R432 is a key residue for the multiple functions of Ndt80p in Candida albicans
Yun-Liang Yang1,2, Chih-Wei Wang3, Shiang Ning Leaw3, Te-Pin Chang3, I-Chin Wang1, Chia-Geun Chen4, Jen-Chung Fan3, Kuo-Yun Tseng3#, Szu-Hsuan Huang3, Chih-Yu Chen5,Ting-Yin Hsiao2, Chao Agnes Hsiung5, Chiung-Tong Chen6, Chwan-Deng Hsiao7 and Hsiu-Jung Lo3,8*
1Department of Biological Science and Technology, National Chiao Tung University, Hsinchu,
Taiwan, 2Institute of Molecular Medicine and Bioengineering, National Chiao Tung University, Hsinchu, Taiwan, 3National Institute of Infectious Diseases and Vaccinology, National Health Research Institutes, Miaoli, Taiwan, 4Institute of Preventive Medicine, National Defense Medical Center, Taipei, Taiwan, 5Division of Biostatistics and
Bioinformatics, Institute of Population Health Sciences, Miaoli, Taiwan, 6Division of Biotechnology and Pharmaceutical Research, National Health Research Institutes, Miaoli, Taiwan, 7Institute of Molecular Biology, Academia Sinica, Taipei, Taiwan, 8School of Dentistry, China of Medical University, Taichung, Taiwan
Running head: R432 for Ndt80p functions
Address correspondence to: Hsiu-Jung Lo, PhD,
35 Keyan Road, Zhunan Town, Miaoli County, 350, Taiwan, R.O.C.
Tel: 886 37 246 166 ext. 35516, Fax: 886 37 586 457, E-mail: [email protected]
Abstract Ndt80p is an important transcription modulator to various stress-response genes in Candida albicans, the most common human fungal pathogen in systemic infections. We found that Ndt80p directly regulated its target genes, such as YHB1, via the mid-sporulation element (MSE). Furthermore, the ndt80R432A allele, with a reduced capability to bind MSE,
failed to complement the defects caused by null mutations of NDT80. Hence, the R432 residue in the Ndt80p DNA-binding domain is involved in all tested functions, including cell separation, drug resistance, nitric oxide inactivation, germ tube formation, hyphal growth, and virulence. Hence, the importance of the R432 residue suggests a novel approach to design new antifungal drugs by blocking the interaction between Ndt80p and its targets.
Keywords: Candida albicans, Ndt80, protein-DNA interaction, MSE, Binding assay
Abbreviation: MSE, mid-sporulation element; NO, nitric oxide; CHIP, chromatin immunoprecipitation assays; CFU, colony-forming unit;
Introduction
In past two decades, the prevalence of invasive nosocomial fungal infections has increased dramatically. Candida albicans is the most common human fungal pathogen causing systemic infections, especially lethal infections in immunocompromised hosts [1-5]. One of the
effective mechanisms for the host innate immune system to defend against microbial pathogens is to produce nitric oxide (NO) [6-8]. Whereas, C. albicans responds to NO by rapidly inducing the expression of YHB1 [9, 10], encoding NO scavengers. Recently, Cta4p was reported to be a positive regulator of YHB1 [11]. Deletion of CTA4 prevented YHB1 transcription during nitrosative stress and increased sensitivity to NO [11].
Candida albicans switches between the unicellular yeast and the filamentous forms to evade unfavorable conditions. The cph1/cph1 efg1/efg1 double mutant fails to form filaments
in vitro and do not cause lethal infections in a mouse model [12, 13]. Tup1p and Nrg1p in C. albicans are negative regulators of filamentation [14, 15]. Null mutations on either one lead
to hyperfilamention and diminished virulence in vivo [14, 16]. These findings suggest that strains possessing the ability to switch between the yeast and filament forms are those
capable of penetrating vital organs and proliferating sufficiently to cause lethal infections. Ndt80p in C. albicans is involved in drug resistance by positively regulating CDR1 [17].
Great similarity is shared between the C-terminus of C. albicans Ndt80p and the N-terminus of Saccharomyces cerevisiae Ndt80p, a novel DNA-binding domain targeting the
mid-sporulation element (MSE, gNCRCAAAA/T) [18]. In contrast, C. albicans Ndt80p does not share the same activation domain with S. cerevisiae Ndt80p [17, 19], suggesting that these two proteins are associated with distinct functions. Recently, Ndt80p has been reported
to regulate different sets of genes in response to different stresses in C. albicans [20]. The present study demonstrated that an R432A substitution blocked the functions of Ndt80p in cell separation, drug resistance, NO inactivation, germ tube formation, hyphal growth, and virulence. Hence, the R432 residue in the DNA-binding domain of Ndt80p is important for pathogenesis in C. albicans and its vicinity a potential target for drug development. These findings prompted us to employ virtual screening approach to identifying compounds blocking the interactions between Ndt80p of C. albicans and its targets.
Materials and methods
Strains, media, and primers
The Candida albicans strains used in the present study are listed in Table 1. Yeast Peptone Dextrose (YPD, 1% yeast extract, 2% peptone, and 2% dextrose), and Synthetic Dextrose (SD, 0.67% yeast nitrogen base without amino acid and 2% dextrose) were prepared as described [21]. Cells were grown in either YPD or SD unless otherwise noted. The
compounds for addition to media were from Difco unless otherwise noted. The primers used in the present study are listed in Table 2.
RNA isolation
To avoid filamentous growth, saturated overnight cultures of the strains were diluted in YP containing 4% glucose [13] pH 6.0 to a starting optical density at 600 nm (OD600) of 0.2 at
concentration of 10 mM when the cell density reached OD600 of 0.9. Then the cells were
harvested after additional incubation at 37ºC for 30 minutes. For real-time PCR, total RNA was isolated from the pellets by glass bead beating in the presence of hot acid phenol pH 4.0 (Sigma, P-4682) and RNA lysis buffer (50 mM NaOAc, 10 mM EDTA, 1% SDS). For
microarray assay, total RNA was isolated using the mechanical disruption method provided in the RNeasy Midi Kit (Qiagen 75144) according to the recommendation of the manufacturer. An off-column DNase digestion (Baseline-ZERO DNase, Epicentre biotechnologies,
Madison, WI, US) was carried out to ensure complete removal of DNA during the RNA isolation. The RNA quality was measured by Agilent 2100 Bioanalyzer (Agilent
Technologies, Santa Clara, CA, US).
