MaryAnnLiebert, Inc.,Publishers Pp. 1029-1036
Transcriptional
Activation
of the
Alpha-1
Acid
Glycoprotein
Gene
by
YY1 Is
Mediated
by
Its
Functional Interaction with
a
Negative Transcription
Factor
YU-MAY
LEE
and SHENG-CHUNG LEEABSTRACT
Regulation
ofalpha-1
acidglycoprotein
(AGP)
geneexpression
involves bothpositive
andnegative
transcription
factors. We havepreviously
identified two dominant factors:positive
andnegative
transcription factors,
AGP/EBP
and factorB, respectively,
involved in thetranscription
ofAGP and otheracute-phase
response genes. In thisreport,
wepresent
evidenceshowing
that thetranscription
of the AGP gene ispositively regulated by
atranscription
factor,
YY1. The activation of AGP geneby
YY1 ismediatedby
anegative
element B in theAGP
promoter
region.
YY1 can also activate the B motif linked to aheterologous
promoter. However,
YY1 does not binddirectly
to the B motifper se.Rather,
our datasuggest
that the activation of AGPgeneby
YY1
may be mediatedby
its functionalinteraction
withfactorB,
whichrecognizes
the B motif.INTRODUCTION
Alpha-1
ACID glycoprotein(AGP)
is one of themost
abundant,
acute-phase-responsive, liver-specific
plasma proteins.
Thebiological
function of AGP isun-known,
but there are indications that it may suppress theimmuneresponse(BennettandSchmid,
1980).
Some exper-imentssuggestthat AGP is anonspecific
antiinfectionagent(Friedmann, 1983),
and that it possesses nervegrowth-promoting activity
(Liu
et al.,1988).
Toinvestigate
theregulation
of AGP geneexpression during physiological
homeostasis and
during perturbed physiological
conditions (i.e.,acuteinflammation),
it isessentialtostudy
thecomplex
interactionsofvariousfratts-acting
factorsin these differentphysiological
states.In rats and
mice,
the levels of liver AGP mRNA andplasma
AGPprotein
increase10-to 100-fold within24hr ofexperimentally
induced inflammation(Baumann
and Held,1981;
Baumannetal., 1983, Kulkarnieío/., 1985;Baumannand
Maquat
1986;Darlington
et al., 1986; Gauldi etal.,1987;
Kleinétal.,1988;
ProwseandBaumann,
1988).
The increase in mRNA isprimarily
attributedtotranscriptional
activationofthe AGPgene
(Kulkami
etal., 1985;Baumannand
Maquat,
1986).
We havepreviously
identified four motifsrecognized by
apositive transcription
factor,
AGP/EBP,
and anegative
eis elementrecognized by
anegative
factor(i.e.,factor
B)
(Change/
al.,1990;
Leeetal.,1993).
During
theacute-phase
response,there is dramatic increase in the levelofAGP/EBP,
coupled
tothe decrease infactorB,
resulting
in the induction of AGP genetranscription
(Leeet al.,1993).
In additiontoAGP/EBP and factorB,
glucocor-ticoids(Klein
et al., 1988) have also been shown to be involved in the induction of AGP gene.Despite
the substan-tial progress inunderstanding
the factors inregulating
the activationof AGPgeneduring
theacute-phase
response,theregulation
ofthis gene in normalphysiological
statesremainstobe
investigated.
Theupstream
regulatory region
of the AGPgenecontainsasequence,GA ACATT TT(
—
121to
-113)
that is similartoamotif
recognized by
atranscription
factor,Y Y1 (consensussequence,
CGACATTTT) (Shi
et al.,1991).
YY1 is ahuman
Kriippel-related protein
that interacts with DNA elements(CGACATTTT,
located at -50 to-70;
CTC-CATTTT, locatedat the
transcription
initiationsite)
of the adeno-associated virus P5 promoter. These DNAelementsare
capable
ofrepressing transcription
directedby
heterolo-Institute of
Biological
Chemistry. AcademiaSinica,and Institute of MolecularMedicine,and Institute of Clinical Medicine,College
ofMedicine,National Taiwan
University, Taipei,
Taiwan.gouspromoters.El Anot
only
relievesrepression
exertedby
YY1 but also stimulatestranscription through
the YY1binding
site(Shi
etal.,
1991).
There is evidencethatYY1 mediates both therepression
and activationresponses(Shi
etal.,
1991).
Theability
ofYY1 to mediateopposite
effectsdepends
on the intracellular milieu(e.g.,
the presence orabsence ofEl
A).
YY1 isrelatedtothe GL1-Krüppel family
ofgenes(Ruppert
etal., 1988).
TheDrosophila Krüppel
protein
canalso repressoractivatetranscription,
depending
onthecontextofitsbinding
site(Frasch
andLevine,1987;
Ruppert
etal.,
1988;
Licht etal., 1990).
Two differentdomains in the
Krüppel protein
havebeenreported
toexhibitrepression activity
when fused to aheterologous
DNA-binding
domain. It remains to be determined whether theKrüppel protein
containsanactivation domain. In additiontoYY1,
severaltranscription
factors identical orhomologous
to YY1 have been identified and cloned.
Among
them are:protein
delta,
which bindstosequenceelements(GCNGC-CATC)
downstream of thetranscriptional
start sites in ribosomalprotein
genes andfunctionsas anactivator(Hari-haran et al.,
1991); NF-El,
which binds to theIgK
3' enhancer(CCACCTCCATCTT)
and functionsas arepressor(Park
andAtchison, 1991); UCRBP,
whichbindstotheUCRcore
(CGCCATTTT)
in thelong
terminalrepeatofMoloney
leukemia virus
(which
also functionsas arepressor)
(Flana-ganetal.,
1992);
andF-ACT1,
which bindscompetitively
tothemost
proximal
serumresponseelement(CGCCATATT)
of theskeletala-actingene and functionsas arepressor
(Lee
etal., 1992).
