中 國 醫 藥 大 學
醫 學 檢 驗 生 物 技 術 學 系 碩 士 班 碩士學位論文
Stenotrophomonas maltophilia 的 ampR-L2 組裝
之特性分析
Characterization of ampR-L2 module of Stenotrophomonas maltophilia
指導教授:楊翠青 博士
Tsuey-Ching Yang, Ph.D
研究生:黃奕瑋
Yi-Wei Huang
中 華 民 國 九 十 八 年 一 月
中國醫藥大學
醫技系碩士班
士 碩 學位 論文
Stenotropho
monas maltophilia的ampR-L2組裝之特性分析
研究 生 奕 黃 瑋 撰 十 九 八年 一月
中 國 醫 藥 大 學
醫 學 檢 驗 生 物 技 術 學 系 碩 士 班 碩士學位論文
Stenotrophomonas maltophilia 的 ampR-L2 組裝
之特性分析
Characterization of ampR-L2 module of Stenotrophomonas maltophilia
指導教授:楊翠青 博士
Tsuey-Ching Yang, Ph.D
研究生:黃奕瑋
Yi-Wei Huang
中 華 民 國 九 十 八 年 一 月
誌謝
本論文的撰寫終於接近尾聲了,意味著離畢業之期已不遠矣。回 想起這兩年多來的碩士生活,我只能說其精彩度與偶像劇相比絲毫不 顯遜色,這其中的曲折說來話長,可是點滴在我的心中卻是永生難 忘。還記得剛進實驗室尚懵懂無知我,跟在老師身邊學習,學習老師 的思考模式、還有對待事情專注、認真的態度,著實讓我受益良多。
而平時老師亦師亦母,也很關心我的生活與人格的發展,我好慶幸能 在這個實驗室,所以首先要感謝的就是我的指導教授楊翠青老師,我 要謝謝老師不厭其煩的教育我、包容我,讓我學會以更寬廣的視野去 看事情、做研究。還要謝謝醫技系林振文老師與張益銍老師不時的關 心、指教。我亦感謝研究室的許多伙伴們:冠傑學長和盈如學姐的指 導與鼓勵;一 路 上 相 互 扶 持 的 凱弘;紹晟、瑜姿熱心的幫忙;還有 許多學長姐、同學、學弟妹們的關心,謝謝你們!因為你們,做研究 的日子並不枯燥乏味。我還要感謝一直很關心我,給我溫暖的舅舅、
舅媽、外婆,以及在天上的外公。 最後,最要感謝的就是我的家人,
尤其是最疼愛我媽媽,因為你們的支持,使我可以無後顧之憂地朝著 夢想步步邁進。
奕瑋謹誌
中華民國九十八年一月
主 目 錄
誌 謝 ... I 主 目 錄... II 圖 目 錄... IV 表 目 錄... VI 中文摘要 ... VII Abstract ... IX
第一章 前言 ... 1
第一節 研究背景 ... 1
第二節 Stenotrophomonas maltophilia 的介紹 ... 2
第三節 S. maltophilia 的抗藥性機轉 ... 3
第四節 β-lactamase 的分類 ... 5
第五節 β-lactam 型抗生素 ... 6
第六節 S. maltophilia 的 L1 與 L2 β-lactamase ... 7
第七節 染色體形式的 ampR-ampC 系統 ... 9
第八節 染色體形式的 ampR-class A β-lactamase 系統 ... 12
第九節 S. maltophilia 染色體上之 ampR-L2 系統 ... 13
第十節 研究目的 ... 14
第二章 研究方法 ... 16
第一節 研究設計 ... 16
第二節 研究材料 ... 17
第三節 實驗方法 ... 21
第三章 研究結果 ... 40
第四章 討論 ... 54
圖 ... 65
表 ... 93
參考文獻 ... 106
附錄 ... 112
圖 目 錄
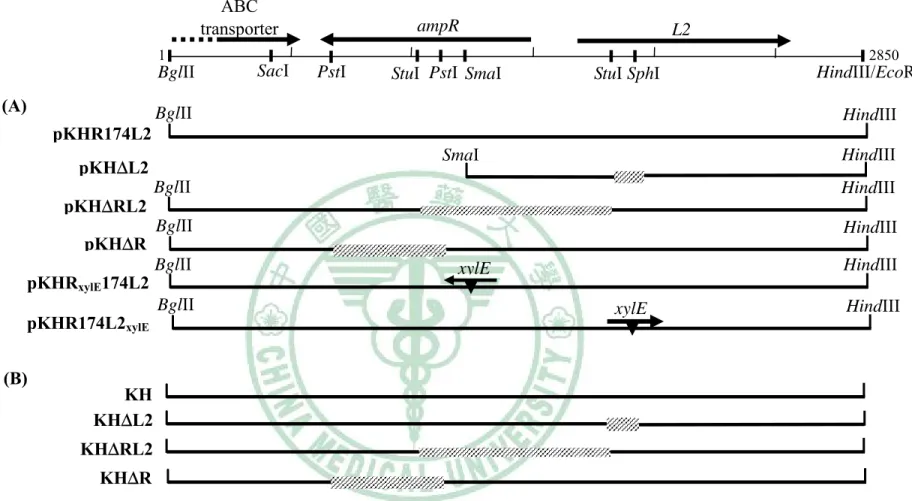
圖一 Restriction enzyme map of a segment of the S. maltophilia KH
chromosome surrounding the ampR-L2 module ... 65
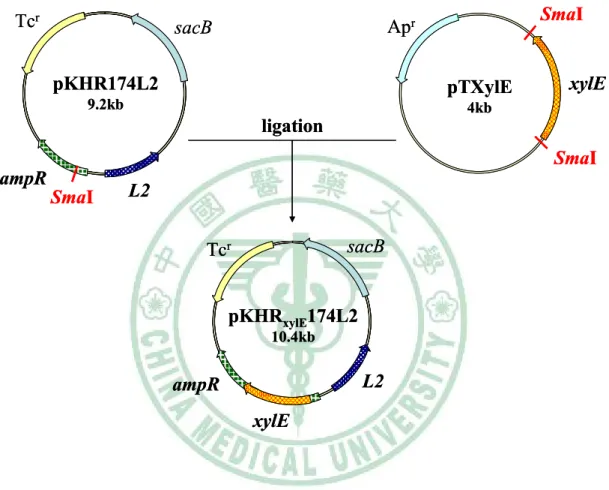
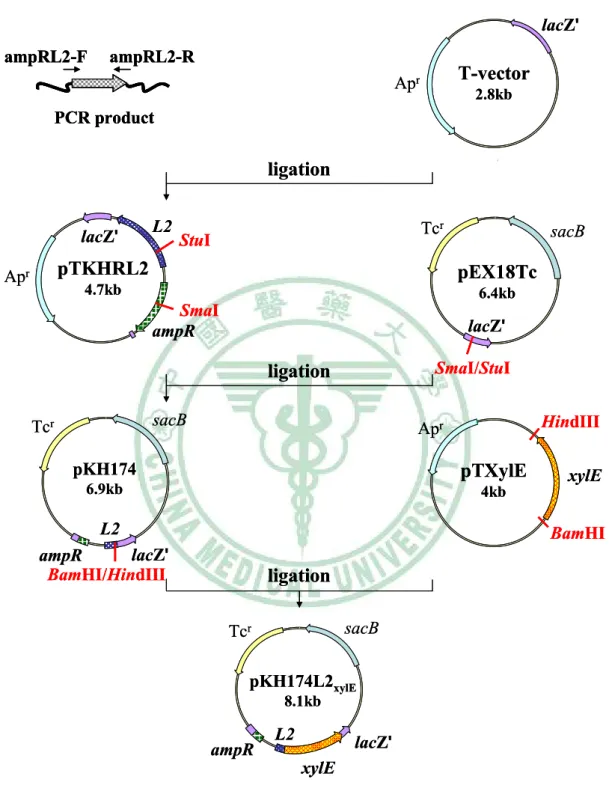
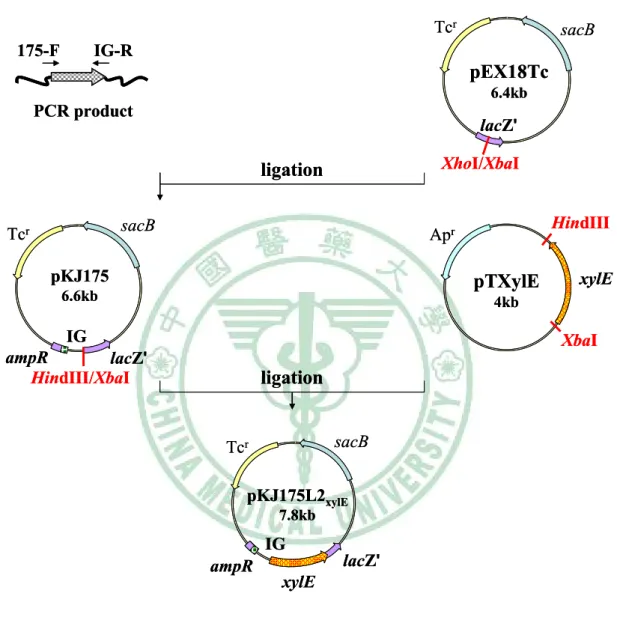
圖二 Construction of pKHRxylE174L2 ... 66
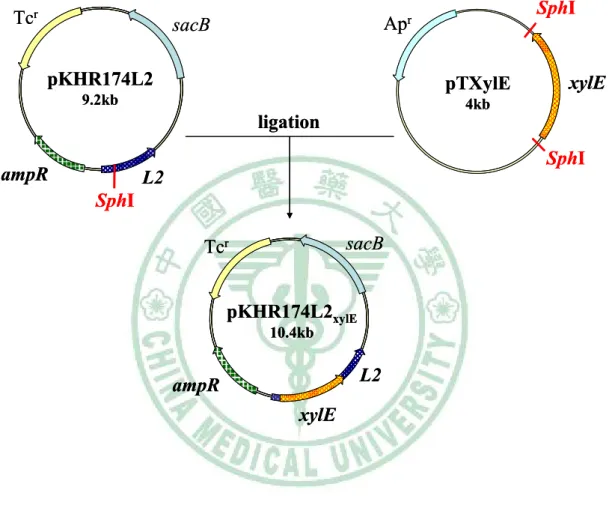
圖三 Construction of pKHR174L2xylE ... 67
圖四 Construction of pKHΔRL2 ... 68
圖五 Construction of pKHΔR ... 69
圖六 Construction of pKHΔL2 ... 70
圖七 Construction of pKHΔL1 ... 71
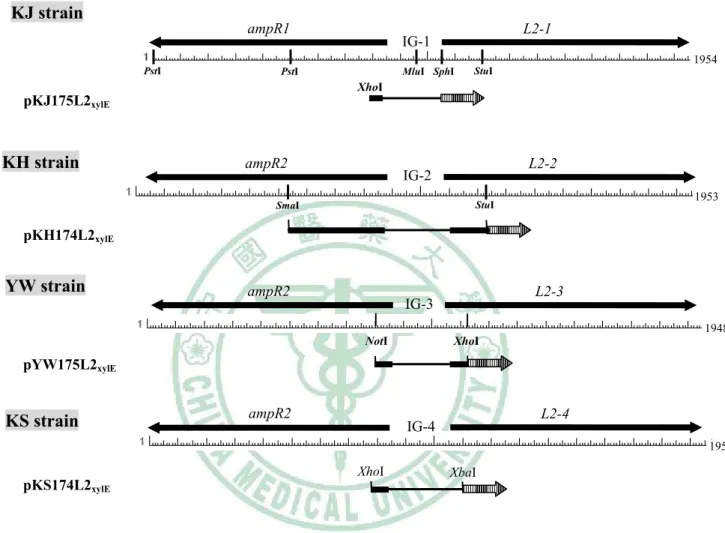
圖八 Restriction enzyme maps of the ampR-L2 modules of the S. maltophilia strains KJ, KH, YW, and KS ... 72
圖九 Construction of pKH174L2xylE ... 73
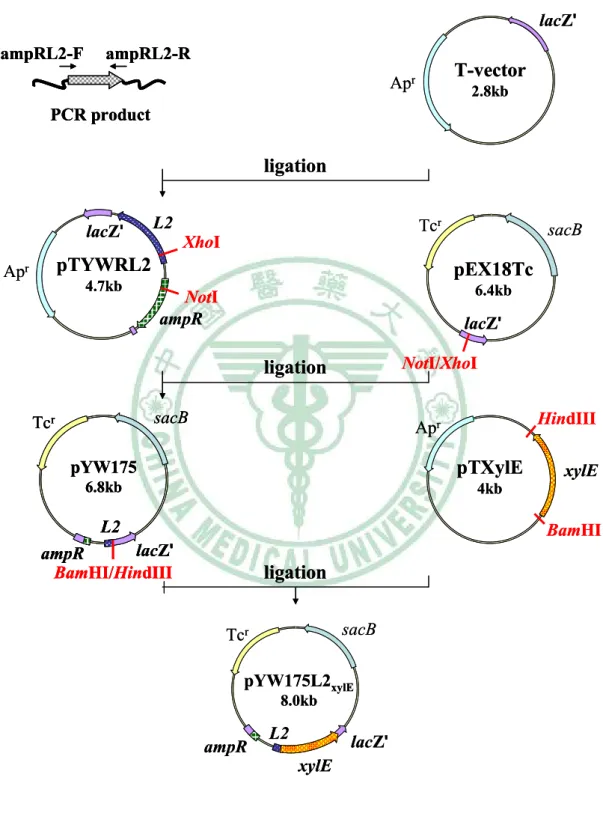
圖十 Construction of pYW175L2xylE ... 74
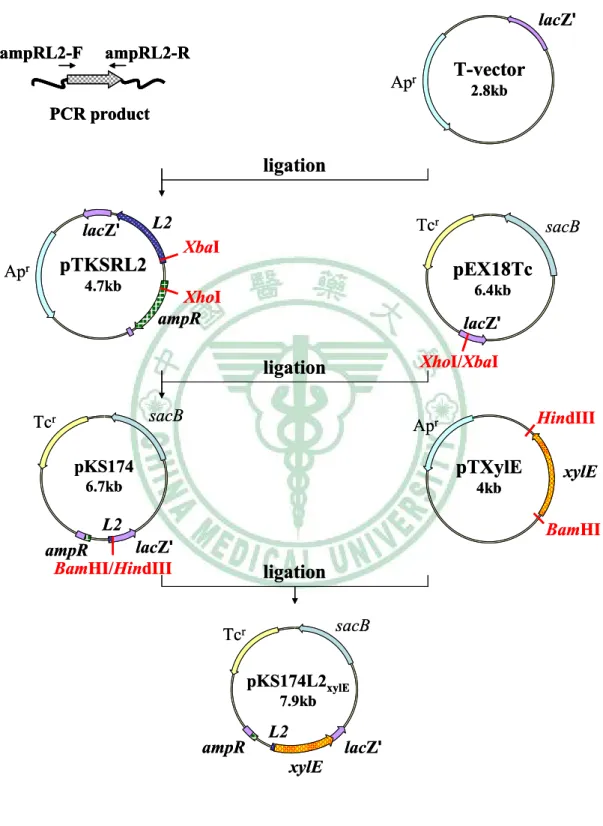
圖十一 Construction of pKS174L2xylE ... 75
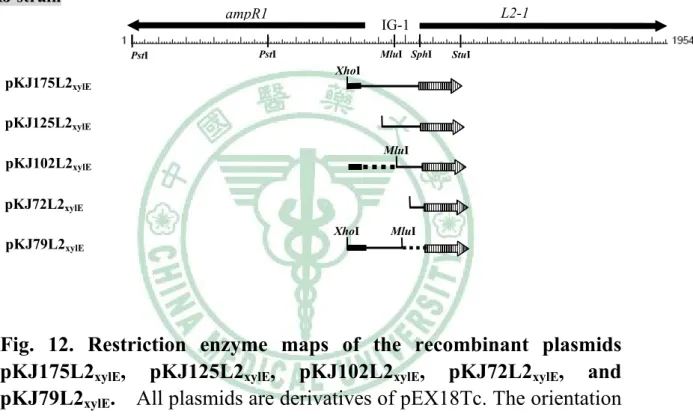
圖十二 Restriction enzyme maps of the recombinant plasmids pKJ175L2xylE, pKJ125L2xylE, pKJ102L2xylE, pKJ72L2xylE, and pKJ79L2xylE ... 76
圖十三 Construction of pKJ175L2xylE ... 77
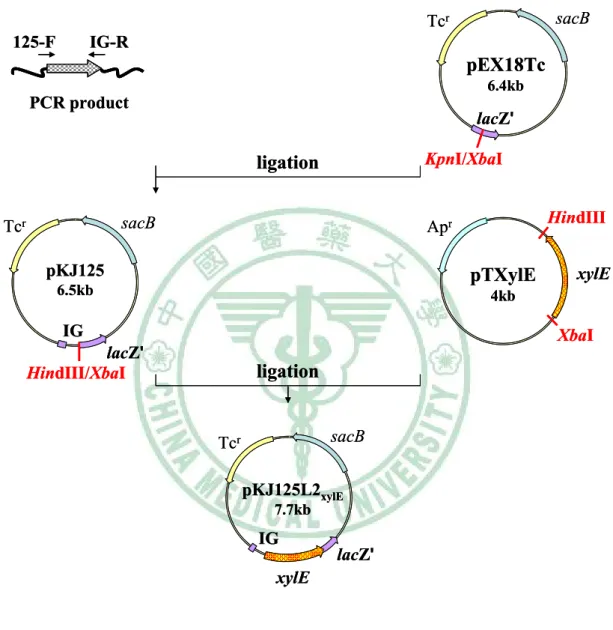
圖十四 Construction of pKJ125L2xylE ... 78
圖十五 Construction of pKJ72L2xylE ... 79
圖十六 Construction of pKJ79L2xylE ... 80