DNA microarray analysis
A total of 25 μg RNAs were directly labeled with Cy3-dUTP or Cy5-dUTP (PerkinElmer Life Science, Waltham, MA, US) using a LabelStarTM Array Kit (Qiagen 28904) according to the manufacturer’s instructions. Incorporation of Cy3-dUTP or Cy5-dUTP was performed during the reverse transcription. The labeling efficiencies for the cDNA were measured using a NanoDropTM 1000 Spectrophotometer (Thermal Scientific, Wilmington, DE, US) before microarray hybridization. The DNA microarray slides were kindly provided by Dr. S. Rupp at the Fraunhofer Institute for Interfacial Engineering and Biotechnology, Germany. The
procedure for prehybridization, hybridization, and wash has been previously described [22]. Finally, the slides were scanned by the GenePix 4000B scanner (Molecular Devices,
Sunnyvale, CA, US). Data were acquired and analyzed by GenePix Pro6.0 (Molecular Devices, Sunnyvale, CA, US).
Comparison of gene expressions in the presence of NaNO2 in the wild-type (SC5314)
versus the ndt80/ndt80 null mutant (YLO133) consisted of two biological replica. One set of replica was also subjected to a technical duplication with dye-swap. To deal with the systemic dye effect, we adapted quantile normalization, which enforced all distributions of the red and green dye intensities to be as similar as possible. From the normalized intensities, we
performed significant analysis [1] to identify the candidate genes in each experiment. The genes with at least 2-fold difference in expression levels between the wild-type and
ndt80/ndt80 cells are listed in Table 3. For the promoter analysis, it was considered to be a
plausible MSE when a sequence matched 8 out of the 9 nucleotides of MSE.
Quantitative analysis of the mRNA level by real-time PCR
A real-time PCR was performedin a Rotor-Gene™ 3000 instrument (Corbett Research, Mortlake, Australia) with a TITANIUM ™ Taq PCR kit (BD Clontech, 639210) and
SYBR®Green I Nucleic Acid Stain (Cambrex, 50513) todetermine the level of mRNA. The sample setup was processed automatically by CAS-1200™ (Corbett Research, Mortlake, Australia). Real-time PCR was performedaccording to the instructions from the manufacturer and the expressions of ACT1 and/or TEF3 in each strain was used as loading controls. The level of mRNA isolated from the wild-type cells in the absence of NaNO2was defined as one
and those from different strains were normalized accordingly.
The assay of NO-mediated growth inhibition and concentrations of NO was modified from a previous report [10]. Freshly made NaNO2 (Sigma, SI-S2252) was added to the culture at time zero. Cell growth was monitored after incubation at 37ºC for 8 hours. Viable cell counts were determined by plate counting on yeast peptone dextrose (YPD) agar medium. The colony-forming unit (CFU) of each strain in the absence of the NaNO2was defined as 100
and those from the same strain grown in various concentrations of the NaNO2 were
normalized accordingly.
Mouse model for virulence
The experiments on the mouse model for virulence were conducted as reported [12]. Briefly, BALB/c mice (white, male) from National Laboratory Animal Center in Taiwan weighing between 18 and 20 g were used to test the virulence of different Candida strains. Each Candida strain was tested for virulence by injecting 0.5 ml of the cell suspension into the tail vein of the mouse (approximately 1 x 106 cells per mouse). Numbers of mice injected with the wild-type cells, the ndt8/ndt80 mutant cells, the ndt80ndt80::NDT80 cells, and the
ndt80/ndt80::ndt80R432A cells were 17, 17, 9, and 8, respectively. The mice were monitored daily for their survival. These studies were carried out in accordance with the NIH Guide to the Care and Use of Laboratory Animals and the Animal Welfare Act in an
AAALAC-accredited program. The animal study protocol for experimental procedures used in the present study was approved by the Institutional Animal Care and Use Committee at the National Health Research Institutes.
Construction of histidine-HA-tagged Ndt80p
Construction of pSFS2A-6H3HA (LOB248): Primers HJL1110 containing a sequence of three tandem repeats from hemagglutinin (3×HA, underlined) and HJL977 were used to amplify a 463 base-pair (bp) fragment from the pSFS2A vector [23]. The 463 bp fragment was further amplified by primers HJL1111 containing a sequence of six histidines and
HJL977 to gain a 529 bp fragment that harbors sequences of two newly introduced restriction sites, BglII and SphI, as well as six histidines and 3×HAs followed by the ACT1 terminator. The 529 bp fragment was cloned to the pSFS2A vector to yield pSFS2A-6H3HA (LOB248).
Construction of pSFS2A-ENO1p-NDT80-6H3HA (LOB302): The ADE2 fragment amplified by HJL1197 and HJL1198 was cloned into the LOB248 plasmid to generate LOB298. The DNA fragment from the 3’UTR region of ADE2 was amplified using primers HJL1199 and HJL1200. The ENO1 promoter region was amplified using primers
HJL1201and HJL1202. The above two DNA fragments were fused by mixing them together as the template and re-amplification with primers HJL1199 and HJL1202 to generate a 1348 bp fragment which was cloned into the LOB298 plasmid to obtain LOB301. The NDT80 gene was amplified using primers HJL1254 and HJL1082 and then cloned into the LOB301 vector to yield the LOB302 construct containing NDT80-HA. Then, the LOB303 was constructed to contain ndt80R432A-HA.
Gene disruption of NDT80 using the SAT1 flipper cassette was performed according to previously described methods [23]. Briefly, a fragment containing NDT80 upstream
sequences from positions −445 to −322 with respect to the translation start codon was amplified with the primers HJL147 and HJL148, and a fragment containing NDT80 downstream sequences from positions +1806 to +2120 was amplified with the primers
HJL716 and HJL717. The upstream and downstream fragments of NDT80 were cloned on either side of the SAT1 flipper cassette in plasmid pSFS2A to result in LOB227. The codon region of NDT80 was replaced by the SAT1 flipper through a double cross-over to generate the YLO386 strain. To tag Ndt80p with histidine-HA via the genomic locus, we applied SacI restriction digestion to LOB302 and LOB303 plasmids and then introduced the resultant DNAs into YLO386 separately to generate YLO387 (ndt80/ndt80::NDT80-HA) and YLO388 (ndt80/ndt80::ndt80R432A-HA), respectively. The DNA fragment containing a full length NDT80 along with its promoter region was amplified by primers HJL1081 and HJL1123. The
DNA fragment was cloned into pGEMT-easy vector and then sub-cloned into LOB227 by
KpnI and XhoI digestion to result in the LOB387. We applied KpnI and SacI restriction
digestion to LOB387 plasmid and then introduced the resultant DNAs into YLO386 to generate YLO464 (ndt80/ndt80::NDT80).