These dataindicate that thebinding specificity
of YY1 oritshomolog
isunusually
diverse.Furthermore,
anegative regulatory
domain in the humanpapillomavirus
type 18 promoter was shown to be the target site ofYY1(Bauknecht
etal.,1992).
Takentogether,
these datasuggest
that,
depending
on the context, YY1 can function as anactivator,
a repressor, or an initiator oftranscription.
Forthesereasons,wedecidedto
investigate
thepossible
involve-mentof the YYl-like motifin the
regulation
of AGP geneexpression.
In thisstudy
we examined theregulation
ofwild-type
and a series of mutants of AGP promoter in transient transfection assays. Wefound that YY1 canacti-vate AGP promoter
by
relieving
thenegative
action ofBelement,locatedat—40to—66.
However,
YY 1 didnotbinddirectly
to thisnegative
element.Instead,
it relieves therepression
of AGP geneby
its functional interaction with factor B.MATERIALS AND METHODS
Preparation of
nuclearextractLiver nuclearextractwas
prepared according
tothe proce-dures of Gorskietal.(1986)
from 200-to300-gram
Wistarratseitheruntreatedortreated with
lipopolysaccharide
(LPS)
(10
p,g/gram
body weight)
for 4 hr. Detailedprocedures
were asdescribed(Gorski
etal.,
1986).
Plasmids and
oligonucleotides
The
AGP/CAT, AGP/EBP/CAT,
and CMV-AGP/EBPwere as described
(Chang
etal.,1990;
Lee etal.,1993).
CMV-YY1
(kindly
provided by
Dr. T.Shenk)
wasdetailedelsewhere
(Shi
etal.,1991).
Various CATconstructscon-taining
the mutated AGPpromotersequenceweregenerated
essentially
asdescribed(Lee
etal.,1993).
pCAT-promoter
(CAT
reporter gene controlledby
SV40promoter)
andpSV-ß-Gal (ß-galactosidase
reporter gene controlledby
SV40
promoter)
wereobtained fromPromega
(WI).
Thesetworeportervectorswereusedfor
constructing
therecombi-nant
plasmids containing
theoligomeric
B motif. All theconstructswereverified
by
sequenceanalysis.
Theoligonu-cleotides used were as follows:
B,
TACTGTCCCTGGCT-TCAGTCCCATGCCCT;
U(UCR),
TAACGCCATTTTG-CAAGGCAT;
mutant UCR(mt
U);
TAAATACATTTT-GCAAGGCAT.Analysis of protein—DNA
interactionGel
mobility-shift
andfootprinting
assays were asde-scribed (Lee et al.,
1993).
Briefly,
an end-labeled DNAfragment
(—180
to +10 from AGPpromoterregion)
(5
ng)
generated by polymerase
chain reactions(PCR)
wasaddedtoa
20-p.l
reaction mixturecontaining
50 mM NaCl, 20 mM Tris-HClpH
7.6,
1 mMMgCl2,
0.2 mMEDTA,
10%glycerol,
5 mMDTT,and 1 p,gpoly(dl-dC).
Nuclearextractor
partially purified
fractioncontaining
factorBwasprein-cubated with recombinantYY 1or
partially
purified
YY 1 for5 minatroomtemperaturefollowed
by adding
theprobe
andincubated for 90 min on ice. DNase I
(30
p.g/ml, Sigma)
freshly
dilutedin 10mMMgCL
and 5 mMCaCl2
wasaddedtothe reaction mixture
(usually
1-3pJ
wasused).Digestion
wasperformed
for2-3 minonice andstopped by
the additionof 80
pd
ofstopsolutioncontaining
75 u-gofyeasttRNA/ml,20 mM EDTA, and 0.5%
NaDodS04.
Thesamples
wereextracted with
phenol
chloroform,
precipitated
with ethanolat -70°C. The
pellets
were dried andresuspended
inse-quencing
solution (95%formamide,
1%xylene cyanol
FF,
and 1%
bromphenol
blue),
heated at 95°C for 3min,
and loadedontoasequencing gel.
For
gel mobility-shift
assays,protein-DNA complexes
wereformed asdescribed forthe
footprinting
assays. Afterincubation for 20 min at room temperature, 1
|xl
of 1%bromphenol
bluewasadded,
and thesample
wasloadedontoa 5%
polyacrylamide gel (acrylamide/bisacrylamide
=30:1)
containing
4%glycerol
inTris-Glycin
buffer.Electro-phoresis
wasperformed
at 150 Vatroom temperaturewithbuffercirculation.
Recombinant YYI and
antibody
to YY1Recombinant YYI was
generated
by cloning
the Ncol-Eco RI
fragment
of YYI cDNA(YYI
plasmid
waskindly
provided by
Dr. T.Shenk,
PrincetonUniversity)
intoplas-mid
pRSET
(Invitrogen,
SanDiego,
CA).
Therecombinant YYIexpressed
in Escherichia coli[BL21 (DE3)]
waspurified
by
Ni column anddetectedby
Western blotusing
rabbitpolyclonal
anti-YYl. Rabbitanti-YYl wasgenerated
by
immunizing
theantigen expressed
inE. coli.Briefly,
acDNA
fragment corresponding
topolypeptide
of232 amino acidscontaining
thecarboxy-terminal
part of YYI wassequence was verified
by
sequenceanalysis.
