doi:10.1152/ajpcell.00019.2005
289:748-756, 2005. First published Apr 20, 2005;
Am J Physiol Cell Physiol
Rachel J. Webb, Jacob D. Judah, Lee-Chiang Lo and Geraint M. H. Thomas
trans-Golgi
reversible protein tyrosine phosphorylation events in
Constitutive secretion of serum albumin requires
You might find this additional information useful...
63 articles, 15 of which you can access free at: This article cites
http://ajpcell.physiology.org/cgi/content/full/289/3/C748#BIBL 3 other HighWire hosted articles:
This article has been cited by
[PDF] [Full Text] [Abstract] , September 14, 2007; 101 (6): 607-616. Circ. Res.
Chapman, J. L. Stow, W. Jessup and L. Kritharides
M. Kockx, D. L. Guo, T. Huby, P. Lesnik, J. Kay, T. Sabaretnam, E. Jary, M. Hill, K. Gaus, J. Calcium-Dependent Pathway Along the Microtubule Network
Secretion of Apolipoprotein E From Macrophages Occurs via a Protein Kinase A and [PDF] [Full Text] [Abstract] , December 1, 2007; 18 (12): 4813-4825.
Mol. Biol. Cell
F. Ferraro, X.-M. Ma, J. A. Sobota, B. A. Eipper and R. E. Mains Secretory Granule Maturation
Kalirin/Trio Rho Guanine Nucleotide Exchange Factors Regulate a Novel Step in [PDF] [Full Text] [Abstract] , June 1, 2008; 28 (6): 1060-1067.
Arterioscler. Thromb. Vasc. Biol.
M. Kockx, W. Jessup and L. Kritharides
Regulation of Endogenous Apolipoprotein E Secretion by Macrophages
including high-resolution figures, can be found at: Updated information and services
http://ajpcell.physiology.org/cgi/content/full/289/3/C748
can be found at: AJP - Cell Physiology
about Additional material and information
http://www.the-aps.org/publications/ajpcell
This information is current as of August 31, 2009 .
http://www.the-aps.org/. American Physiological Society. ISSN: 0363-6143, ESSN: 1522-1563. Visit our website at
a year (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the is dedicated to innovative approaches to the study of cell and molecular physiology. It is published 12 times
AJP - Cell Physiology
on August 31, 2009
ajpcell.physiology.org
CALL FOR PAPERS
Protein and Vesicle Trafficking, Cytoskeleton
Constitutive secretion of serum albumin requires reversible protein tyrosine
phosphorylation events in trans-Golgi
Rachel J. Webb,1Jacob D. Judah,1Lee-Chiang Lo,2and Geraint M. H. Thomas1
1Department of Physiology, University College London, London, United Kingdom;
and2Department of Chemistry, National Taiwan University, Taipei, Taiwan
Submitted 18 January 2005; accepted in final form 12 April 2005
Webb, Rachel J., Jacob D. Judah, Lee-Chiang Lo, and Geraint M. H. Thomas. Constitutive secretion of serum albumin requires
reversible protein tyrosine phosphorylation events in trans-Golgi.
Am J Physiol Cell Physiol 289: C748 –C756, 2005. First published
April 20, 2005; doi:10.1152/ajpcell.00019.2005.—Serum albumin se-cretion from rat hepatocytes proceeds via the constitutive pathway. Although much is known about the role of protein tyrosine phosphor-ylation in regulated secretion, nothing is known about its function in the constitutive process. Here we show that albumin secretion is inhibited by the tyrosine kinase inhibitor genistein but relatively insensitive to subtype-selective inhibitors or treatments. Secretion is also blocked in a physiologically identical manner by the tyrosine phosphatase inhibitors pervanadate and bisperoxo(1,10-phenanthro-line)-oxovanadate. Inhibition of either the kinase(s) or phosphatase(s) leads to the accumulation of albumin between the trans-Golgi and the plasma membrane, whereas the immediate precursor proalbumin builds up in a proximal compartment. The trans-Golgi marker TGN38 is rapidly dispersed under conditions that inhibit tyrosine phosphatase action, whereas the distribution of the cis-Golgi marker GM130 is insensitive to genistein or pervanadate. By using a specifically reac-tive biotinylation probe, we detected protein tyrosine phosphatases in highly purified rat liver Golgi membranes. These membranes also contain both endogenous tyrosine kinases and their substrates, indi-cating that enzymes and substrates for reversible tyrosine phosphor-ylation are normal membrane-resident components of this trafficking compartment. In the absence of perturbation of actin filaments and microtubules, we conclude that reversible protein tyrosine phosphor-ylation in the trans-Golgi network is essential for albumin secretion and propose that the constitutive secretion of albumin is in fact a regulated process.
vesicular trafficking; liver; genistein; pervanadate
PROTEIN SECRETION from mammalian cells occurs in either a
constitutive or regulated fashion and proceeds through the coordination of a series of complex and highly conserved processes (30, 37, 46). In constitutive secretion, cargo proteins are fed into the pathway by translational events at the endo-plasmic reticulum (ER) and they progress through the Golgi and then out of the cell without storage at any stage. On average, the mass of a protein secreted from cells with time is thought to be nearly constant and to depend only on the level of gene transcription. The concentrations of any constitutively secreted protein in the subcompartments of the secretory sys-tem are thought to be in near steady state (25, 34). In regulated
secretion, protein is sorted from the Golgi into inert storage granules and released when stimuli trigger their final fusion with the plasma membrane (13).
One of the major physiological functions of the liver is the constitutive secretion of serum albumin, the principal blood plasma protein by one order of magnitude. Blood albumin is replaced at a rate of⬃2% per day (40) and so by protein mass this process is the largest internal constitutive secretory process in mammals. The primary amino acid sequences, structure, and function of albumins are highly conserved across species as are the processes of albumin synthesis and secretion (42). In all cases, translation produces the short-lived, primary product preproalbumin, which is rapidly co-translationally cleaved in ER lumen, at its NH2terminus, to produce the stable precursor
proalbumin (17). Proalbumin then passes to the Golgi and is converted to mature albumin by a second NH2terminal
cleav-age (17). This process is easily monitored because proalbumin can be resolved experimentally from albumin. The cleavage occurs in a late Golgi compartment that is indistinguishable from the trans-Golgi network (TGN) and depends on Furin (38, 45). Furin and other members of the mammalian subtilisin/ Kex2p-like proprotein convertase family are localized largely in the trans-Golgi network with small amounts in the plasma membrane and recycling endosomes (28, 35) pinpointing the TGN as the crucial compartment in albumin production from proalbumin. In the absence of efficient proteolysis, e.g., be-cause of mutations in the cleavage signal, proalbumin is not trapped but is itself secreted from the cell (30) indicating that passage through the TGN is not dependent on cleavage but that it is a usual feature.
Some enzymes mediating reversible protein phosphorylation are known to play roles in secretory processes. Protein ki-nase(s) C (PKCs) were implicated in Golgi vesicle formation (9, 47, 51, 52, 61) and more recently, PKD has been shown to promote the Golgi budding induced by the ␥-subunits of heterotrimeric G proteins (15, 22, 57). Tyrosine kinases have prominent roles in some forms of regulated secretion acting in signal transduction pathways that can modulate the final exo-cytotic step (36, 41, 48). Protein tyrosine kinases and phospha-tases were not thought to influence the mechanisms of storage/ secretory granule formation preceding exocytosis but studies with inhibitors (2) revealed the action of these enzymes in trafficking between the Golgi and the plasma membrane during
Address for reprint requests and other correspondence: G. M. H. Thomas, Dept. of Physiology, Univ. College London, Rockefeller Bldg., 21 Univ. St., London WC1E 6JJ, UK (e-mail: [email protected]).
