系)。另外以臺農 67 號、臺中秈 10
號、臺中在來
1 號、臺中 65 號及臺 9 號
等
5 個非糯性品種為對照。合計有 10 個不
同來源國家共
140 份種原。其種原代碼、品
種名稱、來源地區等列於
Table 1,在本文
所提到的參試材料均以
Table 1 所列之名稱
代碼表示之。
二、田間試驗
糯稻種原外表性狀之調查,係於
2000
年 第 一 期 作 及 第 二 期 作 在 行 政 院 農 業 委 員
會農業試驗所之試驗田進行,調查性狀係依
據
1996 年國際稻米研究所(IRRI)所發表之
評估標準進行,調查性狀計有葉毛、葉色、
莖基色、葉角度、劍葉型、葉舌色、葉舌形
狀、葉環色、葉耳色、稈角度、節間色、稈
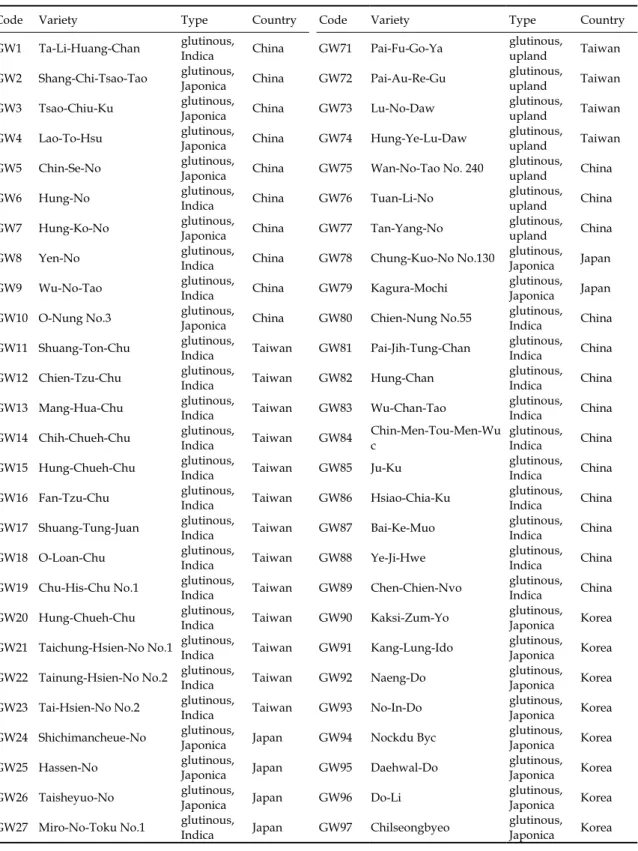
Table 1. Codes, names, types and origins of 135 glutinous and 5 non-glutinous rice germplasm.
Code Variety Type Country Code Variety Type Country
GW1 Ta-Li-Huang-Chan glutinous, Indica China GW71 Pai-Fu-Go-Ya glutinous, upland Taiwan GW2 Shang-Chi-Tsao-Tao glutinous,
Japonica China GW72 Pai-Au-Re-Gu
glutinous,
upland Taiwan GW3 Tsao-Chiu-Ku glutinous,
Japonica China GW73 Lu-No-Daw
glutinous,
upland Taiwan GW4 Lao-To-Hsu glutinous, Japonica China GW74 Hung-Ye-Lu-Daw glutinous, upland Taiwan GW5 Chin-Se-No glutinous,
Japonica China GW75 Wan-No-Tao No. 240
glutinous, upland China
GW6 Hung-No glutinous,
Indica China GW76 Tuan-Li-No
glutinous, upland China GW7 Hung-Ko-No glutinous, Japonica China GW77 Tan-Yang-No glutinous, upland China
GW8 Yen-No glutinous,
Indica China GW78 Chung-Kuo-No No.130
glutinous, Japonica Japan GW9 Wu-No-Tao glutinous,
Indica China GW79 Kagura-Mochi
glutinous, Japonica Japan GW10 O-Nung No.3 glutinous, Japonica China GW80 Chien-Nung No.55 glutinous, Indica China GW11 Shuang-Ton-Chu glutinous,
Indica Taiwan GW81 Pai-Jih-Tung-Chan
glutinous, Indica China GW12 Chien-Tzu-Chu glutinous,
Indica Taiwan GW82 Hung-Chan
glutinous, Indica China GW13 Mang-Hua-Chu glutinous, Indica Taiwan GW83 Wu-Chan-Tao glutinous, Indica China GW14 Chih-Chueh-Chu glutinous, Indica Taiwan GW84 Chin-Men-Tou-Men-Wu c glutinous, Indica China GW15 Hung-Chueh-Chu glutinous,
Indica Taiwan GW85 Ju-Ku
glutinous, Indica China GW16 Fan-Tzu-Chu glutinous, Indica Taiwan GW86 Hsiao-Chia-Ku glutinous, Indica China GW17 Shuang-Tung-Juan glutinous,
Indica Taiwan GW87 Bai-Ke-Muo
glutinous, Indica China GW18 O-Loan-Chu glutinous,
Indica Taiwan GW88 Ye-Ji-Hwe
glutinous, Indica China GW19 Chu-His-Chu No.1 glutinous, Indica Taiwan GW89 Chen-Chien-Nvo glutinous, Indica China GW20 Hung-Chueh-Chu glutinous,
Indica Taiwan GW90 Kaksi-Zum-Yo
glutinous, Japonica Korea GW21 Taichung-Hsien-No No.1 glutinous,
Indica Taiwan GW91 Kang-Lung-Ido
glutinous, Japonica Korea GW22 Tainung-Hsien-No No.2 glutinous, Indica Taiwan GW92 Naeng-Do glutinous, Japonica Korea GW23 Tai-Hsien-No No.2 glutinous,
Indica Taiwan GW93 No-In-Do
glutinous, Japonica Korea GW24 Shichimancheue-No glutinous,
Japonica Japan GW94 Nockdu Byc
glutinous, Japonica Korea GW25 Hassen-No glutinous, Japonica Japan GW95 Daehwal-Do glutinous, Japonica Korea GW26 Taisheyuo-No glutinous,
Japonica Japan GW96 Do-Li
glutinous, Japonica Korea GW27 Miro-No-Toku No.1 glutinous,
Indica Japan GW97 Chilseongbyeo
glutinous, Japonica Korea
(continued)
Code Variety Type Country Code Variety Type Country