The cDNAfragment
was cloned intopRSET
(Invitrogen,
SanDiego,
CA)
andexpressed
inBL21(DE3).
Cell
cultures,
DNAtransfections,
and CAT assaysBaby
hamsterkidney
(BHK)
cells were cultured inDul-becco's modified
Eagle's
mediumsupplemented
with 10% fetal calfserum. DNAtransfectionswereperformed by
thecalcium
phosphate
precipitation technique.
For each 10-cmpetri
dish,
the calciumphosphate-DNA precipitate
con-tained 8 p.g of target
plasmid,
and 3 u,g CMV-YYI oramino-terminal deletedYYI
(CMV-C250),
whichcontains thecarboxy-terminal
250 amino acids.PlasmidpGEM4
wasusedas acarrierto make the finalDNA tobe 15 p-g. Cells
wereharvested48hr
post-transfection.
Cellextract(30u,g)
was usedfor CATassay. CAT
activity
wasquantitated by
densitometric
scanning
of theautoradiogram
orby scanning
ofthe TLC
plate
with BioRadImage
analysis
system.Western blot
analysis
Rat liver nuclear extracts were
prepared
as described(Chang
etal.,1990).
Liver nuclearextracts were obtainedfromcontrol and LPS-treated Wistarrats.
Varying
amountsof nuclear extracts were resolved on a 10%
NaPodS04-polyacrylamide gel.
Proteinswereelectroblottedtonitrocel-lulosemembrane.The
protein
blotwassubjected
tostandard Western blotdevelopment
using
rabbit anti-YYl and goat anti-rabbitIgG
alkalinephosphatase conjugate.
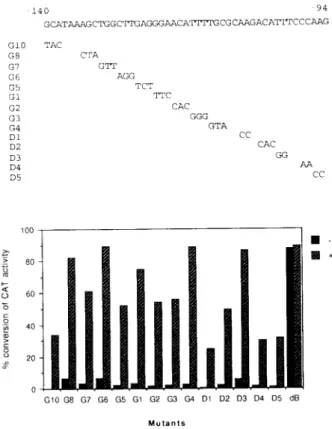
< ü -122 GGAACATTTTGCGCAAGA -115 Gl TTC G2 CAC G3 G4 GGG GTA
FIG. 1. 7Va«s-activation of the AGP gene
by
YYI. Cotransfection assays ofCMV-YY1expression
vectorandwild-type
and mutated AGP sequences linked to CATre-porter gene. The upstream
regulatory
sequence(—122
to
-155)
of the AGPgene and thecorresponding
mutantsareshown in the upper
panel.
The results ofCAT
assay areshown in the lower
panel.
CMV-YYI(3
p,g)
and 10 p,gof reportergenewerecotransfected intoBHK cells.(—
)Cellstransfected with reporter gene without
CMV-YY1;
(
+)
cotransfectedwith CMV-YY1.
RESULTS
Transcriptional
activationof
AGPpromoter
by
YYITo
investigate
thepossible
involvementofYYI in AGP generegulation,
we initiated functional and biochemicalcharacterizations of YYI on AGP
regulation.
BHK cells were used for the transfection assays because BHK andHepG2
cells behavedsimilarly
as demonstratedby
ourprevious
studies(Lee
etal.,
1993).
Transfection ofwild-type
and severalmutantsof AGP-CAT
together
withCMV-YYI showed that YYI could activate AGP promoter(Fig.
1). Mutationsinthesequence,GAACATTTT,
donothave anyeffecton YYl'sactivation. These results indicate thatYYI can activate AGP
transcription,
butindependently
of the sequence, GAACATTTT. Furtherexperiments by gel
mo-bility-shift
orfootprinting
assayusing
recombinant YYIderived fromE.coli have failedtoshowthat Y Y1couldbind
tothissequence
(data
notshown).
Having
demonstrated that YYI couldrra/w-activate AGP gene, we tested itspossible
interaction with themost
prominant
activator for AGPgene, AGP/EBP. Itturnedoutthat activation of AGPgeneby
YYI and AGP/EBPis additive(Fig.
2). Therefore,
the activation ofAGP geneby
YYI and AGP/EBPareindependent
event.To understand the
implications
of these datafurther,
wegenerated
a series ofmutantsspanning
from—
140 to —94 and a B-element-truncated mutant
(Lee
etal.,1993).
Thenucleotidesequence and theco-transfectiondataare shown
in
Fig.
3. YYI could activatewild-type
aswell as every8
F-< 20H
Ü o o + +AGP/EBP
YY1
FIG. 2. The activation of AGP promoter function
by
AGP/EBP and YY-1 is additive.AGP/CAT,
CMV-YY1,
andCMV-AGP/EBPweredetailed in the
previous
sectionorelsewhere. BHKcellswereused forthetransfection assays.
AGP/CAT
(10
p.g)
and 1 pug of YY1 and/or CMV-AGP/EBPwereusedfor the transfectionassay.-140 -94 GCATAAAGCTGGCTTGAGGGAAGATTTTGCGCAAGACATTTCCCAAG GIO TAC G8 CTA G7 GTT G6 AGG G5 TCT Gl TTC G2 CAC G3 GGG G4 GTA Dl CC D2 CAC D3 GG D4 AA D5 CC 100 G10 G8 G7 G6 G5 G1 G2 G3 G4 D1 D2 D3 04 D5 dB Mutants
FIG. 3.