圖十七 Construction of pKJ102L2xylE ... 81
圖十八 Restriction enzyme maps of the recombinant plasmids pKJΔR, pKJ175R, pKJ79R, and pKJ36R ... 82
圖十九 Construction of pKJ175R ... 83
圖二十 Construction of pKJ79R ... 84
圖二十一 Construction of pKJ36R ... 85
圖二十二 Construction of pKJΔR ... 86
圖二十三 Dendrograms showing the relationship of L2 and AmpR proteins of S. maltophilia KH with related β-lactamases and AmpR proteins ... 87
圖二十四 Alignment of ampR-β-lactamase intergenic regions (IGs) with that of S. maltophilia KH ... 88
圖二十五 Dendrograms showing the relationships of L2 β-lactamase proteins and ampR-L2 IG regions among sixteen S. maltophilia isolates ... 89
圖二十六 Protein sequences alignment of the S. maltophilia AmpR proteins from sixteen different clinical isolates ... 90
圖二十七 DNA sequences alignment of ampR-L2 IG regions from S. maltophilia strains YW, KJ, KH, and KS ... 92
表 目 錄
表一 Bacterial strains, plasmid, and primers used in this study ... 93 表二 The C23O activity of E. coli and S. maltophilia harboring
recombinant plasmid pKHR174L2xylE ... 96 表三 The β-lactamase activity of S. maltophilia mutants KHΔL2 and
KHΔRL2 ... 97 表四 The β-lactamase activity of S. maltophilia mutants KHΔR and
KHΔL1 ... 98 表五 The C23O activity of S. maltophilia KH and KHΔRL2
harboring recombinant plasmid pKHRxylE174L2 ... 99 表六 Characteristics of L2 proteins, IG regions and AmpR proteins
of ampR-L2 modules from sixteen S. maltophilia isolates ... 100 表七 Expression of C23O activity and MICs in S. maltophilia KJ and
KH harboring different recombinant plasmids ... 101 表八 Expression of C23O activity in S. maltophilia KJ harboring
different recombinant plasmids... 102 表九 The MIC values of S. maltophilia strains harboring different
recombinant plasmids ... 103 表十 The MIC values of S. maltophilia KJΔR harboring different
recombinant plasmids ... 104 表十一 The MIC values of S. maltophilia strains harboring different
recombinant plasmids ... 105
中文摘要
Stenotrophomonas maltophilia 可 產 生 兩 種 可 被 誘 發 表 現 的
β-lactamases 蛋白,分別為 L1 和 L2 蛋白。而 L1 與 L2 基因的表現通 常受 LysR 型轉錄調控因子 AmpR 蛋白所調控。在分類上,L2 基因 屬於class A 型 β-lactamase,而 ampR 基因位於 L2 基因的上游形成一 ampR-L2 組裝,且兩基因間含有一 174(或 175)-bp 的 intergenic(IG)
區域,此組裝與腸內菌屬的ampR-ampC 組裝相似。 L1 蛋白則屬於 class B 型 β-lactamase,但其鄰近並無類似 ampR 之調控基因。
本論文的第一部份是將S. maltophilia KH 的 ampR-L2 組裝與其它 微生物的 ampR-
β
-lactamase 組裝做進一步的分析,比較其 AmpR 蛋 白、β-lactamase 蛋白和 ampR-β
-lactamase IG 片段。在所有被分析的 菌株中,S. maltophilia 與 Xanthomonas campestris 的演化關係最為相 近。接著在評估 AmpR 蛋白對於 L1 與 L2 基因表現所扮演的調控角 色方面發現,沒有誘發物的情況下,AmpR 蛋白可活化 L1 基因的表 現,抑制 L2 基因的表現;然而在有誘發物的情況下,AmpR 蛋白對 於 L1、
L2 兩基因表現皆扮演著活化者的角色。另外 L1、
L2 兩基 因的誘發表現需要依靠 AmpR 蛋白的幫助(AmpR-dependent),而 ampR 基因的轉録作用是微弱且持續表現的,並無自我調控的現象。本論文的第二部份將 16 株 S. maltophilia 臨床分離菌株的
ampR-L2 組裝進行分析,並且將其 AmpR 蛋白、ampR-L2 IG 區域和
L2 蛋白分組。結果發現,其 L2 蛋白和 ampR-L2 IG 區域之變異度高 達32%,然而 AmpR 蛋白序列在這 16 株 S. maltophilia 分離菌株中卻 被高度保留。接著利用縮短 IG 區域的 DNA 片段來鑑定足以誘發 L2 基因表現之區域和ampR 基因之啟動子區域。結果證實 S. maltophilia 之 ampR 基因的啟動子與 AmpR 蛋白的結合區域並無重疊,說明 AmpR 蛋白對其 ampR 基因並無自我調控。然後將不同的 IG-L2xylE