The costs of publication of this article were defrayed in part by the payment of page charges. The article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. First published April 20, 2005; doi:10.1152/ajpcell.00019.2005.
on August 31, 2009
ajpcell.physiology.org
regulated secretion. More recently, protein tyrosine phospha-tase-MEG2 has been shown to be the protein tyrosine phos-phatase associated with secretory storage granules, which al-lows maturation and fusion by regulating N-ethylmaleimide-sensitive fusion protein tyrosine phosphorylation (59, 63). In addition, studies (1, 18) of stimulated melanosome aggregation in cells from lower vertebrates indicate that protein tyrosine phosphorylation has been shown to be essential for nonsecre-tory vesicular trafficking.
Here we demonstrate that reversible protein tyrosine phos-phorylation is an essential feature of the constitutive albumin secretory pathway in rat hepatocytes, the major natural protein transport system in these cells.
MATERIALS AND METHODS
Materials. Rabbit anti-TGN38 antiserum was the generous gift of
Prof. George Banting (University of Bristol). Mouse anti-GM130 was from BD Biosciences. Both mouse anti-rabbit IgG Alexa 488 and goat anti-mouse IgG Alexa 568 were from Molecular Probes Europe (Leiden, The Netherlands). Genistein, staurosporine, PP2, and BpV-(phen) were from Calbiochem, and wortmannin, SU-6656, rapamycin, and PP1 were from Sigma. Fresh pervanadate (peroxovanadate) was prepared by treating a 10 mM stock of neutralized sodium vanadate with equimolar hydrogen peroxide. Occasionally, 0.1 mg/ml catalase (Sigma-Aldrich, Poole, UK) was added to remove any excess perox-ide and the enzyme was in turn denatured by heating. Mouse mono-clonal anti-phosphotyrosine antibody P-Tyr-100 (Cell Signaling Technologies) was from NEB (Hitchin, UK). Both goat-anti-rabbit IgG-horseradish peroxidase (HRP) and goat-anti-mouse IgG-HRP were from Bio-Rad Laboratories (Hemel Hempstead, UK). Streptavi-din-HRP conjugate and [35S]Promix ([35S]-labeled cysteine and
me-thionine) were from Amersham Pharmacia Biotech. Super Signal West chemiluminescent substrate was from Perbio Science (Tatten-hall, UK). Disodium ATP from Sigma-Aldrich was converted to the magnesium salt by sequential neutralization with Tris base (2 Mol per Mol ATP) and treatment with equimolar MgCl2.
Hepatocyte primary cell culture. Cells were prepared from the
livers of male rats (200 –300 g body wt) by perfusion of the liver with Ringer’s solution containing EDTA (2 mM). The cells were washed twice with HEPES-buffered Ringer’s solution (HBR) containing 1 mM MgCl2, 1 mM CaCl2, 4 mg/ml bovine serum albumin, and 25
mM HEPES (pH 7.4), and then purified on a discontinuous Percoll gradient. The viable cells were suspended in Williams E medium [supplemented with 5% (vol/vol) FCS, 0.04 mg/ml gentamicin, and 2 mM glutathione] at ⬃5 ⫻ 105 cells/ml. Insulin (1 mg/ml) and
hydrocortisone (0.1 mM) were added to the suspension, of which 2 ml was placed in 3.5-cm-diameter collagen-coated culture dishes or 50l on similarly treated glass coverslips. The cultures were incubated overnight and replenished with fresh medium each day if maintenance is required or washed into HBR three times and then left in a final volume of 1 ml for experimentation. For studies of albumin secretion, 3.7 MBq (100 Ci) of [35S]-labeled methionine was added to the
buffer (“pulse”). The cells were incubated for 10 min, after which they were washed with ice-cold saline containing 15 mM unlabeled me-thionine and cysteine (“chase”) and the cells transferred to similar medium containing 10g of rat serum albumin, as a carrier, along with any inhibitors. After 30 min of incubation at 37°C, the medium was removed and kept for the analysis of secreted albumin (proalbu-min is not released under these conditions). The secretion of albu(proalbu-min over this period is strictly linear with time. The cells were then washed once with fresh buffer and then lysed with the same solution contain-ing 0.5% (wt/vol) Triton X-100. The sample was centrifuged briefly, and the supernatant containing the cellular albumin and proalbumin was kept for analysis.
Analysis of albumin and proalbumin secretion. The albumin and
proalbumin in each sample was precipitated by the addition of 110l of sheep anti-rat serum albumin, centrifuged, washed twice with 150 mM NaCl, and then taken up in dilute TCA. Redundant antibody was removed by the addition of ethanol. Albumin and proalbumin were reprecipitated from the acid with diethylether, air dried, redissolved in 20l of water, and analyzed by one-dimensional isoelectric focusing [5% polyacrylamide gels containing 6 M urea, 5% (vol/vol) glycerol, and ampholines]. The gels were then air dried and phosphorimaged for quantification.
Immunocytochemistry. Cells on optical glass coverslips were
treated with indicated compounds in HBR for 10 min at 37°C. After being washed with phosphate-buffered saline (PBS), the cells were fixed with 4% (wt/vol) paraformaldehyde, permeabilized with 0.1% (wt/vol) TX-100, and then blocked with 5% (vol/vol) goat serum. The coverslips were then individually soaked in rabbit anti-rat TGN38 serum (1/200 dilution), or mouse anti-GM130. After being washed to remove the primary antibodies, rabbit IgG-Alexa 488 or anti-mouse IgG-Alexa 568 were added as appropriate and incubation continued for up to 1 h. After being washed, the cells were sealed in Mowiol on glass microscope slides. Routine confocal microscopy was carried out on a Bio-Rad Radiance imaging system fitted with a Zeiss Axiovert 100TV inverted microscope. Alexa 488 was excited with the 488-nm line of an Argon laser and the emission collected with a 520-nm (40 nm bandwidth) band-pass filter. Alexa 546 was excited at 514 nm (shoulder on the excitation spectra) and collected with a 520-nm (40-nm bandwidth) band-pass filter. Identical results could be obtained with excitation at 568 nm on a Zeiss LSM510 microscope with a⫻40 (1.3 numerical aperture) oil-immersion lens. For data analysis, images of either small groups of hepatocytes (⬃10–20 members) or individual cells could be scrutinized using MetaMorph software with qualitatively identical results. Changes in the organized, patterned distributions of Golgi staining were quanti-fied by normalizing the standard deviation of pixel intensity to average pixel intensity. The loss of a discrete staining pattern is revealed as a decrease in this index.
Preparation of rat liver Golgi membranes. In a variation of the
surgical and perfusion procedures for the isolation of hepatocytes, the livers were cleared of blood and then removed and placed in ice-cold 0.5 M sucrose. The tissue was then processed up to the gradient centrifugation step described by Taylor et al. (54). All procedures were conducted on ice and all centrifuge rotors were of the swinging-bucket type unless otherwise stated. Briefly, the tissue was chopped and then minced with a stainless steel press before two successive steps of disruption in a Dounce homgenizer. After low-speed centrif-ugation (1,000 gmax) to remove large debris and nuclei, the
micro-scopic particulate fraction was recovered from the supernantant by further centrifugation at higher speed (5,000 gmax). After resuspension
of this pellet in 250 mM sucrose buffer, the particulate fraction was floated on a discontinuous density gradient consisting of 2, 1.3, 1.1, and 0.85 M sucrose and then ultracentrifuged (200,000 gav, 2 h). The
Golgi-enriched fraction (which was clearly resolved from the ER-rich particles in the 1.3 M fraction) was recovered from the 0.85:1.1 M sucrose interface. This preparation could be snap frozen in liquid nitrogen and stored in aliquots at⫺80°C if required.
