不同雌性素受體對於攝食及體重的調控作用與下視丘神經胜肽表現量的影響; THE EFFECT OF DIFFERENT ESTROGEN RECEPTOR AGONISTS ON FOOD INTAKE,BODY WEIGHT AND HYPOTHALAMIC NEUROPEPTIDE EXPRESSION IN
55
0
0
全文
(2) 圖目錄. 圖一 腦部冠狀切片顯示弓形核(ARC)之解剖位置圖。………………24 圖二 腦部冠狀切片顯示室旁核(PVN)之解剖位置圖。………………25 圖三 給予 0.5 與 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大 白鼠子宮重之影響。……………………………………………………26 圖四 給予 0.5 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大白 攝食之影響。……………………………………………………………27 圖五 給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大白鼠攝 食之響 。………………………………………………………………28 圖六 給予 0.5 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大白鼠 體重之影響。……………………………………………………………29 圖七 給予 1 mg/Kg/days 的甲型及乙型雌性素受體致效劑對大白鼠 體重變化之影響。………………………………………………………30 圖八 皮下注射甲型及乙型雌性素受體致效劑對大白鼠飼料效能之 影響。……………………………………………………………………31 圖九 大白鼠接受 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑後下 視丘室旁核神經胜? Y 的影響。………………………………………32. 2.
(3) 圖十 大白鼠接受 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑後下 視丘弓形核神經胜? Y 的影響。………………………………………33 圖十一 給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大白鼠 下視丘弓形核及室旁核神經胜? Y 的表現量的影響。………………34 圖十二 大白鼠接受 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑後 下視丘弓形核甲型促黑激素的影響。…………………………………35 圖十三 給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對大白鼠 下視丘弓形核甲型促黑激素的表現的響。……………………………36. 3.
(4) 英文摘要. Estrogen does not only control reproduction, but also affects other physiology functions, such as modulation of feeding behavior and fat storage. In order to delineate whether eatrogen receptor alpha (ERa) or beta (ERß) is involved in the action of estrogen in energy balance, we treated ovarietomized rats with a specific ERa agonist PPT (0.5 or 1 mg/Kg) or a specific ERß agonist DPN (0.5 or 1 mg/Kg) for 6 days, and observed changes in food intake and body weight gain. We found that treatment of PPT (0.5 and 1 mg/Kg) suppressed feeding and body weight. In contrast, DPN (0.5 and 1 mg/Kg) only decreased food intake. Hypothalamic levels of neuropeptide Y and a-MSH were detected by immunohistochemistry in the animals. Semiquantification with a computerized image analysis system revealed no significant difference of these peptide levels among groups. These data suggest neuropeptide Y and a-MSH may not be involved in the regulation of energy balance by estrogen.. 4.
(5) 中文摘要. 雌性素(Estrogen)是由雌性動物的卵巢所分泌出來的賀爾蒙,其 在於調控女性的生殖及相關的生理功能上,扮演了很重要的作用。除 此之外,雌性素可以影響部分由中樞所調節的攝食行為(Wallen et al., 2001; Geary et al., 2001)。實驗中,我們給予不同雌性素受 體的致效劑,觀察其對大白鼠攝食及體重的調控作用,並分析下視丘 神經胜? 的表現。我們以皮下注射的方式,連續給予六天 0.5 或 1 mg/Kg/day 的甲型雌性素受體致效劑(PPT)或乙型雌性素受體致效劑 (DPN),結果發現,不論 0.5 或 1 mg/Kg/day 的 PPT 都可以顯著的下 降大白鼠的攝食與體重;然而兩種劑量的 DPN 卻都只能下降攝食,對 體重則不產生影響;另外,以免疫組織染色的方式,觀察下視丘神經 胜? Y 及甲型促黑激素的表現;結果顯示不論給予何種雌性素受體致 效劑,下視丘神經胜? Y 及甲型促黑激素的表現皆不受影響。由以上 結果可以推知,內生性雌性素下降攝食的作用,可能經由活化其兩種 受體;然而它對體重的調控則只活化了甲型雌性素受體;且其對能量 平衡的影響可能不是經由下視丘神經胜? Y 及甲型促黑激素來達成 的。. 5.
(6) 前言. 文明的進步,伴隨著生活水準不斷的提高,人們的飲食生活習慣 也因此而有所改變,隨之而越來越多肥胖的問題,已為人們所重視; 肥胖除了會影響生活外,許多疾病,像是糖尿病、心血管疾病及高血 壓、高血脂,都和肥胖有著密不可分的關係;然而改善肥胖問題,除 了增加身體的活動量之外,飲食的調控也是一門很大的學問,原因在 於人們常常沒辦法有效的控制自己的食慾;食慾的調節中樞位在下視 丘,由許多極為複雜的神經元互相支配調控,另外有一些周邊或腸道 賀爾蒙也參與著這個作用,有些機制是我們已經知道的,但仍有許多 對攝食相關的機轉我們仍不明瞭。 肥胖問題在我們生活中隨處可見,它尤其的困擾著女性,在 1998 的一篇報告中指出(Geary et al.,1998),在女性中度肥胖(超出正 常體重的百分之三十到五十)者佔百分之十一,高出男性的百分之 八;而嚴重肥胖(高於正常體重的百分之五十以上)的比率,女性則 為男性的四倍;因此,肥胖問題顯然的在女性較為嚴重,許多飲食所 引起的問題,像是暴食症或厭食症也往往發生在女性身上,原因為何 至今仍不非常清楚,但這種差異,讓我們聯想到性賀爾蒙和攝食間的 相關。 6.
(7) 雌性素和攝食間的關係 雌性素(Estrogen)是由雌性動物的卵巢所分泌出來的賀爾蒙,其 在於調控女性的生殖及相關的生理功能上,扮演了很重要的作用,其 次和學習記憶、下視丘功能、以及部分癌症的形成等之間的關係也都 有相關報告。除此之外,性賀爾蒙也可以影響部分由中樞所調節的攝 食行為(Wallen et al., 2001),而性賀爾蒙中又以雌性素和攝食行 為的調控最為相關。許多報告也指出在雌性動物的生理週期中,血液 中的雌性素(主要是雌二醇 estradiol)濃度和攝食行為有很大的相 關性。以大白鼠為例,在雌二醇分泌量最高的動情期後,有減少攝食 的情 (Geary & Asarian, 2001);另外在人類也可以發現,動情素 分泌較高時期的排卵前期,會有進食量下降的情況,而動情素較低的 黃體期則伴隨著較高的攝食量,且也較易囤積脂肪(Geary, 2001)。 由以上證據可以知道雌性素的確在攝食上扮演一重要的角色,另 外我們可以在更年期後雌性素量驟減、或失去活性的婦女身上看到代 謝率下降以及白色脂肪組織堆積(Heine et al.,2001)。在實驗動物 上為了研究相關的問題,我們通常以切除卵巢的動物來模擬停經後婦 女的內分泌狀態; Liang 的報告也指出,在切除卵巢的大鼠可見其 體重及腹部脂肪的堆積增加;而這個情形可以在外給雌性素後恢復 (Liang et al., 2002)。 7.