GW28 Kintoki-No glutinous,
Japonica Japan GW98 Suweon 290
glutinous, Indica Korea GW29 Miyuoshin-No glutinous,
Japonica Japan GW99 Rinoldo-Bersoni
glutinous, Indica Italy GW30 Kilien-No glutinous,
Japonica Japan GW100 Kinan-Kuda
glutinous,
Japonica Philippine GW31 Shiasher-No glutinous, Japonica Japan GW101 Lin-Leng glutinous, Indica Philippine GW32 Kuroo-No No.22 glutinous,
Japonica Japan GW102 Legage-In
glutinous, Indica Philippine GW33 Aehegomiz-No glutinous, Japonica Japan GW103 IR 29 glutinous, Indica Philippine GW34 Tosou-No glutinous,
upland Japan GW104 Perurufong Nba
glutinous,
Indica Philippine GW35 Kirosheta-No glutinous, upland Japan GW105 Inaway glutinous, Indica Philippine GW36 Shiroga-No glutinous,
upland Japan GW106 IR3941-9-2
glutinous,
Indica Philippine GW37 Mirozaki-No No.1 glutinous,
upland Japan GW107 TD 52
glutinous,
Indica Thailand GW38 Fujiura-No No.16 glutinous,
upland Japan GW108 KU 81
glutinous,
Indica Thailand GW39 Tokyofhujra-No glutinous, upland Japan GW109 Hawm glutinous, Indica Thailand GW40 Yuronoyuki-No glutinous, upland Japan GW110 Ja-Ma-Bau-Jah glutinous, Indica Thailand GW41 Roku-U-No No.22 glutinous,
upland Japan GW111 Ja-No-Nug
glutinous, Indica Thailand GW42 Kunimiz-No glutinous, upland Japan GW112 K-17044 glutinous, Indica Thailand GW43 Gaisen-No glutinous, upland Japan GW113 K-17054 glutinous, Indica Thailand GW44 Tainung-No No.8 glutinous,
Japonica Taiwan GW114 K-17049
glutinous,
Indica Thailand GW45 Nung-Yu-No No.3 glutinous,
Japonica Taiwan GW115 K-17057
glutinous,
Indica Thailand GW46 Nung-Yu-No No.11 glutinous,
Japonica Taiwan GW116 India 16
glutinous, Indica India GW47 Nung-Yu-No No.13 glutinous, Japonica Taiwan GW117 India 18 glutinous, Indica India GW48 Nung-Yu-No No.20 glutinous,
Japonica Taiwan GW118 ARC 12886
glutinous, Indica India GW49 Nung-Yu-No No.24 glutinous,
Japonica Taiwan GW119 Jhum-Paddy 7
glutinous, Indica India GW50 Nung-Yu-No No.25 glutinous,
Japonica Taiwan GW120 WRC 4
glutinous, Indica India GW51 Nung-Yu-No No.30 glutinous, Japonica Taiwan GW121 KU 16 glutinous, Indica India GW52 Nung-Yu-No No.35 glutinous, Japonica Taiwan GW122 Land-Bauw glutinous, Indica Indonesia GW53 Nung-Yu-No No.54 glutinous,
Japonica Taiwan GW123 Umbang-Sampahiring
glutinous,
Japonica Indonesia GW54 Taichung-No No.46 glutinous,
Japonica Taiwan GW124 Glutinous-Lebonnent
glutinous,
Indica U.S.A. GW55 Hsinchu-No No.4 glutinous, Japonica Taiwan GW125 Khao-Kieng glutinous, Indica Loas
(continued)
Code Variety Type Country Code Variety Type Country
GW56 Taichung-No No.70 glutinous, Japonica Taiwan GW126 Kap-Nhay glutinous, Indica Loas GW57 Taikeng-No No.1 glutinous,
Japonica Taiwan GW127 Mack-Hing-Hoom
glutinous, Indica Loas GW58 Taikeng-No No.3 glutinous,
Japonica Taiwan GW128 Khao-Kam
glutinous, Indica Loas GW59 Taikeng-No No.5 glutinous,
Japonica Taiwan GW129 Khao-Chao-Hom
