利用突變策略、抑制作用與SELEX技術針對氧化鯊烯環化酵素進行功能性之探討
270
0
0
全文
(2) 利用突變策略、抑制作用與 SELEX 技術針 對氧化鯊烯環化酵素進行功能性之探討 Functional Characterization of Oxidosqualene Cyclases Using Mutagenesis Approaches, Inhibition Studies, and SELEX Technology. 研究生:張程翔. Student: Cheng-Hsiang Chang. 指導教授:吳東昆 博士. Advisor: Prof. Tung-Kung Wu Ph.D. 國 立 交 通 大 學 生物科技研究所 博士論文 A Dissertation Submitted to Department of Biological Science and Technology College of Biological Science and Technology National Chiao Tung University in partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy in Biological Science and Technology Hsinchu, Taiwan, Republic of China. July, 2008. 中華民國九十七年七月.
(3) 摘要. 氧化鯊烯環化酵素(EC 5.4.99-) 為一群具有催化受質 (3S)-2,3-氧化鯊烯 ((3S)-2,3-oxidosqulaene)進行一系列的環化與重組反應之同源酵素。在絕大多數的 物種中,氧化鯊烯環化酵素會負責專一性的生物轉化作用,進而生成多樣的固醇 類或三萜類產物。受質所進行的環化機制與催化酵素本身結構間所產生的協同關 係更是極度複雜與引人注目的。為了闡明受質-氧化鯊烯在機制上所進行的環化 與重組反應,我們設計了許多實驗,包括分子生物學上的突變策略以及利用生物 有機化學的方式來加以探討。 在一開始,我們利用“丙氨酸掃描式突變” 以及“定點突變” 的方式,並配 合酵母菌體內的“質體交換篩選法” ,來加以確定在酵母菌中氧化鯊烯環化酵素 (S.c.ERG7) 其內部對於催化反應或結構穩定上重要的氨基酸位置。由實驗的結 果發現,在我們所感興趣的酥氨酸-509 到異白氨酸-513 的這段區域上,酪氨酸 -510 在突變效應的影響下產生非常迥異的催化活性。當酪氨酸-510 置換成色氨 酸或離氨酸時,酵母菌的轉殖株便無法補充其體內本身的氧化鯊烯環化酵素上的 缺失而導致菌株的死亡。但在換成丙氨酸時,酵母菌的突變株卻是可以存活。因 此,我們進行了各轉殖株其體內非皂化酯醇的分離與鑑定,結果發現酵母菌氧化 鯊烯環化酵素酪氨酸-510 的丙氨酸突變株 (S. cerevisiae ERG7Tyr510Ala) 可以產生 單環的蓍醇 A (achilleol A)、四環的羊毛硬脂醇 (lanosterol) 以及 parkeol。而在致 死的色氨酸或離氨酸的突變株中,酵母菌僅能產生兩個單環的產物分別為蓍醇 A (achilleol A)或 camelliol C。為了更進一步的瞭解在不同的氨基酸改變下對於氧化 鯊烯環化酵素酪氨酸-510 所造成突變效應,我們繼而利用“定點飽和突變” 的方 式來進行探討。其結果發現,不同的酪氨酸-510 突變株會產生單環、三環以及多 種四環的生合成產物。此外,在原先所推測對於產物專一性非常重要的“催化氨 基酸鹼性殘基對” (catalytic base-dyad),即酪氨酸-510 與組氨酸-234 所形成的共 i.
(4) 伴效應,可以藉由不同的酪氨酸-510 與組氨酸-234 的雙點突變來加以闡明。另 一方面,我們亦利用類似的定點飽和突變結合生物有機化學的特性,發現更多對 於在不同階段的環化與重組反應中相當重要的一些氨基酸位置,並分別鑑定出多 個中間態產物的結構。為了仔細地瞭解這些氨基酸在酵素活性區域中的重要性, 我們也建構了一系列酵母菌氧化鯊烯環化酵素的蛋白質同源模擬結構,進而合理 地解釋這些關鍵性的氨基酸位置對於其單離產物間的關係。之後,我們亦利用量 子力學的高斯軟體 Gaussian 03 來預測在不同的酵素突變株中,不同的產物其在 理論能階上的相對應位置與其產生的趨勢。另一方面,我們將酵母菌中氧化鯊烯 -羊毛硬脂醇環化酵素與阿拉伯芥中氧化鯊烯-環阿屯醇環化酵素進行一系列嵌 合體酵素資料庫 (chimeric enzyme library) 的建構,期望能找出在這兩個酵素中 專門與產物專一性有關的特定區域。因此,我們一共設計了十種不同的嵌合體 (chimeras),並藉由“質體交換篩選法” 來分析它們在酵母菌中的活性表現。但 在所有的嵌合體轉殖菌株中,其體內的非皂化酯醇並沒有明顯的差異,這也許意 味著過於粗糙的切割可能造成整體酵素在結構上的瓦解。之後,我們進一步地比 較之前在三萜類合成酵素中,其嵌合體酵素資料庫的實驗數據後,我們認為在固 醇類生合成酵素中,產物的專一性也可能僅來自於酵素活性區域上少數幾個關鍵 的功能性氨基酸。 除了分子生物學外,我們也同步進行了不同的生物化學實驗,並且將重點放 在哺乳類動物中氧化鯊烯-羊毛硬脂醇環化酵素其結構與功能間的關係。我們主 要是利用管柱層析、生物有機化學及不同的抑制劑修飾等實驗,對於牛肝中的氧 化鯊烯環化酵素進行一系列的探討。在一開始,我們成功地由牛肝中將氧化鯊烯 -羊毛硬脂醇環化酵素純化出來,並藉由串聯式質譜儀加以鑑定。之後,我們也 順利地將整段序列解讀出來並將其表現在酵母菌中。而由氨基酸的比較上,我們 發現牛肝中的氧化鯊烯環化酵素與其他三種哺乳類動物的氧化鯊烯環化酵素在 序列上有百分之八十以上的相同度。然而,為了更進一步的瞭解其中一個非常有 效的抗氧化鯊烯環化酵素之特性抑制劑-Ro48-8071 其作用機制與其明確的鍵結 ii.
(5) 位置,我們進行了光親和性的分析與設計了五個以 Ro48-8071 為骨架的螢光性衍 生物。然而,由抑制酵素活性的實驗與螢光光譜特性上,我們發現化學螢光修飾 會大幅減弱 Ro48-8071 的抑制活性。另外,我們利用分子入塢實驗(molecular docking) 也合理的解釋經修飾後的螢光化合物在氧化鯊烯環化酵素的活性區域 中,對於酵素立體空間所產生的障礙關係以及經此修飾對於抑制能力的影響。希 望未來能更進一步的研發新型式的 Ro48-8071 衍生型螢光抑制劑,除了能幫助氧 化鯊烯環化酵素在結構與功能性質上進行探討,也能進一步提供在蛋白質體學上 的應用,或對於在降膽固醇藥物的篩選上有所幫助。另一方面,我們也利用了一 個隨機選擇性的組合式核苷酸資料庫篩選法-SELEX 對於氧化鯊烯環化酵素進 行篩選,希望選擇出對其具有一定結合能力的最適體 (aptamer)。經過九次的篩 選循環,我們成功的得到了多個最適體分子。針對這些最適體分子,我們除了證 實了它們與牛肝中氧化鯊烯環化酵素的結合能力外並計算出其大約在 nM 範圍 的親和力。然而最適體與氧化鯊烯環化酵素間的作用方式仍需更進一步的試驗來 加以瞭解。期望這些與氧化鯊烯環化酵素有結合能力的分子,可以在未來不論是 針對羊毛硬脂醇環化酵素或是針對膽固醇的生合成途徑皆能提供在醫療或診斷 上的應用。 綜合以上所述,我們利用了不同的分子生物學實驗,包括丙氨酸掃描式突 變、定點直接/飽和突變、區域置換實驗、同源模擬與量子力學等生物資訊的研 究得以針對氧化鯊烯環化酵素其環化與重組反應在機制上有更深入的瞭解。另外 一方面,藉由生物化學與生物有機化學的相互運用,我們更獲得許多寶貴的資 訊。特別是在哺乳動物其氧化鯊烯環化酵素的取得,以及利用化學性修飾而得以 針對抑制劑 Ro48-8071 其抑制作用有更進一步的瞭解。同時利用體外 SELEX 的 實驗,更幫助我們對於未來針對合理地設計抗真菌類或降低膽固醇藥物的領域上 開創一條展新的大道。. iii.
(6) Abstract Oxidosqualene cyclases (EC 5.4.99-) constitute a family of enzymes that catalyze diverse cyclization/rearrangement reactions of (3S)-2,3-oxidosqualene (OS) into a distinct array of sterols and triterpenes. Notably, the product specificity among most of cyclase enzymes is species-dependent. The relationship between cyclization mechanism and enzymatic structure is extremely complex and attractive. In order to further elucidate the cyclization/rearrangement reaction of oxidosqualene cyclases, experiments including different molecular biological mutagenesis approaches as well as the bioorganic studies were accordingly carried out. First, the alanine-scanning mutagenesis and site-directed mutagenesis coupled with in vivo plasmid shuffling selection were employed to identify the catalytic or structural important residues in oxidosqualene-lanosterol cyclase from Saccharomyces cerevisiae (S. cerevisiae ERG7). Among the investigated sequence segment from Thr-509 to Ile-513, Tyr-510 showed the catalytic discrepancy in the cyclase activity upon mutagenic effect. The yeast transformant failed to complement the cyclase-deficiency when this position was mutated to tryptophan or lysine residues, but still maintained the yeast viability in the S. cerevisiae ERG7Y510A mutant. After analysis of the nonsaponifiable lipid from S. cerevisiae ERG7Y510A, the monocyclic achilleol A, tetracyclic lanosterol and parkeol were identified. Moreover, two monocyclic compounds, achilleol A and camelliol C, were isolated from the lethal S. cerevisiae ERG7Y510K and S. cerevisiae ERG7Y510W mutants. In order to further investigate the mutated effects on this residue, the site-saturated mutagenesis was subsequently performed. Diverse products including monocyclic, tricyclic, and different tetracyclic products were isolated from the S. cerevisiae ERG7Y510X mutants. Moreover, the inherent influence on product specificity via an altered coordinative interaction between the hypothesized catalytic dyad, Tyr-510 and His-234, were further examined in more detail via construction and analysis of a different set of S. cerevisiae ERG7H234X/Y510X double mutations. Moreover, other catalytically important. iv.