transcriptional fusion 質體送入 S. maltophilia 原始菌株中以評估 AmpR 蛋白對不同的 ampR-L2 IG 區域之誘發能力。另外,將一段突變之 102-bp 的 IG 片段送入 S. maltophilia 原始菌株中,即可同時減弱其染 色體上L1 和 L2 基因之表現,此 oligo-nucleotides attenuator 可發展為 控制S. maltophilia 菌株 β-lactam 抗藥性之新策略。
Abstract
Stenotrophomonas maltophilia is known to produce at least two chromosomal-mediated inducible β-lactamases, L1 and L2. Expression of the L1 and L2 β-lactamase genes responsible for β-lactams resistance is generally regulated by a LysR type regulator of AmpR. Gene L2, which encodes a class A β-lactamase, and the ampR gene is located immediately upstream of L2 and is transcribed divergently, forming an ampR-L2 module with a 174 (or 175)-bp intergenic (IG) region. L1 belongs to the class B β-lactamase and has no neighbor ampR-like regulatory gene.
Firstly, the ampR-L2 module from S. maltophilia KH was compared with ampR-β-lactamase modules from several microorganisms with respect to the AmpR and β-lactamase proteins and the intergenic (IG) region. S. maltophilia and Xanthomonas campestris showed the most closely phylogenetic relationship among the microorganisms considered.
The regulatory role of AmpR towards L1 and L2 was further analysed. In the absence of inducer, AmpR acted as an activator for L1 expression and as a repressor for L2 expression, whereas AmpR was an activator for both genes in an induced state. In addition, inducibility of L1 and L2 genes depended on the presence of AmpR. The ampR transcript was weakly and constitutively expressed, but was not autoregulated.
Secondly, the ampR-L2 modules of sixteen S. maltophilia isolates were analyzed in respect of AmpR protein, L2 protein, and the IG region.
The results demonstrate that L2 proteins and IG regions show a significantly genetic diversity (up to 32% diversity) among S. maltophilia species, whereas AmpR proteins are highly conserved with identity of
98%. The DNA segments in the IG region, which are sufficient to support the L2 induction and ampR mutant complementation, were identified by deletion constructs assays. The AmpR binding domain and the ampR promoter were found to be not overlapped in the ampR-L2 IG region of S.
maltophilia, signifying the fact that ampR gene is hardly autoregulated.
Furthermore, the induction potential of AmpR toward the different ampR-L2 IG regions was evaluated by introducing the various IG-L2xylE transcriptional fusion constructs into a wild S. maltophilia strain. The induction levels achieved in the various AmpR-IG pairs display marked quantitative differences. In addition, introduction of a 102-bp mutated ampR-L2 IG segment into the wild strain attenuates the expression of chromosomal L1 and L2 genes. A concept of oligo-nucleotides attenuator for the development of antimicrobial agent is proposed.