glutinous, Indica Loas GW60 Paerizu-Mochi glutinous, upland Taiwan GW130 Khao-Khane glutinous, Indica Loas GW61 Tarunatsu-Mochi glutinous,
upland Taiwan GW131 Khao-Lay-Nhay
glutinous, Indica Loas GW62 Warisan-Mochi No.1 glutinous,
upland Taiwan GW132 Khao-Nane-Noi
glutinous, Indica Loas GW63 Warisan-Mochi No.2 glutinous,
upland Taiwan GW133 Houei-Deng
glutinous, Indica Loas GW64 Komapatai glutinous, upland Taiwan GW134 Khao-Konhdam glutinous, Indica Loas GW65 Pagaitsuitaiyaru glutinous,
upland Taiwan GW135 Pheip
glutinous, Indica Loas GW66 Airaromu glutinous,
upland Taiwan GW136 Tainung No.67
non-glutino
us, Japonica Taiwan GW67 Pazumataharu glutinous,
upland Taiwan GW137 Taichung No.65
non-glutino
us, Japonica Taiwan GW68 Nakara No.2 glutinous, upland Taiwan GW138 Taikeng No.9 non-glutinous, Japonica Taiwan
GW69 Nakabo glutinous,
upland Taiwan GW139 Taichung-Shien No.10
non-glutino
us, Indica Taiwan
GW70 Ya-A-Bi glutinous,
upland Taiwan GW140 Tai-Chung-Native No. 1
non-glutino
us, Indica Taiwan
之強度、穗型、次生支 、穗之伸出度、穗
軸、穗之落粒性、脫粒性、芒、芒色、稃尖
色、柱頭色、內外穎色澤、內外穎之軟毛、
護穎色澤、護穎長度、種皮色等
27 個質的
性狀;及葉長、葉寬、葉舌長度、株高、分
蘗數、莖徑、穗長等七個數量性狀,另外於
2000 年 第 二 期 作 種 植 於 臺 灣 大 學 試 驗 農
場,調查確認其中主要質的性狀之表現,植
株 性 狀 之 調 查 則 依 據 國 家 種 原 庫 稻 種 原 調
查標準實施。
三、統計分析
1.糯稻種原農藝性狀之相關性檢定
利用
SAS 軟體之 CORR Procedure 針
對 各 參 試 品 種 進 行 性 狀 間 之 外 表 型 相 關 係
數 分 析 , 並 進 行 相 關 係 數 之 顯 著 性 測 驗
(SAS Institute 1993)。
2.糯稻種原農藝性狀進行分群分析
根據參試品種農藝性狀,利用歐幾里得
距 離
(Euclidean distance)算出各參試品種
之遺傳歧距離:
m
d
ij2=Σλ
p(y
ip-y
jp)
2 p=1y
ip和
y
jp分別是
i 和 j 在因素軸 p (p=1, 2,
3, …, m) 上 的 因 素 分 數 (component
scores) ; λ
p為 第
p 個 因 素 之 特 徵 根
(eigenvalue)。以 SAS 軟體(SAS Institute
1993) 。 計 算 成 遺 傳 距 離 矩 陣 (genetic
distance matrix),並輸出為 NT-SYS 軟體
所 需 特 定 格 式 之
Excel 檔 案 。 再 利 用
NT-SYS (Rohlf 1989) 以
UPGMA
(unweighted pair group method with
arithmetic mean) 進 行 群 聚 分 群 (cluster
analysis),並繪出樹狀分枝圖,以求出各參
試材料間的差異。
3.糯稻種原農藝性狀之主成分分析
利 用
SAS 軟 體 之 PRINCOMP
Procedure 以各 參試品種農 藝性狀之平均
值進行主成分分析,求得各品種標準化之主
成分數值,分別以第一及第二主成分為座標
軸作族群之散佈圖,以探討解析族群間遺傳
變異之主要因子。
結果
一、農藝外表性狀遺傳變異分析
1.質的性狀調查結果(如 Table 2)
在
8 個葉的性狀調查結果,140 個糯稻
種 原 中 , 其 葉 毛 大 部 份 為 非 光 滑 葉 型
(107
個
),葉色以綠色(125 個)為主。葉角度以橫
平型
(78 個)為主,而劍葉型有直立型、中間
型、橫平行、下垂型等各類型數目相當。葉
舌色主要以白色
(126 個)為主,葉舌形狀以
二 裂 片
(136 個 )為 主 , 葉 環 色 澤 以 淡 綠 色
(127 個)為主,葉耳色以淡綠色(127 個)為
主。
在
8 個莖稈的性狀調查結果,在 140
個 糯 稻 種 原 中 除 穗 軸 無 外 表 型 變 異 均 為 下
垂型而無直立型,其莖基色以綠色
(123 個)
為主,桿角度以中間型
(93 個)為主,節間色
以金黃色(119 個)為主。而桿之強度包括 5
種類型,以中強度
(52 個)到中度強度(47 個)
為主。穗型以中間型(137 個)為主,次生支
以無次生支 為主
(98 個),穗之伸長度以中
伸出
(99 個)為主,穗軸則全部為下垂型。
在
11 個穀粒性狀之調查結果,在 140
個 糯 稻 種 原 中 , 其 穗 之 落 粒 性 以 中
(50 個)
至多
(51 個)為主,脫粒性以容易(81 個)為
主。芒之有無以無芒(87 個)為主,有芒的品
種中,其芒色包括
6 種類型,其中有 13 個
為稻草色,9 個為金黃色,13 個為褐色,2
個為紅色,11 個為紫色,5 個為黑色。稃尖
色 以 稻 草 色
(85 個)為主,柱頭色以淡綠色
(94 個)為主。內外穎色澤以褐色斑紋(57 個)
為主,內外潁之軟毛以短毛
(51 個)為主,護
穎 色 澤 以 黃 色
(59 個 )至 金 黃 色 (53 個 )為
主,護穎長度以中度(128 個)為主,種皮色
以白色
(96 個)為主。
由此可見,葉片及莖稈的各種性狀所分
之類別,除了葉角度、劍葉型及莖桿角度各
類型均有較高比例分布外,其他性狀偏向於
特定外表型;而榖粒性狀則較均勻分佈在不
同類型。
2.數量性狀調查
由
Table 3 結果可知,7 個調查性狀在
一期作與二期作間均有顯著差異。三個葉的
性狀方面:一期及二期作葉長之平均分別為