(8) 因此雌性素,除了調節女性生殖功能之外,對攝食、體重及身體 能量的代謝影響也是不容小覷的。. 雌性素受體 雌性素之作用,主要是透過其接受器而達成的。雌性素受體是屬 於類固醇受體家族的一員,當其與 ligand 結合並活化後,雌性素接 受器會形成雙體(dimer)並與雌性素反應序列(estrogen responsive element,ERE)結合,並改變下游基因的轉錄活動(Moras & Gronemeyer, 1998)。 雌性素受體為 76KD 的蛋白,具有八個 exon。傳統雌性素受體是 於一九八六年由 Green 發表(Green et al., 1996),早期一般都認 為它是唯一的雌性素受體,直到一九九六年,Kuiper 在大鼠的前列 腺及卵巢上又發現另一種新的雌性素受體(Kuiper et al,1996),因 此現在將雌性素受體,分為甲型(estrogen receptor alpha),也就 是傳統的雌性素受體,以及乙型(estrogen receptor beta),也就是 新發現的雌性素受體。 傳統的雌性素受體(甲型雌性素受體)基因序列長度為 2092 個 鹼基對(base pair) ,可轉譯成 595 個氨基酸的蛋白質,它具有六個 功能區(domains),分別為一個基因結合區域(DNA binding domain, 8.
(9) DBD) ,一個樞紐區(hinge area) ,一個賀爾蒙結合區(ligand binding domain, LBD),和兩個執行轉活化(transactivation)區(AF-1 與 AF-2) 。而乙型雌性素受體的基因序列長度為 1560 個鹼基對,可以轉 譯成含有 477 個氨基酸的蛋白質,其與甲型受體相同亦包含六個區域 (Gustafsson, 1999; Pettersson & Gustafsson, 2001)。 乙型雌性素受體與甲型的基因結合區有 97﹪的相同度,賀爾蒙 結合區有 59﹪的相同度,乙型和甲型雌性素受體與雌二醇 (estradiol)的親和力約略相同(Gustafsson, 1999)。 甲型雌性素受體廣泛的表現在子宮、陰道和腦中的下視丘弓形核 (arcute nucleus,ARC)等處;而乙型受體則主要表現在肺臟、卵 巢以及前列腺和下視丘視旁核(paraventricular nucleus, PVN); 另外骨骼及腦下垂體則會同時表現兩種受體(Gustafsson, 1999)。 雖然兩種受體皆可在下視丘不同的區域表現,但是否兩種受體都 參與了攝食及能量代謝的調控仍未明朗。. 雌性素受體致效劑 由與雌二醇與甲型及乙型受體的親合力相仿,無法分辨兩者之作 用,因此必須選擇特異性較高的致效劑,方能了解不同型的雌性素受 體在調控能量平衡之功能中的重要性。 9.
(10) PPT (1,3,5-tris(4-hydroxyphenyl)-4-propyl-1H-pyrazole), 是一個甲型雌性素受體致效劑,它的分子量為 3860.45,其對甲型雌 性素受體的親和力是乙型的 410 倍(Stauffer et al., 2000);另外 DPN(2,3-bis(4-hydroxyphenyl)-propionitrile )則為乙型雌性素 受體致效劑,其對乙型受體有較高的親和力約為甲型的 70 倍左右 (Meyers et al., 2001; Kraichely et al., 2000)。. 神 經 胜 ? Y( neuropeptide Y) 神經胜? Y 不論是在大鼠或是其他哺乳類身上皆是一個強效的 促進攝食的訊號(Clark et al., 1984; Clark et al., 1985; Sahu & Kalra, 1993)。1980 年代即有研究指出,神經胜? Y 能有效的刺 激食慾、增加攝食(Clark et al., 1984; Clark et al., 1987),神 經胜? Y 為一 36 個氨基酸所組成的胜? (peptide),它廣泛地存在中 樞與周邊神經系統。下視丘的神經胜? Y 細胞主要由位於弓形核, 再投射至室旁核與其它神經核,例如:下視丘外側區(LH) 、背側核 (dorsomedial nucleus)等(Billington et al., 1994; Inui, 2000; Kalra et al., 1999)。 當能量缺乏(例如:在禁食情況之下)或代謝增加時,神經胜? Y 神經元受到活化,其合成和分泌量也會增加(Baskin et al., 1999; 10.
(11) Chance et al., 1994)。. 神經胜?. Y 與攝食行為及能量調節之關係. 經由腦室或室旁核注射神經胜? Y 後,會減少棕色脂肪(brown adipose tissue; BAT)的產熱作用(thermogenesis)、降低能量消耗 (energy expenditure)及增加食物之攝取,因而促進脂肪儲存,最後 導致肥胖(Billington et al., 1991; Egawa et al., 1991; Zarjevski et al., 1993 ); 此外,中樞給予神經胜? Y 受體之拮抗劑後,則 會抑制自發性及神經胜? Y 所誘導的攝食行為(Dube et al., 1995; Ishihara et al., 1998; Kanatani et al., 1996; Polidori et al., 2000)。 在神經胜? Y 受體剔除的大白鼠也有攝食量及體重降低之情形 (Inui. 2000; Michel et al., 1998),所以神經胜 Y 在增加攝食調 控作用中扮演極重要之角色。 文獻指出,雌性素對攝食及體重的調控作用可能是經由活化或抑 制下視丘神經元的代謝而達成的(Bonavera et al., 1994); 神經胜 ? Y細胞本體所在的下視丘弓形核及其投射到下視丘的其他區域也 正好是類固醇荷爾蒙的作用區域(Morrel et al.,1986; Sar et al., 1990)。有報告指出,性荷爾蒙可以調控下視丘神經胜? Y的產生與 11.
(12) 釋放(Sahu et al., 1992; Sahu et al., 1994)。2002年的報告指 出動物在切除卵巢後顯著的減少神經胜? Y傳訊者核糖核酸(mRNA) 的表現,而在外給雌二醇後,可使傳訊者核糖核酸下降的情形回復 (Baskin et al.,1995),因此實驗中我們想知道活化甲型或乙型雌性 素受體是否會經由影響下視丘神經胜? Y的神經元表現量,進而對攝 食及體重產生作用。. 甲 型 -促 黑 激 素 荷 爾 蒙 與 攝 食 行 為 之 關 係 在許多的哺乳類動物(包括靈長類)中樞神經系統中皆有黑激素 ( melanocortin ) 的訊息存在( Koegler et al., 2001)。而甲型促 黑激素荷爾蒙是屬於下視丘中促黑激素系統中的一員,它是 pro-opiomelanocortin ( POMC ) 前驅蛋白分裂的一個片段 ( Alvaro et al., 1997 ),為含有 13 個胺基酸的多胜? 類,其與接受器結合 會抑制攝食行為( Grill et al., 1998 ) 。 中樞神經的促黑激素為下視丘中一個調節攝食的重要角色 ( Mountjoy et al., 1994 )。中樞給予選擇性促黑激素第 3/4 受體 的促進劑 MT II 會降低食物的攝取( Fan et al., 1997 ); 而腦室注 入一個選擇性促黑激素第 4 受體的拮抗劑 HS014 則會增加攝食量 ( Kask et al., 1998 )。因此,腦中的促黑激素第 3/4 受體的確參 12.