第一章 前言
第一節 研究背景
自一九二八年英國科學家 Alexander Fleming 發現青黴素後,隨 著時代的進步,有越來越多種抗生素陸續被發現、合成並大量使用於 醫療、畜牧養殖業上。但是「物競天擇,適者生存」,細菌在人類使 用近一世紀抗生素的歲月裡,為了繼續生存,已陸續發展出許多抗藥 機轉來抵抗抗生素的威脅。因此,細菌抗藥性的產生,不僅是公共衛 生上的一大問題,同時也加重人類對疾病治療和新藥研發的壓力與困 難度。
目前在臨床醫療中,我們特別注意造成院內感染排行前幾名的革 蘭氏陰性桿菌,如綠膿桿菌(Pseudomonas aeruginosa)、嗜麥芽醣 黃 單 胞 菌 ( Stenotrophomonas maltophilia ) 、 鮑 氏 不 動 桿 菌
(Acinetobacter baumannii)等非葡萄糖發酵性細菌,其與生俱來之多 重抗藥性,使得在治療上十分困難。因此,研究細菌抗藥性機轉,進 而發展出能對抗細菌抗藥性的藥物或治療方法,實為當前迫切之課 題。
本實驗室以 Stenotrophomonas maltophili 作為研究菌種,並致力 於該菌產生β-lactamase 抗藥性機制之探討。至目前為止,已累積相當
的成果。
第二節 Stenotrophomonas maltophilia 的介紹
Stenotrophomonas maltophilia 最早從環境中分離出來時,被歸類
命名為 Pseudomonas maltophilia(Hugh R. et al., 1961)(1),後來改分 類命名為 Xanthomonas maltophilia(Sutter VL. 1968)(2)。直到近幾年 拜分子生物學技術進步之賜,經科學家重新分析其遺傳資訊和DNA 序列,最後又更名為 Stenotrophomonas maltophilia(Palleroni NJ. et al., 1993)(3)。S. maltophilia屬於葡萄糖非發酵性革蘭氏陰性桿菌,具有單 極鞭毛,廣泛存在於環境中。該菌嗜以麥芽醣為主要之營養碳源,可 以耐受 silverlined catheters(Ubeda P. et al., 1998)(4),其生長溫度範 圍從4~40℃,而最適宜的生長溫度約35℃。S. maltophilia 也普遍生存 在人體中,為常見的 normal flora。一般在健康的正常人身上並非致病 菌,但是在醫院中卻經常在免疫力低下的病人檢體裡被分離出來,許多 的感染症狀和該菌皆有相關性。這是因為在醫院中,S. maltophilia 會 藉由許多媒介物,如處方用的隱形眼鏡鏡片、各類治療插管、空調系統 等,或是因進行器官移植而使用之免疫抑制藥物,而感染免疫力缺陷或 低下之病患,並造成院內感染。導致病人產生呼吸道感染、菌血症、
心內膜炎和尿道感染(Maningo E. et al., 1995)(5)。一些研究認為,
S. maltophilia 所產生的胞外蛋白酶,可能會破壞受感染的組織,是一
種重要的致病因子(Windhorst S. et al., 2002)(6)。S. maltophilia對許 多抗生素皆具抗藥性,如 β-lactam、aminoglycoside、macrolide 類,
這使得 S. maltophilia 在臨床治療上一直是棘手的問題。
關於S. maltophilia 的研究,目前有兩株 S. maltophilia 菌株已完 成 其 基 因 體 定 序 。 一 株 為 S. maltophilia R551-3
(http://www.ncbi.nlm.nih.gov/),G+C content 為 66%,序列全長為 4,544233 bp ; 另 一 株 為 S. maltophilia K279a
(http://www.sanger.ac.uk/Projects/S_maltophilia/ ) 。 G+C content 為 66.32%,序列全長為 4,851,126 bp。
第三節 S. maltophilia 的抗藥性機轉
Stenotrophomonas maltophilia 目前已知有四種抗藥性機制。第一
種是產生可分解或抑制抗生素活性之酵素,例如產生 β-lactamase,
其可破壞 β-lactam 類抗生素的環狀結構,使 S. maltophilia 能夠抵抗 β-lactam 類抗生素的威脅(Richmond MH. et al., 1973)(7)。目前已知 S. maltophilia 能製造出兩種不同型的 β-lactamases,分別為 L1 和 L2
(Krueger TS. et al., 2001)(8)。另外,亦發現S. maltophilia 的染色體 基因上也存在著其他抗藥性基因,能產生水解抗生素的酵素,像是 aminoglycoside acetyltransferases ( Lambert T. et al., 1999 )(9)和
erythromycin inactivating enzyme(Alonso A. et al., 2000)(10)。第二 種是細菌細胞內自行修飾抗生素所辨認的受器標的,例如 β-lactam 類抗生素的殺菌原理主要是藉由抗生素與細菌之 penicillin binding protein(PBP)結合,進而抑制菌體細胞壁的合成,以達到殺菌的效 果。而細菌可透過修飾、改變PBP(penicillin binding protein)的結 構,使抗生素無法與 PBP 結合作用,因而產生抗藥性(Livermore DM.
1995)(11)。第三種抗藥機制是細菌改變自身細胞膜的通透性,使抗生 素難以進入到細胞膜內作用,像S. maltophilia 等革蘭氏陰性菌能改變 細胞膜上的porin,使抗生素無法順利進入細胞內部 (Valdezate S. et al., 2001)(12)。第四種抗藥性機轉則是細菌在細胞膜上發展出特殊的 efflux pump(multidrug pump)system 結構,使抗生素一進入到細胞 膜內部則馬上藉由 pump 打到細胞膜外部 (Li XZ. et al., 2002;
Sanchez P. et al., 2002; Zhang L. et al., 2001)(13)(14)(15)。目前在 S.
maltophilia 已發現兩種多重抗藥排出幫浦系統,分別為 SmeABC 和
SmeDEF 兩型 (Morita Y. et al., 2006)(16)。SmeABC 與 SmeDEF 運作 機制相當類似,它們是細菌消耗自身的ATP 能量或使用鈉-鉀離子差能
(gradient)驅動幫浦而將抗生素自菌體內排出至菌體外。SmeDEF 能 夠將 macrolides、tetracyclines、chloramphenicol 和 quinolones 等多種抗 生素排出菌體外而達到生存的目的。
第四節
β-lactamase 的分類
學術界對 β-lactamase 已發展出許多分類系統,而目前主要沿用 兩種分類系統。一是1986 年提出的 Bush 分類(Bush K. et al., 1986)
(17),另一種是1980 年提出的 Ambler 分類(Ambler RP. 1980)(18)。 Bush 分類法主要是依據受質(Substrate)種類與其活性受何種抑制物
(Inhibiter)抑制這兩種主要因素,分成 Group 1 到 Group 4 四大類。
第 一 類 (Group 1 ) 酵 素 屬 於 cephalosporinase , 其 酵 素 受 質 為 cephaloridine 和 cephalothin,酵素活性不受 clavulanic acid 所抑制。第 二類(Group 2)分為五個亞型,2a 到 2e。Group 2a 屬於 penicillinases;
Group 2b 屬 於 broad-spectrum 和 extended-broad-spectrum 類 β-lactamases;Group 2c 屬於 carbenicillin-hydrolyzing β-lactamases,
其受質為 cephalosporins;Group 2d 是屬於 cloxacillin-hydrolyzing β-lactamases , 受 質 為 cloxacillin ; 最 後 之 Group 2e 則 是 屬 於 cephalosporin-hydrolyzing β-lactamases 。 而 Group 2a 到 2e 型 的 β-lactamases 活性均會被 clavulanic acid 所抑制。第三類(Group 3)
為 metallo-β-lactamase,其活化型的酵素需要鋅離子的加入,所以又 稱為金屬酵素(metallo-enzyme)。此類型的 β-lactamase 活性不受 clavulanic acid 抑制,但是會被 EDTA 所抑制。最後第四類(Group 4)
為penicillin-hydrolyzing β-lactamases,其特性目前尚未完全定論。而
Ambler 的分類法主要是依據 β-lactamase 的胺基酸序列進行分類
(Ambler RP. 1980)(18),分為 Class A 到 Class D 四類;其中 Class A、
C 和 D 三類是以 serine 作為催化機制之酵素(serine active site enzyme),均具有 S-X-X-K motif 的結構。而 Class B 則是需要鋅離 子才具有活性的酵素,所以會受EDTA 所抑制。科學界於 1995 年將 這兩種主要分類法综合整理成一個完整的總表,並列出具有該分型 之β-lactamase 的菌種與基因。其資料整理於附錄一。
第五節 β-lactam 型抗生素
β-lactam 型抗生素是一種廣泛被使用在醫療上的抗生素,而分子 結構中包含 β-lactam ring 的抗生素均屬於 β-lactam 型抗生素。
β-lactam 類抗生素屬於殺菌劑,其殺菌原理是能抑制細菌細胞壁中胜 肽聚醣(peptidoglycan)的形成,而合成肽聚糖則需要 transpeptidase
(penicillin-binding protein, PBP)、carboxypeptidase 和 transglycosylase 等酵素。其中在細菌細胞膜上的 PBP 就是 β-lactam 型抗生素的作用 標 的 。 目 前 β-lactam 型 抗 生 素 主 要 分 為 四 大 類 : penicillin 、 cephalosporin、carbapenem 和 monobactam。Penicillin 類抗生素有 benzylpenicillin(penicillin G)、oxacillin、ampicilli、carbenicillin、