7Vww-activationofAGPgeneby
YYIismediatedby
aB motif. Cotransfection assaysofCMV-YY1expres-sionvectorandmutatedupstream
regulatory
sequence(span-ning
from -140to-94)
of the AGPgene linkedto CATreportergene.Themutated sequencesareshown intheupper
panel.
dB is thedeletedmutantofthe B motif linkedtoCAT,as
reported
inourearlierpublication
(Lee
etal.,1993).
Theamountsof
plasmid
and thecells usedforthe transfectionarethesameasin
Fig.
1.(-
and+)Transfectioninthe absenceandinthepresenceofCMV-YY1.
mutated AGP/CATexceptthe B-truncated mutant
(Fig.
3,
lower
panel,
dB).
These data indicatethat YYI isunlikely
dependent
onasingle
motifspanning
from -140to -94of AGPpromoterforthe activation.However,
whenthe B-mo-tif-deletedmutant wasco-transfected withCMV-YY 1, theactivation was abolished
(Fig.
3,dB).
As detailed in ourearlier
publication
(Lee
et al.,1993),
the B motif is anegative
element that isrecognized by
factorB. The CATactivity
ofB-motif-deletedmutantwassubstantially
elevated ascompared
with thewild-type
orother mutatedAGP/CAT,
consistent with the
negative
effect ofthe B motif.Having
observed that YYI could activate the AGPpromoter but failedtodosowhentheBelementwasdeleted, weinitiatedstudiesonthe
possible recognition
ofthe B motifby
YYI.Extensive
experiments using
a number ofoligonucleotide
probes
andrecombinant YYI orYYIpurified
fromHepG2
failedto show that YYI can bind to the B motif
(data
notshown).
These datasuggestthat the observedactivationof YYI on the AGP promoter isunlikely
to be mediatedby
direct interaction of YYI and the B
element; rather,
afunctional interactionmaybe the
likely explanation.
100
Wt -71 OligoB pCAT-P dB
Reporter
FIG. 4. Transfection assays demonstrate that the activa-tion of AGP
promoter
functionby
YY1 isthrough
the B motif. CAT reporter constructscontaining
thewild-type
AGPpromoter,
deletion mutants of B motif(dB),
andupstreamof-71
(-71),
oligomeric
B motic(BIO)
linkedtoSV40
promoter-CAT
(oligo
B),
and SV40promoter-CAT
(pCAT-P)
wereused fortransfection(-)
orcotransfectionwithCMV-YY1
(+)
into BHK cells.Reporter plasmid
(10
p-g)
and3 p.gof CMV-YY1 wereused.YYI can activate theB
motif
linked toheterologous
promoter
The above
experiments
showedthatthe activationofYY 1onthe AGPpromoterismediated
by
theBelement,butnotby
directprotein-DNA
interactions. To address further the issueofhowtheB motifmight
mediatetheactivationof the AGP promoterby
YYI,
we constructed aheterologous
promoter
(SV40)
containing
oligomeric
Bsequencesaswellas -71/AGP
(upstream
of -71 of AGPpromoter
wasdeleted)
promoterlinkedtoCAT(Lee
etal.,1993).
Wild-type and -71/AGP promoter CAT could be activated
by
CMV-YY1(Fig.
4).
YYI could also activateoligomeric
B-linkedheterologous
promoters. Instriking
contrast tothese
data,
YYI had no effect on promoters that do notcontain the Belement. Consistent with theconclusionthat B isa
negative
element fromprevious
studies,heterologous
promoters
containing oligomeric
B(B10)
element showed reducedactivity.
To rule out thepossibility
that there is adifference in transfection
efficiency
betweenexperiments,
we used
B,0-linked-pSV-ß-Gal
for the transfection and co-transfectionassays.TheB10-linked
pSV-ß-Gal
wasmuchweaker thanthe
pSV-ß-Gal
whenstaining
withX-Gal(data
not
shown).
However,
whenYYI wasco-transfected withB,0-pSV-ß-Gal,
theß-Gal activity
was elevated(data
notshown).
Thisincrease inactivity
isnotduetothe increase in transfectionefficiency
when YYIwaspresent,asevidentby
the apparentnumberofcells
positive
inX-Galstaining
arerelatively
constant(data
notshown).
These datasuggestthat YY 1 isinvolvedinmodulating
thefactorBactivity
eitherby
directprotein-protein
orfunctional interactionwith Bfactor orby repressing
theexpression
oftheBfactorgene.To rule out furtherany
possible
artefactsresulting
from these cotransfectionexperiments,
we constructed a YYIexpression
vector(CMV-C250)
in whichonly protein
con-taining
thecarboxy-terminal
250 amino acids could beZn Finger
I i CMV-C250
1 25 2.S 1 23 1.1 1 29 2.1 1 25 1.2
FIG. 5. The /ra/js-activation of the AGP gene
by
YYIrequires
thefull-length
YYI molecule. The upperpanel
shows the construction of YYI and the truncated YYIexpression
vectors,CMV-YY1 andCMV-C250. Thelowerpanel
shows the trans- activation of AGP/CATby
CMV-YY1 but notby
CMV-C250. Cotransfectionexperiments
weredetailed in the
legends
ofFigs.
1and 2.The numbersontopof the lower
panel
indicate the fold stimulationrelativetothe basal CAT
activity
of each CAT reporter construct(normalized
to1).
(—)
Transfection with the CATreporterplasmid
alone;
(YY)
cotransfection withCMV-YY1; (C)
cotransfection with CMV-C250.
