國立臺灣大學生命科學院動物學研究所 碩士論文
Institute of Zoology College of Life Science
National Taiwan University Master Thesis
果蠅脂肪酸去飽和酶 2 的分子演化
Molecular Evolution of Fatty acid desaturase 2 in Drosophila
朱光曦
Kuang-Hsi Chu
丁照棣 方淑
Chau-Ti Ting Shu Fang 中華民國 97 年 7 月
July, 2008 指導教授:
Advisors:
iii
Acknowledgements
我個人非常喜歡閱讀,生活中其實任何事物都可以以書來做為比擬:今天是 章節多少,劇情如何發展。假如要用書來比喻我的碩士生涯的話,我想史蒂芬.金 的黑塔系列 The Dark Tower (serious) (皇冠出版)應該是再合適不過。書裡描述 故事的主角羅蘭如何在業(Ka)的驅動下展開尋找支撐世界的樑柱-黑塔的旅程。
中文版的第一集在 2007 年 7 月出版,剛好是我碩二要瘋狂的為實驗而貢獻生命的 時候;最後一集則是在 2008 年 6 月發行,而我當時正好準備要口試了。我還記得 當我看到書的結尾,羅蘭在歷經千辛萬苦終於抵達黑塔時,不禁讓我潸然淚下,
因為黑塔與我一直在累積的 Data 實在是太像了(念念看)。
如同許多偉大的旅程,中途一定有許多導師傾囊相助。在這兩年的碩士旅程,
我必須要感謝三位老師,不只是實驗上的指導,還有許多平時生活上的建議:丁照 棣老師,方淑老師以及曹順成老師。丁老師和方老師是我的指導教授,她們從我 初進實驗室便給予莫大的幫助,無論是實驗上還是生活上。討論實驗的時候也會 和我進行充分的腦力激盪,讓我能夠擁有科學的思維與態度(不過我還需繼續努 力)。而我最感謝她們也是感到最不好意思的,就是關於英文寫作,每次都要花很 多時間改我的摘要和壁報,當然這本論文就更不用說了,方老師遠在美國都還特 地用數位相機把她對論文各章節的意見拍下來寄給我。而曹老師在我於中研院做 實驗的過程裡給予非常多的幫助,曹老師會用相當淺顯易懂的方式很幽默的表達 他的意見,請教他任何問題都可以得到滿意的答覆,還有就是關於 PCR 實驗的問
iv
題,曹老師在這方面給予我相當大的幫助,這本論文幾乎就是用 PCR 堆起來的,
所以假如沒有老師的鼎力相助,我恐怕會被這些問題搞得焦頭爛額。而與這三位 老師同時進行討論更是有如醍醐灌頂,每每都感覺自己是如此的渺小,但是又何 其有幸能夠站在這三位巨人的肩上。還有要感謝昆蟲系的張慧羽老師,每次在 meeting 的時候都給我很多很棒的意見。尤其是在我口試的時候,讓我能對我的研 究有新的思考方向並且糾正以往偏頗的立場。還有中研院的博士後研究員:來自印 度的 Paras,法國的 Stephane,日本的三代隆洋,還有王琄嬋博士。他們對我的 實驗都給予許多的意見與幫助並且毫不吝嗇與我分享他們的經驗。還有我的論文 口試委員:王弘毅老師,張慧羽老師,胡哲明老師和蔡玉真老師,非常感謝你們 在口試時給我的建議。
在書中羅蘭擁有數位共業的夥伴(Ka-tet),而我也很榮幸擁有好幾位實驗室 的夥伴在這兩年給予我莫大的幫助。首先是小春,她扮演的是亦師亦友的角色。
內建 Google 搜尋引擎與台灣美食圖鑑,我想不到有什麼事情和好吃的餐廳是她所 不知道的。除了常常跟我討論實驗外,也常依據豐富的人生經驗給予我日常生活 上的指引,還有要特別感謝她的是,對於我的摘要與壁報的幫忙,是的,她也是 我恐怖的摘要和壁報的長期受害者之一。再來是雅仁,我跟她相處的時間並不多,
所以關係並沒有很熟絡,但是她仍然很熱心的幫我看論文,並給予許多改進的建 議,沒能跟她相處得更久一點,算是我碩士這兩年的最大的遺憾之一。小菊算是 搞笑派的角色,他在我初進實驗室和即將要離開實驗室的兩大關鍵期扮演了很重
v
要的地位,他在我碩一的時候曾經把我的筆記本撕掉、果蠅 stocks 拿去急凍、弄 壞我的 kit…無所不用其極打壓新人(他是無心的,我一直都這麼告訴自己),但是 他卻在忙著準備 GRE 考試的期間還抽空幫我檢查論文。還有承叡,他非常優秀,
他所進行的實驗和我的很有關連,每次他的結果都讓我有新的研究方向可以思 考,他即將到美國念書,希望他能順利的發展。還有跟我一樣要畢業的筱涵,非 常單純善良,每次跟她聊天都會讓人非常輕鬆愉快。我的學妹莞儒大小姐和姵君 以及台大的助理文喬,大馬,麒鳴,士又和其駿,他們所有人在有意無意間都幫 了我不少忙,還有已經離職的小笨和佳瑋,她們常以過來人的身分教給我許多碩 士生存法則。中研院果蠅研究室的各位助理也對我助益良多,尤其是祺筠,他幫 我檢查整本論文,還有珈蓉,民濤,圓通,婉婷和雲露。還有已經離開中研院的 家宣,嘉琳和立婷,他們教導了我許多果蠅實驗技巧。還要感謝巧雯,常常在實 驗室傻呼呼的跑來跑去,逗大家開心。以及來中研院果蠅研究室當過實習生的大 同大學的數位同學。張慧羽老師實驗室的小斌,嵩翰與岱樺也常常和我互相交換 心得與討論各自的研究進度。他們所有人豐富了我的研究生涯,在這段漫長的時 光中伴我一起成長與茁壯。
特別感謝輔仁大學生命科學系九十五年畢的某團體,其成員不時的在 MSN 上 給予我鼓勵並且和我嘴砲,讓我能隨時保持向前衝的動力。
最後要感謝我的家人,他們雖然不清楚我每天在幹嘛,但是關心之情往往溢 於言表;還有我的母親,她在我兩年前甫上研究所之際離我而去,但是我卻感覺
vi
她在這段日子裡從未離開過,一直在我身邊給予支持與祝福。
在研究所的日子裡,我最大的收穫不只是完成了一項研究,還有在尋求真理 的過程中所必須具備的邏輯的態度以及思維,這不光只是對於碩士有所幫助,而 是對我的未來同樣有無限的影響。如同在黑塔一書的最後:羅蘭發現自己不但找 到了旅行的終點,同時也找到了起點。人生充滿了一座座的黑塔,我們窮其一生 不斷的重覆追尋它,但是每次我們都帶著更多的信心與勇氣踏出前進的步伐。
“I believe that when we leave a place…
…part of it goes with us, and part of it remains.