46.16 cm 及 40.58 cm,一期及二期作葉寬
之平均分別為
12.62 mm 與 11.60 mm,一
期 及 二 期 作 葉 舌 長 度 之 平 均 分 別 為
16.65
mm 及 5.87 mm。三個莖的性狀方面:一期
及二期作株高之平均分別為
125.67 cm 與
112.7 cm,一期及二期作分蘗數之平均分別
為
8.73 個與 6.51 個,一期及二期作莖徑之
平均分別為
6.31 cm 與 3.97 cm。一期及二
期作穗長之平均分別為
23.96 cm 及 22.66
cm,可知一期作所調查之 7 個數量性狀均
極顯著大於二期作。
二、數量性狀間之相關分析
2000 年一期、二期作於臺中種植的糯
稻 種 原 各 數 量 性 狀 間 之 相 關 分 析 結 果 如
Table 4。兩期作之性狀間相關值顯著性有
Table 2. The clssifications and frequencies (in parentheses) of qualitative traits of 140 glutinous rice
germplasm.
Trait Classification
and
frequency
Leaf blade pubescence
Glabrous (1), Intermediate (32), Pubescent (107)
Color of leaf blade
Light green (0), Green (125), Dark green (12), Purple tips (0), Purple
margins (3), Purple blotch (0), Purple (0)
Leaf angle
Erect (42), Horizontal (78), Droopy (20)
Flag leaf angle
Erect (39), Intermediate (32), Horizental (41), Descending (28)
Ligule color
White (126), Purple lines (12), Purple (2)
Ligule shape
Acute (4), Cleft (136), Truncate (0)
Collar color
Light green (127), Green (1), Purple (12)
Auricle color
Light green (127), Purple (13)
Basal leaf sheath color
Green (123), Purple lines (8), Light purple (2), Purple (7)
Culm angle
Erect (3), Intermediate(93), Open (44), Spreading (0), Procumbent (0)
Internode color
Green (4), Gold (119), Purple lines (7), Purple (10)
Culm strength
Strong (8), Moderately strong (52), Intermediate (47), Weak (21), Very
weak (12)
Panicle type
Compact (0), Intermediate (137), Open (3)
Secondary branching of
panicles
Abscent (98), Light (42), Heavy (0), Clustered (0)
Panicle exsertion
Well exserted (35), Moderately well exserted (99), Just exserted (3)
Partly exserted (3), Enclosed(0)
Panicle axis
Straight (0), Droopy (140)
Shattering habit
Very low (1), Low (9), Intermediate (50), Moderately high (51), High (29)
Panicle threshability
Difficult (1), Moderately difficult (58), Easy (81)
Awning
Absent (87), Short and partly awned (27)
Short and fully awned (7), Long and fully awned (19)
Awn color
Straw (13), Gold (9), Brown (13), Red (2), Purple (11), Black (5)
Apiculus color
White (1), Straw (85), Brown (28), Red (3), Red tip (4)
Purple (9), Purple tip (10)
Stigma color
White (3), Light green (94), Yellow (25), Light purple (17), Purple (1)
Lemma and palea color
Straw (1), Gold (30), Brown spots (8), Brown furrows (57), Brown (17)
Reddish to light purple (5), Purple spots (0), Purple furrows (21),
Purple (1), Black (0)
Lemma and palea
pubescence
Glabrous (19), Hair on lemma keel (36), Hair on upper portion (32),
Short hairs (51), Long hairs (2)
Sterile lemma color
Yellow (59), Gold (53), Red (3), Purple (25)
Sterile lemma length
Short (0), Medium (128), Long (12), Extra long (0), Asymmetrical (0)
Seed coat color
White (96), Light brown (22), Speckled brown (2), Brown (1), Red (13)
Table 3. Testing on mean and variance of quantitative traits of glutinous rice germplasm grown in the
spring and fall of 2000.