(7) residues and their respective premature cyclization products, involved in different cyclization/rearrangement stages, were also discovered by using similarly executed site-saturated mutagenesis, coupled with bioorganic characterization. In order to carefully explore the importance of these crucial residues within the enzymatic active site, plausible homology modeling structures were subsequently created. The diverse array of product profiles which were isolated from various mutated S. cerevisiae ERG7 cyclases was broadly representative. Moreover, the products’ tendency in different mutated enzymes was consequently understood by using the quantum mechanics calculation of Gaussian 03. In addition, the combination of chimeric enzyme library between Saccharomyces cerevisiae lanosterol synthase and Arabidopsis thaliana cycloartenol synthase was also constructed to determine the critical functional domain responsible for the product specificity. Ten diverse domain swapping chimeras were successfully created, and their activities were subsequently confirmed via plasmid shuffling selection. No divergence of the nonsaponifiable lipid patterns was observed among these inactive chimeras, suggesting that the rough partition might disrupt the enzyme structure. After comparison with the previous experimental results from the triterpene synthases, the product specificity-determining residues among these sterol-biosynthetic cyclases might be determined by just several functional crucial residues within the enzymatic active site. In parallel to the ongoing molecular biology approaches, we also performed a number of biochemical studies, including chromatographic purification, bioorganic characterization, and inhibition studies to examine the structure-function relationships for mammalian lanosterol cyclase. After successful purification and tandem mass characterization of bovine liver OSC, the gene encoding bovine liver OSC was subsequently determined. The deduced amino acid sequence showed >80% identity to that of the other three mammalian lanosterol synthases. The bovine liver OSC gene was also successfully cloned and functionally expressed in a yeast erg7 disruption strain. Moreover, in order to better understand the inhibiting mechanism of one potent OSC inhibitor, Ro48-8071, as well as to solve the exact inhibitor binding site, the photoaffinity labeling and chemical fluorescent modification of Ro48-8071 was also v.
(8) carried out. Several Ro48-8071-based fluorescent probes were developed and their inhibitory activity or fluorescence characteristics were analyzed. The results of chemical modification of Ro48-8071 suggested that the fluorescent Ro48-8071 derivatives dramatically reduced its inhibitory activity for purified bovine liver OSC. Moreover, the interactions between fluorescent Ro48-8071 derivative and the active site of bovine liver OSC, as well as the orientation of these probes have obviously changed, based on the molecular docking experimental data. In the future, improved site-specific fluorescent probes should be developed and applied to the chemical proteomic field for effectively screening the OSC drugs. By another approach, a randomly selected combinatorial approach, SELEX, was utilized for screening the potential OSC-binding aptamer molecules of the bovine liver OSC. After nine rounds of SELEX screening, a diverse array of aptamer candidates was isolated from the single-strand DNA library. These aptamer molecules exhibited the definitive interaction with the targeted protein and also revealed the approximate nM range affinity for bovine liver OSC. However, the binding interaction between individual aptamers and the cyclase protein should be explored in the future. These obtained OSC-binding aptamers will be applied in the pharmaceutical or diagnostic applications for lanosterol synthase as well as for studies in the cholesterol pathway in the future. Thus, the combination of results obtained from the molecular biological approaches, including alanine-scanning, site-directed/saturated mutagenesis, domain swapping experiments, homology modeling structure, and quantum mechanics calculation, a better understanding of the cyclization/rearrangement mechanism of oxidosqualene. cyclase. has. been. achieved.. In. addition,. the. biochemical. characterization coupled with the bioorganic studies toward mammalian lanosterol synthase provided valuable information, especially in obtaining the purified cyclase protein, and illustrating the inhibition mechanism of Ro48-8071. Moreover, the in vitro SELEX procedure opens new avenues in rational designing of antifungal agents and hypocholesteremic agents.. vi.
(9) 謝誌 (Acknowledgement) 每個人的一生就像一道道雷射光束般,雖然顏色、能量、強度都不盡相同,但在兩道光束 交會時,卻會依倆人的緣份而綻放出繽紛的火花。宏觀地來看,雖然人與人的際遇就應像網狀結 構般的雜亂無章。但,我卻在這近十年期間,與原本素昧平身的一群人,在交通大學生物科技學 系吳東昆老師所領導的生物有機與分子演化實驗室內,交織出一道道最美妙的光輝。. 而這一切將會是我最難抹滅的回憶!. 首先,最要感謝的是我的指導教授-吳東昆博士,願意在八年前收留一個當時正躊躇不安的 大學畢業生進入他的實驗室。雖然來不及參與實驗室的草創,但我卻何其有幸地可以一路看著它 成長茁壯,至今已成為人人稱羡的實驗室。常常看著滿屋子的實驗設備思考著,有著幾近外面數 間實驗室總合的我們,倘若還是無法做出好的研究成果,那就真的太對不起一直為實驗室默默地 付出的吳老師了。跟隨老師共同學習的這段期間,讓我感受到的是老師長期以來對於學術的熱忱 與堅持。另外,他在實驗設計上的嚴謹與其對實驗結果的辯證和思維,也令我受益良多。每每困 頓怠墮時,老師時而的當頭棒喝,時而的經驗分享與砥礪,總能不知不覺地在我心中興起一股繼 續向上的動力,不斷地激勵我繼續堅持。另外,老師在面對事情時,所表現出來的積極態度與不 畏懼挫折的勇氣,更是我要好好學習的地方,我想這一路上能有老師的幫助與肯定,才是讓我能 獲得學位的一大原因。 另外,從資格考開始到非論文考試,一直到最後的學位口試,一路陪著我度過的幾位老師 包括李耀坤老師、張大慈老師、袁俊傑老師與鄭建中老師。感謝您們對於我在整體實驗邏輯上的 慎密性,總是不吝地給予最中肯的辯證。而對於文法結構上的錯誤,也相當用心地加以釜正,並 提供具體的指導及建議。老師們在口試期間所給予的討論與激盪,我想這是督促自己與檢視自己 最直接的方法,也是促使自己能在研究上更摯嚴謹與完備。而老師們在學理上的雄厚底子,也是 學生學習與效法的對象。 另外,生科系楊裕雄老師對於我在大學時期專題研究上的指導與支持,是讓我能順利地從 化學背景轉行進入生物科技領域的關鍵。而應化系鍾文聖老師在有機化學上悉心地教授,更是讓 我受益頗多。 而我的實驗之所以能順利的完成,在這一個過程中,必須要誠心地感謝許多參與其中並且 不吝給予幫忙的很多人。中央研究院的陳玉如老師與廖信凱同學,對於我們在蛋白質的質譜鑑定 上提供了完善且專業的合作,使得我們能順利的解決膜蛋白定序上的難題。應化系刁維光老師、 駱立揚學長與張智煒同學,在超快雷射技術上以及理論化學上的輔助,更提供我們對於實驗結果 上有了另一個角度之思維方式。生科系林玉淳學妹與陳彥甫學弟,在生物資訊學上的幫助與不厭 其煩的指導,得以讓我拓展自己的研究領域。而應化系陳奎百同學、吳曜杉學弟與何孟寰學弟, 對於我在有機合成上也給予了許多寶貴的建議與幫忙。而交大貴儀中心的李蘊明小姐在氣相層析 儀上的輔助與專業的測量,也是讓我們收穫良多。另外特別要感謝清大貴儀中心的彭菊蘭小姐, 您在 NMR 光譜上提供了我最實質的幫助。除了悉心地指導我使用 NMR 外,也非常體恤我們, 總是額外地犧牲自己的假期來幫助我們進行光譜的測量,我們的期刊得以順利發表,您功不可沒。. vii.
