A Xanthine-Based Epithelium-Dependent Airway Relaxant
KMUP-3
(7-[2-[4-(4-Nitrobenzene)piperazinyl]ethyl]-1,3-dimethylxanthine) Increases Respiratory Performance and
Protects against Tumor Necrosis Factor-
␣-Induced Tracheal
Contraction, Involving Nitric Oxide Release and Expression of
cGMP and Protein Kinase G
Rong-Jyh Lin, Bin-Nan Wu, Yi-Ching Lo, Li-Mei An, Zen-Kong Dai, Young-Tso Lin,
Chao-Shun Tang, and Ing-Jun Chen
Institute of Medicine (R.-J.L.), Department and Graduate Institute of Pharmacology (B.-N.W., Y.-C.L., L.-M.A., I.-J.C.), and Departments of Pediatrics (Z.-K.D.), Cardiovascular Surgery (Y.-T.L.), and Anesthesiology (C.-S.T.), College of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan
Received July 7, 2005; accepted October 14, 2005
ABSTRACT
KMUP-3 (7-[2-[4-(4-nitrobenzene)piperazinyl]ethyl]-1,3-dimethyl-xanthine) was investigated in guinea pig tracheal smooth muscle.
Intratracheal instillation of tumor necrosis factor (TNF)-␣ (0.01
mg/kg/300 l) induced bronchoconstriction, increases of lung
resistance, and decreases of dynamic lung compliance. Instillation of KMUP-3 (0.5–2.0 mg/kg) reversed this situation. In isolated
trachea precontracted with carbachol, KMUP-3 (10 –100
M)-caused relaxations were attenuated by epithelium removal and by
pretreatments with an inhibitor of K⫹channel,
tetraethylammo-nium (10 mM); KATP channel, glibenclamide (1M);
voltage-de-pendent K⫹channel, 4-aminopyridine (100M); Ca2⫹-dependent
K⫹ channel, charybdotoxin (0.1M) or apamin (1 M); soluble
guanylate cyclase (sGC),
1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ, 1M); nitric-oxide (NO) synthase, N-nitro-L-arginine
methyl ester (L-NAME, 100M); and adenylate cyclase, SQ 22536
[9-(terahydro-2-furanyl)-9H-purin-6-amine] (100 M). KMUP-3
(0.01–100M) induced increases of cGMP and cAMP in primary
culture of tracheal smooth muscle cells (TSMCs). The increase in
cGMP by KMUP-3 was reduced by ODQ andL-NAME; the
in-crease in cAMP was reduced by SQ 22536. Western blot analysis
indicated that KMUP-3 (1 M) induced expression of protein
kinase A (PKA)RIand protein kinase G (PKG)1␣1in TSMCs. SQ
22536 inhibited KMUP-3-induced expression of PKARI. On the
contrary, ODQ inhibited KMUP-3-induced expression of PKG1␣1.
In epithelium-intact trachea, KMUP-3 increased the NO release. Activation of sGC, NO release, and inhibition of phosphodiester-ases in TSMCs by KMUP-3 may result in increphosphodiester-ases of intracellular cGMP and cAMP, which subsequently activate PKG and PKA,
efflux of K⫹ion, and associated reduction in Ca2⫹influx in vitro,
indicating the action mechanism to protect against TNF-
␣-in-duced airway dysfunction in vivo.
Xanthine derivatives, including theophylline, have long been recognized as bronchodilators because of their
nonspe-cific inhibition activities of phosphodiesterase (PDE) to in-crease cAMP, and they have been widely used in the therapy of bronchospastic diseases, including acute asthmatic at-tacks, emphysema, and chronic bronchitis, which are often associated with immunoresponses (Caramori and Adcock, 2003). The immunopharmacologic effects of xanthine deriv-atives have recently become a particular subject of interest (Kobayashi et al., 2004). Nevertheless, several important This work was supported by Grants NSC-89-2320-B-037-056-M59 (to
I.-J.C.) from the National Science Council, Taiwan and Vital Pharm Co., Ltd. (Kaohsiung, Taiwan).
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
doi:10.1124/jpet.105.092171.
ABBREVIATIONS: PDE, phosphodiesterase; PKG, protein kinase G; TNF, tumor necrosis factor; TSMC, tracheal smooth muscle cell; KMUP-1,
7-[2-[4-(2-chlorophenyl)piperazinyl] ethyl]-1,3-dimethyl xanthine; TSM, tracheal smooth muscle; KMUP-3, 7-[2-[4-(4-nitrobenzene)piperazinyl]-ethyl]-1,3-dimethylxanthine; NO, nitric oxide; sGC, soluble guanylyl cyclase; PKA, protein kinase A; IBMX, 3-isobutyl-1-methylxanthine; TEA, tetraethylammonium; 4-AP, 4-aminopyridine; ChTX, charybdotoxin; ODQ, 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one; NOS, nitric-oxide syn-thase;L-NAME, N-nitro-L-arginine methyl ester; AC, adenylate cyclase; SQ 22536, 9-(terahydro-2-furanyl)-9H-purin-6-amine; TBS, Tris-buffered saline; ANOVA, analysis of variance; 8-Br-cGMP; 8-bromo-cGMP.
THEJOURNAL OFPHARMACOLOGY ANDEXPERIMENTALTHERAPEUTICS Vol. 316, No. 2 Copyright © 2006 by The American Society for Pharmacology and Experimental Therapeutics 92171/3072683
JPET 316:709–717, 2006 Printed in U.S.A.
709
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
drawbacks remain, and a low therapeutic margin of safety, associated with a pharmacokinetic profile highly influenced by individual factors, results in the need for monitoring blood levels, which makes the drug hard to use widely in clinics (Barnes, 2003).
Likewise, the properties of xanthine derivatives to increase epithelium-dependent cGMP and protein kinase G (PKG)
expression, which suppress tumor necrosis factor-␣
(TNF-␣)-induced response in tracheal smooth muscle cells (TSMCs),
have not been explored thoroughly. TNF-␣ has been
de-scribed to up-regulate 5-hydroxytryptamine2A-mediated and
B1/B2 receptor responses in airway inflammation (Adner et al., 2002; Zhang et al., 2005), to modulate tracheal respon-siveness to G-protein-coupled receptor agonist (Chen et al., 2003), and to act as an inflammatory cytokine involved in asthmatics (Leung et al., 2005). Previously, KMUP-1 has been stated to possess airway relaxation activities (Wu et al., 2004). To create a more hopeful xanthine-based tracheal smooth muscle (TSM) relaxant, KMUP-3 (Fig. 1) was synthe-sized (Wu et al., 2005) and investigated regarding its mech-anism of TSM-relaxing activity and the benefits of its intra-tracheal administration. In routine clinical practice, in
addition to corticosteroids and2-adrenoreceptor agonist
in-halants, however, no xanthine derivatives are inhaled for the treatment of bronchocontraction or airway inflammation.