Go anywhere in the station when it is quiet and just listen.
After a while you will hear the echoes of all our conversations…
…every thought and word we’ve exchanged.
Long after we have gone, our voices will linger in these walls.”
By G’Kar, Babylon 5: The Lost Tales.
vii
摘要
去飽和酶 1,去飽和酶 2 和脂肪酸去飽和酶 2 在具性費洛蒙功用的果蠅表皮碳 氫化合物的合成過程中扮演重要的角色。而脂肪酸去飽和酶 2 在黃果蠅和賽昔爾 果蠅中參與合成雌性所特有的費洛蒙中的二烯化合物。但是我們仍然不了解它在 其他種果蠅中所扮演的角色。我在十二種果蠅的基因體中找出脂肪酸去飽和酶 2 可能的同源基因並加以分析。而結果顯示脂肪酸去飽和酶 2 可能是經由反轉錄移 位所產生的基因,產生的時間點可能在水果果蠅和果蠅亞屬分化之前,但在後者 中失去。我也發現脂肪酸去飽和酶 2 在黃果蠅種亞群中和雌雄二型性表皮碳氫化 合物有直接關連,此基因只會表現在具有二型性表皮碳氫化合物的果蠅;在單型 性果蠅則沒有。為了要研究脂肪酸去飽和酶 2 在雌雄單型性表皮碳氫化合物的果 蠅中是否有發生退化現象,我分析了擬黃果蠅,亞庫巴果蠅和德氏果蠅的族群,
結果在二十個擬黃果蠅的品系中找到一個品系有二十五個鹼基對的缺失,在十六 個德氏果蠅的品系內發現其中四個品系帶有無義突變,而在二十個亞庫巴果蠅品 系中,有十四種插入和刪除的序列缺失,導致只有三個品系具有完整的編碼序列。
脂肪酸去飽和酶 2 在三種果蠅中有不同程度的突變累積,代表它在各種果蠅中的 退化可能是獨立發生的。脂肪酸去飽和酶 2 提供了一個研究基因重複的分子演化 的模式。
關鍵字: 表皮碳氫化合物,脂肪酸去飽和酶 2,反轉錄移位,性雙型
viii
ix
Abstract
Three fatty acid desaturase genes, namely desat1, desat2 and Fad2, in Drosophila involve in the production of cuticular hydrocarbons (CHs) that serve as sex pheromones.
Among them, Fad2 is responsible for female-specific diene pheromone biosynthesis in D. melanogaster and D. sechellia. However, the role of Fad2 in other Drosophila species is still unknown. To address this question, the Fad2 homologs in 12 Drosophila species were analyzed. Based on the sequence information, Fad2 is originated by retrotransposition which took place before the split of Drosophila and Sophophora and subsequently lost in the Drosophila lineage. Furthermore, in D. melanogaster species subgroup, Fad2 is expressed only in the species with dimorphic CHs. These results suggest that Fad2 is responsible for sexually dimorphic CHs in the D. melanogaster species subgroup. To understand whether sequence degeneration also occurred in the non-expressed Fad2, populations of three CH monomorphic species, i.e., D. simulans, D. teissieri, and D. yakuba, were surveyed. A 25-bp deletion was observed in one of 20 D. simulans alleles. One non-sense mutation occurred in 4 of 16 D. teissieri sequences.
In D. yakuba, 14 deletions and non-sense mutation resulted in that only 3 out of 20 samples keep full-length Fad2. The different levels of mutation accumulation indicate that the degeneration of Fad2 happened independently in each species. In summary, Fad2 provides a good model to study the molecular evolution after gene duplication.
Key words: cuticular hydrocarbons, Fad2, retrotransposition, sexually dimorphism
x
Contents
口試委員會審定書………....i
Acknowledgements………..iii
Abstracts Chinese………...vii
English……….…...ix
Chapter 1 General Introduction………...1
Chapter 2 The evolution of Fad2 in Drosophila………... 2.1 Introduction………...9
2.2 Materials and Methods………14
2.3 Results………... 2.4 Discussion……….……... Chapter 3 Pseudogenization of Fad2 in Drosophila melanogaster subgroup………...45 3.1 Introduction………..…...
3.2 Materials and Methods………....
3.3 Results………...
3.4 Discussion………....
Conclusion………...
References………...
Appendix………...
9
52 18 34
45 49
62 69 71 81
xii
xiii
List of Figures
Fig 1.1. The CHs biosynthesis pathway in D. melanogaster………..…………...8
Fig 2.1. Comparison of the main CHs in Drosophilidae………...………..13
Fig 2.2. (A)The phylogenic tree of seven desaturase genes from 12 Drosophila species
(B) The Bayesian tree with GTR model………21 Fig 2.3. The Dotplot of desaturase genes...………23
Fig 2.4. Mapping of Fad2 homologs in Drosophila...………..………..26
Fig 2.5. The alignment of Fad2 homolog amino acid sequences in Drosophila...…….
Fig 2.6. The pairwised dN/dS of DNA sequences among 16 Fad2 homologs………….
Fig 2.7. The phylogenetic relationships of Fad2 homologs expression patterns and
sexually dimorphic CHs in Drosophila species……..………...