Chromatin immunoprecipitation
The C. albicans cells were treated with freshly prepared 10 mM NaNO2 at 37°C for 30 minutes before being harvested. Cells were cross-linked with 1% formaldehyde (Merck, 1.04003.100) and then lysed in lysis buffer (50 mM HEPES, pH 7.5, 500 mM NaCl, 1 mM EDTA, pH 8.0, 0.1% Triton X-100, 0.1% sodium dodecyl sulfate, 0.1% sodium deoxycholate) supplemented with a protease inhibitor cocktail (ROCHE, 04 693 132 001), using the
FastPrep-24 instrument (MP Biomedicals, Illkirch, France) at a speed of 6 m/s for 40 s, four times with cooling intervals. The chromatin was sheared to an average size of 500 bp by a Bioruptor (Diagenode, Liege, Belgium). Portions of the input sample (50 μl) was retained as the genomic DNA control and the remaining (500 μl) was incubated with 2 μg of HA
antibody (Abcam, ab9110), rotated at 4ºC overnight, and subsequently incubated with 100 μl of protein A agarose (Upstate, 16-156) at 4ºC for 4 hours. Agarose beads were then washed and the DNA was eluted by TES buffer (50 mM Tris-HCl, pH 8.0, 10 mM EDTA, 0.1% SDS). Cross-linking was reversed by incubating the DNA solution at 65ºC overnight. Finally,
proteins were removed by the phenol/chloroform/isoamyl alcohol method and DNA precipitation was facilitated by adding 20 μg of glycogen (Invitrogen, 10814-010). To normalize the PCR efficiency for different primers, the amount of promoter DNA amplified from 25 ng of genomic DNA was defined as one and those from the CHIP resultants were normalized accordingly. Ndt80p should not associate with the ADE2 promoter in vivo.
Therefore, the value of the ADE2 DNA fragments in each immunoprecipitation was the result of non-specific background and was used for secondary normalization to determine the ratio of change for particular promoters from the same strain.
Electrophoretic mobility shift assays
Nuclear protein was extracted as described previously [24]. Briefly, spheroplasts were prepared by incubating cells in 20 ml of YPD with 1.1 M sorbitol containing 400 U/ml lyticase (Sigma, L4025) for 30 minutes. Crude nuclei were extracted by douncing the
spheroplast in breaking buffer (18% Ficoll, 1% thiodiglycol, 5mM Tris-HCl, pH 7.4, 20 mM KCl, 2 mM EDTA-KOH, pH 7.4, 0.125 mM spermidine, 0.05 mM spermine) for 20 times and subsequently centrifuging at 5000 xg twice for 12 minutes each at 4ºC to remove non-spheroplasting cells. Finally, crude nuclei were harvested by centrifugation at 24000 xg for 15 minutes at 4ºC followed by suspension in ice-cold buffer B (100 mM Tris-acetate, pH 7.9, 50 mM potassium acetate, 10 mM MgSO4, 20% glycerol, 2 mM EDTA-KOH, 3 mM
DTT, 1x protease inhibitor cocktail), while adding (NH4)2SO4 to a final concentration of 0.9 M before being stirred for 30 minutes at 4ºC. The samples were then centrifuged at 215000 xg for 30 minutes. The supernatant was recovered and desired amount of (NH4)2SO4 was added to attain 75% saturation. The pellet containing nuclear proteins were harvested, dissolved and dialyzed in buffer C (20 mM HEPES, pH 7.6, 10 mM MgSO4, 10 mM EGTA, 20% glycerol, 5 mM DTT, 0.5 mM PMSF).
Gel shift assay was performed by the electrophoretic mobility shift assay “Gel Shift” Kit (Panomics, Fremont, CA, USA) according to the manual. Briefly, 4 μg of total nuclear
proteins or 0.3 μg of purified N, C-terminally tagged Ndt80221-529-hexa-His from E. coli along with 1 μg Poly d(I-C) was incubated in 1x binding buffer at room temperature for 5 minutes. The sample was then combined with the biotin-labeled double-stranded MSE probe (200 fmol) and incubated at 15ºC for 30 minutes. DNA-protein complexes were then resolved by electrophoresis on a 6% native polyacrylamide gel and transferred to the membrane
(GeneScreen Plus® Hybridization Transfer Membrane, PerkinElmer, NEF988001PK). A DNA fragment containing the putative YHB1 MSE (GAA TAT TAC ACA AAA CCG CGG AA) was used as the probe. The shifted bands were detected by the HRP-conjugated Streptavidin. The unlabelled MSE probe or a single base substituted mutant probe (GAA TAT TAC AgA AAA CCG CGG AA) was used to perform competition analyses.
Construction and purification of Ndt80p in Escherichia coli
The N, C-terminally tagged Ndt80221-529-hexa-His plasmids were constructed by cloning a PCR-generated fragment containing the Ndt80 DNA-binding domain, using HJL1619 and HJL1620 primers, into the EcoRI and XhoI sites of pET42a (Novagen). Protein was
expressed in C43 E. coli cells (Novagen), and positive clones were checked by PCR,
restriction analysis and Western blot analysis using anti-hexa-His antibodies (Sigma). Protein expression was induced by the addition of IPTG. Crude extracts were filtered through 0.45 μm and passed over a Ni-NTA column (GE Healthcare). The protein was eluted with an imidazole gradient ranging from 50 to 400 mM. Further purification was carried out with GSTrap (GE Healthcare) with PBS pH 8.0 containing 10 mM glutathione. The proteins in the collected fractions were concentrated with Centricons (Millipore), dialyzed against PBS, and digested by thrombin before further purification by gel-filtration (Superdex 200)
chromatography. The resulting protein was concentrated to 6.8 mg/ml in a buffer containing 10 mM Tris pH 7.8, 100 mM NaCl, and 1 mM DTT.