piperacillin…… 等 。 Cephalosporin 類 抗 生 素 包 括 cephaloridine 、
cephalexin、cephalothin……等。Carbapenem 類抗生素則有 imipenem、
meropenem、faropenem ……等。而 monobactam 類抗生素目前市面上 則主要是aztreonam。
目前發現 β-lactam 類抗生素的副作用包括:腹瀉、頭暈、疹塊、
蕁麻疹、重疊感染(包括念珠菌),偶爾還會導致發燒、嘔吐、紅斑、
皮膚炎、血管性水腫和偽膜性腸炎。β-lactam 類抗生素與 β-lactamase inhibiter 同時使用時,其注射處往往會有疼痛和發炎之情形。臨床上,
約10%的病人對 β-lactam 類抗生素產生過敏,約 5-10%的病人對青黴 素衍生物、頭孢菌素和碳青黴烯類抗生素產生交叉敏感性反應。所以 當病患對一種 β-lactam 類抗生素會產生嚴重過敏反應的話,如果需 要再給予使用其它 β-lactam 類抗生素時就必須慎重考慮(Rossi S. et al., 2004 年)(19)。
第六節 S. maltophilia 的 L1 與 L2 β-lactamase
S. maltophilia 為一革蘭氏陰性桿菌,易感染免疫力低下的病人
(Denton M. et al., 1998 )(20)。 其 可 產 生 兩 種 可 被 誘 發 表 現 之 β-lactamases 蛋白,分別為 L1 蛋白和 L2 蛋白,進而提高 S. maltophilia 對β-lactam 類抗生素之抗藥性(Alonso A. et al., 1997)(21)。目前已知 由S. maltophilia 染色體上產生出來的 β-lactamase 之 N 端大約有 20 個
胺基酸的訊號胜肽(signal peptide)(Walsh TR. et al., 1997)(22)。首 先,在細菌細胞質產生的 β-lactamase 還是屬於前趨物,而到最後成 熟型的 β-lactamase 會被送到細胞膜的外膜與內膜之間。通過細胞內 膜時,β-lactamase 的 signal peptide 會經過修飾作用而被切除,signal peptide 具有一個 hydrophobic core region 方便將 secreted polypeptide 插入內膜,在hydrophobic core region 前 N-terminal 帶有正電荷的胺 基酸,它會與細胞膜上帶有負電荷的phospholipids 互相吸引,而 core region 另一端的 C-terminal 則帶有極性的胺基酸。這時帶正電荷的胺 基酸 N 端與 hydrophobic core region 會形成具有功能性的 signal peptide,在進行修飾作用時會被 signal peptidase 切斷。
L1 β-lactamase 為 一 需 要 二 價 鋅 離 子 ( Zn2+) 才 具 有 活 性 的 metallo-beta-lactamase。它對於抗生素 imipenem 具有很強的水解性,
且酵素活性會受到EDTA 所抑制。目前的文獻指出 L1 β-lactamase 為 一四聚體(tetramer)之蛋白分子,而一個分子可以結合 2 個 Zn2+。 因此一個完整的L1 β-lactamase可以結合8個Zn2+ (Saino Y. et al., 1982)
(23)。依照Bush 分類法,L1 β-lactamase 屬於 Group 3。而依照 Ambler
分類法,則是把L1 β-lactamase 歸類為 Group B。L1 β-lactamase 能水解 的抗生素主要為 penicillin 型抗生素,和部份的 cephalosporin 型抗生 素以及carbapenem 型抗生素,但 L1 β-lactamase 不能水解 monobactam
型抗生素。目前在 S. maltophilia 臨床分離菌株中發現 L1 基因具有 8~12%的變異性,其基因的序列長度約 873 bp,能編碼出 291 個胺基 酸,蛋白分子量約115 kDa,pI 值約為 7,屬於酸性蛋白質(Crowder MW.
et al., 1998)(24)。
而 L2 β-lactamase 主要能水解 cephalosporins 類抗生素,是屬於 serine active-site cephalosporinase。其酵素活性會受到 clavulanic acid、
tazobactam、sulbactamn …等化合物所抑制(Crowder MW. et al., 1998)
(24)。L2 β-lactamase 在 Bush 分類中屬於 Group 2e,能水解的抗生素主
要 為 cephalosporin 型 抗 生 素 , 和 部 份 penicillin 型 抗 生 素 及 monobactam 型抗生素,但無法水解 carbapenem 型抗生素。而在胺基 酸的序列分析中發現 L2 β-lactamase 具有 S-T-D-K、S-D-N、K-T-G 的 高度保留序列(Walsh TR. et al., 1997)(22),以Ambler 之分類為 Group A。目前在 S. maltophilia 臨床分離菌株中發現 L2 基因具有 9~25%的變 異性,其基因序列長度為909bp,能編碼出 303 個胺基酸,蛋白分子量 為31.5 kDa,pI 值約為 8.2,屬於鹼性蛋白質(Avison MB. et al., 2001)
(25)。根據文獻整理 L1 與 L2 β-lactamases 於附錄二。
第七節 染色體形式的 ampR-ampC 系統
革 蘭 式 陰 性 菌 中 常 見 一 LysR 型 轉 錄 調 控 因 子 ( LysR transcriptional regulator)之基因與一可被誘發表現的
β
-lactamase 基因所形成的組裝(ampR-
β
-lactamase module),通常 ampR 基因與其 相鄰的β
-lactamase 基因走向相反,且 ampR 基因與β
-lactamase 基 因中間的基因片段(IG)包含兩基因的啟動子與 AmpR 蛋白結合區 域(Hanson N. et al., 1999)(26)。有兩種不同分型的ampR-β
-lactamase 組 裝 已 被 報 導 , 一 為 ampR-ampC 組裝 ,另一為 ampR-class A β-lactamase 組裝。染色體形式的 ampR-ampC 系統最早是在 Citrobacter freundii
(Lindbery F. et al., 1985) (27)中發現,其後陸續在Enterobacter cloacae
(Honore N. et al., 1986)(28)
、
Yersinia enterocolitica(Seoane A. et al., 1992)(29)、
Morganella morganii(Poirel L. et al., 1999)(30)、
Hafnia alvei(Girlich D. et al., 2000)(31)
、
Ochrobactrum anthropi(Nadjar D. et al., 2001) (32)、
Buttiauxella agrestis (Bauernfeind A. et al., 1996) (33)、
Serratia marcescens(Mahlen SD. et al., 2003)(34) 和 Pseudomonasaeruginosa(Lodge JM. et al., 1990)(35) 中看到其相關報導,這些菌大 多屬於腸桿菌科,報導中對於此系統的調控機制也有相當的了解
(Jacobe C. et al., 1997;Lindberg F. et al., 1987;Normark S. et al., 1995;Wiedemann B. et al., 1998)(36) (37) (38) (39)。以 Citrobacter freundii 為 例,其 ampR-ampC 組裝的誘發機制與胜肽聚醣的循環有關,至少有 三種以上的基因產物與其有關聯,分別為 ampG、ampD 和 ampR 基
因 。 ampG 基 因 產 生 一 穿 膜 蛋 白 , 可 將 細 胞 壁 瓦 解 後 的 anhMurNAc-tripeptide,從細胞間質傳送至細胞質內。ampD 基因產生 一 cytosolic N-acetyl-anhydromuramyl-L-alanine amidase , 可 將 anhMurNAc-tripeptide 水解成 UDP-MurNAc-tripeptide 而促進其循環 再利用。而 C. freundii 的 ampC 基因之誘發機制已被清楚報導,且其 LysR 型轉錄調控因子(LysR transcriptional regulator)如何誘發與其 調控基因相鄰的
β
-lactamase 基因也為一典型的模式。Lindquist et al.(Lindquist S. et al., 1989)(40) 發現 ampR 基因會持續且微弱地表現,
且其產生的 AmpR 蛋白無論有無誘發物的存在,都會直接結合在 IG 區域上,更進一步的清楚指出有種信號 ligand(實際的誘發物)與 AmpR 蛋白的相互作用會決定 ampC 基因表現被抑制或者被誘發;
UDP-MurNAc-pentapeptide 為一抑制 ampC 基因表現之 ligand,反之,
anhMurNAc-tripeptide 使 ampC 基因活化(Hu RM. et al., 2008)(41)。 ampR 基因產生一 LysR 型的調控因子,其可控制 ampC 基因的轉
錄作用,而LysR 型的調控因子通常會結合在其調控基因上游的啟動 子上,再由一小分子(ligand)進一步去活化其調控的基因(Schell MA.
et al., 1993)(42)。另有報導指出,ampR 基因亦與細菌的生長、處於壓 力下之反應、致病因子和抗藥性有關 (Kong KF. et al., 2005;Schell MA. et al., 1993)(42) (43),而LysR 型的調控因子需要結合在 DNA 序列
T-N11-A 上。以 Citrobacter freundii 為例,其 ampR 基因和 ampC 基 因的啟動子位於兩基因之中間區域(IG),走向相反且有重疊之現象,
而LysR-motif T-N11-A 序列位於 ampR 基因的啟動子上(Lindquist S.