Fig 2.8. The hypothesized origin of three desaturase genes in Drosophila…………....
Fig 3.1. The nucleotide diversity with sliding windowsize of 2690 bp of Fad2 in D.
simulans...………..
Fig 3.2. Frequency spectrum of Fad2 alleles in D. simulans strains...……….…..
Fig 3.3. The pseudogenization of Fad2 in (A) D. simulans, (B) D. yakuba and
(C) D. teissieri..……….
Fig S1. The annotated transcription binding sites in the 5’ flanking regions of Fad2
homologs………..……….
61 28 30
33 36
54 56
88
xiv
xv
List of Tables
Table 2.1 The list of flies used for expression analysis………...16
Table 3.1 The Tajima’s D, Fu and Li’s and Fay and Wu’s tests results (A) all strains (B)
Africa strains (C) non-Africa strains…...……….
Table 3.2 The McDonald and Kreitman test result from Fad2 in D. simulans………….57
Table 3.3 The McDonald and Kreitman test result (A) and the Tajima’s D, Fu and
Li’s and Fay and Wu’s tests results (B) for Fad2 in D. teissieri...
Table S1 The primer list………...81
Table S2 Desaturase gene homologs in 12 Drosophila species………..83
Table S3 The introduction of transcription factors in the 5’ flanking regions of Fad2
homologs……….89 Table S4 Species and strains used in this study………..91 60 54
xvi
1
Chapter 1
General Introduction
Specific mate recognition system (SMRS) consists of a set of signals and responses between potential mates. Every species has its unique SMRS that controls the
information exchanged between both sexes during interplay (Patterson 1985). SMRS plays a very important role in animal courtship and mating behavior, because
individuals do not respond to inappropriate signals (Greenspan and Ferveur 2000).
In D. melanogaster, the cuticular hydrocarbons serve as contact pheromones, female-specific 7,11-heptacosadiene can stimulate wing vibrations in males (Antony and Jallon 1982; Antony et al. 1985; Ferveur and Sureau 1996). The 7,11-dienes (fatty acid with two double bonds) are sex pheromone in D. melanogaster, though immature males with little or no 7,11-diens can also induce courtship behavior of mature males (Jallon and Hotta 1979; Pechine, Antony, and Jallon 1988). The study of moth
pheromone showed that there were three key characteristics in the biosynthesis pathway:
desaturation, chain-shortening and formation of functional group. Sex-specific pheromone was produced through these steps (Jurenka 2003). The production of sex-specific CHs pheromone in Drosophila might through similar steps, except it uses chain-elongation instead of chain-shortening. The study of the cuticular hydrocarbons biosynthesis pathway in D. melanogaster (Fig.1.1) suggested that males and females
2
share the same elongation-decarboxylation mechanism (Bjostad, Wolf, and Roelofs 1987; Pennanec'h et al. 1991; Reed et al. 1994; Reed et al. 1995), and several ∆9 desaturases are involved in the pathway (Pennanec’h et al. 1997).
Fatty acid desaturases are nonheme iron-containing enzymes that introduce a double bond to specific positions of fatty-acyl carbon chains. Delta-x (∆x) means the double bond is located on the xth carbon–carbon bond, counting from the carboxylic acid end and omega-x (ωx) means the double bond is located on the xth carbon–carbon bond, counting from the terminal methyl carbon. Some Mammals use ∆5 and ∆6 desaturases to synthesize highly unsaturated fatty acids (Sprecher 2000), and both desaturases were widely expressed in human tissues (Cho, Nakamura, and Clarke 1999a; 1999b). Some plants, the synthesis of linoleic acid and α-linolenic acid from stearic acid are catalyzed by acyl-ACP ∆9, ∆12 and ω3 desaturases (Higashi and Murata 1993; Tocher, Leaver, and Hodgson 1998). In insects, the study on Trichoplusia ni and S. littoralis showed that their ∆11 desaturases presented in pheromone glands had conserved biochemical properties and structures with ∆9 desaturases in animals (Wolf and Roelofs 1986; Rodriguez et al. 1992). There are many active desaturases in pheromone glands, which have specific functions in pheromone production. The desaturases in moth’s sex pheromone biosynthesis pathway have different positional and stereochemical specificities, producing fatty acids with Z9, E9, Z10, Z11, E11, Z12,
3
E12, Z14 and E14 double bonds (Jallon and Wicker-Thomas 2003). Some desaturases have been proved to possess multiple functions. For example, Bombyx mori and
Spodoptera littoralis had ∆11 desaturase with ∆10, 12 utilities (Moto et al. 2004; Serra et al. 2006); Thaumetopoea pityocampa contained bifunctional ∆11/∆13 desaturase with acetylenase activity (Serra et al. 2007).
Eight desaturase genes, desaturase 1 (desat1), desaturase 2 (desat2), fatty acid desaturase 2 (Fad2), CG8630, CG9743, CG9747,CG15531 and infertile crescent have been identified by comparing the amino acid sequences similarity with desaturases in rat and yeast, and they are all on the third chromosome of D. melanogaster except infertile crescent, which locates on the second chromosome. desat1 and desat2 use saturated fatty acids as substrates; Fad2, CG8630, CG9743 and CG9747 use unsaturated ones and they all are expressed in adult flies. The function of CG15531 is still unknown. Fad2 is involved in the female-specific CHs biosynthesis pathway in D. melanogaster and D.
sechellia (Chertemps et al. 2006). The gene infertile crescent, located on the 2nd chromosome were found to be expressed mainly in males (reviewed in Jallon and Wicker-Thomas 2003).
desat1 has been identified based on its highly conserved protein products in carps, rats, ticks, and yeasts. Its peptide sequence was 29% and 43% identical with desaturase gene products in rat and yeast, respectively (Wicker-Thomas, Henriet, and Dallerac
4
1997). It can be expressed in both sexes with a higher expression level in females (Lee, Pariza, and Ntambi 1996). It encodes a ∆9 desaturase and prefers to use palmitic acid (chain length of 16 carbons) as substrate to produce ω7 unsaturated fatty acids (Dallerac et al. 2000). desat1 contains five exons with alternative transcription start sites which produce five different transcripts (Jallon and Wicker-Thomas 2003). The interruption of desat1 transcription by insertion of P-element would reduce the overall production of unsaturated cuticular hydrocarbons in both sexes, with the effect was more pronounced in males. The decrease of unsaturated hydrocarbons is accompanied by an increase of saturated ones, suggesting that desat1 is involved in the first desaturation step (Labeur, Dallerac, and Wicker-Thomas 2002).