It has been reported that epithelial cells release various smooth muscle inhibitory mediators, such as nitric oxide
(NO) and prostaglandin E2, as well as so-called
epithelium-derived relaxing factor detected by the coaxial bioassay sys-tem (Nijkamp et al., 1993; Folkerts and Nijkamp, 1998). Some reports regarding xanthine derivatives on tracheal ep-ithelium, modulated by NO, have been published (Kanoh et al., 2000; Vajner et al., 2002). In the present study, we char-acterize the effects of KMUP-3 on TSM, associated specific epithelium-dependent action, soluble guanylate cyclase (sGC) activation, PDE inhibition, and reciprocation of cAMP-and cGMP-dependent protein kinase pathways, cAMP-and opening
of K⫹-channels. The combination of epithelium-derived
ac-tion (including NO release), PDE inhibiac-tion, sGC
stimula-tion, and K⫹-channel opening activity is suggested to
en-hance the results achieved. Additive effects of those pathways result in increase of cGMP/cAMP, coexpression of PKG/protein kinase A (PKA), and protection activity against
TNF-␣-induced bronchoconstriction.
Materials and Methods
Animals. Male Dunkin Hartley guinea pigs (350 – 450 g) were
provided by the National Laboratory Animal Breeding and Research Center (Taipei, Taiwan) and housed under conditions of constant temperature and controlled illumination. Food and water were avail-able ad libitum. The study was approved by the Animal Care and Use Committee of the Kaohsiung Medical University.
Pulmonary Function. Guinea pigs (350 – 450 g) were
anesthe-tized with pentobarbital (40 mg/kg i.p.) and maintained with further i.v. doses of 2 to 5 mg/kg when required according to methods pre-viously described (Wu et al., 2004). The animal was tethered in a supine position, and the trachea was cannulated below the larynx with a short tracheal cannula via a tracheotomy. Tracheal pressure was measured by a catheter connected to the side arm of the tracheal cannula. The carotid artery and jugular vein were cannulated to monitor blood pressure and heart rate and for drug administration, respectively. Throughout the experiment, the body temperature of the guinea pig was maintained at 37°C. Measurements of pulmonary cardiac function were carried out as previously described (Hsu et al., 1998). Total lung resistance (RL) and dynamic lung compliance (Cdyn) were measured on a breath-by-breath basis using a computer equipped with an A/D interface (DAS 1600; Buxco Electronics, Inc., Wilmington, NC) and software (version 1.5.7; Buxco Electronics, Inc.). Results obtained from the computer analysis were checked for accuracy by comparison with those calculated manually.
Animals were allowed to stabilize for 10 min following surgical manipulations before the test agent was administered. Intratracheal instillation was performed using a catheter placed through the tra-cheal tube into the bronchial system. The catheter connected to a 1-ml syringe was inserted into the trachea cannula extending 1 cm beyond the tip of the cannula for instillation of drug or vehicle. The instillation was followed by rapid intratracheal injection of 0.5 ml of air to facilitate deposition of drug or vehicle into the lungs as in the modified methods of Chung et al. (2001). The animals were pre-treated intratracheally with KMUP-3 (0.5, 1.0, and 2.0 mg/kg/150l of saline) or an equivalent volume (150l) of vehicle 15 min before TNF-␣ (0.01 mg/kg/300 l saline) was instilled to induce acute bron-choconstriction. In another experiments, KMUP-3 or KMUP-1 (1.0 mg/kg i.v.) were used to compare the effects on respiratory perfor-mance, indicated by changes of tidal volume, total RL, and Cdyn.
Isolated Tracheal Preparation and Measurement of Ten-sion. Guinea pigs (350 – 450 g) were anesthetized with pentobarbital
(70 mg/kg) i.p. and killed by a sharp blow on the neck, followed by cervical dislocation. The trachea was excised, cleaned of adhering fat and connective tissue, cut transversely into four to five rings, and then opened by cutting longitudinally through the cartilage rings diametrically opposite the tracheal smooth muscle (Hwang et al., 1999). Then trachea rings were suspended in a 10-ml organ bath containing Krebs’ solution (118 mM NaCl, 25 mM NaHCO3, 4.7 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 11 mM glucose, and 2.5 mM CaCl2, pH 7.3–7.4) maintained at 37°C and aerated with 95% O2and 5% CO2. Each tracheal tissue sample was subjected to 2 g of initial basal tension. All experiments were carried out in the presence of indomethacin (3M) and propranolol (1 M) to prevent the forma-tion of prostanoids and to inhibit-adrenergic responses, respec-tively. Isometric tension was recorded with a force displacement transducer (model FT03; Grass Instruments, Quincy, MA). Before the start of measurements, all preparations were allowed to equili-brate with frequent washing for 1 h. They were first contracted with carbachol (1M) to determine the contractility of preparations. This was also done at the end of each experiment. The preparations were then washed and allowed to equilibrate with Krebs’ solution for 50 min before being contracted a second time with carbachol (1M). When stable constriction to carbachol was reached, concentration responses of KMUP-3 (10, 30, and 100M) were obtained. Data were expressed as percentages of the maximum contractile response to carbachol (1 M). In comparison with KMUP-3, theophylline, Fig. 1. Chemical structures of KMUP-3 and KMUP-1.
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
3-isobutyl-1-methylxanthine (IBMX), milrinone, rolipram, zapri-nast, and levcromakalim, respectively, were examined at 100M.
In experiments to examine the role of epithelium in tracheal relaxation, the epithelial cells were removed mechanically by rub-bing the internal surface of trachea with a fine silver wire, and the removal of the epithelial layer was confirmed by histological exam-ination as previously described (Wu et al., 2004).
To examine the possible mechanism of tracheal relaxant effects of KMUP-3, the tracheal strips were pretreated with a K⫹ chan-nel blocker, tetraethylammonium (TEA; 10 mM); a KATPchannel blocker, glibenclamide (1 M); voltage-dependent K⫹ channel blocker, 4-aminopyridine (4-AP, 100M); Ca2⫹-dependent K⫹ chan-nel blockers, apamin (1M) and charybdotoxin (ChTX; 0.1 M); a sGC inhibitor, 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one (ODQ; 1 M); an NOS inhibitor, N-nitro-L-arginine methyl ester (L-NAME; 100M); and an adenylate cyclase (AC) inhibitor, SQ 22536 (100 M) for 30 min prior to the addition of KMUP-3 (10, 30, and 100 M). In the other experiments, TSMs were precontracted with 60 mM KCl to examine the relaxation achieved by KMUP-3. The KCl solution was prepared by substituting NaCl with KCl (60 mM) in an equimo-lar amount.