Results
Target genes of Ndt80p
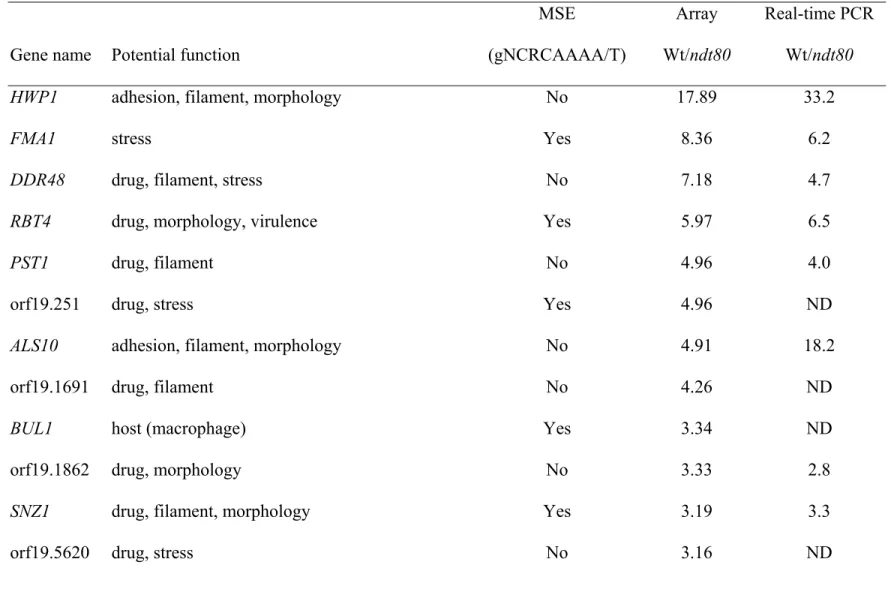
A total of 40 potential targets of Ndt80p (Table 3) were identified by expression profiling based on DNA microarray analyses. The promoters of 22 of them contained a plausible MSE
site. These 40 genes have one or multiple potential functions. There were 3 genes involved in adhesion, 3 in virulence, 4 in host interaction, 13 in filamentous growth, 16 in morphology, and 27 in stress responses (including drug and nitric oxide) response. The expressions of 16 of those 40 potential targets were verified by real-time PCR (Table 3).
In the present study, we found that null mutations in NDT80 reduced the expression of YHB1 (Fig. 1A, bar 1 vs. bar 3), which is consistent with the previous report [20]. Nevertheless, this expression was not completely abolished in the ndt80/ndt80 mutant cells because Ndt80p is not the sole positive regulator of YHB1. As expected, the level of YHB1 expression was restored in the mutants by introducing a wild-type NDT80 allele (Fig. 1A, bar 3 vs. bar 4).
We then sought to investigate the biological effect of the mutated Ndt80p on the NO sensitivity. The ndt80/ndt80 null mutant was more sensitive to NaNO2 than the wild-type was,
a behavior shared with the yhb1/yhb1 mutant (Fig. 1B). A wild-type NDT80 allele restored the growth of the ndt80/ndt80 null mutant (Fig. 1B, ndt80/ndt80::NDT80), indicating that the increased sensitivity was indeed due to the NDT80 knockout. All strains were killed when the concentrations of the NaNO2were higher than 30 mM (Fig. 1B). Interestingly, after NaNO2
treatment, the ndt80/ndt80 mutant cells form as few colonies as yhb1/yhb1 mutant cells did even though the expression of YHB1 in the ndt80/ndt80 mutant cells was approximately half of that in the wild-type cells. The observation that yhb1/YHB1 heterozygous mutant cells have similar NO sensitivity as the wild-type cells [10] rules out the possibility that the sensitivity to NO in ndt80/ndt80 mutant cells solely resulted from 50% reduction of YHB1 expression. Hence, down-regulations of other Ndt80p target genes involved in NO
inactivation, such as IFG3, orf19.1724, and orf19.4690, may also contribute to the sensitivity to NO in ndt80/ndt80 mutant cells.
Ndt80p is important for cell separation, germ tube formation, and hyphal growth
Here we further determined whether Ndt80p was also involved in other functions. The
wild-type cells formed germ tubes in liquid media containing serum (Fig. 2Ba). In contrast,
the ndt80/ndt80 mutant cells formed a chain of cells similar to pseudohyphae and failed to
form germ tubes in the presence of serum (Fig. 2Bb). As expected, the ndt80/ndt80 mutant cells (Fig. 2Cb) were also defective in hyphal growth. A copy of the wild-type NDT80allele restored the ability to form germ-tube (Fig. 2Bc) and hyphae (Fig. 2Cc) of the ndt80/ndt80 mutant cells. Our results demonstrate that Ndt80p is critical for cell separation, germ tube formation, and hyphal growth.
Mutations on NDT80 diminish the virulence in a mouse model
Then, we investigated whether mutations on NDT80 diminish the virulence in a mouse model of systemic infections. After one week, all mice injected with 1 x 106 wild-type cells died as expected (Fig. 3, solid squares). In contrast, all mice injected with the ndt80/ndt80 mutant cells still survived (Fig. 3, open squares). A copy of the wild-type NDT80 allele restored the virulence to the mutant cells (Fig. 3, solid triangles).
Ndt80p binds DNA with MSE
To investigate whether Ndt80p in C. albicans can bind DNA, we constructed strains expressing histidine-HA-tagged Ndt80p (Ndt80-HAp, Fig. 4A, lane 3) in the wild-type SC5314 background for immunoprecipitation against the HA tag. First, we conducted chromatin immunoprecipitation assays (CHIP) to enrich the chromatin DNA bound by Ndt80-HAp. Then, we performed PCR on the resulted DNA (Fig. 4B). The DNA fragments carrying the YHB1 promoters were amplified (Fig. 4B, upper panel, lane 8). In contrast, those
carrying the ADE2 promoters without MSE were not detected (Fig. 4B, bottom panel, lane 8). To further determine the binding efficiency of Ndt80p to MSE, we performed real-time PCR on the resulted DNA from the CHIP experiment to quantify DNA fragments carrying the promoters of various genes (Fig. 4C). As expected, the DNA of the YHB1 promoter was enriched significantly by Ndt80-HAp (Fig. 4C, bar 2 vs. bar 3). The data also showed that the binding of Ndt80p to the YHB1 promoter was sequence-specific since the DNA fragment localized approximately 1.6-1.9 kbp upstream of YHB1 was not enriched by Ndt80-HAp (Fig. 4C, bar 6 vs. bar 7). Furthermore, Ndt80-HAp also enriched other promoters with MSE, such as FMA1, a potential Ndt80p target gene identified by the DNA microarray analysis (Fig. 4C, bar 10 vs. bar 11), but not those without, such as HWP1 (Fig. 4C, bar 14 vs. bar 15).
The electrophoretic mobility shift assays (EMSA) further supported the notion that MSE was the binding site of Ndt80p. When the labeled oligonucleotides containing MSE
sequences were added to the nuclear extract from ndt80/ndt80::NDT80-HA, additional bands were present (Fig. 4D, indicated by the arrow marked protein-DNA). As the amount of the wild-type unlabeled probes increased, the intensity of the bands faded (Fig. 4D, lane 3 vs. lanes 4-6). In contrast, when the unlabeled mutant probe, with a base substitution in MSE, was used to compete with the wild-type labeled probes, the effect on the band intensity could be neglected (Fig. 4D, lane 3 vs. lanes 7-9). Hence, the interaction between Ndt80p and MSE could be titrated out by excessive amount of the unlabeled wild-type probes but not by the mutant ones. To assess whether Ndt80p directly binds MSE instead of being mediated by additional factors in the nuclear extract, we then performed EMSA with recombinant Ndt80p purified from E. coli. Our results showed that purified recombinant Ndt80p from E. coli could bind MSE directly (Fig. 4E, lane 2). Again, the interaction between Ndt80p and MSE could be titrated out by excessive amount of the unlabeled wild-type probes (Fig. 4E, lane 2
vs. lanes 5-6) but not by the mutated probes (Fig. 4E, lane 2 vs. lanes 9-10).