Another ∆9 desaturase in D. melanogaster is desat2. The chromosome location of desat2 is close to desat1 and shows 65% sequence identity between them. desat2 also encodes a ∆9 desaturase and prefers to use myristic acid (chain length of 14 carbons)to produce ω5 unsaturated fatty acids (Dallerac et al. 2000). The analysis on promoter region of desat2 in different D. melanogaster strains, including African and
cosmopolitan strains, showed that a 16 bp deletion near the putative transcription start site resulted in the loss of expression in cosmopolitan strains (Dallerac et al. 2000;
Takahashi et al. 2001; Jallon and Wicker-Thomas 2003). So desat2 is expressed in both sexes of D. melanogaster African strain and other D. melanogaster subgroup species,
5
but not in D. melanogaster cosmopolitan strains (Greenberg et al. 2006). The main CHs in African and Caribbean population females is 5,9-HD, whereas the main CHs in most D. melanogaster population females is 7,11-HD (Jallon and Pechiné 1989).
There is an asymmetric reproductive isolation between African and cosmopolitan strains in D. melanogaster: The Z type female prefer to mate with the Z type male and has lower or no desire to mate with the cosmopolitan strain male and this sexual
isolation probably is in an incipient speciation (Wu et al. 1995). They also had different copulation latency: The M type female with high amount of 7,11-HD mated more rapidly than the female with 5,9-HD (Ferveur et al. 1996). The functional and
nonfunctional desat2 determined the females’ main CHs, which were thought to be sex pheromone. The analysis of dN/dS ratio (the numbers of synonymous and non-
synonymous substitutions per site) also showed the desat2 had higher evolution rate than desat1. desat2 was therefore suggested to involve in the differentiation of Z and M type in D. melanogaster (Greenberg et al. 2006).
The definition of species in biological species concept is “groups of interbreeding natural populations that are reproductively isolated from other such groups” (Mayr 1963). According to this definition, the barriers of genetic exchange must play an important role in speciation. Reproductive isolation can have diverse mechanisms, including pre- and post-mating isolation that prevents gene flow among populations.
6
The behavioral barrier is the most important barrier among the pre-mating mechanism (Coyne and Orr 2004). Since the CHs in Drosophila serve as sex pheromone and strongly affect courtship behavior, the CHs difference between species is a component of reproductive isolation leading to speciation (Coyne and Charlesworth 1997;
Takahashi et al. 2001).
D. melanogaster shows sexually dimorphic CHs: The male has a large amount of 7-T and female with 7,11-HD. The second double bond in females requires an
additional desaturation step that is not involved in male CHs biosynthesis pathway.
Fad2 had been found to be responsible for diene production in females of D.
melanogaster and D. sechellia. Fad2 was not expressed in both sexes of D. simulans that has monoenes as main CHs (Chertemps et al. 2006). The interspecific difference in major female CHs is mainly controlled by a few genes on the third chromosome (Coyne 1996). A recent study showed that the difference of CHs between females in D.
melanogaster and D. simulans was due to the female-specific expression of Fad2 and elongaseF. The latter was a female-specific elongase in D. melanogaster and D.
sechellia (Chertemps et al. 2006; Chertemps et al. 2007). According to these studies, the evolution of Fad2 might be distinct in different Drosophila species. Fad2 is an
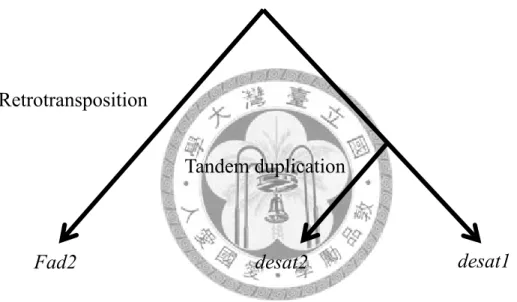
intronless, a characteristic of the product of retrotransporsition (Bai et al. 2007).
However, the role of Fad2 in other Drosophila and the origin of Fad2 are still unknown.
7
When does the retrotransposition happen in the evolution of Drosophila? What are the expression patterns of Fad2 in other Drosophila species? What is evolution of Fad2 in Drosophila? Another important issue is the pseudogenization of non-expressed Fad2 in Drosophila species with sexually monomorphic CHs, such as D. simulans.
In chapter 2, I first searched for the homologs of desaturase genes in Drosophila and tried to reconstruct the phylogeny which can also reveal the origin of Fad2. Then the expression patterns of Fad2 in Drosophila were described. I also discussed their function in CHs synthesis. In chapter 3, I surveyed Fad2 in Drosophila species with non-functional Fad2 to investigate evolutionary patterns during pseudogenization.
These results would tell us the evolutionary fates of duplicated gene in different Drosophila species.