et al., 1989)(40),造成了 ampR 基因的負向自我調控(negative autoregulation)現象。在此組裝中,AmpR 蛋白在沒有抗生素的情況 下當作一抑制者,抑制 ampC 基因的表現;而當抗生素存在時,則 扮演著活化ampC 基因的角色(Jacobe C. et al., 1997;Lindbery F. et al., 1985;Lindberg F. et al., 1987)(27) (36) (37)。
除了染色體上的ampR-ampC 組裝,亦有在細菌中發現由質體帶 來的持續表現、非誘發型的ampC 基因(Bradford PA. et al., 1997;
Gaillot O. et al., 1997;Nadjar DM. et al., 2000;Papanicolaou GA. et al., 1990;Philippon AG. et al., 2002)(44) (45) (46) (47) (48),但是最近又有在臨 床菌株中發現帶有ampR-ampC 組裝之質體,而 ampR 基因的存在使 ampC 基因回復其被誘發性 (Barnaud G. et al., 1998;Fortineau N. et al.,
2001;Nakano R. et al., 2004;Reisbig MD. et al., 2002;Verdet C. et al., 2006)(49) (50) (51) (52) (53)。
第八節 染色體形式的 ampR-class A β-lactamase 系統
除 了 ampR-ampC 系 統 外 , 另 有 一 相 似 之 ampR-class A β-lactamase 組裝,此組裝存在於 Rhodopseudomonas capsulate sp108
(Campbell JIA. et al., 1989)(54)
、
Enterobacter cloacae NOR-1(Naas T.et al., 1994)(55)
、
Proteus vulgaris B317(Datz M. et al., 1994)(56)、
Serratia marcescens S6(Naas T. et al., 1995)(57)、
Burkholgeria cepacia 249(Trepanier S. et al., 1997)(58)
、
Xanthomonas campestris(Weng SF. et al., 2004)(59)、
Citrobacter sedlakii 2596(Saino Y. et al., 1982)(23) 和 Stenotrophomonas maltophilia(Hu RM. et al., 2008)(41) 中,此組裝到 目 前 為 止 只 有 存 在 於 染 色 體 形 式 的 被 報 導 。 在 ampR-class A β-lactamase 組裝的 IG 片段與 ampR-ampC 組裝的 IG 片段相較之下,ampR-class A β-lactamase 組裝的 IG 片段更具多樣化,例如 Serratia marcescens S6 的 ampR 基因與 class A β-lactamase 基因之啟動子非但 沒有重疊,還是一面對面走向(face to face)的結構(Naas T. et al., 1995)(57)。除此之外,在ampR-class A β-lactamase 組裝中,其 AmpR 蛋白的調控規則亦有所不同,以Enterobacter cloacae NOR-1(Naas T.
et al., 1994)(55) 和Serratia marcescens S6(Naas T. et al., 1995)(57) 為 例,AmpR 蛋白不管在有無誘發物的情況下,都當作一活化者;而 Burkholgeria cepacia 249 的 AmpR 蛋白在有無誘發物的條件下都扮演 著一抑制者的角色(Trepanier S. et al., 1997)(58)。
第九節 S. maltophilia 染色體上之 ampR-L2 系統
最近,L1 蛋白和 L2 蛋白的誘發表現證明由 LysR 型轉錄調控因
子(LysR transcriptional regulator)AmpR 蛋白所調控 (Okazaki A. et al., 2008)(60)。而ampR 基因位於 L2 基因的上游,且兩基因的走向相 反,形成一ampR-L2 組裝(Hu RM. et al., 2008)(41)。因為ampR-L2 組裝與ampR-ampC 組裝的構造非常相似,所以 AmpR 蛋白和 ampR-L2 IG 區域之間的結合,可能是 L2 基因表現的關鍵。
第十節 研究目的
Stenotrophomonas maltophilia 為一革蘭氏陰性桿菌,廣泛存在於
週遭環境,其容易伺機性的引發院內感染(Denton M. et al., 1998)(20), 此菌已知有兩型由染色體基因所產生的可誘發型 β-lactamases 蛋 白,分別為L1 與 L2 蛋白(Avison MB. et al., 2001;Saino Y. et al., 1984;Saino Y. et al., 1982)(23) (25) (61) 。此兩個
β
-lactamases 基因之 DNA 序列已在實驗室先前的研究中構築與分析(Hu RM. et al., 2008)(41),但是對於 AmpR 蛋白對 L1 基因與 L2 基因的調控機制還不甚清 楚(Okazaki A. et al., 2008)(60)。本論文將研究主題分為兩部份,第 一部份是要更進一步確認S. maltophilia 在有與無誘發物的情況下,探 討 AmpR 蛋白對於 L1 與 L2 基因的調控情形,與評估 AmpR 蛋白是 否有自我調控現象。另外,在對於ampR-ampC 組裝與 ampR-class A β-lactamases 組裝這兩系統作更深入的探討與比較。此外,在 S.
maltophilia 菌株中,ampR 基因與 ampR-L2 IG 區域的序列差異度尚未
有其研究報導,可以合理的預期在不同分離菌株中,其 AmpR 蛋白 和ampR-L2 IG 基因間的差異,可能會影響 L2 基因的誘發能力。所以,
本論文的第二部份將16 株 S. maltophilia 臨床分離株的 ampR-L2 組裝 進行分析,且將其AmpR 蛋白、ampR-L2 IG 區域和 L2 蛋白之差異度 分組。讓不同群組的AmpR 蛋白和 ampR-L2 IG 區域相互作用,測量 其不同組合的誘發能力。同時,說明ampR-L2 IG 之 DNA 片段對 ampR 基因表現和對 L2 基因誘發之情形。最後,評估傳入一段 trans-acting ampR-L2 IG DNA 片段對宿主菌株染色體上之 L1 與 L2 基因的影響。
第二章 研究方法
第一節 研究設計
本論文之第一部份是先利用生物資訊作 ampR-
β
-lactamase 組裝 之分子演化分析。接著構築 S. maltophilia KH 的 ampR -L2 組裝之質 體,且送入 E. coli 系統中觀察其表現情形。然後利用基因替換(gene replacement)的策略,將欲突變之基因 deletion,形成 KHΔL2、KHΔRL2 、 KHΔR 和 KHΔL1 之 突 變 株 , 之 後 定 量 突 變 菌 株 之 β-lactamase 活性以釐清 AmpR 蛋白對 L1 和 L2 基因表現所扮演的角 色。最後將構築好的 pKHRxy1E174L2 質體送入 KH 和 KHΔRL2 突 變株中,測量其C23O 活性以評估 ampR 基因的自我調控。
本論文之第二部份則是先構築 16 株 S. maltophilia 分離菌株的 ampR-L2 組裝之質體,且利用生物資訊作其 ampR-L2 組裝之分子演
化分析。然後構築一系列包含完整 ampR-L2 IG 片段與 xylE 基因形 成之transcriptional fusion 的重組質體,分別送入 S. maltophilia 分離菌 株KJ 和 KH 中測量其 C23O 活性,以評估兩不同型的 AmpR 蛋白對 不同演化分群的ampR-L2 IG 片段之誘發能力。接著構築五個包含不 同長度ampR-L2 IG 片段與 xylE 基因形成之 transcriptional fusion 的 重組質體,分別送入S. maltophilia KJ 菌株中測量其 C23O 活性,以
定位 ampR-L2 IG 區域中可誘發 L2 基因表現之最小區域。利用基因 替換(gene replacement)的策略,將 ampR 基因 deletion,形成 KJΔR 之突變株,然後將重組質體pKJ175R、pKJ79R 和 pKJ36R 送入 KJΔR 突變株中,利用互補試驗以定位 ampR 基因之啟動子區域。利用 error-prone PCR 對 ampR-L2 IG DNA 片段任意突變,篩選能同時降 低L1 和 L2 基因表現之 IG 突變片段。
第二節 研究材料
一. 本論文所構築與分析之質體與菌株列於表一。
二. PCR 引子
實驗中所使用的PCR 引子序列皆詳列於表一,其中 stock solution concentration 為 100 μM;而 working concentration 為 10 μM。
三. 培養基
本實驗使用的培養基購自 MDBio,Inc.(參照 Sambrook et al.
Molecular Cloning: A Laboratory Manual 所記載之配方)。
1. Luria-Bertani broth(LB):每 1 公升的水中含 10 g tryptone
5 g yeast extract 10 g NaCl pH: 7.0±0.05
2. Luria-Bertani agar(LA):Luria-Bertani broth 成份中額外加
入1.5% agar.