The R432 residue is important for all functions of Ndt80p
The residues responsible for DNA binding capability of Ndt80p in S. cerevisiae have been reported. Among them, the R177 residue is the most important one. When this R177 residue is mutated, the DNA binding capability decreases 27-foldin S. cerevisiae [25]. In a sequence comparison, the R432 residue in C. albicans corresponds to the R177 residue in S. cerevisiae. We recently found that the ndt80R432A allele, containing a mutation on the DNA-binding
domain, failed to complement the drug sensitive phenotype caused by null mutations on
NDT80 in C. albicans [19]. Therefore, in the present study, we investigated whether R432A
substitution also affected other functions of Ndt80p. The ndt80R432A allele, unlike the
wild-type, failed to restore to the ndt80/ndt80 mutant cells the YHB1 expression (Fig. 1A, bar 3 vs. bar 5), NaNO2 sensitivity (Fig. 1B, ndt80/ndt80::ndt80R432A), cell separation (Fig. 2Ad),
germ tube formation (Fig. 2Bd), hyphal growth (Fig. 2Cd) and virulence in a mouse model (Fig. 3, open triangles).
According to the reported crystal structure of the Ndt80p DNA-binding domain of S.
cerevisiae, we modeled the three dimensional structure of Ndt80p DNA-binding domain of C. albicans using Phyre 2. The core structure of both proteins forms a β-sandwich with two
antiparallel β sheets that consist of a three-stranded sheet and a four-stranded sheet. The strands in these two sheets are approximately perpendicular to the direction of those found in the β-sandwich. Besides that, the other structure comprises a continuous polypeptide chain with two short α-helices, helical turns, and also loop regions. Most importantly, like the R117 residue in Ndt80p DNA-binding domain of S. cerevisiae, the R432 in C. albicans is predicted
to interact with DNA directly (Fig. 5). Additionally, the R432A substitution reduced the binding efficiency to MSE (Fig. 4D, lane 3 vs. lane 10), consistent with the CHIP result (Figs. 4B and 4C).
Discussion
Candida albicans may cause invasive infections when there are major dysfunctions in the host’s cellular defense mechanisms. The two most well-defined mechanisms by which C.
albicans can breach innate host defenses are the production of NO scavengers and the ability
to penetrate macrophages by formation of hyphae. In the present study, we found that the DNA-binding domain of Ndt80p binds directly to MSE to regulate the expression of target genes involved in multiple signal transduction pathways in C. albicans. Most importantly, a single R432A substitution on the DNA-binding domain abolished all functions examined.
Initially, the ndt80/ndt80 mutant cells generated from BWP17 were used for microarray and NO sensitivity assay. Since we have to construct new strains for the experiments in the present study, we chose SC5314 as the parental strain since the SAT1 flipper cassette became available. Using the newly constructed strains, we found that Ndt80p bound to various promoters with diverse biological functions, consistent with the previous report [26]. Hence,
C. albicans Ndt80p is an upstream modulator of the gene networks that respond to a variety
of environmental conditions. To cope with stress such as exposures to antifungal drugs and NO, Ndt80p regulates different sets of genes, such as CDR1, ERGs [17, 26], and YHB1. It is interesting to determine whether Ndt80p regulates the basal and/or induced expressions of various targets.
such as YHB1 and CHT3, were also identified in the present study while others were not. Notably Sellam et al did not find differences in the cell wall gene HWP1, which is the most highly down-regulated gene in the present study. We believe that the differences in the listed genes are due to different growth conditions. Cells were grown in YPD at 30ºC by Sellam et al, but in YP containing 4% glucose pH 6.0 with 10 mM NaNO2 at 37ºC in the present study.
Furthermore, cells were grown in the presence of NaNO2 instead of azole drugs.
Subsequently, CDR1 was not identified in the present study.
The eukaryotic nature of fungi reduces the number of potential antifungal targets. Hence, the limited choices of clinically active antifungal drugs and the emerging drug resistance are major issues in managing fungal infections. Our findings further support that the Ndt80p of C.
albicans recognizes MSE just as Ndt80p of S. cerevisiae does [26]. Furthermore, the R432A
substitution in the Ndt80p DNA-binding domain blocks its functions in cell separation, drug resistance, NO inactivation, germ tube formation, hyphal growth, and virulence in C.
albicans, suggesting a novel target for new antifungal drugs.
Acknowledgements We thank Drs. G. Fink, M. Gustin, and A. Mitchell for strains and plasmids. We acknowledge Dr. S. Rupp for DNA microarray slides and Mss. H. I. Shih, C. L. Su, and F. H. Yang for their technical assistance. We thank Drs. J. J. Chiu, Hsiao, C. D., T. S. Huang, J. L. Juang, S. F. Lin, W. S. Lo, S. C. Lee, F. Murad, S. C. Teng, L. Wang, and L. H. Wang for their helpful suggestions. We would like to thank Drs. X. Chen, I. G. Ho, K. Kunin, and T. L. Lauderdale for their critical review of the manuscript. This work was supported in part by grants 98-3112-B-400-006 and 99-2320-B-400-006-MY3 from the National Science Council (NSC) and ID-100-PP-09 from the National Health Research Institutes to HJL, as well as by grants 98-3112-B-009-001 and 99-2320-B-009-001-MY3 from NSC, 99W962
from the ATU program of National Chiao Tung University to YLY.
References
1. Tusher VG, Tibshirani R,Chu G (2001) Significance analysis of microarrays applied to the ionizing radiation response. Proc Natl Acad Sci U S A 98:5116-5121.
2. Cheng MF, Yu KW, Tang RB, Fan YH, Yang YL, Hsieh KS, Ho M,Lo HJ (2004) Distribution and antifungal susceptibility of Candida species causing candidemia from 1996 to 1999. Diagn Microbiol Infect Dis 48:33-37.
3. Pappas PG, Rex JH, Lee J, Hamill RJ, Larsen RA, Powderly W, Kauffman CA,
Hyslop N, Mangino JE, Chapman S, Horowitz HW, Edwards JE,Dismukes WE (2003) A prospective observational study of candidemia: epidemiology, therapy, and
influences on mortality in hospitalized adult and pediatric patients. Clin Infect Dis 37:634-643.