Trait Season
Range
Mean
SE
t-test
C891 26.0 - 72.0
46.2
0.9
Leaf length (cm)
C892 18.7 - 65.6
40.6
0.9
**
C891 8.0 - 19.0
12.6
0.2
Leaf width (mm)
C892 5.2 - 18.8
11.6
0.2
**
C891 7.6 - 41.7
16.7
0.2
Ligule length (mm)
C892 2.0 - 14.2
5.9
0.5
**
C891 87.6 - 175.7
125.7
2.0
Culm length (cm)
C892 65.2 - 166.6
112.7
1.8
**
C891 3.9 - 16.0
8.7
0.2
Culm number (No. pl
-1)
C892 2.4 - 14.6
6.5
0.2
**
C891 4.3 - 8.9
6.3
0.1
Culm diameter (mm)
C892 2.2 - 5.7
4.0
0.9
**
C891 17.1 - 38.9
24.0
0.3
Panicle length (cm)
C892 15.0 - 30.2
22.7
0.3
**
**: Significantly different at 1% level.
Table 4. Significant tests of correlation coefficients among quantitative traits of glutinous rice germplasm
grown in the spring and fall of 2000.
Season Trait
Leaf
length
Leaf
width
Ligule
length
Culm
length
Culm
number
Culm
diameter
C891
0.30**
C892
Leaf width
0.48**
C891
0.67**
0.02
C892
Ligule length
0.75**
0.39**
C891 0.70**
0.15
0.46**
C892
Culm length
0.83** 0.44** 0.59**
C891 -0.37**
-0.57**
-0.06
-0.19*
C892
Culm number
0.17* -0.29** 0.33** 0.12
C891
0.58** 0.64** 0.34** 0.23** -0.69**
C892
Culm diameter
0.47** 0.58** 0.37** 0.38** -0.26**
C891
0.75** 0.37** 0.56** 0.55** -0.51** 0.66**
C892
Panicle length
0.62** 0.57** 0.47** 0.65** -0.22* 0.56**
*,** : Significant at 5% and 1% levels, respectively.
不一致者,如葉寬與葉舌長、葉寬與株高、
葉舌長與分蘗數、株高與分蘗數,其餘性狀
間相關值顯著或極顯著之一致性很高。
三、以不同季節農藝性狀進行糯稻種原
群聚分析
以
2000 年一期、二期兩期作所調查之農
藝性狀利用歐幾里德距離算出各參試品種之
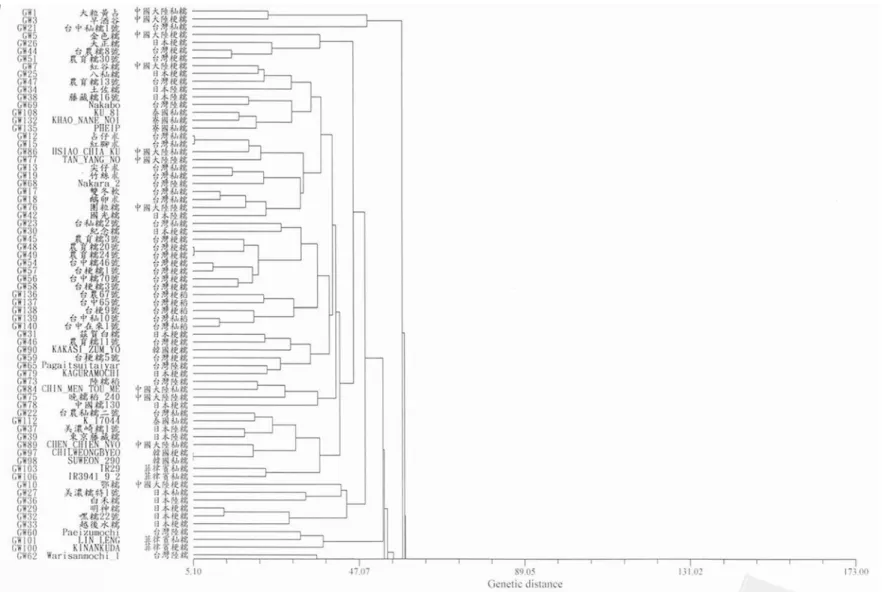
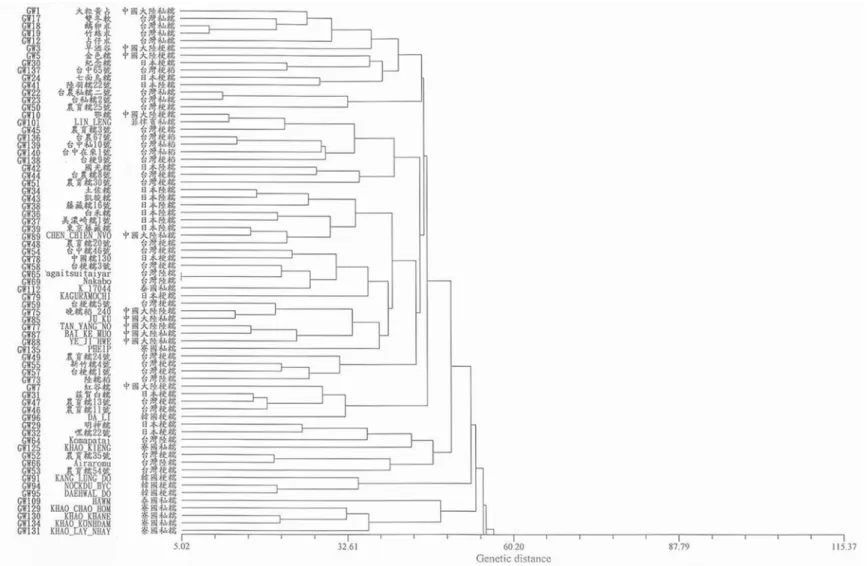
遺傳距離,再進行分群分析,其結果如
Fig. 1
及
Fig. 2。若遺傳距離越遠,則表示其兩個糯