Wild-type
(wt)
or mutantAGP-CAT
(G1,
G2,
andG3)
areshownonthebottomline ofthe lower
panel.
demonstrated thatwhentheanimo-terminal
portion
ofYYIwas
deleted,
it failed to activate the AGP gene(Fig.
5).
Taken
together,
the activation of theAGPgeneby
YYI notonly requires
the B motif but alsoneeds theamino-terminalportion
ofYYI molecule.YYI doesnot
interfere
with thebinding
of
Bfactor
toits
cognate
motif
Having
demonstrated thatYY 1 canactivatetheAGPgenethrough
the Bmotif,
we would liketo knowwhether YYI interactedwith the Bfactorphysically.
Purified YYI(from
liver)
or recombinant YYI was incubated with rat liver-derived B factor. Purified YYI didnotinterfere with theB factorbinding
to itsmotifinfootprinting analysis
(data
not shown) or ingel mobility-shift
assay in the presence orabsence of
specific antibody
to YYI(data
notshown).
Purified AGP/EBP or
protein
derived from other columnfractionsbehaved
similarly.
Furthermore,
recombinantYYI did not inhibitthefootprinting activity
of B factor. These data indicated that YYI did not form aprotein-protein
complex
with factor B; rather, it interacted with B factorfunctionally
withoutinterfering
with theDNA-binding
activ-ity
ofBfactor.The level
of
YYI does notchange
during
theacute-phase
responseWe have shown
previously
that the levelsofAGP/EBP (themostprominant positive
factor)
and Bfactor(a
negative
factor)
are induced and reducedduring
theacute-phase
reaction
(Change
etal., 1990;Leeetal.,
1993).
This may account in part for the net induction of theacute-phase-response genes. Becausewehavenowshown that YYI can
activatethe
transcription
ofAGP gene, it would beinterest-ing
toexamine thechange
intheprotein
levelaswellasthebinding activity
of YYIduring
theacute-phase
response. Liver nuclear extracts wereprepared
from untreated orLPS-treated rats and usedfor
gel
retardationassays(using
UCRoligonucleotide,
which also contains anoverlapping
AGP/EBP-binding
site)
and Western blotanalysis.
Both thebinding
activity
and theprotein
level of YYI remainsconstant whether pre- or
post-acute-phase
response(Fig.
6).
AGP/EBPwasincreasedduring
theacute-phase
response,whichwas anice internalcontrol. Taken
together,
these datasuggest that the
protein
level of YYI does notchange;
however, itsfunctionalactivity
for interactionwithBfactor maybe elevated(e.g.,
through post-translational
modifica-tion)
during
acuteinflammation.DISCUSSION
Mechanism
of
activationof
AGPgeneby
YYI In this paper we have demonstrated that YYI activatestranscription
directedby
the AGPpromoterorheterologous
(SV40)
promotercontaining
thenegative
element B. YYI repressesactivity
ofthe SV40enhancer/promoter containing
theYYI-binding
motif(Shi
etal.,1991).
Thefundamental difference between theYYI motif-mediated and the B-mo-tif-mediatedactivity
of YYI is the involvement of direct Y Y1 -DNA interaction in the formercaseandonly
functionalinteraction of YYI and B-motifin the lattercase. Two lines
of evidence suggest that the B motif of AGP promoter mediates YYl's activation ofAGPgene:
(i)
the activation of AGP promoterby
YYIdepends
on the presence of Bsequence;
(ii)
YYI canactivateB-motif-mediatedrepression
of SV40 promoter when the B motif is inserted into this promoter. An extensive search failed to reveal any DNA
elementinthe AGPpromoter
region
thatcanberecognized
by
Y Y1. HowcantheB sequence mediate the action of YY 1without direct
DNA-protein
interaction? Because the B elementwasshowntoberecognized
by
anegative
transcrip-tion factorB
(Lee
etal.,1993),
fourpossible
mechanismsmay exist:
(i)
direct YYI and factor B interaction at theprotein
level resulted inthe reducedrepressoractivity
of Bfactor;
(ii)
YY 1 could repress theexpression
of theBfactor,
resulting
inareducedprotein
level of factorB; (iii)
there isonly
a functional interaction between YYI and factor B without directprotein-protein
interaction,
e.g.,modulating
offactor B'sactivity by
YYI; (iv)
YYI interacts with factor Bfunctionally
viaanintermediary.
For the firstpossibility,
itcould be a
physical
interaction between YYI and factor B.For the second
possibility,
YYI may bind to thecognate
motif in theregulatory region
ofthe factorB gene and result inrepression
ofexpression
of factorB. For the thirdpossi-bility,
YY 1 mayaffect factorB'sactivity by
modification of factorBpost-translationally.
Forthe fourthpossibility,
there mayexistacorepressorfor theinteraction of YYI andfac-torB.
Twolines of evidence indicatethat YYI interacts with B factor
functionally,
butnotphysically
(i.e.,complex
forma-B LPS N
YY-1_AGP/EBP
L N LNLNLNm
70--351234561
2345
6FIG. 6. Determination of the
protein
level and theDNA-binding activity
of YYI in thenuclearextractsprepared
from normal andLPS-treated liver.A. Westernblotanalysis showing
therelativeconstantprotein
level of YY l in normalratliver nuclearextract(N)
andextractprepared
from LPS-treated liver(LPS).
Rabbitantibody
toYYI wasusedfortheblotanalysis.