Determination of Nitrite. To measure concentrations of nitrite
(NO2⫺) in guinea pig TSM strips, TSM strips were pretreated with KMUP-3 (100M) in Krebs’ solution for 30 min. Then guinea pig TSM strips (2 ml/g tissue of methanol) were ultrasonically homoge-nized in an ice bath and then centrifuged at 10,000g for 10 min at 4°C. Briefly, harvestable supernatants were reacted with an equal volume of Griess reagents (1% sulfonamide/0.1% N-L -naphthylethyl-enediamine dihydrochloride/5% H3PO4) for 10 min to form a colored azo dye. The absorbance at 540 nm was detected by a flow-through visible spectrophotometer, and an equal volume of methanol was used for blank control.
Cell Culture of Tracheal Smooth Muscle. Guinea pigs (350 –
450 g) were injected i.p. with a lethal dose of pentobarbital. The tracheas were excised and cut longitudinally through the cartilage. Using a dissecting microscope, TSM strips were dissected from the surrounding parenchyma. The epithelium was removed from the luminal surface, and bands of TSM were gently separated from the underlying connective tissue. Then TSM strips were chopped into small sections (1 mm3) and incubated in Hanks’ balanced salt solu-tion (138 mM NaCl, 4 mM NaHCO3, 5 mM KCl, 0.3 mM KH2PO4, 0.3 mM Na2HPO4, and 1.0 mM glucose) with 0.05% elastase type IV and 0.2% collagenase type IV (Invitrogen, Carlsbad, CA) for 30 min at 37°C with gentle shaking. The solution of dissociated smooth muscle cells was centrifuged (6 min at 500g), and the pellet was resuspended in 1:1 Dulbecco’s modified Eagle’s medium/Ham’s F-12 medium sup-plemented with 10% fetal bovine serum, 0.244% NaHCO3, and 1% penicillin/streptomycin. Cells were cultured in 25-cm2flasks at 37°C in humidified air containing 5% CO2. Confluent cells were detached with 0.25% trypsin-0.02% EDTA at 37°C and then subcultured to establish secondary cultures. Cultures were maintained for no more than four passages. They were identified as smooth muscle cells by the typical hill-and-valley appearance, and cellular homogeneity was further confirmed by the presence of smooth muscle specific ␣-myo-sin and ␣-actin immunoreactivity. Indirect immunofluorescence staining for a variety of antigens was carried out by first plating the cells on chamber slides, fixing the cells in 3.7% formaldehyde-phos-phate-buffered saline for 10 min, and permeabilizing the cells with phosphate-buffered saline and 0.1% Triton X-100. Cells were then stained with either a mouse monoclonal antibody directed against the amino-terminal 10 amino acids of ␣-smooth muscle actin or ␣-myosin (Roche Diagnostics, Indianapolis, IN). All cells were stained with fluorescein-labeled goat anti-mouse IgG antibody (Roche Diagnostics). Over 95% of the cell preparation was found to be composed of smooth muscle cells.
Phosphodiesterase Activity. PDE activity was determined by
the method of Nicholson et al. (1991). PDE3 and PDE5 purified from human platelets and PDE4 purified from human U-937 pronocytic
cells were used. Test compound (10M) or vehicle was incubated with 0.2g of enzyme and 1 M cAMP containing 0.01 M [3H]cAMP in Tris buffer, pH 7.5, for 20 min at 25°C. The reaction was termi-nated by boiling for 2 min, and the resulting cAMP was converted to adenosine by the addition of 10 mg/ml snake venom nucleotidase, which is followed by a further incubation at 37°C for 10 min. Unhy-drolyzed cAMP was bound to AG1-X2 resin, and the remaining [3H]adenosine in the aqueous phase was quantitated by scintillation counting.
Another test compound (10M) or vehicle was incubated with 3.5 g of enzyme and 1 M cGMP containing 0.01 M [3H]cGMP in Tris buffer, pH 7.5, for 20 min at 25°C. The reaction was terminated by boiling for 2 min, and the resulting GMP was converted to adenosine by the addition of 10 mg/ml snake venom nucleotidase, which was followed by a further incubation at 37°C for 10 min. Unhydrolyzed cGMP was bound to AG1-X2 resin, and the remaining [3H]guanosine in the aqueous phase was quantitated by scintillation counting.
Measurement of cAMP and cGMP. Intracellular
concentra-tions of cAMP and cGMP in TSMCs were measured as in our previ-ous reports (Wu et al., 2001; Lin et al., 2002). In brief, cells were finally grown in 24-well plates (105cells/well). At confluence, mono-layer cells were washed with phosphate-buffered saline and then incubated with KMUP-3 (0.01–100M) for 20 min at 37°C in me-dium. Milrinone, rolipram, zaprinast, theophylline, or aminophylline (100M) was added for 20 min. The reaction was terminated by replacement of medium with 1 ml of ice-cold 1 N hydrochloric acid. Cell suspensions were sonicated and then centrifuged at 2500g for 15 min at 4°C. Then, the supernatants were lyophilized, and the cAMP and cGMP of each sample were determined using commercially available radioimmunoassay kits (GE Healthcare, Little Chalfont, Buckinghamshire, UK).
Expression of PKA and PKG. The expression of PKA and PKG
was determined by Western blot as previously described by Wu et al. (2004) and Murthy et al. (2002). Guinea pig TSMCs were incubated with KMUP-3 for 30 min. Each of the cell lysates, each containing 1 g of cellular protein, was electrophoresed in 7.5% SDS-polyacryl-amide gel electrophoresis and then transferred to polyvinylidene diflouride membrane (Millipore Corporation, Billerica, MA). The membrane was stained with Ponceau S to verify the integrity of the transferred proteins and to monitor the unbiased transfer of all protein samples. The membrane was then washed with 25 ml of TBS (100 mM NaCl, 0.1% and 10 mM Tris-HCl, pH 7.5) for 5 min at room temperature and incubated in 25 ml of blocking buffer (TBS plus 0.1% Tween 20 and 5% nonfat milk) overnight at 4°C. In the mea-surement of protein, the membrane was incubated with appropriate PKARIor PKG1␣1primary antibody (diluted 1:250) in 2 ml of block-ing buffer for 1 h at room temperature, washed three times for 5 min each with 15 ml of Tris-buffered saline/Tween 20 (TBS plus 0.1% Tween 20), incubated with horseradish peroxidase-conjugated sec-ondary antibody (1:10,000) in 15 ml of blocking buffer with gentle agitation for 1 h at room temperature, and finally washed three times for 5 min each time with 15 ml of Tris-buffered saline/Tween 20. The membrane was then subjected to enhanced chemilumines-cence (GE Healthcare) for the detection of the specific antigen.