稻種原間農藝性狀差異越大。以來源地區觀
察樹狀分群圖,同一來源地區之糯稻種原有
群聚的現象發生,在一期作中的
5 個非糯稻
對照品種
(GW136-GW140)群聚於一小群,而
依 來 源 國 家 及 糯 稻 種 類 之 不 同 形 成 若 干 分
群,例如臺灣 稻(GW45、GW48、GW49、
GW54、GW56、GW57、GW58)分別集中於
一小群;日本 稻(GW27、GW29、GW32、
Fig. 1. Dendrogram of cluster analysis based on agronomy characteristics of 135 glutinous and 5 non-glutinous rice germplasm investigated in the
first cropping season of 2000.
Cro
p
, Envi
ro
nme
nt & Bioinf
ormatics, Vol.
2, Marc
h 200
5
Fig.1. (continued).
農
藝性狀
評估糯
稻種原
遺傳歧
異性
21
Fig. 2. Dendrogram of cluster analysis based on agronomy characteristics of 135 glutinous and 5 non-glutinous rice germplasm investigated in the
second cropping season of 2000.
Cro
p
, Envi
ro
nme
nt & Bioinf
ormatics, Vol.
2, Marc
h 200
5
Fig. 2. (continued).
農
藝性狀
評估糯
稻種原
遺傳歧
異性
23
GW33 、 GW36) 及 中 國 大 陸 糯 稻 (GW4 、
GW80、GW81、GW87、GW88)等分別集中
於一小群。此外,亦可發現寮國、泰國和印
度
3 個來源地區的糯稻種原多歸屬於同一群
聚,其餘來源地區的糯稻種原也有群聚的現
象,不過因為群聚零散,不如前面來源的群
聚 現 象 明 顯 。 在 二 期 作 中 除 了 臺 中
65 號
(GW137)以外,4 個非糯稻對照品種歸屬於同
一群聚;日本陸糯(GW34、GW36、GW37、
GW38 、 GW39 、 GW43) 、 中 國 大 陸 糯 稻
(GW75、GW77、GW85、GW87、GW88)、
寮 國 秈 糯 稻
(GW129 、 GW130 、 GW131 、
GW134)的糯稻種原也各別有群聚現象,而其
餘來源地區的糯稻種原之群聚則較零散。由
此可見,以不同期作之數量性狀之分群結果
顯示在兩期作未有明顯大分群出現,而相同
來源國家種原有集中於小群之趨勢,但兩期
作分群結果並不一致。
以秈、 型稻種及陸糯稻分類觀察樹狀
分群圖,於一期作中
糯、秈糯及陸糯稻平
均分佈於各小群;於二期作之結果亦相似。
故兩季分群結果皆未有明顯之
糯、秈糯及
陸糯稻之分群。
四、以不同季節農藝性狀進行糯稻種原
遺傳距離之主成分分析
以 各 參 試 種 原 之 農 藝 性 狀 之 平 均 值 進
行主成分分析,求得各品種標準化之主成分
Table 5. Eigenvalues and proportions of the first 25 principle components among 60 markers on 33 traits
of 140 rice germplasm in the first and second cropping seasons of 2000.