3. Mueller Hinton Ⅱ Agar:每 1 公升的水中含 2 g beef extract
17.5 g acid hydrolysate of casein 1.5 g starch
17 g agar
四. 實驗藥品
本實驗菌種所使用的藥品購自 Difco Laboratories 或 Accumedia manufacturers, Inc. 而其他的化學藥品及有機溶劑則購自 E. Merk, J. T.
Backer Company、Severva Frvafeinrobiochemica、Boehringer mannhrim GmhH Biochemical、Pharmacia、Sigma Chemical Company、Biosolve 及日本和光藥廠。限制酶以及其它酵素是購自 TaKaRa Shuzo Co.
Ltd.、New England Biolabs(NEB)、promega company 和 Bethesda Research Laboratories。T4 DNA ligase 購買自 promega Co.。
五. 抗生素
所有抗生素皆購自sigma 公司,並依照實驗所需配置成適當濃度。
六. 試劑與緩衝溶液
1. 質體DNA 抽取之試劑
抽取質體 DNA 之技術是利用 alkaline lysis method 進行,所需試 劑如下:
(1) Solution Ⅰ:10 mM EDTA(pH 8.0),50 mM Glucose 及
25 mM Tris-HCl(pH 8.0)。
(2) Solution Ⅱ:1% SDS 及 0.2 N NaOH。
(3) Solution Ⅲ:3 M potassium acetate(pH 4.8)及 5 M glacial acetic acid。
2. 染色體抽取之試劑
(1) STE buffer : 10 mM Tris-HCl,100 mM NaCl 及 1 mM EDTA
(pH 8.0)。
(2) Proteinase K(10 mg/ml)。
3. Agarose gel 電泳之試劑
(1) TAE running buffer : 預先配製 50X 濃度的 TAE buffer,取 242 g Tris base,57.1 ml galacial acetic acid,100 ml 之 0.5 M EDTA(pH 8.0),加水至 1 L。使用時加水稀釋至 1X,濃 度為40 mM Tris-acetate,1 mM EDTA(pH 8.0)。
(2) 6X loading dye:溶劑中含 0.25% xylene cyanol,30%
glycerol,0.25% bromophenol blue。
(3) Staining buffer:於染盆中先加入少許之蒸餾水,再加入 5 μl 的ethidium bromide(EtBr)、1 ml 1 mM EDTA(pH 8.0)
及 200 μl RNAse A,最後再加蒸餾水至總體積為 150 ml,
充份混合。於使用時染盆中約含有 0.5 μg/ml ethidium
bromide。
4. 檢測 β-lactamase 活性所需試劑
(1) Sodium phosphate buffer, 0.1 M pH 7.0 預先配製
Solution A : 27.6 g NaH2PO4‧H2O per liter(0.2 M)
Solution B : 28.3 g Na2H2PO4‧H2O per liter(0.2 M)
然後再以solution A 9.8 ml 搭配 solution B 15.3 ml 之比例,
使其pH 值為 7.0,最後加去離子水至總體積為 200 ml 即完 成。
(2) Nitrocefin
1 mg Nitrocefin powder 加入 2 ml phosphate buffer solution,
使其完全溶解後,再加入 2 ml 0.1 M pH 7.0 之 Sodium phosphate buffer,使其 final stock concentration 為 0.5 mM,
最後保存於-20℃。
(3) Protein determination dye
Bio-Rad Protein Assay Dye Reagent Concentrate
(4) EDTA(0.5 M pH 8.0)
取146.125 g EDTA 和 67.41 g NaOH,將兩者溶於 1 L 去離 子水中,最後調整pH 值為 8.0。
6. Catechol 2,3-dioxygenease 活性測試所需試劑
(1) Sodium phosphate buffer, 0.1M pH 7.5 預先配製
Solution A : 27.6 g NaH2PO4‧H2O per liter(0.2 M)
Solution B : 28.3 g Na2H2PO4‧H2O per liter(0.2 M)
然後再以solution A 16 ml 搭配 solution B 84 ml 之比例,使 其pH 值為 7.0,最後加去離子水至總體積為 200 ml 即完成。
(2) Assay buffer
取100 ml 0.1 M pH 7.5 sodium phosphate buffer,加上 80 ml 去離子水,再加入 20 ml acetone,配製成 200 ml assay buffer。
(3) 0.1 M catechol
秤取1.1 g catechol 溶至 100 ml assay buffer,使其完全混和 溶解。最後保存於 4℃冷藏備用。
七. 儀器設備
1. 聚合酶連鎖反應器 2. 恆溫培養箱
3. 恆溫乾浴器 4. 離心機 5. 電泳設備 6. 分光光度計
第三節 實驗方法
一. 菌種的培養與保存
1. 短期保存
將實驗用菌株培養於不含抗生素的Luria-Bertani agar(LA)固態 培養基中,置於37℃隔夜培養後,放置 4℃保存 5~7 天,予以備用。
2. 長期保存
將本實驗所用的菌株培養於含有合適抗生素濃度的Luria-Bertani broth(LB)培養液中,37℃震盪隔夜培養。視菌種的生長速度,在 生長對數期間(log phase)取 0.7 ml 的菌液並加入 0.3 ml 87%無菌 glycerol,混合均勻存放於抗凍管中,並在管上貼上標籤註明編號、
菌種名稱和製作日期,存放於-80℃備用。
二. 洋菜膠體電泳分析(agarose gel electrophoresis)
秤取適量之agarose powder 加入 0.5X TAE buffer 中,利用微波爐 加熱使之溶解,agarose 的濃度依欲分析之 DNA 片段大小而定。一般 使用濃度範圍約0.8%~2.0%(w/v)。等agarose 溶液降溫至 50℃~60℃
時將之倒入鑄膠槽並插上齒梳(comb)。待 agarose 溶液冷卻凝固後 緩慢拔除齒梳,即完成agarose gel 製作。將 agarose gel 置於水平式電 泳槽裝置,並加入 0.5X TAE buffer 至淹蓋過 gel 為止。把欲分析之 DNA 樣品與 6X loading dye 以 5:1 混合之比例混勻後,加入 agarose gel 的溝槽(well)內。開啟電源,以 5V/cm 的電壓進行電泳,泳動 的時間視DNA 片段長度而定。待分離完畢後,將 agarose gel 置於含
ethidium bromide(0.5 μg/ml)的染盆中染色 10~15 分鐘後,再置於紫 外燈箱中觀察DNA 泳動的位置並與 DNA marker 比對,以評估 DNA 片段的大小及濃度。
三. DNA 之製備
1. 質體 DNA 之抽取
將含有質體之菌株培養於 3 ml 液態培養基中,可視情況加入合 適濃度之抗生素以預防污染。經 37℃震盪隔夜培養,視菌種的生長 速度,在生長對數期間(log phase)取出,分裝至 1.5 ml eppendorf 中,以12,000 rpm 5 分鐘離心,去除上清液後加入 100 μl solution Ⅰ 溶液,使菌體重新懸浮,靜置於室溫中5 分鐘。接著加入 200 μl solution
Ⅱ溶液,溫和上下倒置數次後,置於冰上5 分鐘。之後再加入 150 μl solution Ⅲ溶液,上下倒置數次後再置於冰上 10 分鐘。接著以 12,000 rpm 離心 10 分鐘,將上清液取至新的 eppendorf 中,再加入 1:1 之 phenol/chloroform 以 vortex 混合均勻,經 12,000 rpm 離心 5 分鐘後取 上清液至另一 eppendorf 中,重複數次。使溶液分界層無雜質後再加 入等量之chloroform 以 vortex 混合均勻,經 12,000 rpm 離心 5 分鐘,
取上清液至新的eppendorf 中加入二倍體積之 95%酒精,用 vortex 混 合均勻後靜置於冰上10 分鐘。最後再以 12,000 rpm 離心 10 分鐘,去 除酒精後得沉澱之DNA,待酒精揮發後用無菌之去離子水回溶備用。
2. 染色體 DNA 之抽取
將菌株培養於 3 ml 液態培養基中,37℃震盪隔夜培養,視菌種 的生長速度,在生長對數期間(log phase),移至 1.5 ml eppendorf 中,以 8000 rpm 離心 5 分鐘,去除上清液後再到入剩餘的菌液離心 5 分鐘後,去除上清液。加入1 ml 之 1X STE buffer,並 vortex 使菌 體重新懸浮,以12,000 rpm 離心 5 分鐘,重複一次此步驟。去上清液 後,先加入 200 μl 1X STE vortex 均勻。接著緩慢加入 40 μl 10%