Drugs and Chemicals. Levcromakalim was generously supplied
by the GlaxoSmithKline (Uxbridge, Middlesex, UK) and dissolved in ethanol at 10 mM. 4-AP, aminophylline, apamin, carbachol, ChTX, forskolin, glibenclamide, Griess reagents, IBMX, indomethacin, Krebs’ solution reagents,L-NAME, methylene blue, ODQ, propran-olol, SQ 22536, TEA, theophylline, and TNF-␣ were all obtained from Sigma-Aldrich (St. Louis, MO). PKARIand PKG1␣1antibodies were purchased from Calbiochem (San Diego, CA). All drugs and reagents were dissolved in distilled water unless otherwise noted. Apamin was dissolved in 0.05 M acetic acid; indomethacin was dissolved in 100 mM sodium carbonate; ChTX, glibenclamide, milrinone, rolip-ram, zaprinast, IBMX, and ODQ were dissolved in DMSO at 10 mM and serially diluted with distilled water; and KMUP-3, synthesized in our laboratory, was dissolved in 10% absolute alcohol, 10%
pro-KMUP-3 and TNF-␣ Bronchocontraction
711
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
pylene glycol, and 2% 1 N HCl at 10 mM. Serial dilutions were made in distilled water.
Statistical Evaluation of Data. The results are expressed as
mean⫾ S.E. Statistical differences were determined by independent and paired Student’s t test in unpaired and paired samples, respec-tively. Whenever a control group was compared with more than one treated group, the one-way ANOVA or two-way repeated measures ANOVA was used. When the ANOVA manifested a statistical differ-ence, the Dunnett’s or Student-Newman-Keuls test was applied. P⬍ 0.05 was considered to be significant in all experiments. Analysis of the data and plotting of the figures were performed with the aid of software (SigmaPlot version 8.0 and SigmaStat Version 2.03, Chi-cago, IL) run on an IBM-compatible computer.
Results
Pulmonary Function. The baseline values of total lung resistance, dynamic lung compliance, and tidal volume were
0.25⫾ 0.02 cmH2O ml⫺1s⫺1, 0.31⫾ 0.02 ml cmH2O⫺1, and
1.91⫾ 0.11 ml, respectively. TNF-␣-induced
bronchocontrac-tion by intratracheal instillabronchocontrac-tion increased RLand decreased
Cdyn, respectively (Fig. 2). KMUP-3 (0.5, 1.0, and 2.0 mg/kg)
instilled through trachea for 15 min prior to TNF-␣-induced
bronchocontraction could reverse the situation by decreasing
RLand by increasing Cdyn(Fig. 2). Blood pressure and heart
rate were not significantly changed by instillation of KMUP-3 during the recording of lung function. In another experiment, i.v. KMUP-3 (1 mg/kg) more significantly
de-creased RL and increased both Cdyn and respiratory tidal
volume than KMUP-1 (Table 1).
NOS and Epithelium-Derived Relaxation. As shown in
Fig. 3A, KMUP-3 (10, 30, and 100M) produced
concentra-tion-dependent relaxations both in epithelium-denuded and
epithelium-intact (EP⫹) guinea pig TSM, indicating that
KMUP-3 induced epithelium-independent relaxation activ-ity. However, KMUP-3 did demonstrate a significant de-crease in the relaxation response after epithelium denuda-tion, suggesting that at least part of the observed effect is epithelium-dependent. Obviously, KMUP-3 completely
re-laxed the TSM strips at 100M, achieving even up to 130%
relaxation. In addition, the relaxations of guinea pig EP⫹
TSM elicited by KMUP-3 were significantly inhibited by
pre-treatment with an NOS inhibitorL-NAME. The NO2⫺release
reached 160⫾ 15% (n ⫽ 3) by KMUP-3 (100M) in
compar-ison with the basal level (0.8⫾ 0.1M/mg) in EP⫹guinea pig
TSM.
KMUP-3, theophylline, IBMX, milrinone, rolipram, and
zaprinast (100 M) were all found to cause relaxations in
epithelium-intact trachea. Both IBMX and KMUP-3 were
more potent at 100 M concentrations than
theophylline-inducing tracheal relaxation (Fig. 3B). A potassium channel
opener, levcromakalim (100 M), also displayed relaxation
effects on TSM (EP⫹) (Fig. 3B).
Kⴙ Channel Activities. KMUP-3 (10, 30, and 100 M) caused a concentration-dependent relaxation in carbachol- or
high-K⫹-contracted guinea pig EP⫹TSM. KMUP-3 against
carbachol-induced contractions in TSM (EP⫹) was inhibited
by the K⫹channel blocker, TEA; a KATPchannel blocker,
glibenclamide; a voltage-dependent K⫹ channel blocker,
4-AP; and Ca2⫹-dependent K⫹channel blockers, apamin and
ChTX (Fig. 4).
Involvement of sGC and Adenylate Cyclase. The
re-laxations of guinea pig EP⫹TSM elicited by KMUP-3 were
attenuated by pretreatment with a sGC inhibitor, ODQ (1
M); an NOS inhibitor,L-NAME (100M); or an adenylate
cyclase inhibitor, SQ 22536 (100M) (Fig. 5). Additionally,
KMUP-3-induced relaxations were dramatically reduced by
the pretreatment with L-NAME ⫹ SQ 22536, ODQ ⫹ L
-NAME, or SQ 22536⫹ ODQ (Fig. 5).
Inhibition of Phosphodiesterase. The enzyme inhibi-tory activity of KMUP-3 on PDEs was measured and com-pared with KMUP-1. As shown in Table 1, KMUP-3 displays Fig. 2. RLand Cdyneffects of KMUP-3 on TNF-␣-induced (0.01 mg/kg)
bronchoconstriction by intratracheal instillation. Each value represents the mean⫾ S.E. from six independent experiments. ⴱ, P ⬍ 0.05 compared with the KMUP-3 solvent ⫹ TNF-␣ (ANOVA followed by Student-Newman-Keuls test).
TABLE 1
Effects of KMUP-3 and KMUP-1 on PDE inhibitory activity and lung resistance, dynamic compliance, and tidal volume maximum changes
The enzyme inhibitory activity was measured by 10M KMUP-1 and KMUP-3 (n ⫽ 3) on various PDEs. RL, Cdyn, and tidal volume were measured after i.v. injection of
KMUP-1 or KMUP-3 (1 mg/kg).
Compound PDE3 Inhibition PDE4 Inhibition PDE5 Inhibition RL Cdyn Tidal Volume
% maximum change of baseline (%)
KMUP-1 23⫾ 2.2 18⫾ 2.3 29⫾ 2.5 ⫺20 ⫾ 3.5 35⫾ 4.5 2.8⫾ 0.2
KMUP-3 55⫾ 2.5* 48⫾ 2.1* 48⫾ 1.9* ⫺55 ⫾ 4.5* 50⫾ 5.5* 21.7⫾ 0.5*
* P⬍ 0.05 compared with KMUP-1, respectively (n ⫽ 3).