(13) 與降低攝食行為的作用。 在中樞或週邊給予雌二醇後,可改變下視丘 POMC 的表現量 (Pelletier, 1993) ,推論 POMC 的增加可上升甲型促黑激素的分泌, 進而產生抑制攝食的作用。因此實驗中我們想知道甲型或乙型雌性素 受體是否會經由影響下視丘甲型促黑激素的神經元,進而對攝食及體 重產生作用。. 實驗目的:. 1. 分別給予甲型或乙型雌性素受體致效劑觀察雌性素對攝食及體重 的調節,是經由何種雌性素接受體而達成的。. 2. 以免疫組織化學染色的方式測量神經胜? Y 和甲型促黑激素的表 現量,以推測甲型和乙型雌性素受體對攝食和體重的調控作用, 是否經由抑制神經胜? Y 神經元而產生作用。. 13.
(14) 材料方法. 一 .實 驗 藥 物 一般藥物均購自於 Sigma、Merck 等公司;甲型雌性素受體 致效劑 PPT 與乙型受體致效劑 DPN 購於 Tocris;PPT 與 DPN 均溶 於 50%Dulbecco’s phosphate buffered saline (DPBS)+50% DMSO 的溶液中。. 二 .實 驗 動 物 動物均為國科會 Sprague-Dawley (SD)品系的雌性大白鼠, 體重介於 250-300 克,飼養於光照週期 12 小時光照 (0700-1900),12 小時黑暗(1900-0700),室溫則調控在 21± 2 ℃間。實驗前和實驗間均給予其充足的飲水和食物。. 三 .切 除 卵 巢 手 術 動物以 Chloral hydrate (360 mg/kg)麻醉後,由背部將兩 側卵巢去除,傷口縫合後,由腹腔施打 penicilline (3000 IU/rat),手術後將動物個別飼養於塑膠籠中兩週,使其傷口復 原。 14.
(15) 實驗一. 皮下給予甲型及乙型雌性素受體致效劑對切除卵巢的 大白鼠體重、攝食及飼料效能的影響 在切除卵巢手術的兩週後,開始連續每天給予大白鼠皮下注 射甲型雌性素受體致效劑(PPT, 0.5 或 1 mg/ml/Kg)或乙型雌 性素受體致效劑(DPN, 0.5 或 1 mg/ml/Kg),對照組則單獨給予 溶劑(DPBS+DMSO, 1 ml/Kg),藥物施打前均先稱取飼料及體重 的變化;第一次注射藥物當天設定為第 0 天(day0),最後一次 測量飼料及體重的時間為第五天注射藥物之前。 飼料效能的計算方法: 將動物累積的總體重變化除以總進食的克數,結果以百分比 表示 BW gain(g)/ food intake(g)×100%. 以上實驗各組結果以 mean±SEM 表示,並採單向變方分析 、. (ANOVA)及 Tukey s test 來檢測,當機率<0.05 時(P<0.05), 即視為有顯著性的差異。. 15.
(16) 實驗二. 甲、乙型雌性素受體致效劑對神經胜? Y 及甲型促黑激 素表現量的調節作用 部分動物在實驗一結束(第六次給藥)後 4 小時,以 chloral hydrate (360 mg/Kg)麻醉,在呼吸心跳尚未停止前,將胸腔切 開,並以止血鉗夾住下腔靜脈,其後在左心室注入 1000 IU 的肝 素(heparin) ,以 18 號靜脈留置針穿刺左心室至主動脈出口後, 依次注入含 2%亞硝酸鈉(Na nitrite)之生理食鹽水,及 4% 的副福馬林(paraformaldehyde) 。灌流後將腦取出,以 4%的副 福馬林浸泡 24 小時後,更換於 30%的蔗糖(sucrose)溶液中, 並置於 4℃冰箱中直至切片。將組織置於-20℃的冷凍切片機做 40μm 連續冠狀切片,切片後保存在含 0.1M 磷酸氫鈉緩衝液、30 %蔗糖、30% ethylene glycol、1% polyvinyl prolidone 的 抗凍液中。. 組織以每隔六片取一片的方式,進行神經胜? Y 免疫組織化 學法的染色。先將組織從抗凍液中取出,以 0.05M 的 Postassium phosphate buffered saline(KPBS)緩衝液將組織清洗數次, 再以 1%的雙氧水( H2O2)減少內生性過氧化? 的反應,其後再以 KPBS 清洗。以神經胜? Y 的初級抗體(1:100000; Diasorin) 16.
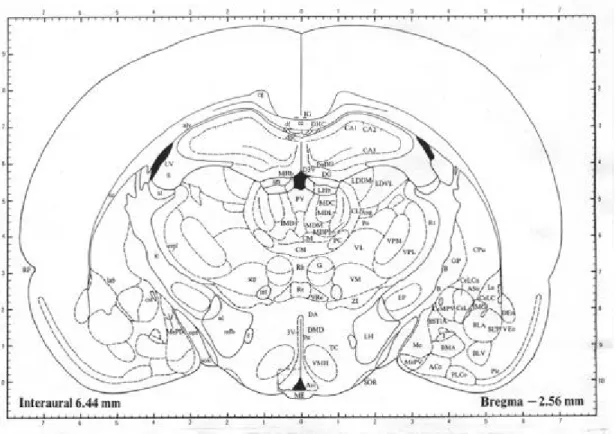
(17) 浸泡 48 小時(4℃下)並以 KPBS 清洗後再與稀釋 600 倍的次級 抗體(biotin-conjuated anti-rabbit IgG; Jackson Immuno Research Laboratories, West Grove, USA)於室溫下反應 1 小 時;清洗後再與結合有過氧化? (horse-radish peroxidase; HRP)的卵蛋白-生物素複合體( Avidin-biotin complex; Vector laboratories; Burlingame, USA)反應 1 小時(室溫下)以放 大反應,之後以含有 175 mM 醋酸鈉、2.5 mg/ml NiSO4、 0.2 mg/ml diaminobenzidine 與 0.025%過氧化氫的溶液呈色。. 甲型促黑激素荷爾蒙的免疫組織化學與神經胜? Y 類似,唯 甲型促黑激素荷爾蒙初級抗體稀釋濃度為 1:50000(Chemicon) , 而次級抗體為抗綿羊之抗體(biotin-conjuated anti-sheep IgG)。. 染色完成後,將組織切片置於光學顯微鏡下依據圖譜上的解 剖位置(Paxinos & Watson, 1986),以影像分析系統(Optimas) 分析下視丘弓形核(圖一;A-P:Bregma -2.30∼-3.30mm,D-V: dura +9.6∼10.2mm,M-L:±0.2∼0.8mm)及室旁核(圖二;A-P: Bregma -1.3∼-2.12, D-V:dura +7.8∼8.4mm,M-L:0∼±0.8mm) 17.