The AGP/EBPlevel
(indicated
by
a35andarrow)wasusedas apositive
controltoshow its increaseduring
theacute-phase
response.B.Gelmobility-shift
assayshowing
theconstantlevel inbinding
activity
of YYI in normal livernuclearextractandextractfrom liver of the
acute-phase
response(lanes
1-6 contain10, 8, 7, 6,4,
and2 p.g of nuclearextract,respectively).
Thearrowindicates the
YYl-DNA
complex;
thearrowheadindicatesthe AGP/EBP-orAGP/EBP-related factors.tion): (i)
bothpurified
andrecombinantYYI cannot inhibit thefootprinting activity
of factor B, andantibody
to YYI failed topattern
ofcomplex
formation inabandshiftassay;(ii)
although
YYI cannotbindtothe Bmotif,
itcanactivategenesthat containaBmotif.The
following examples
showthat YYI could affect gene
expression through
a thirdproteins:
(i)
YYIrepression
of the P5promoterisrelievedby
adenovirus E1Aprotein
(Shi
et al.,1991). (ii)
A fusionprotein
of YYI and the GAL4 DNAbinding
domainre-presses
transcription
of athymidine
kinase promoter withGAL4
binding
sites(Seto
etal.,1991).
(iii)
c-Myc
canformcomplex
withYYI and thus modulates theactivity
ofYYI(Shrivastava
et al.,1993).
Asimple
model toexplain
thefunctional
versatility
of YYI is that its interaction with different cellularproteins
may alter itsactivity. During
theacute-phase
response, thebinding activity
of factor Bde-creases while both the
protein
level and theDNA-binding
activity
of YYI do notchange.
Theactivity
of YYI canbemodulated
during
theacuteinflammationin such waysothatits functional interactionwith factor B isenhanced,whilethe
DNA-binding activity
of factorB is unaffected. However, this modification of YYl'sactivity,
whichultimately
affects theactivity
of factorB,maynotinvolvein the formationof YY1-Bcomplex. Alternatively,
factorBis modifiedduring
theacute-phase
response andresults inthe enhancedfunc-tional interaction with YYI.
Furthermore,
both of thesepossibilities
exist. Futureexperiments
shouldbedesigned
toresolve this issue.
A number ofsystemsin which
protein-protein
interactions resulted inaltered functions ofaspecific
transcription
factorhave been demonstrated
(Lee
etal., 1993;
Nishio et al., 1993;Steinetal.,1993).
The functions ofYYIaremediatedmainly by
itsbinding
toaspecific
DNA motif(Hariharan
etal.,
1991;GualbertOÉ>rer/.,
1992;Montalvoe<a/.,1991;
Park andAtchison,
1991;
Seto et al.,1991;
Shi etai,
1991;
Bauknechtetal., 1992;
Flanagan
etal.,1992;
Lee etal.,
1992). Therefore,
YYI mayalso modulatetheexpression
of factor Btranscriptionally through
itsbinding
to theregula-tory
regions
of factorB gene. Definitiveanswers mustwait until the factor Bgeneisisolated and cloned. Takentogether,
our present data and the
rapid
reactionduring
the acuteinflammationsuggestthatthe functional interaction ofYYI andfactorB,rather than the
transcriptional
mechanismis themore
likely
mechanisminmodulating
thefactorBactivity.
However,
despite vigorous
tests(performed
by
anti-YYlantibody depletion
followedby
footprinting
assayortheuseof anti-YYl
antibody
ingel mobility-shift
assay),
we havefailed to demonstrate that the YYI and factor B
complex
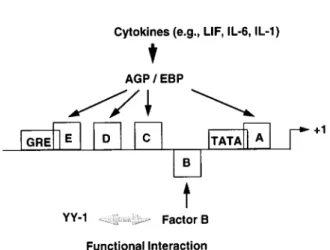
exists.TRANSCRIPTION
OF AGP
GENE
Cytokines
(e.g.,LIF, IL-6,IL-1)
YY"1 Factor B
Functional Interaction
FIG. 7. Schematic
representation
oftheregulation
ofAGPgene
by
varioustranscription
factors.Cytokines
stimulate AGP/EBP- orAGP/EBP-related /ra/w-activators andresultin the induction ofAGP
during
theacute-phase
response. YYIparticipates
in the AGPregulation by repressing
the functionalactivity
of anegative
factor,
B.Thus,
YYI activatesAGPgeneindirectly by
functional interaction with factorB.is its induction
during
theacute-phase
response. We havepreviously
shown the level of AGP/EBP increased several foldduring
theacute-phase
response.Wealso documentedadecreaseinthe
binding activity
of factorB. In thisreport,wepresented
evidence forthe activation of AGPgeneby
YYl,including
that(i)
this activation is mediatedby
Bmotif;
(ii)
the activation of AGP gene
by
YYIrequires
the amino-terminalportion
ofYYI molecule;and(iii)
YYl'sfunction inactivating
AGPgene isindependent
ofAGP/EBP.Boththe
protein
level and thebinding activity
ofYYI donot
change
inratliverduring
theacute-phase
response(Fig.
6).
Becausethemechanism of reduction of factorB's
binding
activity
during
theacute-phase
response remains to besolved,
it ishighly possible
that somepost-translational
mechanisms are involved in
regulating
factor B'sactivity,
suchasinteracting
withYY 1.Ifthisindeedhappens,
then thefunctional
activity
ofYYI undernormalphysiological
con-ditions must be different from that
during
theacute-phase
response. Theregulatory
roleofYYI onfactorB's leveloractivity
offersanattractive model forgeneregulation:
Reg-ulation of a
negative
factorby
anothernegative/positive
(Yin-Yang-1)
factor. A schematic modesummarizing
theregulation
ofAGPgene ispresented
inFig.