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
different inhibition activities on PDE3, PDE4, and PDE5. The result indicated that the enzyme inhibition activity of KMUP-3 appears to be more potent than KMUP-1, but it has no selectivity among PDE3, 4, or 5. We also observed that the
inhibitory effects of theophylline (10M) on PDE3, 4, and 5
were 8⫾ 1.2, 7.9 ⫾ 1.2, and 12 ⫾ 1.5% (n ⫽ 3). The IC50
values of KMUP-3 for PDE3, PDE4, and PDE5 were 8.5⫾
1.5, 14.3 ⫾ 2.2, and 14.5 ⫾ 1.8 M (n ⫽ 3), respectively,
whereas KMUP-1’s IC50 values for those PDEs were⬎100
M. Under this condition, IBMX was used as a reference
agent, and its IC50values for PDE3, PDE4, and PDE5 were
6.4⫾ 1.6, 25.6 ⫾ 4.9, and 30.8 ⫾ 4.5M (n ⫽ 3), respectively.
Accumulation of cAMP and cGMP. cAMP and cGMP levels were examined in guinea pig primary TSMCs. The amounts of basal levels of cAMP and cGMP in the cells were
62.6⫾ 6.4 and 1.18 ⫾ 0.06 pmol/mg protein, respectively (n ⫽
3). KMUP-3 (0.01, 0.1, 1, 10, and 100M) significantly
in-creased both cAMP (80.2 ⫾ 2.5, 91.3 ⫾ 3.4, 97.9 ⫾ 2.5,
110.7 ⫾ 4.5, and 127.5 ⫾ 4.8 pmol/mg protein) and GMP
(1.62⫾ 0.05, 1.73 ⫾ 0.01, 1.75 ⫾ 0.04, 1.86 ⫾ 0.05, and 1.98 ⫾
0.09 pmol/mg protein) levels compared with each basal value in guinea pig primary TSMCs. Moreover, we compared both
cAMP and GMP levels of KMUP-3 with theophylline,
ami-nophylline, milrinone, rolipram, and zaprinast at 100 M
(Fig. 6). KMUP-3, theophylline, aminophylline, and zapri-nast significantly enhanced the amounts of cGMP, but this was not observed in milrinone or rolipram for selective PDE3 and PDE4 inhibitions, respectively. On the other hand, KMUP-3, theophylline, aminophylline, milrinone, and rolip-ram all elicited significant elevations of cAMP accumulation, but this was not so for the selective PDE5 inhibitor zaprinast (Fig. 6). The elevated cAMP levels induced by KMUP-3 and
forskolin (100M), an adenylate cyclase activator, were
in-hibited by pretreatment with the adenylate cyclase inhibitor,
SQ 22536 (100M) (Table 2). Correspondingly, the rise of
cGMP accumulation by KMUP-3 was abolished by
pretreat-ment with the NOS inhibitorL-NAME (100M) or the sGC
inhibitor ODQ (10M), respectively (Table 2).
Expression of PKA and PKG. Expression of
immunore-active PKARI is shown in Fig. 7A. Monoclonal antibody to
PKARIrecognized a band at 49 kDa in extracts of tracheal
smooth muscle. KMUP-3 (1M) stimulated the expression of
immunoreactive PKARI protein. Treatment with SQ 22536
(100 M) 30 min prior to KMUP-3 (1 M) attenuated the
expression of PKARI(Fig. 7A). Both 8-Br-cAMP and KMUP-3
(1M) increased the expression of PKARI; however,
pretreat-ment with ODQ (10M) 30 min prior to KMUP-3 did not
significantly affect the PKA protein expression by KMUP-3 (B. N. Wu and I. J. Chen, unpublished data). Expression of
immunoreactive PKG1␣1 is shown in Fig. 7B. Polyclonal
antibody to PKG1␣1recognized a band at 75 kDa in extracts
of tracheal smooth muscle. Both 8-Br-cGMP and KMUP-3 (1
M) increased the expression of PKG1␣1, and pretreatment
with ODQ (10 M) 30 min prior to KMUP-3 (1 M)
sig-nificantly reduced the PKG protein induced by KMUP-3 (Fig. 7B).
Discussion
Relaxation of smooth muscles are through activation of AC and sGC and accumulation of cAMP and cGMP (Ignarro and Kadowitz, 1985). cAMP and cGMP are degraded by PDE,
which catalyzes the conversion of cAMP to 5⬘-AMP and
cGMP to 5⬘-GMP and therefore leads to decreases in
intra-cellular cAMP or cGMP levels. Among them, PDE3 is inhib-ited by cGMP, indirectly leading to the increase of cAMP. Accumulations of cAMP and cGMP further induced the in-creases of PKG and PKA. In this study, KMUP-3 signifi-cantly increases the expression of PKA and PKG and thus confirms the contents of cAMP and GMP in TSMCs. The rank
order of relaxant activity at 100M was KMUP-3 ⬎ KMUP-1
(Wu et al., 2004)⬎ IBMX ⬎ milrinone ⬎ rolipram ⬎
zapri-nast, theophylline, and levcromakalim (Fig. 3B). This result is adaptable to enzyme inhibition activity and IC50 of KMUP-3 on PDE3, 4, and 5, in comparison with previous KMUP-1. We suggest that the higher inhibition (percentage) of KMUP-3 than KMUP-1 on PDE3, 4 and 5 favors better respiratory performance as shown by increased respiratory
tidal volume, decreased RL, and increased Cdyn(Table 1).
Inhibition of PDE3 with specific inhibitors has resulted in relaxation of TSM, either on spontaneous or carbachol-in-duced tone (Torphy and Cieslinski, 1990). In contrast, the PDE4 inhibitor rolipram did not reduce spontaneous tone but relaxed carbachol-induced tone. To date, some of the highly Fig. 3. A, effects of KMUP-3 on carbachol (1M)-precontracted guinea
pig trachea in the EP⫹and epithelium-denuded (Ep⫺) tissues.ⴱ, P ⬍ 0.05,
n⫽ 8, compared with the EP⫹(ANOVA followed by Tukey test). B, effects of KMUP-3, theophylline, IBMX, milrinone, rolipram, zaprinast, and levcromakalim (100M) on carbachol (1 M)-precontracted guinea pig epithelial-intact trachea.