(18) 兩處神經胜? Y 的分佈濃度,並計算所染到的神經纖維(有顏色 的部分)佔整個分析範圍中的百分比,將結果進行統計分析。 甲 型促黑激素荷爾蒙則只分析弓形核部分的含量,此外亦以人工方 式計算弓形核中甲型促黑激素細胞本體的數量。. 以上實驗各組結果以 mean± SEM 表示,並採單向變方分析 、. (ANOVA)及 Tukey s test 來檢測,當機率<0.05 時(p < 0.05), 即視為有顯著性的差異。. 18.
(19) 三、結果. 在連續六天給予皮下注射 0.5 mg/Kg/day 的甲型雌性素受體 致效劑(PPT)後,可以顯著的增加子宮的重量(圖三 a,與對照 組相比較,p < 0.05),相同的在給予皮下注射 1 mg/Kg/day 的 甲型雌性素受體致效劑(PPT)後,子宮重比起對照組也明顯的 增重(圖三 b,p < 0.05),且在給予兩種劑量的 PPT 後,其子宮 重皆顯著的高於 DPN 的組別(p < 0.05)。. 然而在連續六天給予皮下注射 0.5 或是 1 mg/Kg/day 的乙 型雌性素受體致效劑(DPN)後對子宮重卻沒有顯著的影響(圖 三 a、b)。. 19.
(20) 實驗一、皮下給予甲型及乙型雌性素受體致效劑對切除卵巢的雌 大白鼠攝食、體重及飼料效能的影響. 攝食. 連續每天給予皮下注射 0.5 mg/Kg/day 甲型雌性素受體致效 劑(PPT)可顯著的降低第 1-5 天每天的累積攝食(圖四,p < 0.05) ;在給予乙型雌性素受體致效劑(DPN)後,顯著的減少前 四天的累積攝食(圖四,p < 0.05),但其對累積第 5 天的攝食 則沒有作用;另外在第 3、4 及第 5 天 PPT 和 DPN 的累積攝食量 間有顯著的差異(圖四, p < 0.05)。 在連續給予皮下注射 1 mg/Kg/day 甲型雌性素受體致效劑 (PPT)後,可顯著的減少累積第 1-5 天每天的累積攝食(圖五, p < 0.05);相同的在給予乙型雌性素受體致效劑(DPN)後,顯 著的減少第 1-5 天每天的累積攝食(圖五,p < 0.05)。. 20.
(21) 體重. 給予 0.5 mg/Kg/day 甲型雌性素受體致效劑(PPT)後,可 顯著的減少第 2-5 天的累積體重變化(圖六,p < 0.01),給予 0.5 mg/Kg/day 乙型雌性素受體致效劑(DPN)後,只顯著減少第 4 天的累積體重變化(圖六,p < 0.01);另外在給藥後的第 2-5 天 PPT 和 DPN 的體重增加間有顯著差異(圖六,p < 0.05)。. 給予 1 mg/Kg/day 甲型雌性素受體致效劑(PPT)後,可顯 著的減少累積第 1-4 天的體重變化(圖七,p < 0.01);然而乙 型雌性素受體致效劑(DPN)對體重的影響則並不明顯;另外在 給藥後的第 4 及第 5 天 PPT 和 DPN 的增加體重間有顯著的差異(圖 七,p < 0.05)。. 飼料效能. 給予 0.5 mg/Kg/day 甲型雌性素受體致效劑(PPT)後,可 顯著的減少飼料效能(圖八 a,p < 0.01),且與 DPN 給予的組別 有顯著的差異(p < 0.01),而 0.5 mg/Kg/day 的乙型雌性素受 21.
(22) 體致效劑(DPN) ,卻沒有辦法減少飼料效能(圖八 a) 。給予 1 mg/ Kg/day PPT 或 DPN 後,其結果與低劑量相似(圖八 b)。. 實驗二、甲、乙型雌性素受體致效劑對神經胜? Y 及甲型促黑 激素表現量的調節作用. 神經胜? Y 在下視丘主要以神經纖維分佈為主,因此以電腦 影像分析系統來計算此兩區纖維的分佈百分比,以定性來看兩種 雌性素致效劑對於切除卵巢大白鼠室旁核(圖九)及弓形核(圖 十)神經胜? Y 含量並無太大影響。. 電腦分析顯示,當大白鼠給予六天的甲型雌性素受體致效劑 PPT(1 mg/Kg/day)後,其下視丘神經胜? Y 室旁核與弓形核的 表現量與控制組相比較之下,並不具有顯著性的差異(圖九 a,p > 0.05);相同的,皮下注射乙型雌性素受體致效劑 DPN(1 mg/Kg/day)亦不影響下視丘室旁核與弓形核神經胜? Y 的表現 量(圖十一 b,p > 0.05)。. 甲型促黑激素在下視丘弓形核的表現分別以人工計數神經 22.
(23) 細胞的數目及以電腦影像分析系統來計算此區纖維的分佈百分 比。. 當給予甲型雌性素受體致效劑 PPT(1 mg/Kg/day)後,其下 視丘甲型促黑激素不論是在弓形核的神經元數目(圖十二)或分 佈量(圖十三)與控制組相比較之下,並不具有顯著性統計的差 異(p > 0.05) ;相同的,乙型雌性素受體致效劑 DPN(1 mg/Kg/day) 也不會改變弓形核甲型促黑激素的神經元數目及分佈量(圖十 二、十三,p > 0.05)。. 23.
(24) 圖一 腦部冠狀切片顯示弓形核( ARC)之解剖位置圖。3V:第三腦室;ME: memdian eminece,中央處;方框處為分析部位。. 24.
(25) 圖二 腦部冠狀切片顯示室旁核(PVN)之解剖位置圖。3V:第三腦室;方 框處為分析部位。. 25.
(26) a #**. 0.9 0.8. 子宮重(g). 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0 CON. 子宮重(g). b. PPT. **. 1 0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1 0 CON. 圖三. DPN. DPN. PPT. 給予 0.5 mg/Kg/day (a 圖)與 1 mg/Kg/day (b 圖)的甲型及乙 型雌性素受體致效劑對去除卵巢的大白鼠子宮重之影響( n = 7-12)。 各組以 mean ±SEM 表示,**p < 0.01 vs. CON。(CON: 對照組,DPN: 乙型雌性素受體致效劑,PPT:甲型雌性素受體 致效劑) 26.
(27) CON. 160. DPN PPT. accumulated food intake (g). 140 120 *. 100. *. 80. * *. 60 *. **. 40 20. *. **. *. 0 day1. day2. day3. day4. day5. 圖四 連續給予 0.5 mg/Kg/day 的甲型及乙型雌性素受體致效劑對去除卵巢 的大白鼠攝食之影響( n = 7-12)。 各組以 mean ±SEM 表示,* p < 0.05 vs. CON;** p < 0.01 vs. CON。(CON:對照組,DPN: 乙型雌 性素受體致效劑,PPT:甲型雌性素受體)。 27.