Probing Golgi membranes for tyrosine phosphatases, ATP-binding proteins, and phosphotyrosyl-containing proteins. For these
experi-ments, snap-frozen Golgi preparations were thawed and the sucrose was diluted with an equal volume of water to allow the membranes to be sedimented by centrifugation (100,000 gav, 30 min, fixed angle
rotor). Membranes were then resuspended in an assay buffer com-posed of (in mM) 20 HEPES, pH 7.5, 1 MnCl2, 1 MgCl2, and 1 DTT.
We have already established that freeze-thaw, followed by centrifu-gation in fixed-angle rotors, removes the soluble contents from the lumen of rat liver Golgi (45) providing only the membranes for analysis. The Golgi membrane preparations in assay buffer were then treated with various agents. In one set of investigations membranes
C749
GOLGI TYROSINE PHOSPHORYLATION
AJP-Cell Physiol•VOL 289 • SEPTEMBER 2005 •www.ajpcell.org
on August 31, 2009
ajpcell.physiology.org
were treated with various concentrations of MgATP in the presence or absence of 100M pervanadate and a second set of membranes were treated with 1 mM LCL2 (diluted from a 50 mM stock in water). In both cases, after 1 h at 37°C, the membranes were recovered by brief ultracentrifugation (100,000 gav, 10 min), taken up in SDS-PAGE
sample buffer, and heated (80°C for 5 min). Golgi membrane proteins were displayed on 10% SDS-PAGE gels and transferred to nitrocel-lulose. After a blocking step with bovine serum albumin or milk protein, as appropriate, the membranes were probed with either mouse monoclonal anti-phosphotyrosine IgG or streptavidin-HRP as needed. In membranes probed with anti-phosphotyrosine, a secondary goat anti-mouse HRP conjugate antibody was added after being washed. In each experiment, reactive bands were revealed by a chemiluminescent reaction. The apparent molecular weight of any reactive bands were estimated conventionally: we modeled the migration distance in relation to log10molecular weight using either first-, or occasionally,
second-order polynomial functions and regression interpolation.
RESULTS
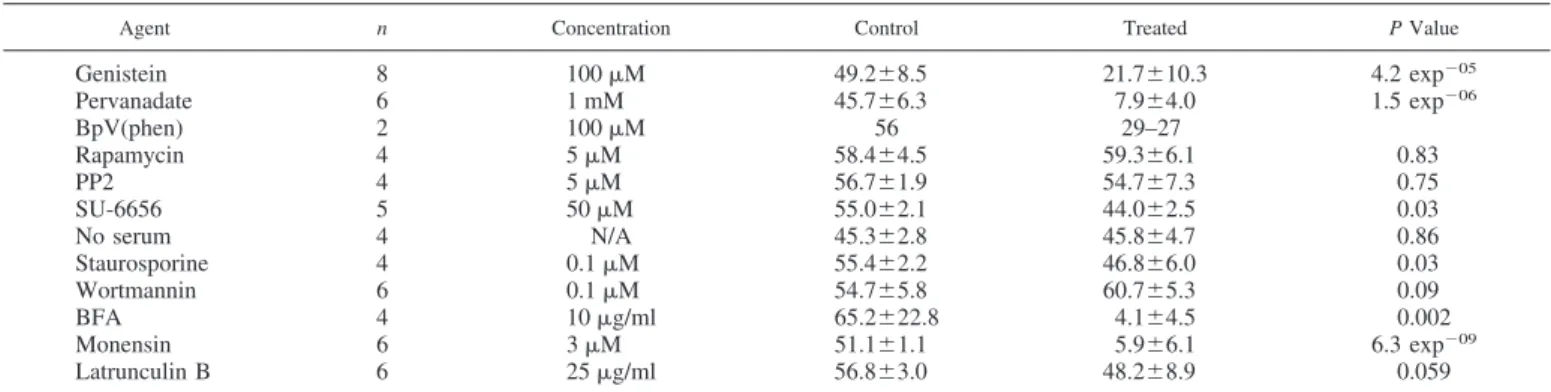
Effects of inhibitors on albumin secretion. A range of
inhib-itors was applied to primary rat hepatocytes and the constitu-tive secretion of pulse-chase-labeled albumin monitored along side untreated paired controls. Table 1 shows the impact of these compounds on the total secretion of albumin, which is in the range of 45 to 65% over 30 min in the controls. The
inhibitor concentrations used are in line with those for physi-ological or pharmacphysi-ological investigations reflecting variables like cell permeability, cellular catabolism (especially in hepa-tocytes) and high intracellular ATP concentrations (competi-tion). The established inhibitors of protein secretion, brefeldin A (BFA), and monesin, were tested as positive controls and to provide authentic blockades for comparison with the effects of other inhibitors. Genistein, a broad-spectrum inhibitor of pro-tein tyrosine kinases, caused a strong inhibition of albumin secretion across the concentration range 10 to 100M (Fig. 1, Table 1, and Fig. 2). Approximately 60% of constitutive albumin secretion could be inhibited with 50 M genistein (Table 1 and Fig. 2A). Indeed, genistein was the most potent agent among all of the protein kinase inhibitors tested (see below). Following the study by Austin and Shields (2), we tested the effects of pervanadate (peroxovanadate), an inhibitor of protein tyrosine phosphatases, and also found this com-pound to be an inhibitor of albumin secretion. More than half of albumin secretion could be eliminated with 50M pervana-date (Table 1 and Fig. 2B). Hepatocytes contain large amounts of catalase, therefore, it is unlikely that the effects of pervana-date are due to free hydrogen peroxide; however, to guard against artefacts, we used two strategies. First, pervanadate
Fig. 1. The inhibition of protein serum albumin secretion from primary rat hepatocytes by genistein. Cell cultures were established and duplicate samples treated with genistein at the concentrations indicated. After 30 min of incubation, both albumin and proalbumin were collected by immunoprecipitation from the culture medium and corresponding cell lysates. Samples were displayed by one-dimensional isoelectric focusing and migration compared with a radiolabeled authentic albumin standard (AS). Data below the figure indicate the impact of genistein on albumin secretion obtained by densitometric analysis of the phosphorimaged gels. The fraction of the total radiolabeled albumin-like protein synthesized by the hepatocytes (proalbumin⫹ albumin) that was secreted from the cells is described as %total albumin. The fraction of the mature albumin alone that was ultimately secreted is shown as %albumin.