4. Yang YL, Ho YA, Cheng HH, Ho M,Lo HJ (2004) Susceptibilities of Candida species to amphotericin B and fluconazole: the emergence of fluconazole resistance in
Candida tropicalis. Infect Control Hosp Epidemiol 25:60-64.
5. Yang YL, Li SY, Cheng HH,Lo HJ (2005) Susceptibilities to amphotericin B and fluconazole of Candida species in TSARY 2002. Diagn Microbiol Infect Dis 51:179-183.
6. Blasi E, Pitzurra L, Puliti M, Chimienti AR, Mazzolla R, Barluzzi R,Bistoni F (1995) Differential susceptibility of yeast and hyphal forms of Candida albicans to
macrophage-derived nitrogen-containing compounds. Infect Immun 63:1806-1809. 7. MacMicking J, Xie QW,Nathan C (1997) Nitric oxide and macrophage function.
Annu Rev Immunol 15:323-350.
8. Rementeria A, Garcia-Tobalina R,Sevilla MJ (1995) Nitric oxide-dependent killing of
Candida albicans by murine peritoneal cells during an experimental infection. FEMS
Immunol Med Microbiol 11:157-162.
9. Hromatka BS, Noble SM,Johnson AD (2005) Transcriptional response of Candida
albicans to nitric oxide and the role of the YHB1 gene in nitrosative stress and
virulence. Mol Biol Cell 16:4814-4826.
10. Ullmann BD, Myers H, Chiranand W, Lazzell AL, Zhao Q, Vega LA, Lopez-Ribot JL, Gardner PR,Gustin MC (2004) Inducible defense mechanism against nitric oxide in
Candida albicans. Eukaryot Cell 3:715-723.
11. Chiranand W, McLeod I, Zhou H, Lynn JJ, Vega LA, Myers H, Yates JR, 3rd, Lorenz MC,Gustin MC (2008) CTA4 transcription factor mediates induction of nitrosative stress response in Candida albicans. Eukaryot Cell 7:268-278.
12. Chen CG, Yang YL, Cheng HH, Su CL, Huang SF, Chen CT, Liu YT, Su IJ,Lo HJ (2006) Non-lethal Candida albicans cph1/cph1 efg1/efg1 transcription factor mutant establishing restricted zone of infection in a mouse model of systemic infection. Int J Immunopathol Pharmacol 19:561-565.
13. Lo HJ, Kohler JR, DiDomenico B, Loebenberg D, Cacciapuoti A,Fink GR (1997) Nonfilamentous C. albicans mutants are avirulent. Cell 90:939-949.
14. Murad AM, d'Enfert C, Gaillardin C, Tournu H, Tekaia F, Talibi D, Marechal D, Marchais V, Cottin J,Brown AJ (2001) Transcript profiling in Candida albicans reveals new cellular functions for the transcriptional repressors CaTup1, CaMig1 and
CaNrg1. Mol Microbiol 42:981-993.
Talibi D, Marechal D, Tekaia F, d'Enfert C, Gaillardin C, Odds FC,Brown AJ (2001)
NRG1 represses yeast-hypha morphogenesis and hypha-specific gene expression in Candida albicans. EMBO J 20:4742-4752.
16. Braun BR,Johnson AD (1997) Control of filament formation in Candida albicans by the transcriptional repressor TUP1. Science 277:105-109.
17. Chen CG, Yang YL, Shih HI, Su CL,Lo HJ (2004) CaNdt80 is involved in drug resistance in Candida albicans by regulating CDR1. Antimicrob Agents Chemother 48:4505-4512.
18. Chu S,Herskowitz I (1998) Gametogenesis in yeast is regulated by a transcriptional cascade dependent on Ndt80. Mol Cell 1:685-696.
19. Wang JS, Yang YL, Wu CG, Ouyang KJ, Tseng KY, Chen CG, Wang H,Lo HJ (2006) The DNA binding domain of CaNdt80p is required to activate CDR1 involved in drug resistance in Candida albicans. J Med Microbiol 55:1403-1411.
20. Sellam A, Askew C, Epp E, Tebbji F, Mullick A, Whiteway M,Nantel A (2010) Role of transcription factor CaNdt80p in cell separation, hyphal growth, and virulence in
Candida albicans. Eukaryot Cell 9:634-644.
21. Sherman F (2002) Getting started with yeast. Methods Enzymol 350:3-41.
22. Sohn K, Senyurek I, Fertey J, Konigsdorfer A, Joffroy C, Hauser N, Zelt G, Brunner H,Rupp S (2006) An in vitro assay to study the transcriptional response during adherence of Candida albicans to different human epithelia. FEMS Yeast Research 6:1085-1093.
23. Reuss O, Vik A, Kolter R,Morschhauser J (2004) The SAT1 flipper, an optimized tool for gene disruption in Candida albicans. Gene 341:119-127.
biology, part C. Methods Enzymol 351:190-192.
25. Montano SP, Cote ML, Fingerman I, Pierce M, Vershon AK,Georgiadis MM (2002) Crystal structure of the DNA-binding domain from Ndt80, a transcriptional activator required for meiosis in yeast. Proc Natl Acad Sci U S A 99:14041-14046.
26. Sellam A, Tebbji F,Nantel A (2009) Role of Ndt80p in sterol metabolism regulation and azole resistance in Candida albicans. Eukaryot Cell 8:1174-1183.
27. Gillum AM, Tsay EY,Kirsch DR (1984) Isolation of the Candida albicans gene for orotidine-5'-phosphate decarboxylase by complementation of S. cerevisiae ura3 and E.
Figure legends
Fig. 1 The level of YHB1 mRNA and nitric oxide inactivation. A, Real-time PCR was applied
to measure the mRNA level. Values on the vertical axis represent the relative mRNA expression level of YHB1 genes in various strains grown in the presence of 10 mM NaNO2. Bar 1, wild-type cells (SC5314); bar 2, yhb1/yhb1 mutant cells (YLO270); bar 3, ndt80/ndt80 mutant cells (YLO133); bar 4 ndt80/ndt80::NDT80 cells (YLO137), and bar 5,
ndt80/ndt80::ndt80R432A cells (YLO263). B, NO sensitivity assay. Values on the horizontal axis represent the concentrations of NaNO2 and those on the vertical axis represent relative colony-forming units per ml.
Fig. 2 Assays for cell separation, germ tube formation, and hyphal growth. Four strains, including (a) wild-type (SC5314), (b) ndt80/ndt80 (YLO386), and (c) ndt80/ndt80::NDT80 (YLO464), and (d) ndt80/ndt80::ndt80 R432A (YLO388 ) were studied. A, Cell separation.