(10) 而在我初入實驗室時,那種惶恐忐忑的心情至今尚令我記憶猶存。感謝當初幸如學姐、安 堡學長與逢弦學長總是能以輕鬆風趣的方式來帶領我,使我能很快的進入狀況,對於實驗的本身 也無私地傾囊相受,使我收穫良多。而乾吉學長與素華學姐,您們的經驗傳承也在潛移默化中讓 我對於我的未來有更明確的領悟。此外,我要特別感激喬盈對於我在實驗上與生活上的幫忙,我 們曾共同地為了純化蛋白質而長期的焚膏繼晷,那種刻苦銘心的一再堅持與嘗試,真的讓我印象 深刻。也謝謝妳在論文其中文部份的潤飾。而在生活上,妳也是一直給予我最溫暖的支持,讓我 得以堅強地唸完博士,真的謝謝妳在這一段期間的鼓勵。而青穎與任民是跟我一起進入實驗室的 同學,感謝你們跟我共同地分擔當時的大小庶務。你們現在都有好的工作與學業,身為同學的我 也與有榮焉。 再來則是要感謝震宇學弟,也是因為你的推薦才讓我選擇進入吳老師實驗室。在實驗上, 你總是跟我一起分擔實驗室的工作,也是當時我看棒球的最佳伙伴。貼心的你總是會適時地給予 我久違的關心與問候。另外,媛婷學妹則是跟隨我最久的弟子,反應敏捷的妳總是給我最多的激 盪,與妳共同建立的實驗流程,以及絞盡腦汁後所鑑定出來的化合物,我想是我們留在實驗室的 寶藏,期待妳可以進一步地發揚光大,記得要適時的肯定自己-妳是很優秀的。裕仁學弟則是我 見過最為聰明的孩子,跟我辛苦地完成專題研究也不時的給予建議,並幫助我建立了一系列的基 因資料庫,期望你可以早日突破困境完成博士學位。 吳老師實驗室與我同屬 Cyclase Project 的每一個學弟妹,也都是我在博士班生涯的好幫手。 分生部份的媛婷、美婷、文暄、皓宇、怡伶、文祥與采婷,你們辛苦地培養與分離實驗來源,並 與我共同地進行光譜分析,才使我們得以成功的鑑定出幾個關鍵結構,也才能發表不錯的結果, 其箇中的辛勞是不可言喻的。喬盈、震宇、佳宜、大景、皓宇、裕仁與柏輝更是不辭辛勞並無願 無悔的陪同我徹夜地固守純化的崗位。我們一同守護著這些實驗室得來不易的珍寶,有時令人洩 氣沮喪的結果,有時歡欣鼓舞的收割,這一切一切的滋味,你們都陪我走過。雖然遺憾的是一直 沒有好的結果可以報導,但我深深的覺得純化組在我心中一直是最勞苦功高的。 而其他分屬不同計畫的學弟妹,也分別給予許多有形或無形的幫助。其中裕國一直是老師 不可獲缺的另一個支柱,你在細胞與抗體技巧上的專業也是我協尋幫忙與請益的對象,特別感謝 你與文鴻幫忙建構出我們的單株抗體。晉豪則是幫我在儀器設備上做了無法取代的分憂,尤其是 你在電腦資訊上的長才,也常幫助我化解因為我本身沒有 update 自己資訊技術,而無法解決老 師問題的窘境。文鴻,謝謝你替我分擔了許多庶務管理上的責任,你是一個不可多得,並執著努 力且具有責任感的人。雖然偶遇困境,但希望你能自我突破,以後不管做什麼都能大放異彩。晉 源則是一個幽默風趣,但又不擾其內心沉著與冷靜,並充滿智慧的孩子。一直以來,你常與我在 實驗室外對於實驗有令人印象深刻的討論。生活上,你是我的室友也給予了很多的幫助,謝謝你。 聖慈,妳是一個對自我有高度期許的孩子,對於實驗室的營運也富含想法,謝謝妳提出的許多寶 貴建議,也盼妳能帶領這一個實驗室走出另一個高鋒,對於研究也希望妳能百尺竿頭。Mili is a kindly friend in our lab. Thank your for your joining, concern, and friendship. 另外,謝謝衣鵑平日與 我的討論,使我獲益良多,期許妳能早日發表論文,實驗也越來越順遂。 怡親小妹,妳是個認真有想法的孩子,雖然有時後會想太多。但妳積極進取與上進獨立的 心態卻是無法抹煞的。謝謝妳跟大景、文暄、文祥在這一年多來所帶來的歡樂,學長也預祝妳在 未來博士生涯可以有一個好的開始、好的過程與順利的結果,並常保愉快。另外還有一些已經畢. viii.
(11) 業的碩士班學弟妹,其中,宏城在電腦模擬上的指導,庭翊對於實驗上的執著認真,宏明所帶來 的歡樂以及其他許多的學弟妹,一直都是我向上提升的一股正面力量。另外,目前還在實驗室努 力的良瑋、天昶、育勳、禕庭與亦諄,你們在許多雜事上的分擔也是辛勞,期待你們能再多放點 心思在實驗室上,讓它越來越好,謝謝你們。 而在生科所這一路走來,我遇到的許多貴人,你們總是在最適當的時機給我幫忙,使我不 致於停滯不前。其中,盈蓉學姐是我最後一個階段的戰友,不時給我打氣與鼓勵,也很阿薩力的 幫忙修改我的英文論文,謝謝妳!也希望妳能一直都很開心並舞藝精進。靖婷學姐則是給予了許 多實驗技巧上的指導與幫忙,尤其是放射性實驗的部份真得讓我受益匪淺。美惠學姐與建龍學長 則是在我初入研究所時,總是不吝的與我分享經驗與促膝長談,使我能早日的下定決心。另外, 也因為因緣際會的關係,我可與你們同屆畢業,我也殊感開心。恩仕學長與天木學長則是我在專 題研究時的指導學長,謝謝你們在當時的悉心教授,讓我有一個好的開始也明瞭如何伴演學長的 角色。繼鋒學長與文亮則是我談天的對象,他們努力的方式也令我深感慚愧。另外,生科系辦的 郭珍佑小姐、呂聖鈴小姐、郭淑卿小姐、吳佳文小姐以及葉連發先生總是在許多事務上對我給予 幫助,謝謝你們。而我們的師母-賴美伶小姐一直以來,總是在實驗上與經驗上給予我許多的寶 貴建議,您親切與開朗的態度也常伴演我們與老師間的潤滑劑,謝謝您。 大學同學伯凱是我在生物科技方面的嚮導員,謝謝你總是很熱情地丟給我很多東西,希望你 身體越來越好並早日榮獲學位。春發、秉恒、志豪、哲一則是我一直以來的好友,另外明妮、姿 惠、怡珊、智華對於我的博士班生活也常保關心。小黃家族的每一個成員都是非常在乎我的人, 也是我這段期間能夠放鬆休憩的地方,你們的關懷我都充滿感激,也都是我心中的力量。 最後,更要感謝的是我的家人-我的父母親、兩個弟弟還有我的奶奶以及已經過世的爺爺。 父親的身教與言教是督促我向上努力的最大後盾。您當初為了我的學業,拖著疲憊的身體參與考 試並通勤上班,至今的記憶我依然清晰難忘。母親的關心則是我心中最大的暖流,是我永遠的避 風港,我終於可以告訴您︰妳的儍兒子終於拿到學位了。兩個聰明的弟弟則是我的驕傲,你們一 直都比我優秀也很有成就,是身為大哥的我能安心唸書的原因,謝謝你們。爺爺與奶奶永遠都是 最疼我們的人,也是我心中最為牽掛的人。 在交大十二年的光陰要感謝的人實在是太多了,深怕不慎有所遺漏。若有為之,真的甚感 抱歉!感謝上蒼賜與的這一切。感謝所有關心我、陪伴著我的人,僅能以小小的版面真的不足以 聊表我最深忱的謝意。未來的我會更加的努力,也在此給與您們最大的祝福。 願大家都能平安、順心、健康、愉悅。 程翔 2008/7/25. ix.
(12) Table of Contents Chapter 1. General introduction. ..........................................................................................................1 1.1 Overview of oxidosqualene cyclase ..............................................................................................1 1.2 Historical hypothesis of cyclization mechanism............................................................................6 1.2.1 The mechanistic and stereochemical insights of oxidosqualene cyclization..........................6 1.2.2 The theoretical enzyme-templated cyclization reactions .....................................................15 1.3 The biological studies of 2,3-(oxido)squalene cyclase enzymes .................................................21 1.3.1 Enzymology of 2,3-(oxido)squalene cyclase enzymes ........................................................21 1.3.2 Molecular cloning of 2,3-oxidosqualene cyclase .................................................................23 1.3.3 Crystallization and structural characterization of cyclase ....................................................27 Chapter 2. Theis organization .............................................................................................................33 Chapter 3. Mutagenesis approach to investigate the putatuve active-site residues from oxidosqualene cyclase .......................................................................................................35 3.1 Introduction .................................................................................................................................35 3.2 Results and Discussion. ...............................................................................................................42 3.2.1 Generation of the alanine-scanning mutations or the site-directed mutations of S. cerevisiae ERG7 gene....................................................................................................................................42 3.2.2 Principle of plasmid shuffle methodology ...........................................................................43 3.2.3 Screening of inactive ERG7 mutants by using the plasmid shuffle method ........................45 3.2.4. Lipid extraction, column chromatography, and product characterization ...........................46 3.2.5 The construction of novel gene disruption strain TKW14 for analysis of inactive mutated cyclase...........................................................................................................................................49 3.2.6 Characterizing the mutant products in the novel gene disruption strain ..............................51 3.2.7 Site-saturated mutagenesis approach to investigate the functional importance of the critical Tyr-510 residues............................................................................................................................55 3.3.8 Homology modeling illustration of the functional critical residues .....................................64 Chapter 4. Construction of chimeric oxidosqualene cyclase gene to study the product specificity ............................................................................................................................................72 4.1 Introduction. ................................................................................................................................72 4.2. Results and Discussion. ..............................................................................................................76 4.2.1 Construction of a chimeric library between oxidosqualene-lanosterol cyclase and oxidosqualene-cycloartenol synthase............................................................................................76 4.2.2 Functional analysis of chimeric enzymes library via plasmid shuffle and the product characterization. ............................................................................................................................80 Chapter 5. Mechanistic insights into oxidosqualene cyclization through homology modeling and quantum mechanism calculation.....................................................................................86 5.1 Introduction. ................................................................................................................................86 5.2 Results and Discussion. ...............................................................................................................87 5.2.1 Sequence alignment, secondary structure prediction, and homology modeling ..................87 5.2.2 Geometrical verification of the generating homology models .............................................91 5.2.3 Comparsion between homology models and mechanistic hypothesis..................................95 5.2.4 Correlation of the OSC model to the experimental mutagenesis studies ...........................106 5.2.5 Quantum mechanism calculation for illustration of the lanosterol synthesis .....................121 5.3 CONCLUTION AND FUTURE PERSPECTIVE. ..................................................................................125 x.