Table 1. The effects of inhibitors on secretion of serum albumin
Agent n Concentration Control Treated P Value
Genistein 8 100M 49.2⫾8.5 21.7⫾10.3 4.2 exp⫺05 Pervanadate 6 1 mM 45.7⫾6.3 7.9⫾4.0 1.5 exp⫺06 BpV(phen) 2 100M 56 29–27 Rapamycin 4 5M 58.4⫾4.5 59.3⫾6.1 0.83 PP2 4 5M 56.7⫾1.9 54.7⫾7.3 0.75 SU-6656 5 50M 55.0⫾2.1 44.0⫾2.5 0.03 No serum 4 N/A 45.3⫾2.8 45.8⫾4.7 0.86 Staurosporine 4 0.1M 55.4⫾2.2 46.8⫾6.0 0.03 Wortmannin 6 0.1M 54.7⫾5.8 60.7⫾5.3 0.09 BFA 4 10g/ml 65.2⫾22.8 4.1⫾4.5 0.002 Monensin 6 3M 51.1⫾1.1 5.9⫾6.1 6.3 exp⫺09 Latrunculin B 6 25g/ml 56.8⫾3.0 48.2⫾8.9 0.059
Values are means⫾ SE for control incubations and with drug treatment are shown for the percent secretion of albumin hepatocytes; n, number of replicates of each experiment. BpV(phen), bisperoxo(1,10-phenanthroline)-oxovanadate; NA, not applicable; BFA, brefeldin A. Cell cultures were established and the secretion of albumin estimated, as described inMATERIALS AND METHODS. Drugs, at the various concentrations indicated, were added to the hepatocytes at the start of a 30-min albumin secretion assays without preincubation. The statistical significance of differences between paired control and experimental incubations are shown as P values from paired t-tests.
on August 31, 2009
ajpcell.physiology.org
preparations were pretreated with catalase to consume re-sidual hydrogen peroxide, and second, we examined the effects of bisperoxo(1,10-phenanthroline)-oxovanadate [BpV(phen)], another inhibitor of protein tyrosine phosphatases. In both cases, albumin secretion was inhibited.
To examine the possibility that members of the Src family of nonreceptor tyrosine kinases are the site of genistein’s action, hepatocytes were treated with either PP2 or SU-6656 (4). PP2 used at 5 M was without effect, but 50 M SU-6656 de-creased albumin secretion by up to 20%.
We also studied the effects of serum starvation to determine if surface receptor kinases or phosphatases or any of the many coupled downstream kinases might be loci for genistein’s action. We found that serum can be withdrawn for up to 20 h (the longest time tested) without any measurable effect on albumin secretion. The subsequent readdition of serum did not produce any rebound effect. Many serum factors stimulate the synthesis of 3-phosphorylated polyphosphoinositides, but there was no effect of wortmannin on albumin secretion even at high, nonspecific, concentrations (1–10M). Similar tests with LY-294002, a complimentary phosphatidylinositol 3-kinase inhibitor (4), also failed to disrupt albumin secretion from hepatocytes (data not shown).
Once we obtained inhibition of secretion with broad-spec-trum inhibitors of tyrosine kinases and phosphatases, we con-trasted the effects of genistein and pervanadate with broad-spectrum inhibitors of other classes of protein kinases. Stau-rosporine caused a relatively small inhibition of albumin secretion from hepatocytes (⬃20% of total). Staurosporine is
known to inhibit various serine/threonine kinases, including isoforms of protein kinases C (PKCs) (62), cAMP-dependent protein kinase (PKA) (44), calmodulin kinase II (65), and myosin light chain kinase (32).
Finally to exclude the possibility that we have characterized a novel, upregulated de novo radioprotein synthesis rather than the normal constitutive albumin production, we examined the effects of rapamycin and found no significant inhibition.
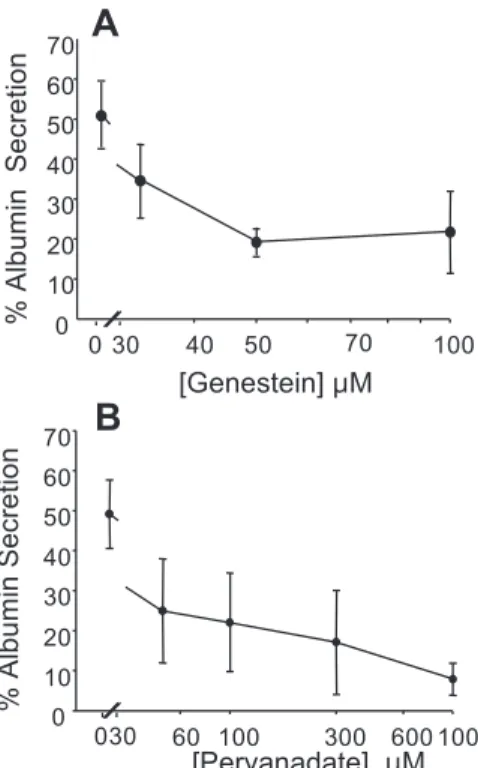
Both genistein and pervanadate inhibit constitutive secre-tory pathway at the level of trans-Golgi. Because both
genistein and pervanadate have a similar effect on the secretion of albumin, we can hypothesize that a cycle of protein phos-phorylation and dephosphos-phorylation might be important in the function of the constitutive secretory pathway. By analyzing the relationship between the observed inhibition of albumin secretion (%inhibition) and the distribution of albumin and proalbumin between the inside and the outside of the cells, we were able to establish important features of the inhibitory processes using two complimentary indexes. The first index reports the ratio of mature, cleaved albumin found inside and outside the hepatocytes (albumin in/albumin out, Fig. 3, A and
C). The second index is the ratio of mature, cleaved albumin
inside the cells to the total albumin-like protein inside the cells [the albumin-like protein is taken as the sum of albumin and proalbumin (Fig. 3, B and D)]. We also confirmed that albumin trapped inside the hepatocytes is not degraded by intracellular proteases. With the use of either index it is immediately apparent that both genistein and pervanadate cause similar changes in the distribution of the proteins between the inside and outside of the cells (compare Fig. 3A with 3C and 3B with 3D). Genistein and pervanadate increase the mass of unse-creted albumin with increasing inhibition of secretion indicat-ing a blockade after the cleavage of proalbumin to albumin, a TGN-mediated process. Proalbumin also accumulates inside the cells showing that there is no alternative pathway that bypasses the blockaded TGN. This indicates that once the export of albumin is blocked a compartment proximal to the TGN accommodates the stalled precursor pool.
Pervanadate causes redistribution of trans-Golgi marker proteins. Because the analysis of secretion indicated a site of
action for genistein and pervanadate at the level of the trans-Golgi, we investigated the effects of these compounds on the morphology of the Golgi. TGN-38 was usually found in relatively large, discrete “bodies” but this distributed to a much more highly dispersed punctate pattern in the presence of pervanadate (Fig. 4). For comparison, BFA had no significant effect on this marker and digital analysis of the change in the staining patterns with pervanadate and BFA showed a statisti-cally significantly change in both cases (P⬍ 0.05, not shown). Genistein did not affect TGN38 in six separate experiments (Fig. 4). We find GM130, a component of the Golgi scaffold always enriched in the cis-Golgi, to be distributed as discrete ribbon-like structures, reminiscent of the forms described by others (33, 55). Characteristically, treatment with BFA rapidly dissolved this pattern (Fig. 3) but no similarly gross rearrange-ment could be seen with either genistein or pervanadate. We evaluated the possibility that gross disruptions of either actin filaments or microtubule networks explain the effects of genistein or tyrosine phosphatase inhibitors on albumin secre-tion from rat hepatocytes. We find that nocodazole abolishes the extensive cytoplasmic microtubule network within our
Fig. 2. The concentration-dependent inhibition of protein serum albumin secretion from primary rat hepatocytes by genistein and pervanadate. A: effects of increasing genistein concentrations. B: effects of increasing pervanadate concentrations. Cell cultures were established and the secretion of albumin estimated exactly, as described inMATERIALS AND METHODS. Without treatment, albumin secretion is normally⬃50% within 30 min. The data shown are the mean of 3 individual experiments within which determinations were made in duplicate. Bars indicate means⫾ SE.