Cells were grown on agar YPD medium at 30ºC overnight. B, Germ tube formations. Cells were grown in liquid YPD medium containing 10% fetal bovine serum at 37ºC for 3 hours. C, Hyphal growth. Cells were grown on agar medium containing 4% fetal bovine serum at 37ºC for 4 days. The scale bar is 50 um.
Fig. 3 Virulence assay in a mouse model. The BALB/c mice were injected with
approximately 1 X 106 cells of the wild-type (SC5314, ■), ndt80/ndt80 (YLO 386, □),
ndt80/ndt80::NDT80 (YLO464 , ▲), or ndt80/ndt80::ndt80R432A-HA (YLO388, △).
(SC5314, lane 1), ndt80/ndt80 (YLO386, lane 2), ndt80/ndt80::NDT80-HA (YLO387, lane 3), and ndt80/ndt80::ndt80R432A-HA (YLO388, lane 4) was determined by western blot analyses. The lower bands represent the loading control histone H4. B, DNA fragments of the YHB1 and ADE2 promoters from different strains after CHIP were amplified by PCR. Lanes 2, 4, 6, and 9, input genomic DNA alone; lanes 3, 5, 8, and 11, CHIP in the presence of HA antibody; and lanes 7 and 10, CHIP in the presence of IgG non-specific antibody. C, DNA fragments of various promoters from different strains after CHIP were amplified by real-time PCR. YHB1 (lanes 1-4), YHB1 upstream: 1.6-1.9 kbp upstream of YHB1 (lanes 5-8), FMA1 (lanes 9-12), and HWP1 (lanes 13-16). D, Ndt80p interacting with MSE. The labeled probes were
incubated in the absence (lane 1) or presence of nuclear extracts from ndt80/ndt80 (A, YLO386, lane 2), ndt80/ndt80::NDT80-HA (B, YLO387, lanes 3 to 9), and
ndt80/ndt80::ndt80R432A-HA (C, YLO388, lane10) in the absence of unlabeled probe (lanes
1-3 and 10), presence of unlabeled probes (lanes 4-6), or unlabeled mutant probes (lanes 7-9). E, Direct interaction between recombinant Ndt80p from E. coli and the MSE sequences. The labeled probes were incubated in the absence (lane 1) or presence (lanes 2-10) of recombinant Ndt80p from E. coli, in the absence of unlabeled probes (lanes 1-2), presence of unlabeled probes (lanes 3-6), or unlabeled mutant probes (lanes 7-10).
Fig 5 Predicated three dimensional structures of Ndt80p DNA-binding domains. A, Crystal structure of the Ndt80p DNA-binding domain in S. cerevisiae (PDB ID: 1mnn). B, Predicted structure of the Ndt80p DNA-binding domain in C. albicans based on the structure of the homologue of S. cerevisiae. The R432 residue of C. albicans and its corresponding residue in
Figure 1A
Figure 2A
Figure 2B
Figure 4A
Figure 4C
Table 1 Candida albicans strains used in the present study
Strain Genotype Comment Reference
SC5314 Wild-type [27]
YLO133 ura3∆::λimm434/ura3∆::λimm434 his1::hisG/his1::hisG
arg4::hisG/arg4::hisG
ndt80::GFP-Arg4/ndt80::URA3-dpl200
ENO1/eno1::ENO1-tetR-ScHAP4-3xHA-HIS1
ndt80/ndt80 [17]
YLO137 ura3∆::λimm434/ura3∆::λimm434 his1::hisG/his1::hisG
arg4::hisG/arg4::hisG
ndt80::GFP-Arg4/ndt80::URA3-dpl200::NDT80-HIS1
ndt80/ndt80::NDT80 [17]
YLO263 ura3∆::λimm434/ura3∆::λimm434 his1::hisG/his1::hisG
arg4::hisG/ arg4::hisG
ndt80::GFP-ARG4/ndt80::URA3-dpl200::ndt80R432A-HIS1
ndt80/ndt80::ndt80R432A [19]
YLO270 ura3∆::λimm434/ ura3∆::λimm434 his1::hisG/ his1::hisG
yhb1::HIS1/yhb1::URA3
yhb1∆/yhb1∆ [10]
YLO386 ndt80∆-FRT/ ndt80∆-FRT ndt80/ndt80 the present study
YLO387 ndt80∆-FRT/ ndt80∆-FRT::NDT80-HA-FRT ndt80/ndt80::NDT80-HA the present study
YLO388 ndt80∆-FRT/ ndt80∆-FRT::ndt80R432A-HA-FRT ndt80/ndt80::ndt80R432A-HA the present study
Table 2 Primers used in the present study
name sequence comment
HJL147 cgg cgg tac cAG GAG ATA TCA TAT CGT Ta NDT80 fragment
HJL148 cgt tct cga gCT GTC TGG ACT TTG TGA NDT80 fragment
HJL319 AGA GTT GCC TGA CCA C real-time PCR for NDT80
HJL320 ATC TGC AAG TCC TCG T real-time PCR for NDT80
HJL687 ACT AGA GCT TTC GGC A real-time PCR for YHB1
HJL688 AGG ATC GTA AGG ACC AA real-time PCR for YHB1
HJL716 ttt tcc gcg gTT GAT GAT GAC GAA GAA AGG NDT80 fragment
HJL717 tgt cga gct cGG GTC ATA CTT CAG CAA TG NDT80 fragment
HJL 1081 ggtaccTTGTGCCCGAAATAAACCTC cloning for full length NDT80
HJL1082 ggg ccc CTG TGG AGG AGT AGG GGT TG NDT80 gene
HJL1110 GGA TAC GAC GTA CCA GAT TAC GCT TAC GAC GTA CCA GAT TAC GCT TAC GAC GTA CCA GAT TAC GCT TAA gag tga aat tct gga aat ctg ga
HJL1111 GGT ACC GGG CCC CCC CTC GAG AGA TCT GCC GCA gca tgc GGA TGC CAC CAC CAC CAC CAC CAC GGT GGA TAC GAC GTA CCA GAT TAC GCT
Histidine-HA-tagged protein
HJL 1123 ctcgagCCAGTACTTGACGAGTCACT cloning for full length NDT80
HJL1174 ACT CCT CCA TCA TTC ATT CAT TCT CHIP for YHB1