8
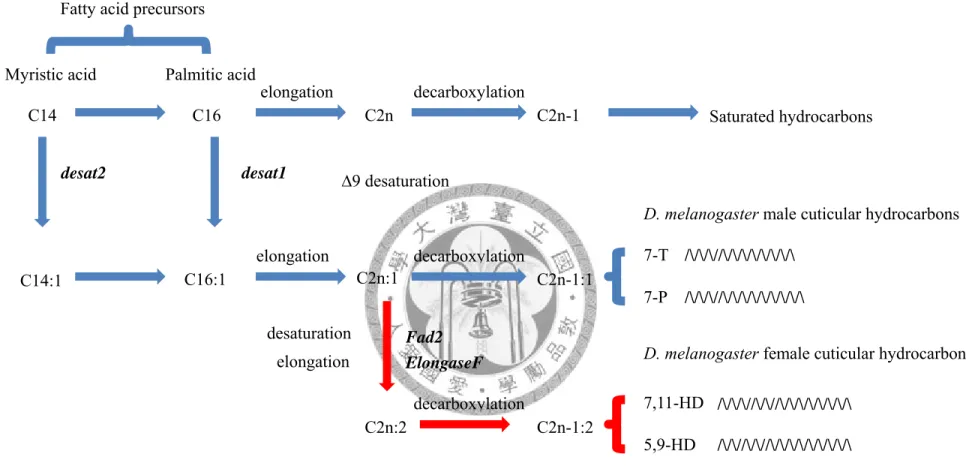
Fig 1.1. The CHs biosynthesis pathway in D. melanogaster (redraw from Legendre et al. 2008). The pathway used in both sexes is shown as blue lines and female–specific pathway is red.
Fatty acid precursors
C14 C16
C14:1 C16:1
C2n
C2n:1
C2n-1
C2n-1:1
C2n:2 C2n-1:2
Saturated hydrocarbons
D. melanogaster male cuticular hydrocarbons
7-T 7-P
D. melanogaster female cuticular hydrocarbons
7,11-HD 5,9-HD elongation
elongation
decarboxylation
decarboxylation
decarboxylation desaturation
∆9 desaturation
desat2 desat1
Fad2
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\/\
/\/\/\//\/\//\/\/\/\/\/\/\/\
/\/\//\/\//\/\/\/\/\/\/\/\/\
elongation ElongaseF Palmitic acid
Myristic acid
9
Chapter 2
The evolution of Fad2 in Drosophila
2.1.
Introduction
The desaturase genes play an important role in the sex pheromone production in insect. Desaturases are particularly significant in the generation of structural diversity of sex pheromone components, since these enzymes have evolved diverse substrate specificities, regiospecificities, and stereospecificities to produce unsaturated fatty acid precursors with a range of chain lengths, variable positions and numbers of double bonds, and both Z and E double-bond geometries (Knipple et al. 2002). As a consequence, studying the evolution of desaturases can help us to understand how signal divergence has occurred in the face of strong selection pressures against small changes in the signal (Roelofs and Rooney 2003). Current knowledge of the evolution of desaturases came from studies on moths.
The desaturase genes in the moth pheromone glands might have evolved from genes involved in normal fatty acid metabolism (Dugdale 1997). The desaturase multigene family is composed of at least four gene clusters, namely ∆9, ∆10, ∆11 and
∆14 desaturase genes, evolved at different rates due to different functional constrain.
The ∆9 (Z9-16:Acid > Z9-18:Acid) group, which contains metabolic desaturases had
10
the slowest evolution rate, was thought to retain the ancestral gene functions. The ∆9 (Z9-16:Acid < Z9-18:Acid) and ∆10,11 groups are composed of desaturase genes that are involved in sex pheromone biosynthesis and evolved at faster rates because of a functional shift from metabolism to reproduction. The ∆14 group, including functional and nonfunctional desaturase genes in sex pheromone production possessed the fastest evolution rate among the four groups. The evolution of desaturase multigene family has been shown to fit the birth-and-death model (Hughes and Nei 1989; Nei and Hughes 1992; Nei, Gu, and Sitnikova 1997; Rooney, Piontkivska, and Nei 2002). Those gene families were created by gene duplication during the evolution processes, in which some genes persisted, and some were deleted or degenerated into pseudogenes. There were three characteristics which would be observed in the phylogeny of these
desaturase gene families: first, sequences were clustered together by genes or
duplication orders not by species; second, the similarity between different genes were low; third, the signatures of gene loss, deletion, and degeneration would be observed (Roelofs and Rooney 2003).
Desaturases has been proved in some Drosophila species to be involved in sex-specific steps in CH biosynthetic pathway (Jallon and Wicker-Thomas 2003;
Chertemps et al. 2006), and played an important role in courtship behavior of D.
melanogaster as its expression is responsible for female-specieifc CHs (Chertemps et al.
11
2006; Legendre et al. 2008). Genetic sequence of a region on the third chromosome, located through genetic studies and screening of deficiencies, was found to be
responsible for female CHs production (Coyne 1996; Wicker-Thomas and Jallon 2000;
Gleason et al. 2005). A putative desaturase gene Fatty acid desaturase 2 (Fad2), whose peptide sequence is 53% and 54% identical with those of desat1 and desat2, was
detected. The putative transmembrane domains and histidine boxes were also found to be highly conserved among the three genes. Using RNA interference to knockdown Fad2 expression in D. melanogaster females significantly increased the monoenes level and decrease the diene production. Those transgenic flies induced less courtship
behavior from normal males. Fad2 was not expressed in males of D. melanogaster and D. sechellia but was involved in the production of 5,9-HD in females of D.
melanogaster African strains. Both sexes in D. simulans did not express Fad2. It played an important role in courtship behavior of D. melanogaster as its expression is
responsible of female-specieifc CHs (Chertemps et al. 2006; Legendre et al. 2008).
Some researchers have proposed that desaturases in D. melanogaster may experience similar evolution progress with moth desaturases (Knipple et al. 2002).
Nevertheless, the evolution of desaturase genes in Drosophila species still remains unclear due to the lack of information of desaturases in most Drosophila species. In this chapter, I aimed to study the evolution of desaturase genes in Drosophila, with special
12
emphasis on Fad2. My first objective is to see if the evolution of desaturases in Drosophila species also follows the birth-and-death model demonstrated in moths. I searched for the desaturase homologs in 12 available Drosophila genomes and reconstructed the phylogeny of desaturase genes.
My second objective is to understand how Fad2 arosed by gene duplication.
Because Fad2 is an intron-less gene, it is most likely the product of retrotransposition (Bai et al. 2007). A gene duplicated by retrotransposition has the following four
characteristics: lost of introns, lost of regulatory regions, a poly A tail and short flanking direct repeats (Rogers 1985). We will examine Fad2 homologs in 12 Drosophila species for those characteristics and infer the origin of Fad2 from our results.