C751
GOLGI TYROSINE PHOSPHORYLATION
AJP-Cell Physiol•VOL 289 • SEPTEMBER 2005 •www.ajpcell.org
on August 31, 2009
ajpcell.physiology.org
hepatocytes but there was no quantifiable disruption with either 100M genistein or pervanadate (not shown). Latrunculin B eliminated the cytosolic actin filaments (mostly found imme-diately below the cell surface) and that either 100M genistein or pervanadate had any effect on these structures in side-by-side incubations (not shown). Despite the impact of latrunculin B on the hepatocellular actin distribution, this compound was without effect on albumin secretion (Table 1).
Isolated rat liver Golgi membranes contain protein tyrosine kinases, phosphatases, and corresponding substrates. To
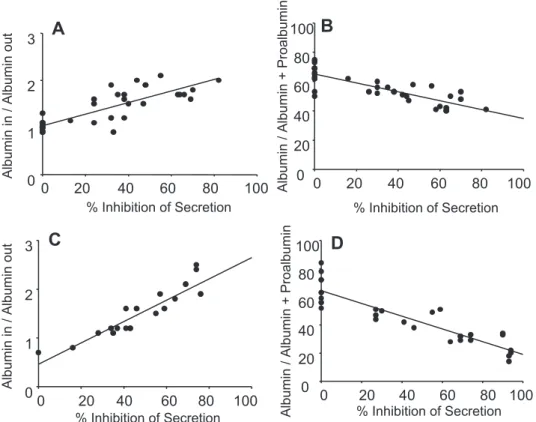
sub-stantiate the hypothesis that the machinery for a cycle of reversible protein tyrosine phosphorylation plays a role in the trafficking of albumin through the Golgi, we probed this organelle for elements of this machinery. For a cycle to be localized in the Golgi membranes, it is obligatory for at least one member of the group of cycle elements, either a tyrosine kinase, tyrosine phosphatase, or a shared protein substrate to be a resident Golgi component. Treatment of Golgi membranes individually with either ATP or pervanadate did not reveal any detectable phosphotyrosyl proteins. However, the addition of ATP in the presence of pervanadate produced a number of these species (Fig. 5A). Figure 5A also shows that by increasing the concentration of ATP over the range of 0 to 100M in the presence of pervanadate increases the amounts of detectable phosphotyrosyl proteins. Further analysis reveals apparent sat-uration kinetics for the tyrosine phosphorylation of at least 13 proteins (not shown).
To investigate further the presence of protein tyrosine phos-phatases in Golgi we used the probe LCL2 (26). This com-pound contains both a phosphotyrosyl recognition head and a trapping group (a transiently generated quinone methide) that efficiently biotinylates only proteins that hydrolyze the tyrosine phosphate group rather than merely bind to it. As a result,
LCL2 is a selective probe for active protein tyrosine phospha-tases and Fig. 5B clearly shows that Golgi membranes contain several LCL2-reactive proteins and that interaction is resistant to the blockade of nonspecific interactions with phosphate. Estimates of the apparent molecular weights range from ⬃34 to 68 KDa, a span known to contain many species of nonre-ceptor phosphotyrosine and dual-specificity phosphatases (21). DISCUSSION
The observation that both pervanadate and genistein inhibit albumin secretion from primary rat hepatocytes is surprising given the predominance of evidence for the role of tyrosine phosphorylation and dephosphorylation in regulated secretion but the complete absence of any corresponding evidence in the constitutive pathway. The postulation of a cycle of protein tyrosine phosphorylation during constitutive secretion follows immediately from these results but the precise kinases and phosphatases are not identifiable with these broad-spectrum inhibitors. The comparatively poor effects of the complimen-tary agents PP2 and SU-6656 (and PP1, data not shown) probably exclude as genistein’s targets the well-described Src family tyrosine kinases, and all coupled downstream actors (4). Because cell-surface receptors often rely on tyrosine kinase or phosphatase activity and continue to signal when endocytosed into the vesicular trafficking pathways (7), we investigated the withdrawal and readdition of serum but neither intervention affected albumin processing or secretion. With the possible exception of hepatocyte surface ligands, our cell-washing pro-tocol, combined with the strict linearity of albumin secretion mitigates against paracrine or autocrine agents unless they are secreted rapidly to response-saturating concentrations by ice-cold hepatocytes. Unlike bile production and volume
regula-Fig. 3. The effects of inhibition of protein serum albumin secretion by either genistein or pervanadate are biochemically indistinguish-able. Two different indexes were used to as-sess the nature of the effects of genistein (A and B) and pervanadate (C and D) on serum albumin from primary rat hepatocyte cultures.
A and C: fraction of total synthesized albumin
that is released from the cells after its forma-tion by proteolysis in the trans-Golgi network (TGN). B and C: efficiency with which albu-min is formed from proalbualbu-min as the ratio of albumin to the total albumin-like protein. In all cases, each point represents one individual duplicate experiment and data from up to 29 different experiments are included. In some cases data points closely overlap and this re-dundancy has been eliminated for clarity.
on August 31, 2009
ajpcell.physiology.org
tion in hepatocytes (19, 31), there is no role for class-1 phosphatidylinositol 3-kinases in the trafficking, processing, or secretion of albumin. Because the activity of these enzymes is often linked to receptor activation this also argues against some forms of receptor tyrosine kinase signaling. Because it is established that PI(3,4,5)P3and PI(3,4)P2have major roles in
the regulated plasma membrane fusion events required for hepatocyte bile secretion and volume regulation, then this lack of effect establishes a clear difference between regulated and constitutive pathways. Among the other broad-spectrum inhib-itors tested, only staurosporine, which inhibits many serine/ threonine kinases, had any effect, and this accords with earlier investigations of Golgi function (51). This might indicate a role for some permutation or combination of PKCs, PKA,
calmod-ulin kinase II, myosin light chain kinase, or even PKD. PKD is recruited to diacylglycerol containing membranes [including the TGN (27)] along with PKC-␣, -⑀, or - kinases, which phosphorylate the PKD activation loop (6, 57, 58), and which are also recruited by diacylglycerol. The activities of PKC-␣, -⑀, -, and PKD are inhibited by staurosporine (12, 16, 50) and alternative routes to PKD activation by tyrosine phosphoryla-tion (53) can be expected to be staurosporine sensitive (Src-PKC␦ path) (29) or alternatively PP2 and SU-6656 sensitive (Src-Abl pathway). Hence, the effects of staurosporine may be due to multiplicative action on several or all of these targets, but significantly, the effect of staurosporine and therefore the importance of its targets are small compared with the targets of genistein and pervanadate. Finally, because rapamycin, a rel-atively specific compound (8), had no effect then we can probably exclude changes in ribosome function mediated through ribosomal S6 kinase and other members of the rapa-mycin sensitive pathways, i.e., mitogen receptor kinases, phos-phatidylinositol 3-kinase, PDK1, and mTOR (43) from consid-eration. We also discount roles for ERKs 1/2, p38-SAPK, and JNKs 1/2. Strong activation of these kinases in primary rat hepatocytes induced with either IL-1 or IL-6 (20, 23, 24, 64) downregulating albumin expression after 24 h, but notably is without short-term effect (3, 11). Furthermore, MAP kinases require serine/threonine phosphorylation and their activation in hepatocytes can be sensitive to inhibitors of PKCs (39).