KMUP-3 and TNF-␣ Bronchocontraction
713
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
selective PDE inhibitors have been disappointing. One rea-son for this may be the presence of significant amounts of other PDE isoenzymes, existing in target tissues (Giembycz et al., 1996). Combination inhibition on PDE3 and PDE4 thus may be more effective than either alone in relaxing TSM, suggesting that the additive effects of PDE3 and PDE4 inhi-bition are able to optimize the bronchodilatory benefits (Giembycz and Barnes, 1991; de Boer et al., 1992). KMUP-3 is found to have PDE3 and PDE4 inhibitions; therefore, we suggest that it can effectively relax TSM. KMUP-3 showed a stronger inhibition on PDE3 and PDE4 than KMUP-1. In-deed, inhibitors with PDE3 and PDE4 inhibition have at-tracted considerable interest as potential antiasthmatic agents because they possess both bronchodilatory and anti-inflammatory activities (Barnes, 1996). However, in addition
to PDE inhibition, previous epithelium-dependent cGMP en-hancing activity of KMUP-1 encourages us to further inves-tigate the benefits of KMUP-3, administered by intratracheal instillation, for the treatment of airway dysfunction caused by bronchocontraction.
In guinea pig trachea, the NO donor has been proven to increase cGMP, and it enhances the increase of cAMP in-duced by the PDE4 inhibitor rolipram (Turner et al., 1994). Moreover, the PDE5 inhibitors sildenafil and zaprinast, with their cGMP-increasing activities, have been proven to induce relaxation in TSM (Ellis and Conanan, 1995; Hirose et al., 2001). KMUP-1, with NO-independent sGC activation and PDE inhibition activities, also leads to cGMP-mediated inhi-bition of TSMs (Wu et al., 2004). In the present study, KMUP-3 is further suggested to have PDE inhibition and Fig. 4. Effects of KMUP-3 on carbachol (1 M)-pre-contracted guinea pig epithelial-intact trachea in the absence or presence of various potassium channel blockers.ⴱ, P ⬍ 0.05, n ⫽ 8, compared with the control (ANOVA followed by Tukey test).
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
action of epithelium-derived NO to increase cGMP and cAMP. KMUP-3 retains partial PDE4 inhibition activity, which increases cAMP, and subsequently the expression of PKA in TSMs. Inhibition of PDE5 activity by KMUP-3 may increase cGMP, which has proven to inhibit PDE3 activity, resulting increase of cAMP, and subsequently the expression of PKG and partial PKA (Fig. 8).
NO is capable of stimulating K⫹channels in smooth
mus-cle cells, such as those that are Ca2⫹-activated (Bolotina et
al., 1994; Koh et al., 1995), leading to hyperpolarization and consequently muscle relaxation (Lincoln and Cornwell, 1991; Robertson et al., 1993), including in airway (Abderrahmane
et al., 1998). K⫹channel openers, acting by liberation of NO,
have been shown to relax TSM and to produce bronchodila-tion. Based on these results, we sought to explore whether Fig. 5. Effects of KMUP-3 on carbachol (1M)-precontracted guinea pig
epithelial-intact trachea in the absence or presence ofL-NAME (100M), ODQ (1M), or SQ 22536 (100 M). ⴱ, P ⬍ 0.05, n ⫽ 8, compared with the control; #, P⬍ 0.05, n ⫽ 4, compared with theL-NAME, ODQ, or SQ 22536, respectively (ANOVA followed by Tukey test).
Fig. 6. Effects of KMUP-3, milrinone, rolipram, zaprinast, theophylline,
and aminophylline (100M) on cGMP and cAMP levels in guinea pig tracheal smooth muscle cells. Each value represents the mean⫾ S.E. from three independent experiments.ⴱ, P ⬍ 0.05 compared with the vehicle (ANOVA followed by Dunnett’s test).
TABLE 2
Effects of KMUP-3 and forskolin (100M) on cAMP and cGMP levels in TSMCs in the absence and presence of SQ 22536 (100M),L-NAME (100M), or ODQ (10 M)
Each value represents the mean⫾ S.E.M. of three independent experiments. Treatment
cAMP (pmol/mg protein)
Vehicle 62.60⫾ 6.40 SQ 22536 74.32⫾ 7.56 Forskolin 119.15⫾ 5.11* Forskolin plus SQ 22536 94.33⫾ 3.42** KMUP-3 128.72⫾ 4.57* KMUP-3 plus SQ 22536 74.51⫾ 3.18**
Cyclic GMP (pmol/mg protein)
Vehicle 1.18⫾ 0.06
KMUP-3 1.98⫾ 0.08*
L-NAME 1.12⫾ 0.02
KMUP-3 plusL-NAME 1.28⫾ 0.01**
ODQ 1.12⫾ 0.03
KMUP-3 plus ODQ 1.35⫾ 0.04**
* P⬍ 0.05 compared with the vehicle, ANOVA followed by Dunnett’s test. ** P⬍ 0.05 compared with forskolin or KMUP-3.
KMUP-3 and TNF-␣ Bronchocontraction
715
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
the opening of K⫹ channels and associated expression of cGMP-dependent protein kinase accounts for the relaxation caused by KMUP-3 in the guinea pig trachea. Evidence indi-cates that KMUP-3-induced relaxation was sensitive to
var-ious K⫹channel blockers. Therefore, it is likely that
KMUP-3-induced relaxation is partly mediated by NO-induced
effects, including stimulating K⫹efflux resulting in
hyperpo-larization (Wu et al., 2005) and thereafter accumulation of cGMP leading to the expression of PKG and reduction of
Ca2⫹influx. KMUP-3-induced TSM relaxation was
signifi-cantly reduced by pretreatment with ODQ. This fact suggests activation of sGC and stimulation of cGMP-dependent mech-anism could be involved in KMUP-3-induced relaxation. Ba-sically, the cAMP pathway was involved in KMUP-3-induced TSM relaxations, which were inhibited by a selective AC inhibitor SQ 22536 (Fig. 5). However, in addition to PDE4 inhibition by KMUP-3 to increase cAMP, we cannot neglect the contribution from cGMP-induced inhibition on PDE3.
Furthermore, SQ 22536 abolished the cAMP accumulation in response to both KMUP-3 and forskolin in TSMCs (Table 2). These facts indicate that KMUP-3-induced relaxation is par-tially mediated by activation of AC. PKG is the action target of cGMP (Komalavilas and Lincoln, 1994). It is reasonable that the expression of PKG accompanies the increased cGMP. Since KMUP-3 induced TSM relaxation and increased cGMP, the expression of PKG is thus realizable.
Pretreat-ment with ODQ abolished the KMUP-3-induced PKG1␣1
expression; moreover, the PKAR1 expression caused by
KMUP-3 also showed similar responses but was inhibited by the AC inhibitor SQ 22536 (Fig. 7A). Notably,
KMUP-3-induced TSM relaxation is associated with K⫹channel
open-ing activity (Somlyo and Somlyo, 1994), resultopen-ing in in-creased expression of PKA and PKG (Fig. 8).