7.Additive
effect of
activationby
YYIand
AGP/EBPon the AGP
promoter
We have shown
previously
that AGP/EBP couldactivate AGPtranscription
and that AGP/EBP is thekey
nuclear factor inconveying
thesignals
[i.e.,
cytokines
like interleu-kin-1(IL-1),
IL-6,
or leukemiainhibitory
factor]
to their targetgenes(e.g.,
AGP)
during
theacute-phase
response.Todetermine whether the activation effects of AGP/EBPand YYI onAGPare
independent
events,weperformed
trans-fection
experiments using
wild-type
AGP/CAT,
CMV-AGP/ EBP, and CMV-YY1. As shown inFig.
2,
AGP/EBPand YY1could activate theAGPpromoter,and theeffect of thesetwo
expression
vectors seems to be additive. These datasuggest that AGP/EBP and YYI donot interact with one
another to activate the AGP gene, but ratheract
indepen-dently
inactiviating
the AGPgene.These resultsareconsis-tentwith thedata that the
activity
ofY Y1depends
onthe Bmotifwhereas AGP/EBP
depends
on other motifs. Takentogether,
YYI,AGP/EBP,
C/EBP-a, and factor B areimportant trans-acting
factors inregulating
AGPgenetran-scription.
The data present in this paper areparticularly
important
in thepotential
modulation of YYl'sactivity
during
theacuteinflammation and itssubsequent
interaction withfactorB. Furtherunderstanding along
these lines mayclarify
notonly
factor B'sregulation
but alsoYY 1'$mode of action andregulation.
ACKNOWLEDGMENTS
WethankDr.
Tung-Tien
Sunforacriticalreview of themanuscript,
Mr.Tso-pang
Yao forperforming
thePCR of YYI cDNA,Dr.Wuh-Liang
Hwufortechnicalhelp,
Wen-Hai Tsai fornuclearextracts, and Drs.T. Shenkfor CMV-YY1andK. Ozato fortheexpression
vectorofUCRBP. This research wassupported by
grants NSC82-0412-B002-251fromtheNationalScience Council.
REFERENCES
BAUKNECHT, T., ANGEL, P., ROYER, HD., and ZUR
HAUSEN, H. (1992). Identification ofa
negative
regulatory
domain in the human
papillomavirus
type 18promoter: interac-tion with thetranscriptional
repressor YYI. EMBO J. 11,4607-4617.
BAUMANN, H., and HELD, W.A. (1981).
Biosynthesis
andhormone-regulated expression
ofsecretoryglycoproteins
in ratliver and
hepatoma
cells.J. Biol. Chem.256, 10145-10155.BAUMANN, H., and
MAQUAT,
L.E. (1986). Localization of DNAsequences involvedindexamethasone-dependent
expres-sionoftheratalpha-1
acidglycoprotein
gene. Mol. Cell. Biol.6,2551-2561.
BAUMANN, H.,FIRESTONE,G.L.,BURGESS, TL., GROSS, K.W., YAMAMOTO,K.R., andHELD, W.A. (1983). Dexa-methasone
regulation
ofalpha-1
acidglycoprotein
and otheracute
phase
reactants in ratliver andhepatoma
cells. J. Biol. Chem.258,563-570.BAUMANN, H., HILL, R.E.,SAUDER, D.N., andJAHREIS,
G.P.(1986).
Regulation
ofmajoracutephase plasma proteins by
hepatocyte stimulating
factors of human squomas carcinoma cells. J. Cell Biol.102,370-383.BENNETT, H., andSCHMID,K. (1980).
Immunosuppression
ofhuman
plasma alpha-1
acidglycoprotein: Importance
of thecarbohydrate
moiety.
Proc. Nati. Acad. Sei. USA 77,6109-6113.
CHANG, C.J., CHEN, T.T., LEI, H.Y., CHEN, D.S., andLEE,
S.C. (1990). Molecular
cloning
ofatranscription
factor, AGP/EBP,that
belongs
tomembersof the C/EBPfamily.
Mol. Cell. Biol. 10,6642-6653.DARLINGTON, G.D., WILSON. R.. and LACHMAN, L.B.
(1986).
Monocyte-conditioned
medium, interleukin-1, andtu-mornecrosis factor stimulate theacute
phase
response in humanhepatoma
cells in vitro. J. Cell Biol. 103,787-793.FLANAGAN,J.R.,BECKER, K.G., ENNIST,D.L.,GLEASON, S.L., DRIGGERS, P. H., LEVI, BZ., APPELLA, E., and
OZATO, K. (1992).
Cloning
ofanegative transcription
factorthat bindstotheupstreamconserved
region
ofMoloney
murine leukemia virus. Mol. Cell. Biol. 12,38-44.FRASCH,M.,andLEVINE, M.(1987).
Complementary
patternsof
even-skipped
and fushitarazuexpression
involve their differ-entialregulation
by a common set ofsegmentation
genes inDrosophila.
Genes & Dev. 1,981-995.FRIEDMANN, M.L. (1983). Control of malaria virulence by
alpha-1
acidglycoprotein
(orosomucoid),anacutephase
(inflam-matory)reactant.Proc. Nati. Acad. Sei. USA80,5421-5424.
GAULDI,J.,RICHARDS, C,HARNISH. D.,LANDSORP,P.,
and BAUMANN, H. (1987). Interferon beta-2 shares
identity
with monocytederived
hepatocyte-stimulating
factor and regu-lates themajor
acutephase protein
response in liver cells. Proc.Nati. Acad. Sei. USA84,7251-7255.