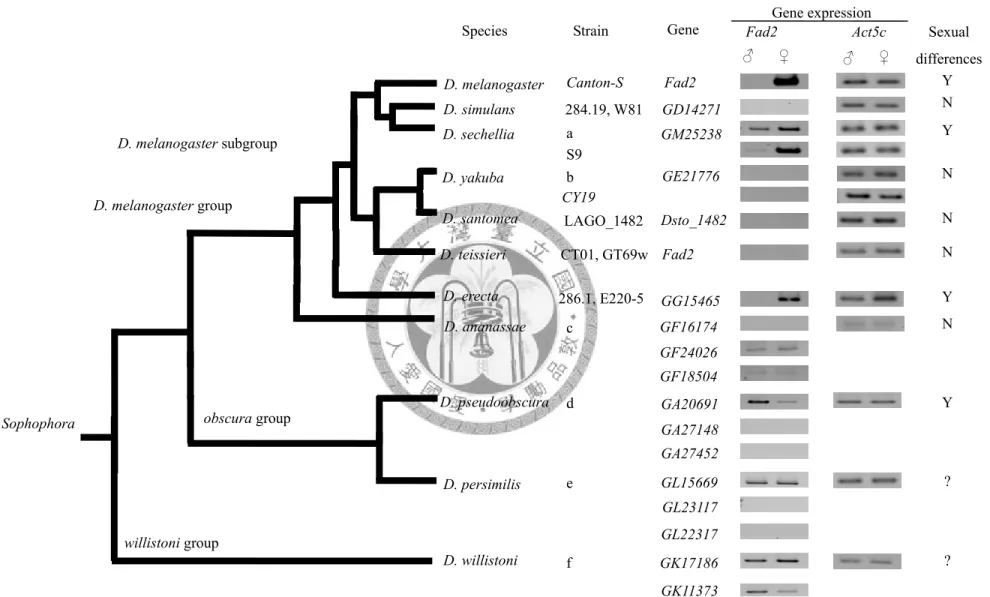
The last objective of this chapter is to study the functional perspective of Fad2 expression in Drosophila species. To do so, the expression patterns of Fad2 homologs were examined and compared with available CHs data (Fig 2.1). Based on the Fad2 function in D. melanogaster, it is responsible for female-specific diene production with unsaturated substrates. If the functions of Fad2 homologs are conserved in other
Drosophila species, I would expect to link the expression of Fad2 to the CH phenotypes that female should possess female-specific diene product.
13
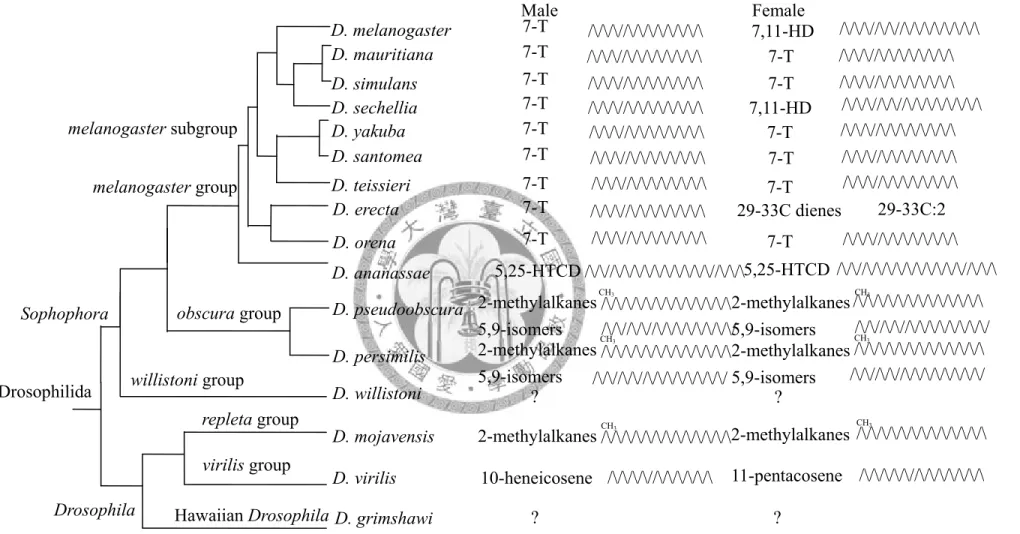
Male Female D. melanogaster
D. mauritiana
D. sechellia D. santomea D. teissieri D. simulans D. yakuba
D. erecta D. orena D. ananassae
7,11-HD
7-T 7-T
7-T 7,11-HD
29-33C dienes 7-T
7-T
7-T 5,25-HTCD 2-methylalkanes 5,9-isomers 2-methylalkanes 5,9-isomers
?
2-methylalkanes 11-pentacosene
?
/\/\/\//\/\/\/\/\/\/\/\ /\/\/\//\/\//\/\/\/\/\/\/\/\
29-33C:2
/\/\//\/\/\/\/\/\/\/\/\/\//\/\/\
/\/\/\/\/\/\/\/\/\/\/\/\/\
/\/\//\/\//\/\/\/\/\/\/\/\/
/\/\/\/\//\/\/\/\/\/\
10-heneicosene /\/\/\/\/\//\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\/\/\/\/\/\/\
/\/\/\//\/\//\/\/\/\/\/\/\/\
CH3 CH3
/\/\/\/\/\/\/\/\/\/\/\/\/\
/\/\/\/\/\/\/\/\/\/\/\/\/\
CH3
CH3
CH3
CH3
/\/\//\/\//\/\/\/\/\/\/\/\/
virilis group
D. virilis
?
Fig 2.1. Comparison of the main CHs in Drosophilidae (Ferveur 2005; Wicker-Thomas 2007). 2-Methylalkanes including methyl branched alkanes, range from 25C-31C, 5,9-Isomers range from 25-29C.
D. pseudoobscura D. persimilis D. willistoni D. mojavensis melanogaster subgroup
melanogaster group
obscura group Sophophora
Drosophilida willistoni group
repleta group
Drosophila
7-T 7-T 7-T 7-T 7-T 7-T 7-T 7-T 7-T 5,25-HTCD
2-methylalkanes 5,9-isomers 2-methylalkanes 5,9-isomers
?