(28) CON DPN. 160. PPT. accumulated food intake (g). 140 120. *. 100. * **. 80. * **. 60 *. 40 20. *. **. **. *. 0 day1. day2. day3. day4. day5. 圖五 連續給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對去除卵巢的 大白鼠攝食之影響( n = 8-10)。 各組以 mean ±SEM 表示,* p < 0.05 vs. CON;** p < 0.01 vs. CON。(CON:對照組,DPN: 乙型雌性素受 體致效劑,PPT: 甲型雌性素受體致效劑). 28.
(29) 20. CON DPN. Body weight gain (g). 15. PPT. 10 *. 5 0 day1 -5 -10. day2. *. day3. day4. day5. *. * **. -15. 圖六 連續給予 0.5 mg/Kg/day 的甲型及乙型雌性素受體致效劑對去除卵巢 的大白鼠體重之影響( n = 7-12)。 各組以 mean ±SEM 表示,* p < 0.05 vs. CON;** p < 0.01 vs. CON。(CON:對照組,DPN: 乙型雌 性素受體致效劑,PPT: 甲型雌性素受體致效劑)。. 29.
(30) CON. 10. *. DPN PPT. 5 Body weight gain (g). *. 0 day1. -5. day2 **. day3. day4. day5. **. -10. ** **. -15. 圖七 連續給予 1 mg/Kg/days 的甲型及乙型雌性素受體致效劑對去除卵巢 的大白鼠體重變化之影響( n =8-10)。 各組以 mean ±SEM 表示,* p < 0.05 vs. CON;** p < 0.01 vs. CON。(CON:對照組,DPN: 乙型 雌性素受體致效劑,PPT: 甲型雌性素受體致效劑). 30.
(31) a. feed efficiency %. 15 10 5 0 CON. -5. DPN. PPT. -10 -15. **. b. feed efficiency %. 10 5 0 -5. CON. DPN. PPT. -10 -15 **. -20. 圖八 皮下注射甲型及乙型雌性素受體致效劑對去除卵巢的大白鼠飼料效 能之影響( n = 7-12)。 各組以 mean ±SEM 表示,** P < 0.01 vs. CON。a 圖為給予 0.5 mg/Kg/day;b 圖為 1 mg/Kg/day 的 PPT 或 DPN。 (CON:對照組,DPN: 乙型雌性素受體致效劑,PPT:甲型雌性素受體 致效劑)。 31.
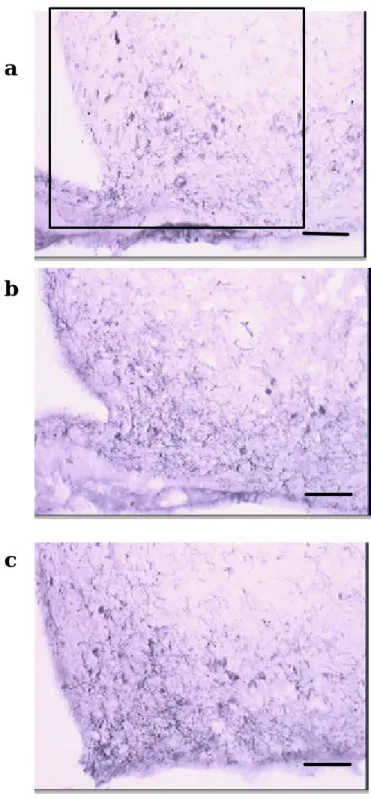
(32) a. b. c. 圖九 去除卵巢的大白鼠接受 1 mg/Kg/day 的甲型(PPT)及乙型(DPN) 雌性素受體致效劑後下視丘室旁核神經胜? Y 的影響。a 圖為 control;b 圖為 DPN 處理;c 圖為 PPT 處理。黑色橫線(. )相當. 於 80μm,(□)涵蓋的區域為分析的位置 相當於 0.012mm2。 32.
(33) a. b. c. 圖十 去除卵巢的大白鼠接受 1 mg/Kg/day 的甲型(PPT)及乙型(DPN) 雌性素受體致效劑後下視丘弓形核神經胜? Y 的影響。a 圖為 control;b 圖為 DPN 處理;c 圖為 PPT 處理。黑色橫線(. )相當. 於 80μm,(□)涵蓋的區域為分析的位置 相當於 0.012mm2。 33.
(34) NPY level (% area). a 50 45 40 35 30 25 20 15 10 5 0. ARC. CON. DPN. PPT. DPN. PPT. b. NPY level (% area). 35. PVN. 30 25 20 15 10 5 0 CON. 圖十一 給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對去除卵 巢的大白鼠下視丘弓形核及室旁核神經胜? Y 神經元的表現 情形( n = 8-10)。 各組以 mean ±SEM 表示。a 圖為弓形核 (ARC);b 圖為室旁核(PVN)神經胜? Y 的表現量,各組間 並無統計上之差異。(CON:對照組,DPN: 乙型雌性素受體致 效劑,PPT: 甲型雌性素受體致效劑) 34.
(35) a. b. c. c. 圖十二. 去除卵巢的大白鼠接受 1 mg/Kg/day 的甲型(PPT)及乙型(DPN)雌 性素受體致效劑後下視丘弓形核甲型促黑激素的影響。a 圖為 control;b 圖為 DPN 處理;c 圖為 PPT 處理。黑色橫線(. )當於. 80μm,(□)涵蓋的區域為分析的位置 相當於 0.012mm2。 35.
(36) a α-MSH細胞數/section. 50 40 30 20 10 0 CON. DPN. PPT. CON. DPN. PPT. α-MSH level (% area). b 18 16 14 12 10 8 6 4 2 0. 圖十三 給予 1 mg/Kg/day 的甲型及乙型雌性素受體致效劑對去除卵 巢的大白鼠下視丘弓形核甲型促黑激素的表現 ( n =8-10)。 各組以 mean ±SEM 表示。a 圖為甲型促黑激素神經元的表現 數量;b 圖為甲型促黑激素神經纖維分佈情形,各組間並無 統計上之差異。(CON:對照組,DPN: 乙型雌性素受體致效 劑,PPT: 甲型雌性素受體致效劑)。 36.