(13) Chapter 6. Purifiation, tandem mass characterization, and molecular cloning of oxidosqualene lanosterol cyclase enzyme from bovine liver ..................................................................130 6.1 Summary....................................................................................................................................130 6.2 Introduction. ..............................................................................................................................131 6.3 Results and Discussion. .............................................................................................................135 6.3.1 Purification of bovine liver OSC........................................................................................135 6.3.2 Identification of OSC by tandem mass spectrometry.........................................................139 6.3.3 Molecular cloning of OSC from bovine liver.....................................................................142 6.3.4 Genetic complementation of recombinant bovine liver OSC in yeast erg7 deficient strain: ....................................................................................................................................................147 6.3.5 Mass spectrometry determination of the photoaffinity labeling bovine liver OSC with Ro48-8071. .................................................................................................................................147 6.3.6 Monoclonal antibody preparation and Western blotting analysis.......................................150 Chapter 7. Development and applization of fluorescent Ro48-8071 derivative probes to study oxidosqualene-lanosterol cyclase.....................................................................................154 7.1 Summary....................................................................................................................................154 7.2 Introduction. ..............................................................................................................................155 7.3 Results and Discussion. .............................................................................................................157 7.3.1 Synthesis of newly Ro48-8071-based fluorescent probes..................................................157 7.3.2 The inhibitory activity of the Ro48-8071-based fluorescent probes. .................................162 7.3.3 Spectroscopy of these newly Ro48-8071-based fluorescent probes. .................................165 7.3.4 Homology modeling and ligand docking studies of newly Ro48-8071-based fluorescent probes..........................................................................................................................................170 7.3.5 Time-resolved fluorescence studies of Ro48-8071-based fluorescent probes ...................173 Chapter 8. Identification of DNA aptamers for bovine liver oxidosqualene-lanosterol cyclase via SELEX ...............................................................................................................................176 8.1 Introduction ...............................................................................................................................176 8.2 Results and Discussion ..............................................................................................................184 8.2.1 In vitro selection of DNA aptamer for bovine liver OSC...................................................184 8.2.2 Gel mobility shift assay of the binding between DNA aptamer and OSC .........................187 8.2.3 Gel mobility shift assay provide alternative selection strategy for SELEX .......................190 8.2.4 Molecular cloning and DNA sequencing analysis of individual aptamer ..........................191 8.2.5 The binding characterization of the individual aptamer molecules....................................199 8.3. Conclusion ................................................................................................................................200 Chapter 9.Experimental section........................................................................................................201 9.1 Materials: ...................................................................................................................................201 9.1.1 General bacterial strains and molecular cloning/expression vectors:.................................201 9.1.2 Enzymes, chemicals, equipments, and reagents:................................................................201 9.2 General experimental procedures: .............................................................................................206 Chapter 3.....................................................................................................................................206 Chapter 4.....................................................................................................................................210 Chapter 5..................................................................................................................................... 211 Chapter 6.....................................................................................................................................213 Chapter 7.....................................................................................................................................219 Chapter 8.....................................................................................................................................221 xi.
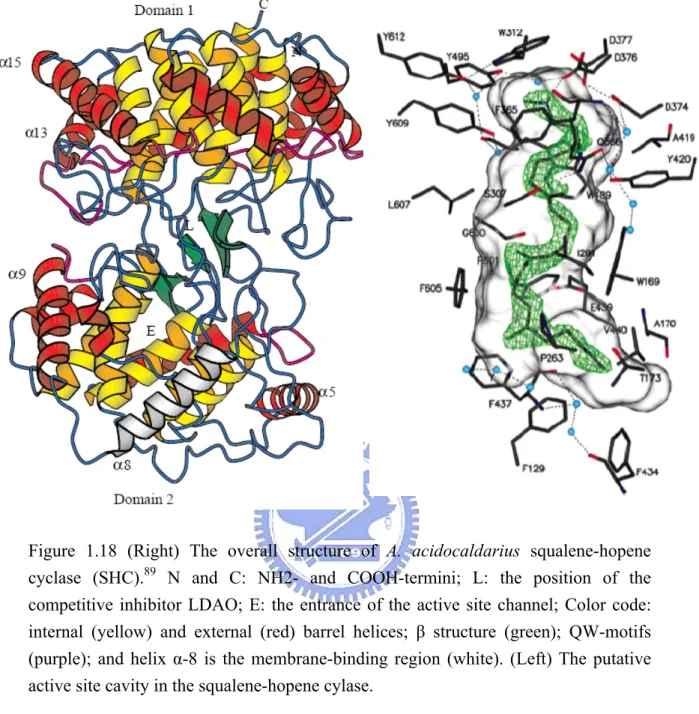
(14) References. ..........................................................................................................................................225. Table of Figures Figure 1.1 The product diversity of (oxido)squalene cyclase in different species....................................2 Figure 1.2 The cyclization cascades of oxidosqualene to lanosterol and the end product, cholesterol in the human cholesterol biosynthetic pathway...................................................................................3 Figure 1.3 The cyclized products are classified according to the stereochemistry configuration of carboncationic intermediates………………………………………………………………………4 Figure 1.4 The postulated cyclization mechanism of oxidosqualene .......................................................5 Figure 1.5 Squalene cyclization model for lanosterol production………………………………….........7 Figure 1.6 The acylic azasqualenes mimic the C-8 and C-20 carbonoum ion intermediates……………9 Figure 1.7 Structure of potent inhibitors of oxidosqualene cyclase enzymes…………….... …………...9 Figure 1.8 Some of the substrate derivatives inhibitors..........................................................................10 Figure 1.9 The geometrical isomers of substrate analogs....................................................................... 11 Figure 1.10 Minor products with a 6,6,5,4 tetracyclic scaffold and 6-6-5 scaffold were isolated from the enzymatic transformation with various analogs............................................................................13 Figure 1.11 Cyclization of oxidosqualene analogs produce 17β-protosteryl derivatives .......................13 Figure 1.12 The hypothesized cyclization cascade of oxidosqualene he acylic .....................................14 Figure 1.13 Johnson proposed three cation-stabilizing auxiliaries in the cyclase. .................................15 Figure 1.14 Hypothetical model based on the cyclization mechanism...................................................17 Figure 1.15 Potential modes of mechanism-based inactivation..............................................................19 Figure 1.16 Affinity labeled cyclase fragment with mechanistic suicide inhibitors. ..............................19 Figure 1.17 Structures of various squalene and dodecyl derivatives of thiol-modifing agents…..………………………… ……………………………………………………………...20 Figure 1.18 Overall structure of A. acidocaldarius squalene-hopene cyclase (SHC)………………... . 30 Figure 1.19 Crystal structure of human OSC .........................................................................................31 Figure 1.20 Local view of crystallized structure of human OSC............................................................32 Figure 3.1 Thin-layer chromatographic analysis of the nonsaponifiable lipid extract............................48 Figure 3.2 Acetylation with acetic anhydride/ pyridine was used to modify the polarity of alcohol moiety............................................................................................................................................48 Figure 3.3 The homology recombination strategy for exchanging the HEM1 gene with a drug geneticin resistance gene ..............................................................................................................................50 Figure 3.4 GC analyses of the nonsaponifiable lipid (NSL) products ....................................................53 Figure 3.5 The X-ray crystallized structure of (a) SHC or (c)OSC and the respective homology modeling structures of (b) wild-type ERG7 and (d) human OSC .................................................67 Figure 3.6 Local view of the homology modeled S.cerevisiae ERG7 structure .....................................68 Figure 3.7 Local view of superimposition of homology modeled S.c. ERG7 structure and different single-point mutations or double-positions mutations. .................................................................70 Figure 3.8 Local views of Asp-456, Trp-443, Lys448, and Phe445 positions in S. cerevisiae ERG7 structure. .......................................................................................................................................71 Figure 4.1The analysis of the preliminary NSL extract from respective ten chimeras on the TLC plate. ......................................................................................................................................................82 Figure 4.2 The GC-FID spectrums of the partially purified NSL from the TKW14 strain transformed with the respective chimera enzymes............................................................................................82 Figure 4.3 Sequence alignment and secondary structure elements comparison between β-amyrin synthase and lupeol synthase or lanosterol synthase and cycloartenol synthase...........................85 Figure 5.1 Multiple sequence alignment and secondary structure elements comparison .......................90 xii.
(15) Figure 5.2 The 3D-1D profiles of the homology modeling structure of yeast ERG7 (Upper plot) and human OSC crystal structure (Downer plot).................................................................................93 Figure 5.3 The Ramachandran plot of the homology model of yeast S.c.ERG7 ....................................94 Figure 5.4 Superimposition of the S.c.ERG7 homology modeling structure with crystal structures .....96 Figure 5.5 Local views of the superimposition of homology modeled S.c. ERG7 structure with human OSC crystal structure ....................................................................................................................97 Figure 5.6 Local views of the superimposition of homology modeled S.c. ERG7 structure with SHC crystal structure...........................................................................................................................100 Figure 5.7 The hypothetical enzyme-triggered stereic control for the formation of the boat B-ring and Local view of S.c.ERG7 homology modeling structure. ............................................................103 Figure 5.8 The different orientation of Trp232 from S.c.ERG7 and Phe601 from A.a.SHC involved in the formation of C ring. ..............................................................................................................104 Figure 5.9 Local view of the homology modeling S.c.ERG7 structure................................................106 Figure 5.10 The chemical structure of the possible producing cationic reactive intermediates or various annulated products. .....................................................................................................................108 Figure 5.11 The isolated respective products from various S.c ERG7X mutates in this thesis ............109 Figure 5.12 Stereo representations of S. cerevisiae wild-type ERG7 homology modeling structure ...112 Figure 5.13 The steretic representation of various S.c ERG7 mutated homlogy modeling structures. 114 Figure 5.14 Putative functional residues in the π-electron pocket of S.c ERG7, in which Phe-445 is near the B/C ring junction...................................................................................................................117 Figure 5.15 Local view of the homology model of S.c. ERG7.............................................................118 Figure 5.16 The partially relative energy profiles among various intermediate stages, derived from quantum mechanics calculations.................................................................................................124 Figure 6.1 Silver-stained SDS–PAGE gel and Coomassie-stained non-denaturing PAGE gel of bovine liver OSC ....................................................................................................................................138 Figure 6.2 Tandem mass spectrum of the doubly charged tryptic peptide from purified bovine liver OSC protein. ...............................................................................................................................140 Figure 6.3 Sequence alignment of trypsin-digested peptides from bovine liver OSC observed by tandem mass spectrometry. .........................................................................................................141 Figure 6.4 The nucleotide and deduced amino acid sequences of cDNA from bovine liver OSC ......144 Figure 6.5 Multiple sequence alignments of oxidosqualene-lanosterol cyclase (OSC)........................146 Figure 6.6 TLC analysis of cyclase activity before and after incubated with Ro 48-8071 ...................149 Figure 6.7 Peptide coverage of bovine liver OSC from LC-MS/MS analysis......................................150 Figure 6.8 Crystal structure of human OSC in complex with lanosterol And Homology modeling structure of bovine liver OSC .....................................................................................................152 Figure 6.9 (a) SDS-PAGE and (b) Western blotting analysis of purified bovine liver OSC.................153 Figure 7.1 TLC analysis of cyclase activity from which OSC was pre-incubated with different Ro48-8071-based derivatives......................................................................................................164 Figure 7.2 The wavelength scanning of fluorescent probes in aqueous solution..................................166 Figure 7.3 The fluorescence spectrum of Ro4-DPA fluorescent probe in aqueous solution, ethanol, and DMSO.........................................................................................................................................168 Figure 7.4 Fluorescence characteristic of Ro48-8071-based fluorescent probes on either (a) TLC plate, or (b)in the aqueous and in the DMSO solution..........................................................................169 Figure 7.5 Fluorescence spectra of Ro4-DPA with addition of bovine liver OSC…………..169 Figure 7.6 The molecular ligand docking results of the (a) Ro4-NA1, (b) Ro4-NA2, (c) Ro4-BP3, and (d) Ro4-BP4 for the bovine liver OSC active site cavity. ...........................................................172 Figure 7.7 The molecular ligand docking results of the Ro4-DPA for the bovine liver OSC active site cavity...........................................................................................................................................173 Figure 7.8 (a) The laser induced fluorescence decay or (b) the anisotropy analysis of the Ro4-NA1 xiii.