HJL1175 TAT GGA AAT CAT GGG GAA GTA AAT CHIP for YHB1
HJL1195 GGG TCA CGT GAT GGT ATT TAA TTT CHIP for ADE2
HJL1196 ATG CGT GTG TGA TGA GAA TAA GAT) CHIP for ADE2
HJL1197 CTA ACC CCT CAA TAG CAT TGA TTC ADE2 fragment
HJL1197 CTA ACC CCT CAA TAG CAT TGA TTC ADE2 fragment
HJL1199 ggt acc gcg gag ctC TAC ATT AGA TGG GGT TGA TT ADE2 fragment HJL1200 GCG ACA CCA CTA AGA AAC TAG
AGA TGT GGT GGA TTG GTA TTT CTT TCT GTG
HJL1201 CAC AGA AAG AAA TAC CAA TCC ACC ACA TCT CTA GTT TCT TAG TGG TGT CGC
ENO1 fragment
HJL1202 GGT ACC GTG GTG GTG GTG GTG GTG CAT TGT TGT AAT ATT CCT GAA TTA
ENO1 fragment
HJL1230 ATT AGA ATC AGT TCC ACT CAT GCA real-time PCR for HWP1
HJL1231 ATA CCA ATA ATA GCA GCA CCG AAA real-time PCR for HWP1
HJL1234 ATC ATT ACC ATC AGG ATC GA real-time PCR for IFG3
HJL1235 TCA CCC CAA CTT AGA ACA TT real-time PCR for IFG3
HJL1254
ggg ccc AAT CAA ACT CTT GTC AGA AT NDT80 gene
HJL1265 CGG TAA GAG GTC ATC GTC ATA C real-time PCR for RTA2
HJL1266 TCA GCC AAT TCT GCC ACT C real-time PCR for RTA2
HJL1271 GGT TAT TGG CTT CAA AGA CA real-time PCR for GIT1
HJL1277 CTT CGT CGA CAA GTT TAT CTT C real-time PCR for CHT3
HJL1278 CAT CTG GAA TAG TAG CAA CAG T real-time PCR for CHT3
HJL1301 TCG ATG ATG CCG ATT TGT TA real-time PCR for orf19.1862
HJL1302 TCG ATG ATG CCG ATT TGT TA real-time PCR for orf19.1862
HJL1303 TTA TCA AGT TGC CGA AAC AT real-time PCR for PST1
HJL1304 GCA GGG AAA TTA CCA AAT CT real-time PCR for PST1
HJL1305 TGG TAG TGG ACA TGG CGA TT real-time PCR for ASR1
HJL1306 TCT TTG TGT GCT TCC TTT TGC real-time PCR for ASR1
HJL1307 TGA GAA GAA TCA ACG AAG GT real-time PCR for SNZ1
HJL1308 CAA CAA AAA CAC CAT CAC AT real-time PCR for SNZ1
HJL1340 GGT TTG GTT ACT TTA GCG TTT C real-time PCR for orf19.308
HJL1341 TGT GAA TGT GGG TGT GAT ATT T real-time PCR for orf19.308
HJL1348 TTG CTA ATG CTG GTG TGT TG real-time PCR for FMA1
HJL1349 TGA CTT TAC CGT TGG TTT TC real-time PCR for FMA1
HJL1373 CAA AAA ACG TGC TAG ACA TG real-time PCR for RBT4
HJL1375 GAA AAT GAA ACC ATA GCA ACA A real-time PCR for ALS10
HJL1376 AGC ATT ACC ACC ACC ATT ATT A real-time PCR for ALS10
HJL1393 ACA CTG ACA GTT ACG GTT CT real-time PCR for DDR48
HJL1394 CGT ATG AAT CAT CAT TTG AA real-time PCR for DDR48
HJL1397 TGG CCT GTT TTC TAA AGC CAA GT CHIP for FMA1
HJL1398 CTG CGC TAT ACG GGT TTC C CHIP for FMA1
HJL1409 TCG GAA ATT CTG ACG ATA AAT G CHIP for HWP1
HJL1410 GCG GAT ACA GGT GAT ACA A CHIP for HWP1
HJL1417 AGG CTA TGA TCA ATG GAA AA real-time PCR for SOU1
HJL1418 TGG CTA AAT GAG TAC ATG CA real-time PCR for SOU1
HJL1452 TTT CGG CAA CTT TTT CGC CHIP for SNZ1
HJL1453 TCT AAG CCT TTA CAC GTC TC CHIP for SNZ1
HJL1454 AAC CAG AGT GGA GAA TGT CHIP for YHB1
HJL1455 CAA TGT TGG CTT AAT GAA CTT T CHIP for YHB1
HJL1619 gaa ttc TTG GCT CAG CAA CA NDT80 binding domain fragment
Table 3 Genes positively regulated by Ndt80p in Candida albicans
MSE Array Real-time PCR
Gene name Potential function (gNCRCAAAA/T) Wt/ndt80 Wt/ndt80
HWP1 adhesion, filament, morphology No 17.89 33.2
FMA1 stress Yes 8.36 6.2
DDR48 drug, filament, stress No 7.18 4.7
RBT4 drug, morphology, virulence Yes 5.97 6.5
PST1 drug, filament No 4.96 4.0
orf19.251 drug, stress Yes 4.96 ND
ALS10 adhesion, filament, morphology No 4.91 18.2
orf19.1691 drug, filament No 4.26 ND
BUL1 host (macrophage) Yes 3.34 ND
orf19.1862 drug, morphology No 3.33 2.8
SNZ1 drug, filament, morphology Yes 3.19 3.3
GIT1 drug, filament, host (macrophage) Yes 3.02 3.2
PGA23 drug Yes 3.00 ND
FAD3 drug, stress No 2.98 ND
MUM2 filament, morphology, stress No 2.92 ND
IFG3 nitric oxide (oxidase), filament Yes 2.84 2.0
CSP37 adhesion, virulence Yes 2.82 ND
HGT20 glucose transporter Yes 2.81 ND
CIS2 filament Yes 2.80 ND
PRN4 morphology No 2.70 ND
YHB1 drug, morphology, nitric oxide, virulence Yes 2.59 2.1
FRP6 host (neutrophils), morphology Yes 2.58 ND
orf19.4043 stress Yes 2.56 ND
DUT1 stress No 2.52 ND
orf19.4216 drug, filament, morphology Yes 2.51 ND
orf19.6840 drug Yes 2.48 ND
orf19.1152 morphology, stress Yes 2.45 ND
NIP1 host (macrophage) No 2.38 ND
RTA2 drug, stress No 2.37 2.8
orf19.4690 nitric oxide Yes 2.37 ND
CHT3 filament, stress No 2.31 2.4
orf19.2125 morphology No 2.26 ND
orf19.308 morphology Yes 2.25 1.8
orf19.7350 drug, stress No 2.23 ND
orf19.2496 drug Yes 2.20 ND
ASR1 drug, morphology, stress No 2.14 1.8
SOU1 drug, filament, morphology Yes 2.12 2.6
PLB4.5 morphology Yes 2.11 ND