(37) 討論 a. 報告指出甲型雌性素受體廣泛的表現在子宮、陰道和腦中的下視 丘弓形核等處;而乙型雌性素受體則主要表現在肺臟、卵巢以及前列 腺和下視丘室旁核;另外骨骼及腦下垂體則會同時表現兩種受體 (Gustafsson, 1999)。由此可知,只有甲型雌性素受體會在子宮表現, 實驗中,為了證實藥物的確與特異的受體結合,我們亦觀察大白鼠子 c 宮重量的變化。結果顯示,在給予 PPT 的大白鼠子宮重顯著高於對 照組及給予 DPN 的組別,由此知 PPT 的確是專一的作用在甲型雌性 素受體,而 DPN 給藥組並沒有子宮增重的情形,因子我們也可推知, DPN 是具專一性的乙型雌性素受體致效劑。. 在Geary等(2001)的研究指出,雌性素可以調控雌性動物各生 理週期中的攝食行為變化。Liang等人 (2002) 的報告也顯示在切除卵 巢後的大白鼠有增加攝食、體重和腹部脂肪累積的情況產生。而這個 不良影響可以在外給雌二醇的治療後回復( Bonavera et al., 1994) 。由 此可知雌性素在動物的攝食及能量平衡上扮演了重要的角色。為了探 討雌性素影響攝食的作用是經由甲型或是乙型雌性素受體而達成 的,我們分別給予切除卵巢的大白鼠甲型雌性素受體致效劑PPT或是 37.
(38) 乙型雌性素受體致效劑DPN,以觀察雌性素對攝食及體重的影響;結 果發現在連續給予PPT六天後,不論是0.5 mg/Kg或是1 mg/Kg,皆可 顯著的減少動物的攝食及體重。在2001年 Geary等的研究指出,缺乏 甲型雌性素受體的動物,其雌性素原本對攝食及體重的調控作用便消 失(Geary et al., 2001):因此我們可以推測,雌性素對大白鼠攝食及 體重的調控,是需要經過甲型雌性素受體的作用。另外,實驗中我們 發現在給予大白鼠0.5 mg/Kg的乙型雌性素受體致效劑DPN後,可以 看到大白鼠有顯著減少攝食,但體重卻沒有明顯的改變;而1 mg/Kg 的DPN也有相同的結果。在Liang Y-Q (2002)的報告指出,乙型雌性素 受體與雌性素所調控的厭食行為是有相關性的,因此,可能雌性素可 以部分藉由活化乙型雌性素受體來達到減少攝食的作用,但這個下降 攝食的結果,並不會伴隨著體重的減少;亦有報告指出雌性素對體重 的調控作用,主要是經由甲型雌性素受體而非乙型雌性素受體的作用 (Roesch., 2004);因此甲型雌性素受體在調節體重上較乙型受體為 重要。 我們分別將動物體重的改變量(g)除以進食量(g),以推算動物的 飼料效能,也就是計算每吃進一克飼料所增加的體重,我們把飼料效 能的結果當作一個代謝率及能量轉換率的指標,當飼料效能越高時, 表示其代謝率越低或者有較高之能量轉換率,反之亦然 ;文獻指出雌 38.
(39) 性素可以直接影響動物的身體代謝作用( Wallen et al., 2001) ;也有報 告認為雌性素可以藉著增加肝臟胰島素的清除速率來增加碳水化合 物的代謝率(Cagnacci et al.,1992);另外,在雌性素處理後可以看到 動物有減少蛋白與脂肪的儲存,以及加速能量支出的情況(Dagnault et al., 1994)。我們的結果顯示給予 PPT 後顯著的下降飼料效能,在給 予 PPT 後因提高了身體的代謝作用,使得吃進去的能量轉換成熱能 的形式而非儲存起來,這和給藥之後我們看到體重下降的結果符合, 也有研究指出甲型雌性素受體可以增加身體的基礎代謝率(Heine et al.,),因此可以知道,雌性素可藉由活化甲型受體來達到減少攝食及 提升身體代謝率,以及下降體重的作用;另外在給予 DPN 後飼料效 能間雖然比對照組低但無顯著差異,所以 DPN 經由活化乙型雌性素 受體後,雖然可以顯著的下降攝食,但可能相對的減低了身體的代謝 率或提高能量轉換率,而使得體重沒有改變。 性賀爾蒙可以影響部分由中樞所調節的攝食行為(Wallen et al., 2001)。中樞神經系統中有許多與攝食相關的神經元,在進食的過程 可以被活化(Eckel et al.,2001),其中神經胜? Y 是一個促進攝食的 胜? 類。在 2002 年 Baskin 等的文章指出,切除卵巢可顯著增加大白 鼠下視丘弓形核神經胜? Y 的傳訊者核糖核酸( mRNA)的表現量, 且在外給雌二醇後,可將此上升的情形回復(Baskin et al., 1995)。另外 39.
(40) 也有報告指出雌性素的厭食作用可能是經由抑制下視丘室旁核神經 胜? Y 的表現與釋放而達成的(Bonavera et al., 1994)。因此我們假 設雌性素甲型或乙型致效劑可會經由減少下視丘神經胜? Y 的表現 而產生的厭食作用。. 但是在本實驗中,在給了兩種雌性素受體致效劑後,我們以免疫 組織染色的方式,並沒有看到下視丘弓形核或是室旁核的神經胜? Y 神經纖維表現增加,有可能是神經胜? Y 的變化量未達到免疫染色 能偵測的靈敏度範圍內,我們必須進一步用西方點墨法或測量傳訊者 核糖核酸,才能作最後結論。. 甲型促黑激素也是位在下視丘一個調控攝食作用的重要胜? ,其 它主要的作用在抑制攝食。報告指出雌性素可以下降下視丘甲型促黑 激素的前驅物(POMC)的傳訊者核糖核酸(mRNA)的表現量(Wilcox & Roberts, 1985),而切除卵巢的大鼠 POMC 的傳訊者核糖核酸的表 現會顯著的上升,且這個增加的情形可以被外給雌二醇後回復 (Pelletier, 1993)。這些結果顯然無法支持動情素可經由提升甲型促 黑激素的作用而達到抑制攝食的目的。在本實驗中不論給予甲型或乙 型的雌性素荷爾蒙致效劑後,皆無法改變甲型促黑激素的表現,因此 40.
(41) 初步推測,雌性素對攝食的影響可能並非經由調節甲型促黑激素而達 成的。推測內生性的雌性素對攝食及體重的調控作用,可能是經由其 他的機轉而達成的。近來有報告指出,雌性素可以藉由活化升糖素及 膽囊收縮素等飽足的訊號,來達到下降攝食的作用(Eckel et al., 2002; Geary et al., 2001)。也有報告認為雌性素減少脂肪的作用,可能藉由 活化交感神經系統而達成的(Lazzarini et al., 1991) 。 且有報告指出, 下視丘室旁核的損傷並不會阻斷雌性素對能量平衡的作用(Dagnault & Richard., 1994)。因此雌性素是否可直接影響到下視丘相關神經元 的活性,進而調控能量平衡,需更深入的研究才能得到答案。. 總結以上實驗,我們可以知道雌性素可以藉由活化甲型及乙型雌 性素受體來達到下降攝食,且可經由活化甲型雌性素受體,而使體重 下降,另外也會增加身體的代謝率及降低能量的轉換率,目前結果顯 示雌性素並不會經由任一種受體而影響下視丘神經胜? Y 或甲型促 黑激素的表現,但仍可能需更進一步的實驗,才可以下完整的定論。. 41.