We examined the effects of genistein and pervanadate on the cleavage of proalbumin to albumin. Because cleavage occurs in the hepatocyte TGN, the sensitivity of this step can then discriminate between blockade upstream or downstream of this compartment. We find that both agents prevent the transfer of
Fig. 5. Components of cyclic protein tyrosine phosphorylation and dephos-phorylation are resident in purified Golgi membranes. Golgi membranes were isolated, as described inMATERIALS AND METHODS. A: membranes were treated 100M pervanadate and with increasing final concentrations of ATP from 0 to 100M as shown. B: membranes were left as controls or treated with combinations of 1 mM LCL2 and 50 mM phosphate, as indicated. Phosphate was included to reduce any nonspecific reactivity. In each experiment after incubation at 37°C for 60 min, the samples were treated with hot SDS-PAGE sample buffer to stop the reactions. Samples were then displayed by SDS-PAGE (10% gels) and analyzed by Western blotting with (A) antiphosphoty-rosine monoclonal antibody or (B) for biotin-containing proteins using strepta-vidin. The apparent molecular mass of reactive proteins was estimated by reference to standards included in the same gel. Prefixes pp and pLCL2 indicate phosphoprotein and LCL2-binding protein, respectively.
Fig. 4. The effects of genistein and pervanadate and other drugs on the distribution of the Golgi marker (GM) proteins TGN38 and GM130. Primary rat hepatocyte cultures on glass coverslips were treated for up to 30 min with the following drugs: control (A and B, no treatment), genistein (C and D, 100 M), pervanadate (E and F, 100 M), brefeldin A (BFA; G and H, 10 min, 5 g/ml), monensin (I and J, 10 M). Cultures were fixed, permeabilized, stained either with antibodies to TGN38 or GM130, and analyzed, as described inMATERIALS AND METHODS. The effects of the drug vehicles (ethanol 0.01% vol/vol final and DMSO 0.02% vol/vol final) had previously been found to have no effect on the distribution of the target proteins.
C753
GOLGI TYROSINE PHOSPHORYLATION
AJP-Cell Physiol•VOL 289 • SEPTEMBER 2005 •www.ajpcell.org
on August 31, 2009
ajpcell.physiology.org
the newly formed mature albumin from the TGN to the outside of the cell and that proalbumin is trapped in a proximal compartment (for examples, see Figs. 1 and 3). The common factor with both genistein and pervanadate is failure of the secretory pathway at the level of the trans-Golgi.
Hepatocytes account for ⬃93% of liver volume, and Golgi represents ⬃2% of hepatocellular membrane (5), making the rat liver one the best-described tissues for the biochemical study of Golgi function. We isolated Golgi membranes free of luminal contents (including albumin and presumably any sol-uble protein kinases, phosphatases, or substrates that might be in transit) from Golgi preparations proven to authentically catalyze the cleavage of proalbumin (45). To test our case in general (before proceeding to the specific) and to avoid bias in searches for specific proteins, we screened these membranes with nonspecific probes, particularly broad specificity an-tiphosphotyrosine antibodies to detect tyrosine kinases and endogenous substrates and with an activity probe for tyrosine phosphatases. In principle, at least one component of a cycle for protein tyrosine phosphorylation (one of either the family of tyrosine kinases, phosphatases, or suitable substrates) must be resident in the Golgi to anchor a cycle in this compartment. The absence of all three elements would allow the hypothetical cycle to be immediately dismissed. We found that isolated membranes do not contain a pool of phosphotyrosyl proteins that survives postmortem subcellular fractionation. However, because we could cause phosphotyrosyl proteins to be gener-ated by ATP in vitro but only in the presence of pervanadate then Golgi membranes must contain active systems for both the rapid generation and removal of resident phosphotyrosyl pro-teins. Phosphoprotein synthesis showed apparent saturation kinetics with ATP over the 0 –100 M range (not shown) which is significant because virtually all well-described protein tyrosine kinases have micromolar Kms for ATP. The apparent
molecular weights of some phosphoproteins are roughly in accord with elements already described by Austin and Shields (2) in regulated secretion. A probe of these same membranes for active tyrosine phosphatases revealed several candidate Golgi tyrosine phosphatases and consequently it is established that all of the machinery for reversible protein tyrosine phos-phorylation is present in rat liver Golgi membranes.
Correct Golgi function is associated with the structural and organisational integrity of this dynamic compartment. We found that pervanadate but not genistein caused major changes in the distribution of the trans-Golgi marker TGN38 and that neither compound induced any change in the localization of the
cis-Golgi marker GM130. A simple hypothesis is that the
accumulation of one or more unphosphorylated proteins stabi-lizes the TGN structure and blocks cargo exit (and also import from distal compartments), whereas the accumulation of the corresponding phosphotyrosyl protein(s) permits outward traf-fic but prevents final exit from the cell.
Gross disruptions to either actin filaments or microtubule networks might explain the effects of genistein or tyrosine phosphatase inhibitors. However, we discount genistein and pervanadate sensitivity in the actin cytoskeleton because this can be entirely dispersed with latrunculin B without significant effect on albumin secretion. This is surprising given the pro-posed role of myosins and actin in the organization and function of Golgi during protein secretion from cultured cell lines (56, 60). Microtubules are essential for the operation of
the secretory pathway providing tracks for migrating transport intermediates. Colchicine and nocodazole disrupt hepatocellu-lar microtubules and block albumin secretion (10, 14, 49), but we find no gross perturbation of microtubules with either genistein or pervanadate. This is not surprising because there is little evidence for an obligatory role for any tyrosine kinase or phosphatase in the organization of microtubules except possi-bly effects mediated through MAP (serine/threonine) kinases, which are discussed above.
In conclusion, rat liver Golgi membranes contain all of the elements for the endogenous cyclic generation of resident phosphotyrosyl proteins. Inhibition of this cycle at either one of two different points (phosphorylation or dephosphorylation) causes an identical trafficking lesion at the level of the trans-Golgi and in the case of phosphatase blockade can cause the redistribution of an established TGN marker. Despite the absence for precise targets for the effective kinase and phos-phatase inhibitors our results indicate a previously unacknowl-edged, permissive role for protein tyrosine phosphorylation at the level of the TGN during the constitutive secretion of serum albumin from the liver. Because this new role for reversible tyrosine phosphorylation is established for a large-scale, con-stitutive secretory process in primary cells then this mechanism is highly likely to be universal.
ACKNOWLEDGMENTS
We thank Ana Knezevic, Fauzia Hussain, and Safi Hussain for valuable technical assistance as part of undergraduate studies in the Molecular Cell Biology Degree Programme.
GRANTS
L.-C. Lo was supported by the National Science Council of Taiwan Grant NSC 93-3112-B-002-001 and G. M. H. Thomas was supported by a Royal Society University Research Fellowship. This study was also funded by a donation from the Gower Street Secretory Mechanisms Group.
REFERENCES
1. Aspengren S, Skold HN, Quiroga G, Martensson L, and Wallin M. Noradrenaline- and melatonin-mediated regulation of pigment aggregation in fish melanophores. Pigment Cell Res 16: 59 – 64, 2003.
2. Austin CD and Shields D. Formation of nascent secretory vesicles from the trans-Golgi network of endocrine cells is inhibited by tyrosine kinase and phosphatase inhibitors. J Cell Biol 135: 1471–1483, 1996. 3. Bader A, Rinkes IH, Closs EI, Ryan CM, Toner M, Cunningham JM,
Tompkins RG, and Yarmush ML. A stable long-term hepatocyte culture
system for studies of physiologic processes: cytokine stimulation of the acute phase response in rat and human hepatocytes. Biotechnol Prog 8: 219 –225, 1992.