(16) fluorescent probes. ......................................................................................................................175 Figure 8.1. The schematic diagram for the scelection of DNA aptamer to target molecule, i.e. the general SELEX procedure ..........................................................................................................179 Figure 8.2. PCR amplicon of the resultant DNA which was eluted from the surface of the microtitter plates during SELEX-3rd-cycle ..................................................................................................186 Figure 8.3 Gel mobility shift assay of different amount of DNA probe interacted with the fixed bovine liver OSC ....................................................................................................................................188 Figure 8.4 Gel mobility shift assay of fixed amount of DNA probe with the various amount of bovine liver OSC. ...................................................................................................................................188 Figure 8.5 Gel mobility shift assay of competition experiment by using the unlabeled DNA probe. ..189 Figure 8.6 (a), (b) Gel mobility shift assay of DNA molecules isolated from microtitter plate for bovine liver OSC. ...................................................................................................................................190 Figure 8.7 Sequence alignments of the respective aptamer clones.......................................................193 Figure 8.8 The representative predicted secondary structures of aptamer DNAs from the SELEX-9th-cycle via the mfold sever prediction. ......................................................................196 Figure 8.9 Gel mobility shift assay of individual DNA aptamer molecules for bovine liver OSC.......197 Figure 8.10 The concentration dependent OSC-binding response was revealed on the SPR analysis of aptamer-125-9-13........................................................................................................................199. Table of Schemes Scheme 3.1 The respective A. thaliana CAS1 mutants changed their product specificity from cycloartenol to lanosterol………………………………………………………………………...37 Scheme 3.2 Screening of inactive ERG7 mutants using a plasmid shuffle method……………..…......44 Scheme 3.3 Oxidosqualene-lanosterol cyclase mutants converted oxidosqualene to a variety of monocyclic and deprotonated compounds ....................................................................................54 Scheme 3.4. Proposed cyclization/rearrangement pathway of oxidosqualene in TKW14C2 expressing ERG7Y510X site-saturated mutagenesis. .........................................................................................60 Scheme 4.1 The schematic representation of the partial chimera library between β-amyrin synthas and lupeol synthase enzymes...............................................................................................................74 Scheme 4.2 The schematic representation of the entire chimerical library between oxidosqualene-lanosterol cyclase gene (OSC) and oxidosqualene-cycloartenol cyclase gene (CAS) ......................................................................................................................................................78 Scheme 4.3 The diagrammatic explanation for the PCR-based construction strategy ...........................79 Scheme 5.1 The hypothesized cyclization mechanism of oxidosqualene or squalene ...........................98 Scheme 5.2 The proposed model of Tyr-99 for the stereochemical control…………………… … …119 Scheme 6.1 Three major photoreactive groups .................................................................................. 134 Scheme 6.2 Formation of covalent adducts from photoexcited benzophenone group..........................134 Scheme 7.1 Synthesis routine of the aromatic boronic pinacol ester....................................................158 Scheme 7.2 The scheme shows the first published Suzuki coupling, which is the palladium-catalyzed cross coupling between organoboronic acid and halides ............................................................159 Scheme 7.3 Derivation reaction of Ro48-8071 with DPA or other four boronic pinacol ester compounds ....................................................................................................................................................160 Scheme 7.4 The chemical structures of five newly Ro48-8071-based fluorescent probes. ..................161. xiv.
(17) Table of Tables Table 1.1 OSCs that have been cloned and functionally characterized in yeast .....................................26 Tabel 3.1 Complementation result of cyclase mutants in a ERG7 knockout strain, CBY57, by using plasmid shuffling counter selection. .............................................................................................46 Table 3.2 Product ratios of the Saccharomyces cerevisiae OSC single mutants.....................................54 Table 3.3 The product profiles of S. cerevisiae TKW14 expressing the ERG7Y510X site- saturated mutants and two ERG7H234WY510X mutants....................................................................................56 Table 5.1 Statistical occurrence of φ, ψ angles in the PDB bank compared with the φ, ψ angles distribution in the homology modeling structure of yeast S.c.ERG7............................................94 Table 6.1 Purification and properties of 2,3-oxidosqualene cyclase from vertebrates, higher plants, and yeast ............................................................................................................................................137 Table 6.2 The purification of oxidosqualene-lanosterol cyclase from bovine liver..............................137 Table 7.1 The inhibitory activity assay of the newly Ro48-8071-based fluorescent probes for bovine liver OSC ....................................................................................................................................163 Table 8.1 The comparison of functional characteristics between aptamers and antibodies..................180 Table 8.2. The concentration of OSC or DNA library load into the each SELEX round. ....................186 Table 9.2.4.1 Primers sequence in the chimerical library construction………………………………..211. List of Abbreviations A, Ade APS bp BSA cDNA dH2O ddH2O DEPC DMF DMSO DNA dsDNA dNTP DPA DTT EDTA EMSA ELISA GC H, His HAP HCl HPLC IPTG kb(s) kDa(s) KPi. Adenine Ammonium Persulfate Base Pair Bovine Serum Albumin Complementary DNA Distilled Water Double Distilled Water Diethylpyrocarbonate Dimethylformamide Dimethylsulfoxide Deoxyribonucleic acid Double-stranded DNA Deoxynucleoside triphosphate 4-(4,5-diphenyl-lH-imidazol-2-yl) phenylboronic acid Dithiothreitol Ethylenediaminetetraacetic acid Electrophoresis Gel mobility shift assay Enzyme-Linked Immunosorbent Assay Gas Chromatography Histidine Hydroxyapatite Hydrogen Chloride High Performance Liquid Chromatography Isopropyl-1-thio-β-D-galactopyranoside kilobase(s) kiloDalton(s) Potassium phosphate xv.
(18) L, Lys LA LB LiOAc mAb M, Met MS NaCl NaN3 NMR NaOH NC OS pAb PBS PCR PEG PMSF PPh3 PVDF RT rpm SDS-PAGE SELEX SPR ssDNA T, Trp TCA TEMED TFA THF TLC Tris base TX-100 U, Ura UV/Vis wt X-gal YNB. Lysine Lanosterol Luria-Bertani Lithium Acetate monoclonal antibody Methonine Mass Spectromerty Sodium Chloride Sodium Azide Nuclear Magnetic Resonance Sodium Hydroxide Nitrocellulose Oxidosqualene polyclonal antibody Phosphate Buffered Saline Polymerase Chain Reaction Polyethylene Glycol Phenylmethylsufonyl fluoride Triphenylphosphine Polyvinylidene difluoride Room Temperature revolutions per minute Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis Systematic Evolution of Ligands by Exponential Enrichment Surface Plasmon Resonance Single-stranded DNA Tryptophan Trichloroacetic Acid N,N,N',N'-tetramethylethylenediamine Trifluoroacetic Acid Tetrahydrofuran Thin Layer Chromatography Tris(hydroxymethyl)-aminomethane Triton X-100 Uracil Ultraviolet/Visible Wild type 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside Yeast Nitrogen Base. xvi.
(19) List of Genes or Proteins ADE2 bAS b. t. LSS B. l. OSC CAS1 CAS A. t. CAS1 ERG1 ERG7 S. c. ERG7 ERG11 HIS3 KanR/NeoR LUP1 LUS LEU2 PNY2 PSY OSC PNK SE TRP1 URA3. Phosphoribosylaminoimidazole Carboxylase gene. β-Amyrin Synthase Bos Taurus 2,3-oxidosqualene-lanosterol cyclase gene; lanosterol synthase gene Bos Taurus 2,3-oxidosqualene-lanosterol cyclase; lanosterol synthase Cycloartenol Synthase gene Cycloartenol Synthase Arabidopsis thaliana Cycloartenol Synthase Squalene Epoxidase gene Oxidosqualene Cyclase gene; Lanosterol Synthase gene Saccharomyces cerevisiae lanosterol synthase Lanosterol 14-α-demethylase gene Imidazoleglycerol-Phosphate Dehydratase gene Neomycin Phosphotransferase; Kanamycin/neomycin resistance gene; G418 Resistance gene Lupeol synthase 1 gene Lupeol synthase β-isopropylmalate Dehydrogenase (IMDH) gene Panax ginseng β-Amyrin Synthase gene Pisum sativum β-Amyrin Synthase gene Oxidosqualene-Lanosterol Cyclase T4 polynucleotide kinase Squalene Epoxidase Phosphoribosylanthranilate Isomerase gene Orotidine-5’-Phosphate (OMP) Decarboxylase gene. xvii.