(42) 伍、參考文獻. Alvaro JD., Tatro JB., Duman RS. Minireview : melanocortins and opiate addiction. Life Sci. 1997; 61: 1- 9.. Bai FL., Yamano M., Shiotani Y., Emson PC., Smith AD., Powell JF., Tahyama M. An arcuato-paraventricular and dorsomedial hypothalamic neuropeptide Y-containing system which lacks noradrenalin in the rat. Brain Res 1985; 331:172-5.. Baskin DG., Breininger JF., Schwartz MW. Leptin receptor mRNA identifies a subpopulation of neuropeptide Y neurons activated by fasting in rat hypothalamus. Diabetes. 1999; 48: 828-33.. Baskin DG., Norwood BJ., Schwartz MW., Koerker DJ. Estrogen inhibits the increase of hypothalamic neuropeptide Y messenerger ribonucleic acid expression induce by weight loss in ovariectomized. Endocrinology. 1995; 12:5547-54. 42.
(43) Billington CJ., Briggs JE., Grace M., Levine AS. Effects of intracerebrovennricular injection of neuropeptide Y on energy metabolism. Am J Physiol. 1991; 260:R321-7.. Billington CJ., Briggs JE., Harker S., Grace M., Levine AS., Neuropeptide Y in hypothalamic paraventricular nucleus : A center coordinating energy metabolism. Am J Physiol. 1994; 266 : R1765-70.. Bonavera JJ., Dube MG., Kalra PS., Kalra SP. Anorectic effects of estrogen may be mediated by decreased neuropeptide- Y release in the hypothalamic paraventricular nucleus. Endocrinology, 1994; 134: 2367-70.. Cagnacci A., Soldani R., Carriero PL., Paoletti AM., Fioretti P., Melis GB. Effects of low doses of transdermal 17 beta-estradiol on carbohydrate metabolism in postmenopausal women. J Clin Endocrinol. 1992; l74: 1396-400.. Chance WT., Balasubramaniam A., Dayal R. Hypothalamic concentration and release of neuropeptide Y into the microdialysates is reduced in anorectic tumor –bearing rats. Life Sci. 1994;. 43. 54: 1869-74..
(44) Clark JT., Kalra PS., Crowley WR., Kalra SP. Neuropeptide Y and human pancreatic polypeptide stimulate feeding behavior in rats. Endocrinology. 1984; 115: 427-9.. Clark JT., Kalra PS., Crowley WR., Kalra SP. Neuropeptide Y stimulates feeding, but inhibits sexual behavior in rats. Endocrinology 1985; 117: 2435-42.. Clark JT., Sahu A., Kalra PS., Balasubramaniam A., Kalra SP. Neuropeptide Y-induced feeding behavior in female rats: comparison with human YY. Regu Pept. 1987; 17: 31-9.. Dagnault A., Richard D. Lesions of hypothalamic paraventricular nuclei do not prevent the effect of estradiol on energy and fat balance. Am J Physiol Endocrinol Metab 1994; 267: E32-E38.. Dube MG., Kalra PS., Crowley WR., Kalra SP. Evidence of physiological role for neuropeptide Y in ventromedial hypothalamic lesion-induced hyperphagia. Brain Res. 1991; 690: R328-34.. 44.
(45) Eckel LA., Geary N. Estradiol treatment increases feeding-induced c-Fos expression in the brains of ovariectomized rats. Am J Physiol, 2001; 281: R738-46.. Eckel LA.,, Houpt TA., Geary N. Estradiol treatment increases CCK-induced c-Fos expression in the brains of ovariectomized rats. Am J Physiol. 2002; 283: R1378-85.. Egawa M., Yoshimatsu H., Bray G. Neuropeptide Y suppresses sympathetic activity to interscapular brown adipose tissue in rats. Am J Physiol. 1991; 260:R328-34.. Everitt BJ., Hakfelt T. The coexistence of neuropeptide Y with other peptides and amines in the central nervous system. In: Mutt V, Fiixe K, HBkfelt T, Lundberg J (eds) Neuropeptide Y. Raven Press, New York, 1989; pp 61-72.. Fan W., Boston BA., Kesterson RA., Hruby VJ., Cone RD. Role of melanocortinergic neurons in feeding and the Agouti obesity syndrome. Nature. 1997; 385:165-8. 45.
(46) Geary N. The Effect of Estrogen on Appetite. Medscape Women Healthy. 1998; 3:3.. Geary N. Estradiol, CCK and satiation. Peptides. 2001; 22: 1251-63.. Geary N., Asarian L. Estradiol increases glucagon's satiating potency in ovariectomized rats. Am J Physiol Regul Integr Comp Physiol, 2001; 281: R1290-94.. Geary N., Asarian L., Korach KS., Pfaff DW., Ogawa S. Deficits in E2-dependent control of feeding, weight gain, and cholecystokinin satiation in ER-alpha null mice. Endocrinology. 2001; 11: 4751-57.. Grill HJ., Ginsberg AB., Seeley RJ., Kaplan JM. Brainstem application of melanocortin receptor ligands produces long-lasting effects on feeding and body weight. Neurosci. 1998; 18: 10128-35.. Gustafsson JÅ. Estrogen receptor ß - a new dimension in estrogen mechanism of action" J Endocrinol 1999; 163: 379-83.. Heine P. A., Jensen D.R., Taylor J.A., Lubahn D.B., Eckel R.H., Cooke P.S., Estrogen receptor alpha (ERa) regulates the effect of estrogen. 46.
(47) on metabolic rate (MR).The Endocrine Society Research Summaries Book. 2001; P3-110, (Abstract).. Inui A. Transgenic approach to the study of body weight regulation. Pharmacol Rev., 2000; 52: 35-55.. Ishihara A., Tanaka T., Kanatani A., Fukami T., Ihara M., Fukuroda T. A potent neuropeptide Y antagonist, 1229U91, suppressed spontaneous food intake in Zucker fatty rats. Am J Physiol. 1998; 274:R1500-4.. Kalra SP., Bube MG., Pu S., Xu B., Horvath TH., Karlra SP. Interacting appetite-regulating pathways in the hypothalamic regulation of body weight. Endocr Rev. 1999; 20: 68-100.. Kanatani A., Ishihara A., Aahi S., Tanaka T., Ozaki S., Ihara M. A potent neuropeptide Y Y1 receptor antagonist, 1229U91 :blockade of neuropeptide Y-induced and physiological food intake. Endocrinology. 1996; 137: 3177-82.. Kask A., Rgo L., Phkla R., Wikber JES., Schioth HB. Selective. 47.