First cropping season, 2000 Second cropping season, 2000 Component Eigenvalue
Difference Proportion Cumulative Eigenvalue Difference Proportion Cumulative
1 502.601 441.128 0.780 0.780 634.705 601.044 0.862 0.862 2 61.472 38.264 0.095 0.875 33.661 17.592 0.046 0.907 3 23.208 10.439 0.036 0.911 16.069 5.249 0.022 0.929 4 12.769 2.813 0.020 0.931 10.819 2.989 0.015 0.944 5 9.956 1.878 0.016 0.947 7.831 2.353 0.011 0.955 6 8.078 3.809 0.013 0.959 5.478 1.448 0.007 0.962 7 4.269 0.192 0.007 0.966 4.030 0.538 0.006 0.968 8 4.077 1.036 0.006 0.972 3.492 0.242 0.005 0.972 9 3.041 0.564 0.005 0.977 3.250 0.387 0.004 0.977 10 2.477 0.363 0.004 0.981 2.863 0.304 0.004 0.981 11 2.114 0.436 0.003 0.984 2.559 0.200 0.004 0.984 12 1.678 0.320 0.003 0.987 2.359 0.681 0.003 0.987 13 1.358 0.239 0.002 0.989 1.679 0.107 0.002 0.990 14 1.119 0.091 0.002 0.991 1.572 0.626 0.002 0.992 15 1.027 0.244 0.002 0.992 0.946 0.096 0.001 0.993 16 0.784 0.092 0.001 0.993 0.850 0.123 0.001 0.994 17 0.692 0.034 0.001 0.994 0.727 0.076 0.001 0.995 18 0.658 0.071 0.001 0.995 0.651 0.101 0.001 0.996 19 0.587 0.109 0.001 0.996 0.550 0.102 0.001 0.997 20 0.478 0.086 0.001 0.997 0.448 0.090 0.001 0.997 21 0.392 0.036 0.001 0.998 0.358 0.048 0.001 0.998 22 0.356 0.136 0.001 0.998 0.310 0.067 0.000 0.998 23 0.220 0.010 0.000 0.999 0.243 0.008 0.000 0.999 24 0.210 0.053 0.000 0.999 0.235 0.033 0.000 0.999 25 0.158 0.008 0.000 0.999 0.202 0.044 0.000 0.999
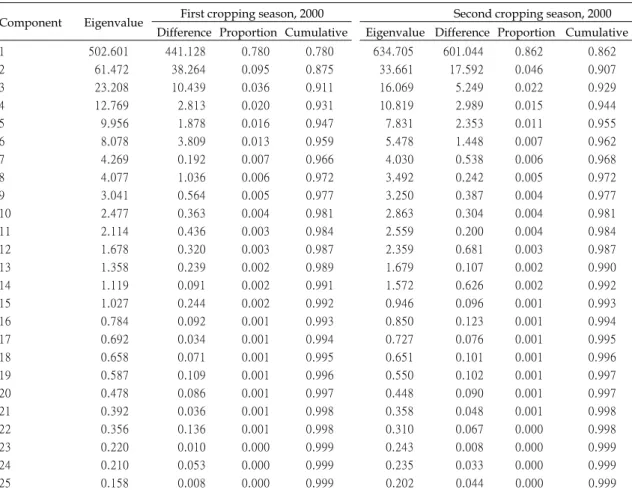
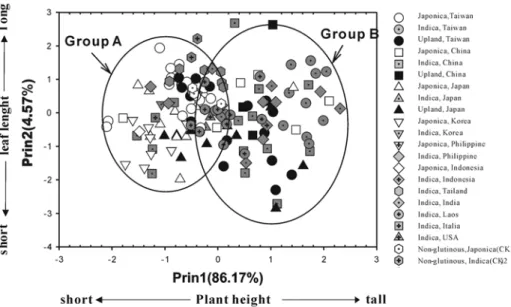
數值,兩個期作結果如
Table 5。由主成分
分析結果,在
2000 年一期作所得的特徵根
(eigen value),由 Table 5 可知前三個主成
分所累加的變異量已達
91.14%。由 Table 6
可 知 , 第 一 主 成 分 裡 特 徵 向 量
( e i g e n
vectors)以株高為最主要的影響因子,第二
主成分以葉長為最主要的影響因子,第三主
主成分以葉舌長度為主要影響因子。由於第
一 及 第 二 成 分 之 累 積 解 釋 變 異 量 已 達
87.5%,故由各品種以第一與第二主成分為
座標軸作品種之
2D 散佈圖,如 Fig. 3。可
分 別 以 農 育 糯
20 號 品 系 之 株 高 (平 均 約
121.8 cm)為 分 界 點 , 以 菲 律 賓 糯 品 系
Kinankuda 之葉長(平均為 45.8 cm)為分群
分 界 點 。 以 來 源 地 區 分 析 , 大 約 可 以 分 為
A、B 兩群。韓國糯稻、日本糯稻、日本陸
糯稻、臺灣
糯稻、美國糯稻(僅 1 個種原)、
義大利糯稻(僅 1 個種原)及 5 個非糯稻對照
品種主要分布於
A 群中;而寮國糯稻、印
度糯稻、臺灣秈糯稻、泰國糯稻、及中國大
Table 6. Eigenvectors of the 32 traits on the first to the 5
thprincipal components on 140 rice accessions the
first and second cropping seasons of 2000.