SDS,慢慢上下倒置 eppendorf,直至澄清。接著靜置 65℃ 30 分鐘,
待降溫後加入Proteinase K(2 mg/ml)20 μl(final conc. 40 ng/ml)
於37℃作用 3~4 小時,接著加入 400 μl 1X STE buffer 放大體積。加 入1:1 比例之 phenol/chloroform 用 vortex 混合均勻,以 12,000 rpm 離心 5 分鐘後取上清液至另一 eppendorf 中,重複此步驟數次使溶液 分界層無雜質。再加入等量之chloroform 用 vortex 混合均勻,以 12,000 rpm 離心 10 分鐘,取上清液至另一 eppendorf 中加入二倍體積之 95
%酒精,用 vortex 混合均勻後置於冰上 10 分鐘,最後利用 tip 捲出染 色體晾乾,再加入無菌去離子水 200 μl 回溶,以 65℃加熱 30 分鐘去 除DNase 後,置於 4℃保存備用。
3. DNA 片段之回收
DNA 片段的回收使用 GeneMark DNA Clean/Extraction Kit,將要
分離的 DNA 片段先以洋菜膠體電泳分離,於 EtBr 染色 10 分鐘,置 於紫外線箱上觀察。使用刀片將目標片段切取下來放入 eppendorf 中,加入 500 μl Binding solution 置於乾浴器 65℃ 30 分鐘,時而上下 倒置搖晃。再吸取溶液至Kit 提供的 Spin column 中,Spin column 下 接Collection column,以 12,000 rpm 離心 3 分鐘。再加入 500 μl Washing Solution 12,000 rpm 離心 5 分鐘,重複此步驟二次。打開 Spin column 蓋子等待 3 分鐘使酒精揮發,最後加入適量無菌去離子水於 Spin column 中,靜置數分鐘後以 12,000 rpm 離心 10 分鐘,即得回收產物。
回收所得之DNA 保存於 4℃備用。
4. DNA 切割反應
選擇合適的限制酶,配合廠商建議的緩衝溶液,和所建議的反應 溫度,進行DNA 切割反應。反應時的 DNA 濃度約為 1 μg/50 μl,而 反應時間則視選擇的限制酶而調整,完成後利用洋菜膠體電泳分析切 割情形。
5. DNA 黏合反應(ligation)
將所選擇的載體經限制酶切割處理後,再與需接黏的DNA 片段 以適當比例混合均勻,並加入廠商建議的 ligation buffer 和 1 μl T4 ligase,再補上無菌去離子水使最後總體積為 20 μl。混合均勻後,置 於16℃下作用 12~16 小時,即可完成黏合作用。
四. 聚合酶連鎖反應(Polymerase chain reaction, PCR)
取1/10 X 體積的 DNA 當做模板,加入 1/10 X 體積的 dNTP mixture
(dATP, dGTP, dCTP, dTTP)、1/10 X 體積的 10 X Taq buffer 以及分 別為1/10 X 體積最終濃度 1 μmole 的正、反向引子,和 1/10 X 體積 的DMSO 與 1 μl 的 Taq DNA polymerase 2.5 U/ml,最後加無菌去離 子水到總體積為20 μl。利用 GeneAmp PCR system 9600 進行聚合酶 連鎖反應。設定條件為第一階段 94℃,10 分鐘,使模板 DNA 產生 變性(denaturation);再以第二階段 94℃,1 分鐘;(50℃~65℃)
適 合 引 子 之 溫 度 ,1 分鐘,使引子與模板 DNA 進行黏合作用
(annealing); 72℃,0.5 分鐘到 5 分鐘(視 PCR amplicon 片段長 度而定)進行 DNA 延長作用(extension),此反應重複 25 到 35 循 環;最後第三階段72℃,10 分鐘使 DNA 充分延長。
五. E. coli 勝任細胞(competent cell)之備製
將E. coli 菌株(DH5α 或 S17-1)接種於 3 ml LB 之培養液中,
置37℃震盪隔夜培養。之後取 500 μl 菌液到 20 ml LB 中,於 37℃震 盪培養。培養至O.D600約為 0.8~0.9 時,將菌液於 8000 rpm 4℃離心 5 分鐘收集菌體。去上清液,加入約 15 ml 之 4℃ 0.1 M CaCl2使菌體 懸浮於其中,靜置冰浴30 分鐘。之後再以 8000 rpm 4℃離心 10 分鐘。
小心倒去上清液,最後加入1 ml 4℃之 0.1 M CaCl2,搖勻後即可冷藏
備用。未使用之勝任細胞可以加入 87% 經滅過菌的甘油(Glycerol)
至最終濃度為15%,輕輕混勻後置入-80℃下保存備用。
六. 勝任細胞之轉形作用(transformation)
取適量之 DNA 或質體加入 100 μl 勝任細胞中,於冰上作用 10 分鐘後,快速放入42℃水浴槽中進行熱休克(heat shock)反應 2 分 鐘,接著再快速移回冰上靜置5 分鐘。然後加入 500 μl LB,於 37℃
震盪培養約 2 小時。吸取適量菌液塗於含合適抗生素的固態培養基 中,置 37℃隔夜培養。如果質體之載體上帶有 lacZ 基因,則視所需 加入50 μl IPTG(20 mg/ml)及 50 μl X-gal(20 mg/ml)均勻塗在含 適 當 抗 生 素 的 培 養 基 中 隔 夜 培 養 , 進 行 藍 白 篩 選 (Blue-white selection)。
七. 接合作用(conjugation)
首先,將 recipient 菌株(S. maltophilia)與 donor 菌株(E. coli S17-1)分別以 3 ml 的 LB broth 於 37℃震盪培養約 16 小時,而後把 recipient 菌液與 donor 菌液以 1.5 ml 分別加入內含 20 ml LB 的三角錐 瓶(flask)中,以 37℃震盪培養至菌液量約為 OD450 0.7~0.8。接著將 兩菌液分別倒入離心管,以5000 rpm 離心 5 分鐘後去除上清液,再 各加入2 ml LB broth 回溶菌體。然後將 recipient:donor 以 4:1 之 比例混合於 1.5 ml 之微量附蓋離心管,並使其混合均勻。再以 5000
rpm 離心 5 分鐘,去除大部分上清液後,預留少量的 LB broth 與菌體 充分混合。最後以微量吸管將混合之菌液置於 LA 瓊脂培養基內之無 菌的nitrocellulose membrane 上,且以 37℃隔夜培養。經接合作用後 之菌株以含有tetracycline(40 μg/ml)和 norfloxacin(2.5 μg/ml)抗 生素的瓊脂培養基(LB agar plate)進行篩選。所得的轉殖菌株
(transconjugant)為含有外來質體之 S. maltophilia 菌株。
而若為得特定的 isogenic 突變株,需將經接合作用所得之轉殖 菌株(transconjugant)再進一步以含有 10% sucrose 的 LB 瓊脂培養 基(LB agar)進行篩選,之後利用 PCR 和 DNA 定序來確定其正確 性。
八. 質體快速篩檢法(Rapid screen method)
使用已滅菌的牙籤挑選單一菌落的轉型菌株,接種於含有合適抗 生素1 ml 的液態培養基中,37℃震盪培養 6-8 小時。以 12,000 rpm 離 心 5 分鐘,去除上清液。加入含有 1% SDS 的 loading dye,並劇烈 震盪使沉澱物完全打散,接著加入等體積比例的 phenol/choroform,
混合均勻後,以12,000 rpm 離心 10 分鐘。吸取上層溶液 10 μl,利用 agarose gel 電泳分析,也可將僅含有載體的轉型菌株依上述同樣方法 製備後,注入同一agarose gel 中,做為對照組。
九. 抗生素感受性試驗
本論文中之抗藥性試驗主要使用Mueller-Hinton II 培養基,以瓊 脂稀釋法(agar dilution method)進行。作法是將新鮮且隔夜培養之 菌液,以濁度為 McFarland 0.5 做 3 倍稀釋,利用多點接種器
(multipoint inoculator)將其接種至含不同梯度濃度抗生素的培養基 上。抗生素分別是cefoxitin 和 aztreonam,濃度範圍為 8~2048 μg/ml。
而每一接種點上有1.0 x 104 的菌落數(104/spot),接著於 37℃培養 24 小時後觀察並記錄其生長情形,藉此判讀該菌株對 cefoxitin 和 aztreonam 抗 生 素 的 最 低 抑 制 濃 度 ( minimum inhibitory concentration)。
十. 菌落快速檢驗聚合酶連鎖反應(Colony PCR)
1. 檢驗大腸桿菌(E. coli)
每管PCR tube 中加入 dNTP 2 μl,引子(primer-F, primer-R)各 2 μl,DMSO 2 μl,10X Taq buffer 2 μl,Taq polymerase 1 μl,最後補 上無菌去離子水使總體積為 20 μl。之後,利用無菌牙籤沾取菌落在 PCR tube 內稍微攪動,然後進行 PCR 反應。反應完後取 PCR 產物進 行agarose 電泳和 EtBr 染色後,將膠片置於紫外燈箱觀察 DNA 片段 大小。
2. 檢驗嗜麥芽醣黃單胞菌(S. maltophilia)
每管PCR tube 中加入 dNTP 4 μl,引子(primer-F, primer-R)各