TNF-␣ has been shown to induce hyper-responsiveness
and bronchocontraction of TSM (Wu et al., 2004). As a
member of cytokines, TNF-␣ attracts and activates
non-specific inflammatory macrophages and neutrophils dur-ing occurrence of type IV hypersensitivity, usually synthe-sized in response to infection, including that found in airway system. Since KMUP-3 has PDE3, PDE4, and PDE5 inhibition activities, we thus investigated the
ben-efits of KMUP-3 to protect against intratracheal TNF-
␣-induced bronchoconstriction. In the present study,
exoge-nous TNF-␣ certainly increased RL components and
decreased Cdyn factor. However, these situations are
sig-nificantly reversed by KMUP-3 (Fig. 2). These facts imply
that KMUP-3 has the potential to protect against TNF-
␣-induced tracheal overcontractility in the airway.
KMUP-3 is suggested to accumulate cAMP/cGMP and
en-hance K⫹efflux, leading to reduction of Ca2⫹
influx-associ-ated contractility in TSM (Fig. 8). Whether accumulation of
cAMP or cGMP caused by KMUP-3 is attributable to K⫹
efflux in TSMC remains to be resolved. In this study, the
expression of PKG1␣1and PKARI, able to be activated by
cGMP/cAMP, was suggested to mediate the K⫹channel
open-ing activity through their cGMP/cAMP-increasopen-ing activities. The combination of the multiple actions of KMUP-3 above thus may contribute to significant relaxation of carbachol-induced TSM contraction in vitro and protect against
TNF-␣-induced increases of RLand decreases of Cdynin vivo. It is
suggested that PDE inhibition, cAMP/cGMP increasing, and Fig. 8. Proposed mechanism of action of KMUP-3 on the intracellular
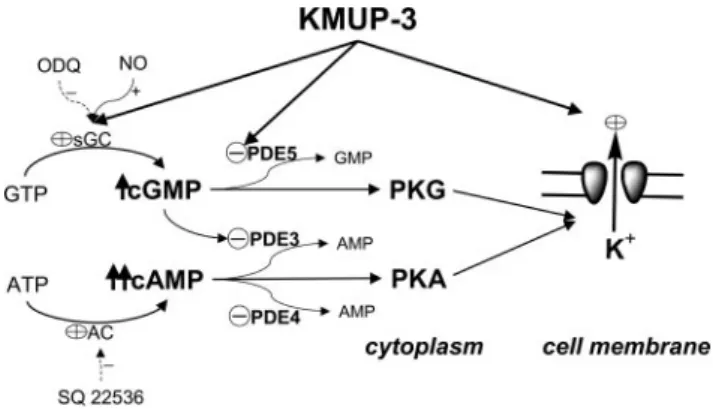
cGMP and cAMP synthesis and metabolism, K⫹channel opening, and phosphodiesterase inhibition. Increased cAMP and cGMP then activate the PKA and PKG cascades and thus enhance the K⫹efflux, leading to attenuation of Ca2⫹ influx-associated contractility in tracheal smooth muscle cells. Cycle symbols indicated as KMUP-3 effects.
Fig. 7. Representative Western blots and the corresponding group data
depicting PKARIand PKG1␣1protein expression in cultured guinea pig tracheal smooth muscle cells incubated for 30 min in the absence (vehicle) and presence of: A, 8-Br-cAMP (1M), SQ 22536 (100 M), SQ 22536 ⫹ KMUP-3 (1M), or KMUP-3 (1 M); or B, 8-Br-cGMP (1 M), ODQ (10 M), ODQ ⫹ KMUP-3, or KMUP-3. SQ 22536 or ODQ was treated 30 min prior to KMUP-3.ⴱ, P ⬍ 0.05 compared with the vehicle; #, P ⬍ 0.05 compared with the SQ 22536⫹ KMUP-3; ⫹, P ⬍ 0.05 compared with the ODQ⫹ KMUP-3 (n ⫽ 3) (ANOVA followed by Dunnett’s test).
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org
K⫹ channel activities of KMUP-3 are the crucial determi-nants for its relaxation effects on TSM. Particularly, more PDE4-inhibiting activity by KMUP-3 than by KMUP-1 may favor the former as useful in increasing respiratory perfor-mance. We also suggest that the in vivo activities of KMUP-3 in airway are consistent with the proposed mode of action derived from the in vitro experiments. Intratracheal instilla-tion with KMUP-3, leading to increase of respiratory
perfor-mance with protection activity against TNF-␣-induced
tra-cheal contractility, may indicate its considerable usefulness and may be the challenge for future inhalation research. References
Abderrahmane A, Salvail D, Dumoulin M, Garon J, Cadieux A, and Rousseau E (1998) Direct activation of K(Ca) channel in airway smooth muscle by nitric oxide: involvement of a nitrothiosylation mechanism? Am J Respir Cell Mol Biol 19:485– 497.
Adner M, Rose AC, Zhang Y, Sward K, Benson M, Uddman R, Shankley NP, and Cardell LO (2002) An assay to evaluate the long-term effects of inflammatory mediators on murine airway smooth muscle: evidence that TNFalpha up-regulates 5-HT(2A)-mediated contraction. Br J Pharmacol 137:971–982.
Barnes PJ (1996) New drugs for asthma. Clin Exp Allergy 26:738 –745.
Barnes PJ (2003) Theophylline: new perspectives for an old drug. Am J Respir Crit
Care Med 167:813– 818.
Bolotina VM, Najibi S, Palacino JJ, Pagano PJ, and Cohen RA (1994) Nitric oxide directly activates calcium-dependent potassium channels in vascular smooth mus-cle. Nature (Lond) 368:850 – 853.
Caramori G and Adcock I (2003) Pharmacology of airway inflammation in asthma and COPD. Pulm Pharmacol Ther 16:247–277.
Chen H, Tliba O, Van Besien CR, Panettieri RA Jr, and Amrani Y (2003) TNF-[alpha] modulates murine tracheal rings responsiveness to G-protein-coupled re-ceptor agonists and KCl. J Appl Physiol 95:864 – 872.
Chung WH, Bennett BM, Racz WJ, Brien JF, and Massey TE (2001) Induction of c-jun and TGF-beta 1 in Fischer 344 rats during amiodarone-induced pulmonary fibrosis. Am J Physiol 281:L1180 –L1188.
de Boer J, Philpott AJ, van Amsterdam RG, Shahid M, Zaagsma J, and Nicholson CD (1992) Human bronchial cyclic nucleotide phosphodiesterase isoenzymes: biochem-ical and pharmacologbiochem-ical analysis using selective inhibitors. Br J Pharmacol 106:1028 –1034.
Ellis JL and Conanan ND (1995) Modulation of relaxant responses evoked by a nitric oxide donor and by nonadrenergic, noncholinergic stimulation by isoenzyme selec-tive phosphodiesterase inhibitors in guinea pig trachea. J Pharmacol Exp Ther 272:997–1004.
Folkerts G and Nijkamp FP (1998) Airway epithelium: more than just a barrier!