First cropping season Second cropping season Trait Prin1 Prin2 Prin3 Prin4 Prin5 Prin1 Prin2 Prin3 Prin4 Prin5 Leaf length (cm) 0.361 0.740 0.170 -0.354 0.063 0.363 0.855 0.097 0.261 0.087 Leaf width (mm) 0.018 0.067 0.191 -0.085 0.048 0.045 0.088 0.249 -0.245 -0.301 Leaf pubescence 0.000 -0.017 -0.026 -0.021 -0.046 0.003 -0.005 -0.064 0.088 -0.046 Leaf color 0.001 0.007 -0.022 0.015 0.005 -0.001 -0.002 -0.003 0.003 -0.007 Leaf sheath color 0.007 0.002 -0.023 0.030 0.029 0.010 -0.007 0.071 -0.048 0.079 Leaf angle 0.020 0.049 0.302 0.293 0.454 0.001 -0.025 0.358 -0.180 0.463 Flag leaf angle 0.030 0.016 0.272 0.244 0.296 -0.007 -0.020 0.305 -0.155 0.316 Ligule length (mm) 0.141 0.416 -0.577 0.579 -0.198 0.057 0.183 -0.089 -0.037 -0.017 Ligule color 0.004 0.002 -0.010 0.016 0.015 0.007 -0.007 0.050 -0.038 0.046 Ligule shape -0.002 -0.003 0.000 -0.001 0.011 0.001 0.000 0.002 0.003 0.010 Collar color 0.006 0.002 -0.019 0.013 0.031 0.011 0.002 0.075 -0.083 0.054 Auricle color 0.003 0.001 -0.009 0.007 0.017 0.005 -0.003 0.051 -0.039 0.027 Plant height (cm) 0.912 -0.388 -0.043 -0.006 0.020 0.923 -0.355 -0.074 -0.069 0.021 Tilling number -0.027 -0.097 -0.336 -0.072 0.061 0.013 0.070 -0.439 0.126 0.341 Culm angle 0.013 -0.027 -0.065 0.038 0.015 0.006 -0.028 0.022 0.077 0.073 Culm diameter (mm) 0.014 0.074 0.074 -0.017 -0.027 0.011 0.031 0.044 -0.016 -0.107 Internode color 0.005 -0.003 -0.015 0.021 0.030 0.013 -0.030 0.038 -0.027 0.075 Culm strength 0.031 -0.123 -0.137 0.115 0.102 0.034 -0.076 -0.053 0.059 -0.049 Panicle length (cm) 0.109 0.246 0.201 0.176 -0.180 0.088 0.063 0.314 -0.154 -0.533 Panicle type 0.005 0.006 -0.002 0.001 0.028 0.002 -0.003 -0.010 -0.012 0.013 Secondary panicles -0.003 0.000 -0.001 -0.039 -0.036 -0.001 0.013 -0.013 0.000 -0.015 Panicle exsertion -0.008 0.033 -0.050 -0.025 0.010 -0.002 0.101 0.003 -0.091 -0.056 Shattering habit -0.006 -0.011 -0.064 0.199 0.381 0.007 0.005 -0.071 -0.119 0.106 Panicle threshability -0.002 0.016 -0.044 0.215 0.418 0.017 -0.009 -0.149 -0.250 0.165 Awning 0.017 -0.131 0.367 0.322 -0.471 -0.003 -0.235 0.262 0.709 -0.017 Awn color 0.018 -0.044 0.204 0.200 -0.198 0.005 -0.116 0.171 0.348 -0.011 Apiculus color 0.015 0.048 0.140 0.143 -0.037 0.011 -0.015 0.290 -0.009 0.109 Stigma color 0.006 0.010 0.067 0.079 0.003 0.012 -0.024 0.150 -0.024 0.076 Lemma and palea color 0.006 0.034 0.166 0.225 -0.069 0.011 0.001 0.320 -0.061 0.273 Lemma and palea pubescence -0.014 -0.073 -0.015 -0.035 -0.122 -0.004 -0.039 -0.011 0.157 0.049 Sterile lemma color -0.002 0.023 0.108 0.093 -0.056 -0.004 0.000 0.178 0.010 0.088 Sterile lemma length 0.001 0.012 0.037 0.036 0.024 0.004 -0.010 0.011 -0.026 0.012 Seed coat color 0.020 0.016 0.027 0.114 0.004 0.008 -0.042 0.093 0.002 -0.045