(48) antagonist for the melanocortin 4 receptor (HS014) increase food intake in free-feeding rats. Biochem Biophys Res Commun. 1998; 245; 90-3.. Koegler FH., Grove KL., Schiffmacher A., Smith MS., Cameron JL. Central melanocortin receptors mediate changes in food intake in the rhesus macaque. Endocrinology. 2001; 142: 2586-92.. Kraichely DM., Sun J., Katzenellenbogen JA., Katzenellenbogen BS. Conformational changes and coactivator recruitment by novel ligands for estrogen receptor-alpha and estrogen receptor-beta: correlations with biological character and distinct differences among SRC coactivator family members. Endocrinology 2000; 141:3534-45.. Kuiper GGJM., Enmare E., Markku PH., NilssonS., Gustafsson JA. Cloning of a novel estrogen receptor expressed in rat prostate and ovary. Biochemistry1996; 93: 5925-30.. Lazzarini SJ., Wade GN. Role of sympathetic nerves in effects of estradiol on rat white adipose tissue. Am J Physiol. 1991; 260: R47-R51. 48.
(49) Liang Y.Q., Akishita M., Kim S., Ako J., Hashimoto M., Iijima K., Ohike Y., Watanabe T., Sudoh N., Toba K., Yoshizumi M. and Ouchi Y. 1 Estrogen receptor ß is involved in the anorectic action of estrogen . Int J Obes. 2002; 26: 1103?-98.. Meyers MJ., Sun J., Carlson KE., Marriner JA., Katzenellbogen BS., Katzenellbogen JA. Estrogen receptor-ß potency-selective ligands: structure-activity relationship studies of diarylpropionitriles and their acetylene and polar analogues. J Med Chem. 2001; 44: 4230-51.. Michel M., Beck-Sickinger A., Cox H., Doods HN., Herzog H., Larhammar D., Quiroon R., Schwartz T. Westfall T. International of pharmacology recommendations for the nomenclature of neuropeptide Y, peptide YY, and pancreatic polypeptide receptors. Pharmacol Rev. 1998; 50: 143-9.. Moras D., Gronemeyer H. The nuclear receptor ligand-binding domain: structure and function. Curr Opin Cell Biol. 1998; 10: 384-91.. 49.
(50) Morrell JI., Krieger MS., Pfaff DW. Quantitative autoradiographic analysis of estradiol retention by cells in the preoptic area, hypothalamic and amygdala. Exp Brain kes. 1986; 62: 343-54 a.. Mountjoy KG., Mortrud MT., Low MJ., Simerly RB., Cone RD. Localization of the melanocortin-4 receptor (MC4-R) in neuroendocrine and autonomic control circuits in the brain. Mol. J Endocrinol. 1994; 8: 1298-308.. Paxinos G, Watson C. The rat brain in stereotaxic coordinates, 2nd Ed. Academic Press Inc; 1986.. Pelletier G. Regulation of proopiomelanocortin gene expression in rat brain and pituitary as studied by in situ hybridization. Ann N Y Acad Sci. 1993;. 680: 246-259.. Pettersson K., Gustafsson JÅ. Role of estrogen receptor beta in estrogen action. Ann Rev Physiol. 2001; 63: 165-92.. Polidori C., Ciccocioppo R., Regoli D., Massi M. Neuropeptide Y. 50.
(51) receptors mediating feeding in the rat : characterization with antagonists. Peptides. 2000; 21: 29-35.. Roesch D. Estrogen receptor alpha, not beta, is the primary mediator of the attenuating effects of estrogens on body weight gain. Annual Meeting of Society for the Study of Ingestive Behavior. 2004, (Abstract).. Sahu A., Crowley WR., Kalra SP. Hypothalamic neuropeptideY gene expression increases before the onset of the ovarian steroidinduced luteinizing hormone surge. Endocrinology. 1994; 134: 1018-22.. Sahu A., Kalra SP. Neuropeptidergic regulation of feeding behavior: neuropeptide Y. Trends Endocrinol Metab. 1993; 4: 217-24.. Sahu A., Phelos CP., White ID., Crowlev WR., Kalra SP., Kalra PS. Steroidai regulation of- hypothala& neuropeptide Y release and gene expression. Endocrinolog. 1992; 130: 3331-6.. Sar M., Sahu A., Crowley WR., Kalra SP. Localization of 51.
(52) neuropeptide Y (NPY) immunoreactivity in estradiol concentrating cells in the hypothalamus. Endocrinolog, 1990; 127: 2752-56.. Stauffer SR.,Coletta CJ., Tedeco R., Nishiguchi G., Carison K.,Sun J.,Katzenellebogen JA. Pyrazole ligands: structure-affinity/activity relationships and estrogen receptor-alpha-selective agonists. J Med Chem 2000; 43: 4934.. Wallen WJ., Michael P., Belange., Wittnich C. Sex hormones and the selective estrogen receptor modulator tamoxifen modulate weekly body weights and food intakes in adolescent and adult rats. J Nutr. 2001; 131: 2351-7.. Wilcox JN., Roberts JL. Estrogen decreases rat hypothalamic proopiomelanocortin messenger ribonucleic acid levels. Endocrinology. 1985; 117: 2392-96.. Zarjevski N., Cusin I., Rohner-Jeanrenaud F., Jeanrenaud B. Chronic intracerebroventricular neuropeptide Y administration to normal rats. 52.
(53) mimics hormonal and metabolic changes of obesity. Endocrinology. 1993 ; 133: 1753-8.. 53.
(54) 作 者 簡 歷. 姓名:葉玲瑜. 出生日:69.9.6. 出生地:台灣省台北縣. 學歷:私立中國文化大學畜產系. 私立中國醫藥大學基礎醫學研究所. 會議論文:. 54.
(55) 1. J.W. Lee, L.Y. Yeh, Y.S. Chen, C.S.J Yang, H-J. Wang.(2004) :Roles of nitric oxide and melanocortin pathways in the anorectic effect of exendin-4.生物醫學聯合學術年會(Abstract).. 2. M.J. Yeh, L.Y. Yeh, J.W. Lee, C.S.J Yang, H-J. Wang.(2004) :The. effect of adenosine receptor antagonists on feeding.生物醫學聯合學 術年會(Abstract).. 55.
(56)
數據

Outline
相關文件
The first row shows the eyespot with white inner ring, black middle ring, and yellow outer ring in Bicyclus anynana.. The second row provides the eyespot with black inner ring
In this paper, we evaluate whether adaptive penalty selection procedure proposed in Shen and Ye (2002) leads to a consistent model selector or just reduce the overfitting of
During the Sejong era of the fifteenth century in particular, Korean science and technology had reached its climax, thus its accomplishments could reveal themselves clearly in the
Robinson Crusoe is an Englishman from the 1) t_______ of York in the seventeenth century, the youngest son of a merchant of German origin. This trip is financially successful,
fostering independent application of reading strategies Strategy 7: Provide opportunities for students to track, reflect on, and share their learning progress (destination). •
Strategy 3: Offer descriptive feedback during the learning process (enabling strategy). Where the
Now, nearly all of the current flows through wire S since it has a much lower resistance than the light bulb. The light bulb does not glow because the current flowing through it
03/2011 receiving certificate of Honorary Chair Professor from National Taiwan University of Science & Technology... 05/2013 receiving certificate of Honorary Chair Professor