Trends Pharmacol Sci 19:334 –341.
Giembycz MA and Barnes PJ (1991) Selective inhibition of a high affinity type IV cyclic AMP phosphodiesterase in bovine trachealis by AH 21–132: relevance to the spasmolytic and anti-spasmogenic actions of AH 21–132 in the intact tissue.
Biochem Pharmacol 42:663– 677.
Giembycz MA, Corrigan CJ, Seybold J, Newton R, and Barnes PJ (1996) Identifica-tion of cyclic AMP phosphodiesterases 3, 4 and 7 in human CD4⫹ and CD8⫹ T-lymphocytes: role in regulating proliferation and the biosynthesis of interleu-kin-2. Br J Pharmacol 118:1945–1958.
Hirose R, Okumura H, Yoshimatsu A, Irie J, Onoda Y, Nomoto Y, Takai H, Ohno T, and Ichimura M (2001) KF31327, a new potent and selective inhibitor of cyclic nucleotide phosphodiesterase 5. Eur J Pharmacol 431:17–24.
Hsu TH, Lai YL, and Kou YR (1998) Acetylcholine and tachykinin receptor antag-onists attenuate wood smoke-induced bronchoconstriction in guinea pigs. Eur
J Pharmacol 360:175–183.
Hwang TL, Wu CC, and Teng CM (1999) YC-1 potentiates nitric oxide-induced relaxation in guinea-pig trachea. Br J Pharmacol 128:577–584.
Ignarro LJ and Kadowitz PJ (1985) The pharmacological and physiological role of
cyclic GMP in vascular smooth muscle relaxation. Annu Rev Pharmacol Toxicol 25:171–191.
Kanoh S, Kondo M, Tamaoki J, Kobayashi H, Motoyoshi K, and Nagai A (2000) Effects of reactive oxygen species on intracellular calcium in bovine tracheal epithelium: modulation by nitric oxide. Exp Lung Res 26:335–348.
Kobayashi M, Nasuhara Y, Betsuyaku T, Shibuya E, Tanino Y, Tanino M, Takamura K, Nagai K, Hosokawa T, and Nishimura M (2004) Effect of low-dose theophylline on airway inflammation in COPD. Respirology 9:249 –254.
Koh SD, Campbell JD, Carl A, and Sanders KM (1995) Nitric oxide activates multiple potassium channels in canine colonic smooth muscle. J Physiol 489:735– 743.
Komalavilas P and Lincoln TM (1994) Phosphorylation of the inositol 1,4,5-trisphosphate receptor by cyclic GMP-dependent protein kinase. J Biol Chem 269:8701– 8707.
Leung TF, Wong GW, Ko FW, Li CY, Yung E, Lam CW, and Fok TF (2005) Analysis of growth factors and inflammatory cytokines in exhaled breath condensate from asthmatic children. Int Arch Allergy Immunol 137:66 –72.
Lin RJ, Wu BN, Lo YC, Shen KP, Lin YT, Huang CH, and Chen IJ (2002) KMUP-1 relaxes rabbit corpus cavernosum smooth muscle in vitro and in vivo: involvement of cyclic GMP and K(⫹) channels. Br J Pharmacol 135:1159–1166.
Lincoln TM and Cornwell TL (1991) Towards an understanding of the mechanism of action of cyclic AMP and cyclic GMP in smooth muscle relaxation. Blood Vessels 28:129 –137.
Murthy KS, Zhou H, and Makhlouf GM (2002) PKA-dependent activation of PDE3A and PDE4 and inhibition of adenylyl cyclase V/VI in smooth muscle. Am J Physiol 282:C508 –C517.
Nicholson CD, Challiss RA, and Shahid M (1991) Differential modulation of tissue function and therapeutic potential of selective inhibitors of cyclic nucleotide phos-phodiesterase isoenzymes. Trends Pharmacol Sci 12:19 –27.
Nijkamp FP, van der Linde HJ, and Folkerts G (1993) Nitric oxide synthesis inhib-itors induce airway hyperresponsiveness in the guinea pig in vivo and in vitro: role of the epithelium. Am Rev Respir Dis 148:727–734.
Robertson BE, Schubert R, Hescheler J, and Nelson MT (1993) cGMP-dependent protein kinase activates Ca-activated K channels in cerebral artery smooth muscle cells. Am J Physiol 265:C299 –C303.
Somlyo AP and Somlyo AV (1994) Signal transduction and regulation in smooth muscle. Nature (Lond) 372:231–236.
Torphy TJ and Cieslinski LB (1990) Characterization and selective inhibition of cyclic nucleotide phosphodiesterase isozymes in canine tracheal smooth muscle.
Mol Pharmacol 37:206 –214.
Turner NC, Lamb J, Worby A, and Murray KJ (1994) Relaxation of guinea-pig trachea by cyclic AMP phosphodiesterase inhibitors and their enhancement by sodium nitroprusside. Br J Pharmacol 111:1047–1052.
Vajner L, Konradova V, Uhlik J, and Zocova J (2002) The effects of intravenously administered methylxanthine preparations on the glycoconjugate composition of goblet cells in rabbit tracheal epithelium. Acta Histochem 104:107–112. Wu BN, Chen IC, Lin RJ, Chiu CC, An LM, and Chen IJ (2005) Aortic smooth muscle
relaxants KMUP-3 and KMUP-4, two nitrophenylpiperazine derivatives of xan-thine, display cGMP enhancing activity: roles of endothelium, phosphodiesterase and K⫹-channel. J Cardiovasc Pharmacol 46:600 – 608.
Wu BN, Lin RJ, Lin CY, Shen KP, Chiang LC, and Chen IJ (2001) A xanthine-based KMUP-1 with cyclic GMP enhancing and K⫹channels opening activities in rat aortic smooth muscle. Br J Pharmacol 134:265–274.
Wu BN, Lin RJ, Lo YC, Shen KP, Wang CC, Lin YT, and Chen IJ (2004) KMUP-1, a xanthine derivative, induces relaxation of guinea-pig isolated trachea: the role of the epithelium, cyclic nucleotides and K⫹channels. Br J Pharmacol 142:1105– 1114.
Zhang Y, Adner M, and Cardell LO (2005) Glucocorticoids suppress transcriptional up-regulation of bradykinin receptors in a murine in vitro model of chronic airway inflammation. Clin Exp Allergy 35:531–538.
Address correspondence to: Dr. Ing-Jun Chen, Department and Graduate Institute of Pharmacology, College of Medicine, Kaohsiung Medical Univer-sity, 100 Shih-Chuan 1st Road, Kaohsiung 807, Taiwan. E-mail: ingjun@ kmu.edu.tw
KMUP-3 and TNF-␣ Bronchocontraction
717
at Kaohsiung Med Univ Lib on September 27, 2012
jpet.aspetjournals.org