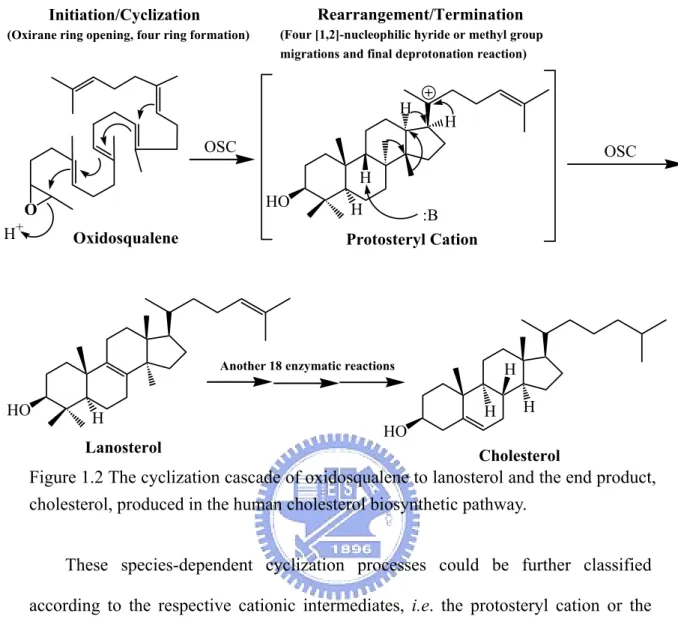
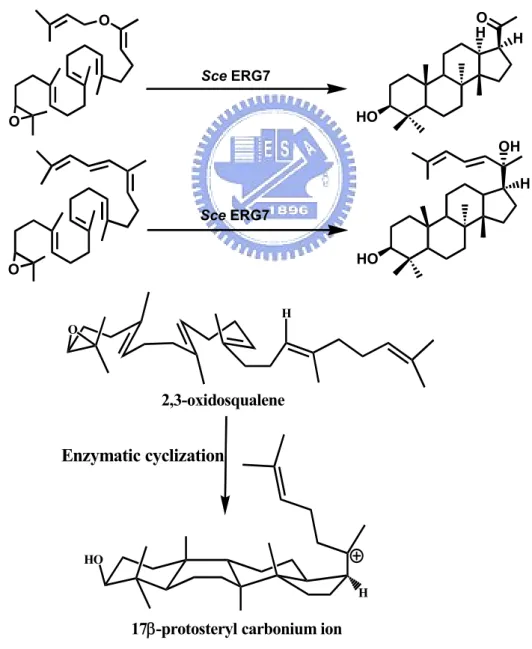
(20) Chapter 1 General Introduction. 1.1 Overview of oxidosqualene cyclase. Oxidosqualene cyclases (OSCs) (EC 5.4.99.-) constitute a family of enzymes which catalyze the common substrate, oxidosqualene, into vast skeletal diversity of cyclization products, acting as the biosynthetic precursors for sterols, related membranous components, steroid hormones, and other secondary metabolites. Up to date, over one hundred different cyclic triterpene alcohols are respectively formed via their individual oxidosqualene cyclases.1 The species-dependent product specificity of cyclase is well-known and fascinating. For example, oxidosqualene is solely transformed into tetracyclic lanosterol in animals and fungi, whereas a variety of polycyclic triterpenoid products, including cycloartenol, lupenol or β-amyrin are produced in higher plants or algae. In another example, squalene cyclase converts the precursor of oxidosqualene, squalene, into pentacyclic hopene or hopanol, which act as their corresponding membranous triterpenoid lipid in lower plants or bacteria (Figure 1.1). This kind of single protein capable of multiple functional enzymatic transformations is one of the most impressive naturally existing reaction-phenomena. The oxidosqualene cyclization reaction is remarkable for its complexity. One acyclic substrate with only one stereogenic center is converted into polycyclic products containing multiple rings and several chiral centers. The conversion from the linear oxidosqualene into polycyclic products contains at least ten steps of covalent bond cleavage and formation. The cyclization cascade comprises one oxirane ring opening, 1.
(21) four or five stereochemically controlled ring formations, four nucleophilic hydride or methyl group migrations, and the final deprotonation reaction. Although the end product, cholesterol, produced in the human cholesterol biosynthetic pathway is required by another eighteen enzymatic reactions, the oxidosqualene cyclases are undoubtedly responsible for the most fascinating step (Figure 1.2).. Figure 1.1 The product diversity of (oxido)squalene cyclase in different species.. 2.
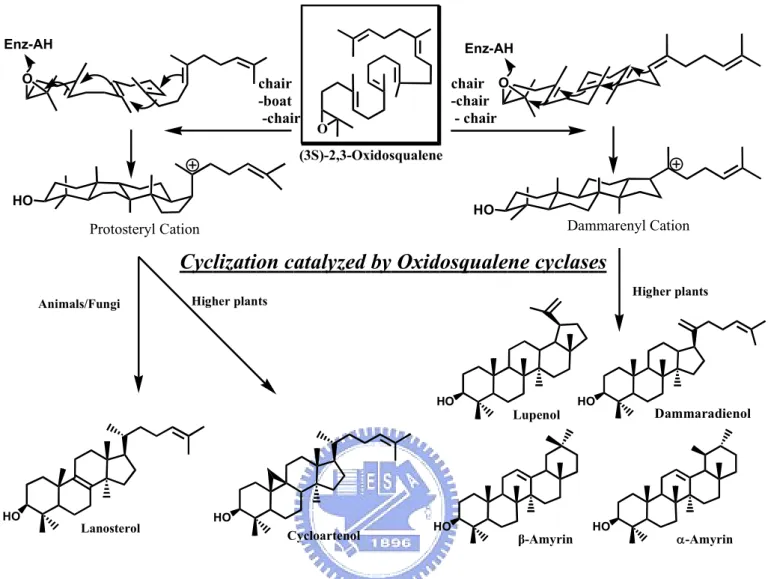
(22) Rearrangement/Termination. Initiation/Cyclization (Oxirane ring opening, four ring formation). (Four [1,2]-nucleophilic hyride or methyl group migrations and final deprotonation reaction). H. H. OSC. OSC H HO. O H. +. H. :B Protosteryl Cation. Oxidosqualene. Another 18 enzymatic reactions. HO. H H. H. H. HO. Lanosterol. Cholesterol. Figure 1.2 The cyclization cascade of oxidosqualene to lanosterol and the end product, cholesterol, produced in the human cholesterol biosynthetic pathway. These species-dependent cyclization processes could be further classified according to the respective cationic intermediates, i.e. the protosteryl cation or the dammarenyl cation. The main difference between these two major intermediate cations is the stereochemical configuration of the second cyclohexyl B-ring. In the protosteryl pathway, the “boat” six-membered B-ring is produced, whereas the “chair” cyclohexyl B-ring is found in the dammarenyl cationic pathway (Figure 1.3). Thus, after the formation of protosteryl cation via the “chair-boat-chair” pre-folded conformation, the substrate is triggered to yield either lanosterol or cycloartenol followed by a series of [1,2]-methyl/hydride groups shifts, and the final alternative elimination. In parallel, the “all-chair” dammarenyl cation processes the similar rearrangement, ring-expansion, and final elimination to give dammaradienol, lepenol, α-amyrin or β-amyrin in higher plants or algae (Figure 1.4).2 3.
(23) Enz-AH. Enz-AH. O. chair -boat -chair. chair O -chair - chair. O. (3S)-2,3-Oxidosqualene HO. HO. Dammarenyl Cation. Protosteryl Cation. Cyclization catalyzed by Oxidosqualene cyclases Animals/Fungi. Higher plants. Higher plants. HO. HO. Lanosterol. HO. HO. Cycloartenol. Lupenol. β-Amyrin. HO. HO. Dammaradienol. α-Amyrin. Figure 1.3 The cyclic products are classified according to the stereochemical configuration of carbocationic intermediates.. 4.
(24) 18. 6 3. O. 10. chair - boat - chair. H. + H. H. 15. H. 2. H. + H. HO Protosteryl Cation. chair -chair - chair. HO. Cycloartenol. HO. Lanosteryl Cation. +. H. +. HO Dammarenyl Cation. Lanosterol. HO. HO. Dammarenyl Cation. +. +. H. HO Lupenyl Cation. H H. HO. Baccharenyl Cation. +. Oleanyl Cation. HO. +. H H. HO. H. H. HO. Dammaradienol. HO. Lupenol. HO. HO. α -Amyrin. β -Amyrin. Figure 1.4 The postulated cyclization mechanism of oxidosqualene into diverse products via different cationic intermediates. The respective products are shown in frame. 5.