4. Bain J, McLauchlan H, Elliott M, and Cohen P. The specificities of protein kinase inhibitors: an update. Biochem J 371: 199 –204, 2003. 5. Blouin A. Anatomy, ultrastructure and morphometry of the liver. In:
Plasma Protein Secretion by the Liver, edited by Glaumann H, Peters T
Jr., and Redman C. New York: Academic, 1983, p. 31–53.
6. Brose N, Betz A, and Wegmeyer H. Divergent and convergent signaling by the diacylglycerol second messenger pathway in mammals. Curr Opin
Neurobiol 14: 328 –340, 2004.
7. Burke P, Schooler K, and Wiley HS. Regulation of epidermal growth factor receptor signaling by endocytosis and intracellular trafficking. Mol
Biol Cell 12: 1897–1910, 2001.
8. Davies SP, Reddy H, Caivano M, and Cohen P. Specificity and mechanism of action of some commonly used protein kinase inhibitors.
Biochem J 351: 95–105, 2000.
9. De Matteis MA, Santini G, Kahn RA, Di Tullio G, and Luini A. Receptor and protein kinase C-mediated regulation of ARF binding to the Golgi complex. Nature 364: 818 – 821, 1993.
10. Dorling PR, Quinn PS, and Judah JD. Evidence for the coupling of biosynthesis and secretion of serum albumin in the rat. The effect of colchicine on albumin production. Biochem J 152: 341–348, 1975.
on August 31, 2009
ajpcell.physiology.org
11. Gauldie J, Sauder DN, McAdam KP, and Dinarello CA. Purified interleukin-1 (IL-1) from human monocytes stimulates acute-phase protein synthesis by rodent hepatocytes in vitro. Immunology 60: 203–207, 1987. 12. Geiges D, Meyer T, Marte B, Vanek M, Weissgerber G, Stabel S,
Pfeilschifter J, Fabbro D, and Huwiler A. Activation of protein kinase
C subtypes ␣, ␥, ␦, ⑀, , and by tumor-promoting and nontumor-promoting agents. Biochem Pharmacol 53: 865– 875, 1997.
13. Hannah MJ, Schmidt AA, and Huttner WB. Synaptic vesicle biogen-esis. Annu Rev Cell Dev Biol 15: 733–798, 1999.
14. Iwai M, Ishii Y, Tada K, Kitagawa Y, Kashiwadani M, Kashima K,
and Ibata Y. Effects of colchicine on the synthesis and transport of
albumin: a combined electron cytochemical and autoradiographic study. J
Gastroenterol Hepatol 9: 257–263, 1994.
15. Jamora C, Yamanouye N, Van Lint J, Laudenslager J, Vandenheede
JR, Faulkner DJ, and Malhotra V. G␥-mediated regulation of Golgi
organization is through the direct activation of protein kinase D. Cell 98: 59 – 68, 1999.
16. Johannes FJ, Prestle J, Dieterich S, Oberhagemann P, Link G, and
Pfizenmaier K. Characterization of activators and inhibitors of protein
kinase C. Eur J Biochem 227: 303–307, 1995.
17. Judah JD. Synthesis and secretion of serum albumin. In: Plasma Protein
Secretion by the Liver, edited by Glaumann T, Peters T Jr, and Redman C.
New York: Academic, 1983, p. 311–330.
18. Karlsson AM, Lerner MR, Unett D, Lundstrom I, and Svensson SP. Melatonin-induced organelle movement in melanophores is coupled to tyrosine phosphorylation of a high molecular weight protein. Cell Signal 12: 469 – 474, 2000.
19. Kipp H and Arias IM. Trafficking of canalicular ABC transporters in hepatocytes. Annu Rev Physiol 64: 595– 608, 2002.
20. Li D, Zimmerman TL, Thevananther S, Lee HY, Kurie JM, and
Karpen SJ. Interleukin-1 beta-mediated suppression of RXR:RAR
trans-activation of the NTCP promoter is JNK-dependent. J Biol Chem 277: 31416 –31422, 2002.
21. Li L and Dixon JE. Form, function, and regulation of protein tyrosine phosphatases and their involvement in human diseases. Semin Immunol 12: 75– 84, 2000.
22. Liljedahl M, Maeda Y, Colanzi A, Ayala I, Van Lint J, and Malhotra
V. Protein kinase D regulates the fission of cell surface destined transport
carriers from the trans-Golgi network. Cell 104: 409 – 420, 2001. 23. Ling PR, Smith RJ, Kie S, Boyce P, and Bistrian BR. Effects of protein
malnutrition on IL-6-mediated signaling in the liver and the systemic acute-phase response in rats. Am J Physiol Regul Integr Comp Physiol 287: R801–R808, 2004.
24. Ling PR, Smith RJ, Mueller C, Mao Y, and Bistrian BR. Inhibition of interleukin-6-activated Janus kinases/signal transducers and activators of transcription but not mitogen-activated protein kinase signaling in liver of endotoxin-treated rats. Crit Care Med 30: 202–211, 2002.
25. Lippincott-Schwartz J, Roberts TH, and Hirschberg K. Secretory protein trafficking and organelle dynamics in living cells. Annu Rev Cell
Dev Biol 16: 557–589, 2000.
26. Lo LC, Pang TL, Kuo CH, Chiang YL, Wang HY, and Lin JJ. Design and synthesis of class-selective activity probes for protein tyrosine phos-phatases. J Proteome Res 1: 35– 40, 2002.
27. Maeda Y, Beznoussenko GV, Van Lint J, Mironov AA, and Malhotra
V. Recruitment of protein kinase D to the trans-Golgi network via the first
cysteine-rich domain. EMBO J 20: 5982–5990, 2001.
28. Mayer G, Boileau G, and Bendayan M. The proprotein convertase furin colocalizes with caveolin-1 in the Golgi apparatus and endosomes of hepatocytes. Cell Tissue Res 316: 55– 63, 2004.
29. McGlynn E, Liebetanz J, Reutener S, Wood J, Lydon NB, Hofstetter
H, Vanek M, Meyer T, and Fabbro D. Expression and partial
charac-terization of rat protein kinase C-␦ and protein kinase C- in insect cells using recombinant baculovirus. J Cell Biochem 49: 239 –250, 1992. 30. Mellman I and Warren G. The road taken: past and future foundations
of membrane traffic. Cell 100: 99 –112, 2000.
31. Misra S, Ujhazy P, Gatmaitan Z, Varticovski L, and Arias IM. The role of phosphoinositide 3-kinase in taurocholate-induced trafficking of ATP-dependent canalicular transporters in rat liver. J Biol Chem 273: 26638 –26644, 1998.
32. Mobley PL, Hedberg K, Bonin L, Chen B, and Griffith OH. Decreased phosphorylation of four 20-kDa proteins precedes staurosporine-induced disruption of the actin/myosin cytoskeleton in rat astrocytes. Exp Cell Res 214: 55– 66, 1994.
33. Mogelsvang S, Marsh BJ, Ladinsky MS, and Howell KE. Predicting function from structure: 3D structure studies of the mammalian Golgi complex. Traffic 5: 338 –345, 2004.
34. Muth TR and Caplan MJ. Transport protein trafficking in polarized cells. Annu Rev Cell Dev Biol 19: 333–366, 2003.
35. Nakayama K. Furin: a mammalian subtilisin/Kex2p-like endoprotease involved in processing of a wide variety of precursor proteins. Biochem J 327: 625– 635, 1997.