GORSKI, K., CARNEIRO, M., and SCHIBLER. U. (1986).
Tissue-specific
in vitrotranscription
from the mouse albuminpromoter.Cell47,767-776.
GUALBERTO, A., LePAGE, D., PONS, G., MADER, S.L., PARK, K., ATCHISON, M.L.,andWALSH,K.(1992). Func-tional
antagonism
between YY 1 and theserumresponse factor. Mol. Cell. Biol. 12,4209-4214.HARIHARAN, N., KELLEY, D.E., and PERRY, R.P. (1991). Delta,a
transcription
factor that bindstodownstream elements in severalpolymerase
IIpromoters, isafunctionally versatile zincfinger protein.
Proc. Nati. Acad. Sei. USA88,9799-9803.KLEIN, E.S., DlLORENZO, D., POSSECKERT. G.. BEATO, M.,andRINGOLD,G. M.(1988).
Sequences
downstream of theglucocorticoid
regulatoryelement mediatecycloheximide
inhibi-tion of steroid-inducedexpression
from the ratalpha-1
acidglycoprotein
promoter:evidence foralabiletranscription
factor.MoL Endocrinol.2, 1343-1351.
KULKARNI, A.B., REINKE, R., andFEIGELSON, P. (1985).
Acute
phase
mediators andglucocorticoids
elevatealpha-1
acidglycoprotein
genetranscription.
J. Biol. Chem. 260, 15386-15389.LEE, J.S., GALVIN, K.M.. and SHI, Y. (1993). Evidence for
physical
interaction between thezinc-finger transcription
factor YYIandSpl.
Proc. Nati. Acad. Sei. USA90,6145-6149.LEE, T.C., SHI, Y.,andSCHWARTZ,R.J. (1992).
Displacement
of BrdUrd-induced YYI by serum response factor activates skeletal
alpha-actin transcription
inembryonic myoblasts.Proc. Nati. Acad. Sei. USA89,9814-9818.LEE, Y.M., TSAI, W.H., LAI, M.Y.,CHEN, D.S., andLEE,
S.C. (1993). Induction of liver
alpha-1
acidglycoprotein
geneexpression
involves bothpositive
andnegative transcription
factors. Mol. Cell. Biol.13,432-442.LICHT,J.D.,GROSSEL,M.J..FIGGE,J.,andHANSEN,U.M.
(1990).
Drosopkila Krüppel protein
isatranscriptionalrepressor.Nature346,76-79.
LIU,H.TAKAGAKI, M.K., andSCHMID, K. (1988). In vitro
nerve-growth-promoting activity
of humanplasma alpha-1
acidglycoprotein.
J. Neurosci. Res. 20,64-72.MONTALVO, E.A., SHI, Y., SHENK, T., andLEVINE, A.J.
(1991).
Negative regulation
of theBZLF1 promoterofEpstein-Barr virus. J. Virol.65,3647-3655.
NISHIO, Y„ ISSHIKI, H., KISHIMOTO, T., and AKIRA. S.
(1993). A nuclear factor for interleukin-6
expression
(NF-IL6)and the
glucocorticoid
receptorsynergistically
activatetranscrip-tion of therata-1 acid
glycoprotein
gene via directprotein-protein
interaction. Mol. Cell. Biol. 13, 1854-1862.PARK, K.,andATCHISON,M.L.(1991).Isolation ofacandidate
repressor/activator,
NF-E1(YY-1, delta),that bindstotheimmu-noglobulin
k 3' enhancer and theimmunoglobulin
heavy-chain uEl site. Proc. Nati. Acad. Sei. USA88,9804-9808.PROWSE,K.R.,and BAUMANN,H.(1988).
Hepatocyte-stimu-lating
factor, beta-2-interferon, and interleukin-1 enhanceex-pression
of the ratalpha-1
acidglycoprotein
gene via adistalupstream
regulatory region.
Moll. Cell. Biol.8,42-51.RUPPERT, J.M., KINZLER, K.W., WONG, A.J., BIGNER, S.H., KAO, FT.,LAW, F.L., SEUANEZ, H.N., O'BRIAN, S.J.,andVOGELSTEIN,B.(1988).
TheGLl-Kriippel family
of human genes. Mol. Cell. Biol.8,3104-3113.SETO, E., LEWIS, B., and SHENK, T. (1993). Interaction between
transcription
factorsSpl
and YYI. Nature 365, 462-464.SETO, E., SHI, Y., and SHENK,T. (1991). YYI is aninitiator
sequence-binding protein
that directs and activatestranscription
in vitro. Nature354, 241-245.
SHI, Y., SETO, E., CHANG, L.S., and SHENK, T. (1991).
Transcriptional repression by
YY-1, a humanGLl-Kriippel-related
protein,
and relief ofrepression by
adenovirus El Aprotein.
Cell67,377-388.SHRIVASTAVA, A., SALEQUE, S., KALPANA, G.V., ARTANDI.S., GOFF, S.P.,andCÁLAME,K.(1993). Inhibi-tion of
transcriptional
regulatorYin-Yang-1 by
association withc-Myc.
Science262, 1889-1892.STEIN, B.. COGSWELL, P.C., and BALDWIN, A.S. (1993).
Functional and
physical
associations between NF-kB and C/EBPfamily
members: A Rel domain-bZIP interaction. Mol. Cell. Biol. 13,3964-3974.Address
reprint
requests to: Dr.Sheng-Chung
Lee Instituteof
Molecular MedicineCollege of
MedicineNationalTaiwan