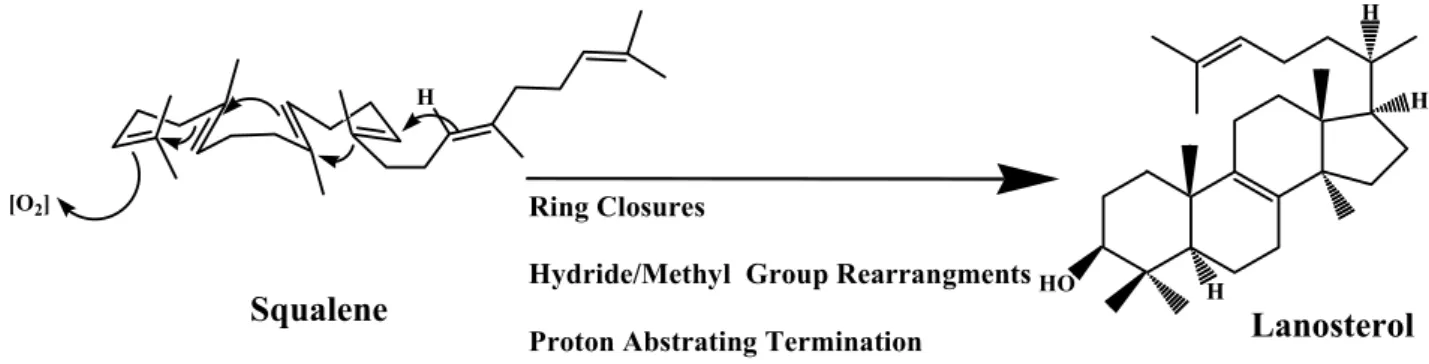
(25) 1.2 Historical hypothesis of cyclization mechanism. 1.2.1 The mechanistic and stereochemical insights of oxidosqualene cyclization From the view of chemical reaction, diverse products should be obtained after the intramolecular cyclization reaction of the flexible linear polyolefinic substrate, oxidosqualene. The highly exothermic process during the formation of novel carbon-carbon bonds from the nucleophilic attack of olefinic π-electrons on carbocationic atoms also cause the difficulty in controlling the reaction tendency in the non-enzymatic cyclization of oxidosqualene. However, the biological system utilized the species-dependent oxidosqualene cyclases to rigorously trigger the cyclization reaction for generations of specific biosynthetic products. The substrate folding manners, the substrate-enzyme interactions, even the processes of substrate immigration or products released are thought to be subject to regulation. Many organic and bioorganic experiments have been designed to understand the relationships between the cyclization mechanism and the cyclase enzyme itself. Because of the intrinsic difficulty, the cyclization mechanism of squalene or oxidosqualene has intrigued scientists for over half of century. After the first pioneering model was established in 1934, different radioisotope feeding experiments were further used to examine the biosynthetic source of cholesterol.3,. 4. With the. successful structural determination of lanosterol, Woodward and Bloch proposed a putative model for lanosterol production from squalene, and also confirmed its biological importance (Figure 1.5).5-8 The following experiments established that the oxidosqualene rather than the squalene is the direct precursor of lanosterol, while the 3S isomer was further proved to be the exclusive precursor.9-11. 6.
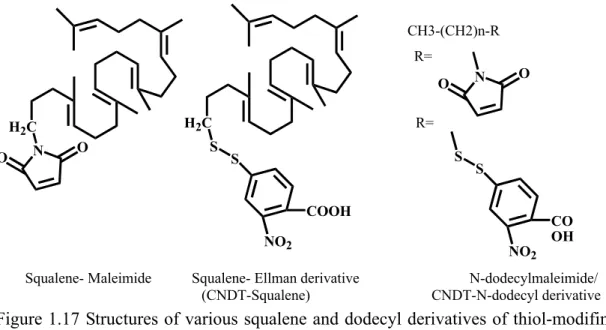
(26) H H. [O2]. H. Ring Closures. Squalene. Hydride/Methyl Group Rearrangments HO. H. Lanosterol. Proton Abstrating Termination. Figure 1.5 Squalene cyclization model for lanosterol production, proposed from Woodward and Bloch.6. Subsequently, various enzymatic incubations of chemically synthesized substrate analogs or inhibitors provided much valuable information regarding the mechanism of oxidosqualene cyclization. For example, a series of structural analogs were utilized to elucidate the conformational requirement or the molecular geometry of the substrate within the enzymatic active sites. These chemical derivatives not only indicated the importance of internal methyl groups/unsaturated double bonds for the oxidosqualene cyclization, but also illustrated the enzymatic toleration for the modification on the oxidosqualene molecule. For example, substrate derivatives, which are modified in the. distal. terminus,. including. the. double. epoxide. rings. derivative. (2,3-22,23-dioxidosqualene), the saturation of terminal double bond derivative (22,23-dihydro-2,3-oxidosqualene), and distal functional modification derivatives (22,23-dihydro-22-methylene-2,3-oxidosqualene or 30-acetyl-2,3- oxidosqualene), were all successfully cyclized via an enzyme triggered reaction.12-15 Similarly, neither the hydroxyl, trimethylsilyl, nor methylidene modification at C-29 methyl group appear to influence the cyclase activity (Figure 1.8).16 Nevertheless, the substrate derivatives with saturation modification of the internal double bond, either 10,11-dihydro-2,3-oxidosqualene. or. 14,15-dihydro-2,3-oxidosqualene,. are. not. suitable substrates for the cyclization reaction. Cyclase also only partially catalyzed 7.
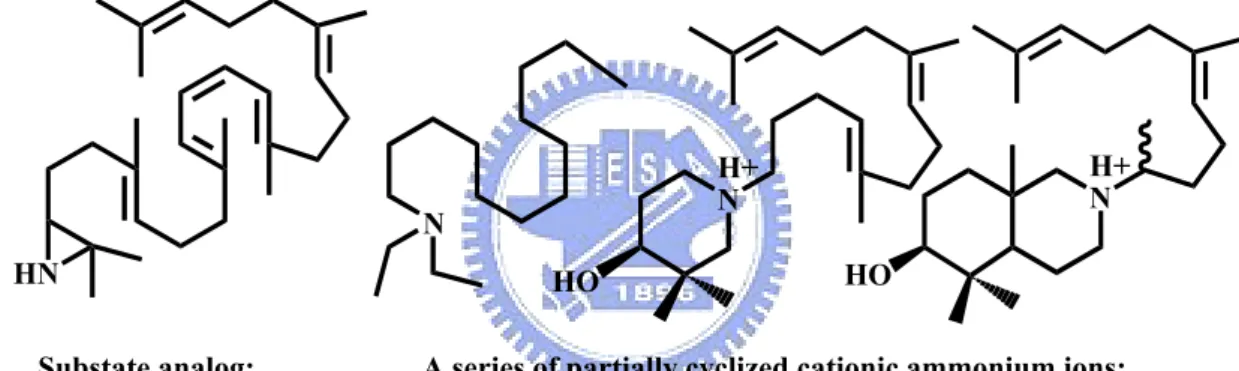
(27) the double demethyl analogs, 10,15-demethyl-2,3-oxidosqualene, into 6,6,5,4 tetracyclic product, suggesting the functional importance of internal methyl group (Figure 1.8).17,18 The modification near the epoxide terminus of substrate, such as 2,3-iminosqualene,. 2,3-cis-1’nor-2,3-oxidosqualene,. 2,3-trans-1’nor-2,3-. oxidosqualene, or 1,1’-bisnor- 2,3-oxidosqualene were used to illustrate the enzymatic tolerance (Figure 1.8).19-21 The necessity of epoxide ring for the OSC-catalyzed reaction was further supported via two acyclic azasqualenes analogs in Figure 1.6.22 The 19-aza-18,19,22,23-tetrahydrosqualene-2,3-epoxide and its non-epoxide ring analogs, 19-aza-18,19,22,23-tetrahydrosqualene, were applied for the cyclase mediated reaction. Only 19-aza-18,19,22,23-tetrahydrosqualene-2,3-epoxide exhibited the inhibitory activity for the oxidosqualene cyclase, indicating the importance of the epoxide ring. In other experiments, a series of substrate analogs or ammonium ions that mimic the transition state in the different stages of cyclization were also synthesized and used to illustrate their inhibition effect (Figure 1.7).19, 23-26 The conformational requirements of oxidosqualene for the cyclase-triggered reaction were further observed by using the different acyclic azasqualene derivatives (Figure 1.9). For example, two geometrical isomers, (6E or 6Z)-10-aza-10,11dihydrosqualene 2,3-epoxide, were used to interact with cyclase.27,28 Only the 6E-isomer which corresponds to the natural all E-form conformation in oxidosqualene was functionally active. Moreover, one pair of isomers, (18Z or 18E)-29methylidene-2,3-oxidosqualene were also utilized as the mechanistic inhibitors for oxidosqualene cyclase.29 The (Z)-form methylidene derivative showed the potent and irreversible inhibitory activity, whereas the (E)-form revealed no inhibition. The large difference in the IC50 value between these isomers indicated the steric requirement for the protersteryl cation in the enzymatic cyclization process. These geometrical isomers, E-form or Z-form derivatives, confirmed that the natural substrate should be 8.
(28) folded with all E-form conformation except for the carbon-18, and be cyclized via all-trans geometry for the cyclization reaction in the enzyme active site.27-29. N+. N+. H. H. O. 19-aza-18,19,22,23-tetrahydrosqualene-2,3-epoxide and its analogues without epoxide. Figure 1.6 Acylic azasqualenes mimic the C-20 carbonium ion intermediates.. N HN Substate analog: 2,3-iminosqualene. H+ N. H+ N HO. HO. A series of partially cyclized cationic ammonium ions:. Figure 1.7 Structure of potent transition state inhibitors of oxidosqualene cyclase.. 9.
(29) O. O (3S)-2,3-oxidosqualene. O. O. 2,3-22,23-dioxidosqualene. 22,23-dihydro-2,3-oxidosqualene 29. Acetyl. R. O. O. O 22,23-dihydro-22-methylene -2,3-oxidosqualene. 30-acetyl-2,3-oxidosqualene. R= hydroxyl, trimethylsilyl, or methylidene derivatives. H H O. O 10, 11-dihydro-2,3oxidosqualene. O. 14,15-dihydro-2,3oxidosqualene. O H 2,3-cis-1'nor-2,3oxidosqualene. O 10,15-demethyl-2,3oxidosqualene. O. H. H. 2,3-trans-1'nor-2,3oxidosqualene. Figure 1.8 Some substrate derivative inhibitors.. 10. H. 1,1'-bisnor-2,3oxidosqualene.