36. Naucler C, Grinstein S, Sundler R, and Tapper H. Signaling to localized degranulation in neutrophils adherent to immune complexes.
J Leukoc Biol 71: 701–710, 2002.
37. Nickel W. The mystery of nonclassical protein secretion. Eur J Biochem 270: 2109 –2119, 2003.
38. Oda K and Ikehara Y. Monensin inhibits the conversion of proalbumin to serum albumin in cultured rat hepatocytes. Biochem Biophys Res
Commun 105: 766 –772, 1982.
39. Oliver BL, Sha’afi RI, and Hajjar JJ. Transforming growth factor-alpha increases tyrosine phosphorylation of microtubule-associated protein ki-nase in a small intestinal crypt cell line (IEC-6). Biochem J 303: 455– 460, 1994.
40. Papageorgopoulos C, Caldwell K, Shackleton C, Schweingrubber H,
and Hellerstein MK. Measuring protein synthesis by mass isotopomer
distribution analysis (MIDA). Anal Biochem 267: 1–16, 1999.
41. Parravicini V, Gadina M, Kovarova M, Odom S, Gonzalez-Espinosa
C, Furumoto Y, Saitoh S, Samelson LE, O’Shea JJ, and Rivera J. Fyn
kinase initiates complementary signals required for IgE-dependent mast cell degranulation. Nat Immun 3: 741–748, 2002.
42. Peters T Jr. Serum albumin: recent progress in the understanding of its structure and biosynthesis. Clin Chem 23: 5–12, 1977.
43. Peterson RT and Schreiber SL. Translation control: connecting mito-gens and the ribosome. Curr Biol 8: R248 –R250, 1998.
44. Prade L, Engh RA, Girod A, Kinzel V, Huber R, and Bossemeyer D. Staurosporine-induced conformational changes of cAMP-dependent pro-tein kinase catalytic subunit explain inhibitory potential. Structure 5: 1627–1637, 1997.
45. Quinn PS and Judah JD. Calcium-dependent Golgi-vesicle fusion and cathepsin B in the conversion of proalbumin into albumin in rat liver.
Biochem J 172: 301–309, 1978.
46. Rothman JE and Wieland FT. Protein sorting by transport vesicles.
Science 272: 227–234, 1996.
47. Sabatini DD, Adesnik M, Ivanov IE, and Simon JP. Mechanism of formation of post Golgi vesicles from TGN membranes: Arf-dependent coat assembly and PKC-regulated vesicle scission. Biocell 20: 287–300, 1996.
48. Sanchez-Mejorada G and Rosales C. Signal transduction by immuno-globulin Fc receptors. J Leukoc Biol 63: 521–533, 1998.
49. Saucan L and Palade GE. Differential colchicine effects on the transport of membrane and secretory proteins in rat hepatocytes in vivo: bipolar secretion of albumin. Hepatology 15: 714 –721, 1992.
50. Schaap D and Parker PJ. Expression, purification, and characterization of protein kinase C-epsilon. J Biol Chem 265: 7301–7307, 1990. 51. Simon JP, Ivanov IE, Adesnik M, and Sabatini DD. The production of
post-Golgi vesicles requires a protein kinase C-like molecule, but not its phosphorylating activity. J Cell Biol 135: 355–370, 1996.
52. Simon JP, Ivanov IE, Shopsin B, Hersh D, Adesnik M, and Sabatini
DD. The in vitro generation of post-Golgi vesicles carrying viral envelope
glycoproteins requires an ARF-like GTP-binding protein and a protein kinase C associated with the Golgi apparatus. J Biol Chem 271: 16952– 16961, 1996.
53. Storz P, Doppler H, and Toker A. Protein kinase Cdelta selectively regulates protein kinase D-dependent activation of NF-kappaB in oxida-tive stress signaling. Mol Cell Biol 24: 2614 –2626, 2004.
54. Taylor JA, Limbrick AR, Allan D, and Judah JD. Isolation of highly purified Golgi membranes from rat liver. Use of cycloheximide in vivo to remove Golgi contents. Biochim Biophys Acta 769: 171–178, 1984. 55. Trucco A, Polishchuk RS, Martella O, Di Pentima A, Fusella A, Di
Giandomenico D, San Pietro E, Beznoussenko GV, Polishchuk EV, Baldassarre M, Buccione R, Geerts WJ, Koster AJ, Burger KN, Mironov AA, and Luini A. Secretory traffic triggers the formation of
tubular continuities across Golgi sub-compartments. Nat Cell Biol 6: 1071–1081, 2004.
C755
GOLGI TYROSINE PHOSPHORYLATION
AJP-Cell Physiol•VOL 289 • SEPTEMBER 2005 •www.ajpcell.org
on August 31, 2009
ajpcell.physiology.org
56. Valderrama F, Babia T, Ayala I, Kok JW, Renau-Piqueras J, and
Egea G. Actin microfilaments are essential for the cytological positioning
and morphology of the Golgi complex. Eur J Cell Biol 76: 9 –17, 1998. 57. Van Lint J, Rykx A, Maeda Y, Vantus T, Sturany S, Malhotra V,
Vandenheede JR, and Seufferlein T. Protein kinase D: an intracellular
traffic regulator on the move. Trends Cell Biol 12: 193–200, 2002. 58. Van Lint JV, Sinnett-Smith J, and Rozengurt E. Expression and
characterization of PKD, a phorbol ester and diacylglycerol-stimulated serine protein kinase. J Biol Chem 270: 1455–1461, 1995.
59. Wang X, Huynh H, Gjorloff-Wingren A, Monosov E, Stridsberg M,
Fukuda M, and Mustelin T. Enlargement of secretory vesicles by protein
tyrosine phosphatase PTP-MEG2 in rat basophilic leukemia mast cells and Jurkat T cells. J Immunol 168: 4612– 4619, 2002.
60. Warner CL, Stewart A, Luzio JP, Steel KP, Libby RT,
Kendrick-Jones J, and Buss F. Loss of myosin VI reduces secretion and the size of
the Golgi in fibroblasts from Snell’s waltzer mice. EMBO J 22: 569 –579, 2003.
61. Westermann P, Knoblich M, Maier O, Lindschau C, and Haller H. Protein kinase C bound to the Golgi apparatus supports the formation of constitutive transport vesicles. Biochem J 320: 651– 658, 1996. 62. Wilkinson SE, Parker PJ, and Nixon JS. Isoenzyme specificity of
bisindolylmaleimides, selective inhibitors of protein kinase C. Biochem J 294: 335–337, 1993.
63. Wimmer M, Tag C, and Hofer HW. A non-receptor-type protein phosphotyrosine phosphatase is enriched in secretory vesicles of gluca.
Histochem Cell Biol 111: 135–142, 1999.
64. Yamamoto T, Kojima T, Murata M, Takano K, Go M, Chiba H, and
Sawada N. IL-1 regulates expression of Cx32, occludin, and claudin-2
of rat hepatocytes via distinct signal transduction pathways. Exp Cell Res 299: 427– 441, 2004.
65. Yanagihara N, Tachikawa E, Izumi F, Yasugawa S, Yamamoto H,
and Miyamoto E. Staurosporine: an effective inhibitor for Ca2⫹ /calmod-ulin-dependent protein kinase II. J Neurochem 56: 294 –298, 1991.
on August 31, 2009
ajpcell.physiology.org