2-methylalkanes
/\/\//\/\/\/\/\/\/\/\/\/\//\/\/\
/\/\/\/\/\/\/\/\/\/\/\/\/\
/\/\/\/\/\/\/\/\/\/\/\/\/\
/\/\/\/\/\/\/\/\/\/\/\/\/\
/\/\//\/\//\/\/\/\/\/\/\/\/
/\/\//\/\//\/\/\/\/\/\/\/\/
Hawaiian Drosophila D. grimshawi
14
2.2.
Materials and Methods
Fad2 homologs in 12 Drosophila genomes
The nucleotide and amino acid sequences of Fad2 and other desaturase genes (desat1, desat2, CG8620, CG9743, CG9747 and CG15531) were obtained from D. melanogaster genome sequences (R5.7) (Kaminker et al. 2002; Misra et al. 2002; Celinker et al. 2002) and performed BLAST in database Genome Assembly (NT), Annotated genes (NT), and
Annotated proteins (AA). The BLAST results of other 11 Drosophila genomes were from D.
pseudoobscura genome (R2.2) (Stark et al. 2007; Richards et al. 2005; Clark et al. 2005) and comparative analyses freeze 1 (CAF1) of D. simulans, D. sechellia, D. yakuba, D. erecta, D.
ananassae, D. persimilis, D. willistoni, D. mojavensis, D. virilis and D. grimshawi genomes (Stark et al. 2007; Richards et al. 2005). I searched 500 bp of Fad2 homologs flanking regions at the syntenic positions for retrotransposition characteristics. I also searched repeat sequences by CENSOR (Kohany et al. 2006) and used Compare and Dotplot program in Genetic
Computer Group package (GCG) to compare those sequence regions to verify if there was Fad2 homologs in the other 11 species. Compare program compares two proteins or nucleic acid sequences and creates a file of the points of similarity between them and the output file will be made a dot-plot by DotPlot (http://www.hku.edu/bruhk/gcgdoc/gcgmanual.html). To
15
identify their chromosome locations surveyed at least five genes in the flanking regions of annotated Fad2 homologs.
Sequence analyses
Sequences downloaded from FlyBase were analyzed by multiple sequences alignment in MegAlign program of DNASTAR (Lasergene software package). The genealogy amino acid sequences of seven desaturase genes were reconstructed by the Molecular Evolutionary Genetic Analysis 4.0 (MEGA 4.0) software using the neighbor-joining (Saitou and Nei 1987) with Poission correction model and maximam parsimony methods (Fitch 1971). Both of them were performed with 1000 bootstrap replications. I used MrBayes (Ronquist and Huelsenbeck 2003) to infer the phylogeny with two variable-rate models, GTR and F80, and running six Markov chains to approximate posterior probabilities with 1500000 generations. The GTR model allows the stationary state frequencies and substitution rates of amino acids to be different and F80 model also allows the stationary state frequencies but assume the same substitution rates. I used infertile crescent (ifc) from D. melanogaster as outgroup. The pairwise dN/dS ratios were estimated by PAML4 (Yang 2007) (runmode= -2, CodonFreq = 2 in codeml.ctl).
Putative transcription factor binding sites in the 5’ flanking regions of Fad2 homologs
16
were analyzed by PATCH program in BIOBASE
(http://www.gene-regulation.com/cgi-bin/pub/programs/patch/bin/patch.cgi?). We only presented the results that were perfectly matched.
Fly stocks
Fourteen strains from 11 Drosophila species were used (Table 2.1). Four to five-day-old virgin flies were used for expression analysis. All flies were kept on cornmeal medium at 22°C with a 12-hr light/dark cycle. I added D. santomea and D. teissieri to RT-PCR study
because they and D. simulans are all belong to D. melanogaster subgroup and share similar CHs phenotype.
Table 2.1. The list of flies used for expression analysis.
Species Strain D. melanogaster Canton-S
D. simulans W81 (284.19)
D. sechellia TSC# 14021-0248.25, S9 D. yakuba TSC# 14021-0261.01, CY19 D. santomea LAGO 1482
D. teissieri CT01, GT69w
D. erecta E220-5 (286.1) D. ananassae TSC# 14024-0371.13 D. pseudoobscura TSC# 14011-0121.94 D. persimilis TSC# 14011-0111.49 D. willistoni TSC# 14030-0811.24
17
RNA preparation and reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA were extracted from five virgin flies of each sex from each strains by using TRIZOL® Reagent (Cal. No. 15596-026, Invitrogen), following the manufacturer’s
instructions. RNA was dissolved in DEPC water and stored at -70°C. The dissolved RNA was treated by DNase I, Deoxyribonuclease I, Amplification Grade (Cal. No. 18068-015,
Invitrogen).
The first-strand cDNA synthesis reactions were performed by SuperscriptTM III First-Strand Synthesis System for RT-PCR (Cal. No. 18080-51, Invitrogen). Gene specific primers (Table S1) were used to amplify target cDNA sequences by PCR. Amplification was performed in a 20 μl reaction volume. Reactions were run with 30 cycles of 30 s at 95°C for
denaturation, 30 s at 55 and 60°C for annealing, 1 min at 72°C for extension, and the final extension at 72°C for 10 min. The PCR amplification products were analyzed by
electrophoresis in 1% agarose gels.
18
2.3.
Results
Desaturase genes in Drosophila
Twelve Drosophila species contained one homolog for each desaturase gene with some exceptions (Table S2): desat2 was absent in D. erecta and multiple Fad2 homologs were found in D. ananassae, D. pseudoobscura, D. persimilis and D. willistoni. The phylogeny of seven desaturase genes in 12 Drosophila species (Fig 2.2A) showed that each desaturase genes formed a monophyletic group. The lineage of desat1 and desat2 were closely-related and Fad2 formed another distinct but closely-related branch. For the other four desaturase genes (CG8630, CG9743, CG9747 and CG15531), CG8630 was the closest one to the lineage of desat1, desat2 and Fad2. CG9743 and CG9747 were closely-related and joined together into an individual branch. CG15531 was the most distinct one from other six desaturase genes. The Bayesian tree with F80 model showed similar results with neighbor-join and maximum parsimony tree (Fig 2.2A). However, the Bayesian tree with GTR model, Fad2 cluster was closely-related to the ourgoup (Fig 2.2B).