(30) O N+. N+. H. H. O (6E or 6Z)-10-aza-10,11-dihydro-squalene 2,3-epoxide. H. O. H. O. (18Z or 18E)-29-methylidene-2,3-oxidosqualene Figure 1.9 The geometrical isomers of substrate analogs.. The argument about a stepwise or a concerted mechanism for the oxidosqualene cyclization has also been considered as the result of direct experimental evidence. After the initial epoxide ring opening, the anchimeric assistance from the neighboring double bond (C-6/C-7) has been observed for the formation of the A-ring.30,31 Subsequently, the non-concerted pathway including a series of conformational rigid and partial cyclic cationic intermediates have been proposed for the remaining process of cyclization cascade.32,33 The bicyclic to pentacyclic triterpenoid products corresponding to the truncated cyclization intermediates were isolated from different natural sources such as P. resins or T. pyriformis, indicating the possibility of a stepwise mechanism for the oxidosqualene cyclization.34,35 The nonstop, concerted cyclization hypothesis, i.e., conversion from oxidosqualene into lanosterol process without mono-, bi-, or tricyclic carbocationic intermediates, was further against by 11.
(31) Corey and coworkers in 1995.36 A minor product with a 6-6-5-4 tetracyclic scaffold was isolated from the enzymatic 20-oxa-2,3-oxidosqualene transformation (Figure 1.10). Thus, Corey proposed that the cyclization of 2,3-oxidosqualene for the tetracyclic protosteryl cation must proceed through a discrete tricyclic 6-6-5 Markovnikov carbocationic intermediate and a sequential ring expansion process with the formation of an anti-Markovnikov cyclohexyl C-ring intermediate.36 Several truncated cyclization products with a 6-6-5 scaffold obtained from the enzymatic incubation with substrate analogs further supported the evidence for the C-ring expansion (Figure 1.10).37-39 Moreover, Corey and coworkers also illustrated the stereochemistry of C-20 during the oxidosqualene cyclization process.40,. 41. The. 20-oxa-2,3-oxidosqualene and the (20E)-20,21-dehydro-2,3- oxidosqualene were respectively synthesized and used in the enzymatic transformation to produce protosteryl derivatives with 17β side chain conformation (Figure 1.11). These experimental results excluded the previously hypothesized 17α side chain conformation, and also facilitated the understanding of a natural R-form configuration at C-20. After the formation of the 17β side chain, the smaller rotation angle (60°) of the side chain is required for producing the natural R-form configuration at C-20. The hindered rotation angle (120°) was required while the side chain conformation appeared with the 17α conformation. Therefore, after decades of efforts by chemists with the representative chemical studies on the respective stages of cyclization processes, the broadly accepted mechanistic hypothesis for the formation of lanosterol is summarized in Figure 1.12.. 12.
(32) O H. O. H. Sce ERG7. (minor) HO. O. H Pig OSC. (minor). HO. O. Figure 1.10 Minor products with a 6-6-5-4 tetracyclic scaffold and a 6-6-5 tricyclic scaffold were isolated from the enzymatic transformation with various analogs. O H. O. H. Sce ERG7 HO. O. OH H Sce ERG7 HO. O H O. 2,3-oxidosqualene. Enzymatic cyclization. HO H. 17β-protosteryl carbonium ion. Figure 1.11 Cyclization of oxidosqualene analogs produce 17β-protosteryl derivatives. 13.
(33) H+ O Prefolded substrate conformation Epoxide Opening/ A-Ring Closure HO. O H+. Monocyclic C-10 Cation. Oxidosqualene. Markovnikov C-Ring Closure. B-Ring Closure. HO. HO 6,6,5-Tricyclic C-14 Cation. Bicyclic C-8 Cation. Anti-Markovnikov C-Ring Expasion. D-Ring Closure. HO. HO Protosteryl C-20 Cation H. 6,6,6-Tricyclic C-13 Cation H Side Chain Rotation. H. H Rearrangement. HO. HO. Lanosteryl C-8/C-9 Cation H. H B: H. H. H. Deprotonation. HO. HO. Lanosteryl C-8/C-9 Cation. Lanosterol. Figure 1.12 The hypothesized cyclization cascade of oxidosqualene.. 14.
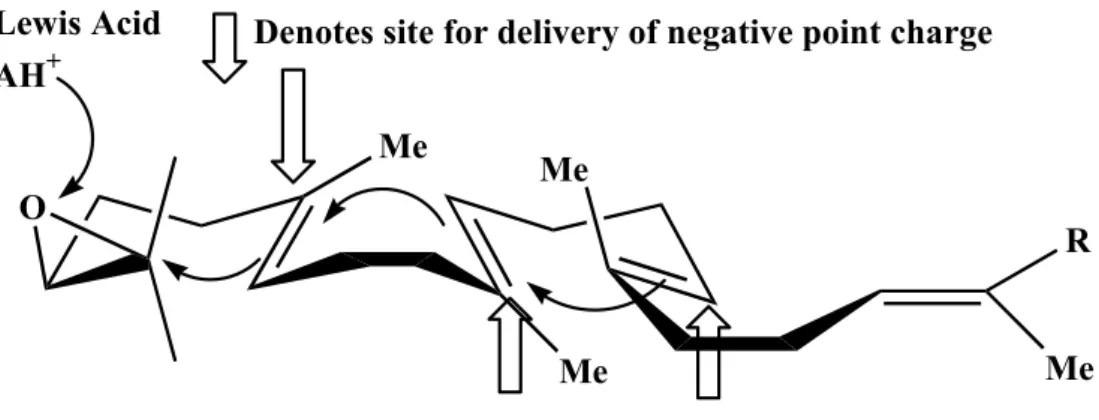
(34) 1.2.2 The theoretical enzyme-templated cyclization reactions In addition to chemical studies on the cyclization mechanism, the critical issues concerning the enzyme itself historically have been poorly understood. After observation of the common structural feature of triterpene molecules, Ourisson proposed a phylogenetic hypothesis for the cyclases.42 It was proposed that a primitive ancestral cyclase containing several essential catalytic residues exists, yet some of the critical active site residues have undergone changes, indicating evolutionary divergence for different species.43,44 Moreover, according to the studies of substrate specificity and the stereochemistry of the cyclization reaction, Johnson has proposed another theoretical model for illustrating the functional role of cyclase enzyme. One Lewis acid residue, which acts as the proton donor for epoxide ring opening, and three axial negative charge residues facing the highly energetic intermediate cations were proposed for the enzymatic cation-olefin cyclization.45,46 These negative charge residues might lower the activation energy for the formation of the boat B-ring and the anti-Markovnikov closure of C-ring (Figure 1.13).. Lewis Acid AH+. Denotes site for delivery of negative point charge Me. O. Me R. Me. Me. Figure 1.13 Johnson proposed three cation-stabilizing auxiliaries in the cyclase.45, 46. 15.
(35) In addition, the “Q-W motif theory” was the hypothetical result, according to the careful comparison of the amino acid sequence’s constitution in the bacterial cyclases as well as in the eukaryotic cyclases.47,48 Sixteen amino acid repeats, Arg/Lys Gly/Ala X2-3 Tyr/Phe/Trp Leu X3 Gly X2-5 Gly X Trp, were highly conserved within the carboxyl region of cyclase gene. One of the Q-W motifs was further labeled with a mechanistic inhibitor, 29-methylidene-2,3-oxidosqualene.49,50 These Q-W motifs were considered for stabilizing the electron-deficient cationic intermediates and for the structural importance in the cyclase enzyme.51 In addition to the “Q-W motif theory”, Griffin proposed the “Aromatic hypothesis” for the cyclase enzymatic active site.52 They suggested that the electron-rich indole rings from tryptophan residues or the phenyl groups from tyrosine/phenylalanine residues, which occurred frequently in the Q-W motifs, are more suitable for the role of stabilizing delivery force than those negative point charges postulated by Johnson. In this “Aromatic hypothesis”, two electron-rich residues are assumed to occupy the position above or below the high energy cationic intermediate in each stage of cyclization processes (C-2, C-8, C-13, and C-20).52 The cation-π interaction in the enzymatic active site is theoretically responsible for stabilizing these electron-deficient cationic intermediates. Therefore, according to these theoretical models, cyclases from different origins might be very similar in their major structural conformation. However, for the purpose of the phylogenetic derivation, small but critical mutational changes exist in these cyclase enzymes, especially in the region spatially near the carbocationic intermediates (Figure 1.14).. 16.
數據

+7
Outline
The mechanistic and stereochemical insights of oxidosqualene cyclization From the view of chemical reaction, diverse products should be obtained after From the view of chemical reaction, diverse products should be obtained after
Biological studies of 2,3-(oxido)squalene cyclases
Crystallization and structural characterization of cyclase
The construction of novel gene disruption strain TKW14 for analysis of inactive mutated cyclase inactive mutated cyclase
Homology modeling illustration of the functional critical residue
Functional analysis of chimeric enzymes library via plasmid shuffle and the product characterization product characterization
Comparison between homology models and mechanistic hypothesis
Correlation of the OSC model to the experimental mutagenesis studies
Conclusion and Future Perspective
Bovine Liver
相關文件
The first row shows the eyespot with white inner ring, black middle ring, and yellow outer ring in Bicyclus anynana.. The second row provides the eyespot with black inner ring
Health Management and Social Care In Secondary
printing, engraved roller 刻花輥筒印花 printing, flatbed screen 平板絲網印花 printing, heat transfer 熱轉移印花. printing, ink-jet
• Content demands – Awareness that in different countries the weather is different and we need to wear different clothes / also culture. impacts on the clothing
(1) Western musical terms and names of composers commonly used in the teaching of Music are included in this glossary.. (2) The Western musical terms and names of composers
Other than exploring the feasibility of introducing a salary scale for KG teachers, we also reviewed the implementation of the Scheme in different areas including funding
• Strange metal state are generic non-Fermi liquid properties in correlated electron systems near quantum phase transitions. • Kondo in competition with RVB spin-liquid provides
We have also discussed the quadratic Jacobi–Davidson method combined with a nonequivalence deflation technique for slightly damped gyroscopic systems based on a computation of