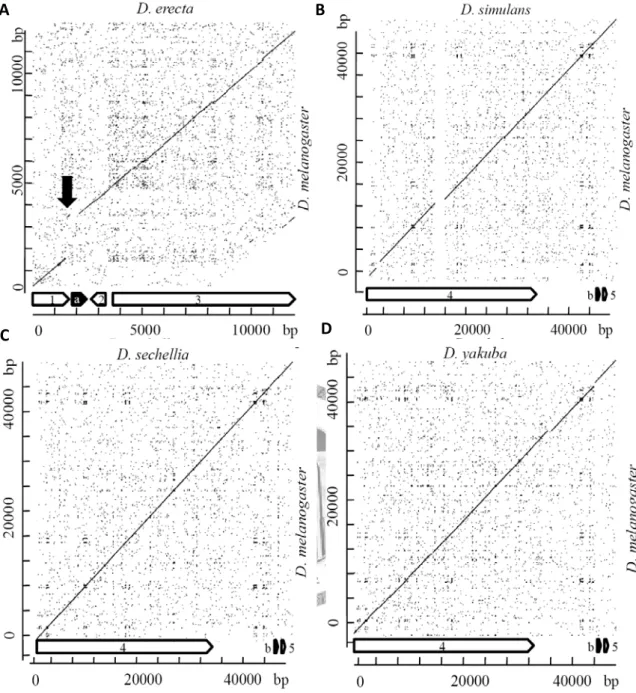
There was no annotated desat2 in D. erecta and the Dotplot result (Fig 2.3) showed that only a small fragment of desat2 sequence was found in the syntenic position. There were three annotated Fad2 homologs in D. ananassae, D. pseudoobscura, D. persimilis and two in
19
D. willistoni. In D. ananassae, all of them (GF16174, GF24026, and GF18504) were not in the syntenic position. GF16174 and GF18504 were in the Muller element B and GF24026 was on the Muller element C, homologous to the right and left arm of the third chromosome in D. melanogaster. In D. pseudoobscura and D. persimilis, there were also three Fad2 homologs. Two Fad2 homologs (GA20691 in D. pseudoobscura and GL15669 in D.
persimilis) were in the syntenic position and the others (GA27148 and GA27452 in D.
pseudoobscura, GL23117 and GL22317 in D. persimilis) were on the Muller element E, homologous to third chromosome right arm in D. melanogaster. The two Fad2 homologs (GK17186 and GK11373) in D. willistoni were both on the Muller element D, homologous to third chromosome left arm in D. melanogaster. However, they were not in the syntenic position. The precise chromosome location of GK11373 was hard to identify due to the variation of flanking genes. The flanking genes of GK11373 were on the same chromosome arm but their sequences were discontinued. The similar pattern is also observed in GF18504 of D. ananassae. There were no annotated desaturase genes in the chromosome locations of those additional Fad2 homologs in other Drosophila species. There was no annotated Fad2 homolog in Drosophila subgenus including D. mojavensis, D. virilis and D. grimshawi. The Dotplot results also revealed that there was no missing annotated Fad2 homolog in the syntenic position of D. mojavensis, D. virilis and D. grimshawi.
20
Retrotranspositions possessed four molecular features: the lost of regulatory regions, no introns, poly-A tail and flanking by short direct repeats. The CENSOR results showed poly-A sites were found out of cases although no repeat sequence within 500 bp flanking regions of the syntenic Fad2 homologs could be identified.
21
Fig 2.2. The phylogenic tree of seven desaturase genes from 12 Drosophila species. (A) The numbers above the nodes are bootstrap values of Neighbor-Joining, Maximum Parsimony method (higher than 50 are shown) and posterior probability of Bayesian method with F81 model (higher than 0.85 are shown). (B) The Bayesian tree with GTR model. The numbers above the nodes are posterior probability (higher than 0.85 are shown). *= 1.00
Dmel CG5887 Dyak GE21698 Dere GG18949 Dana GF17961 Dpse GA19204 Dper GL27316 Dwil GK14407
Dmoj GI10485 Dvir GJ23166 Dgri GH21229
Dwil GK14406 Dsim GD18836 Dsec GM24035 Dmel CG5925 Dyak GE26197
Dpse GA19234 Dper GL27314
Dmoj GI10484
Dwil GK17186 88/99/*
73/79/*
64/76/*
100/75/*
53/61/0.99 66/68/*
100/99/*
100/99/*
98/91/*
78/65/- 99/97/*
57/-/- 99/93/- 67/-/-
100/100/* 100/100/*
100/100/*
85/68/0.99
74/77/- 64/84/0.87
72/-/0.87 71/51/0.90
76/-/0.89 73/61/*
100/99/*
99/81/*
98/89/*
100/100/*
100/100/*
100/100/0.99
100/100
90/76/0.76
89/58/- 98/93/0.94 100/100/*
87/-/- 100/97/*
85/82/0.99 98/66/*
93/84/*
65/77/*
100/100/*
92/96/*
89/69/0.95 100/99/*
90/50/*
81/51/-
100/100/0.95 100/100/*
91/91/*
100/79/0.93 100/90/0.99 91/-/- 100/100/0.89
94/56/- 93/67/0.97
84/99
58/99/*
97/93/*
94/98/*
96/71/*
63/68/*
74/-/0.98 99/97/*
100/99/*
99/93/0.99 -/99/*
-/99/*
-/-/-
desat1 cluster
desat2 cluster
Fad2 cluster
CG8630 cluster
CG9743 cluster
CG9747 cluster
CG15531 cluster
81/-/*
99/70/*
86/85/-
0.2
Dsim GD18837 Dsec GM24036
Dwil GK11373
Dpse GA27148 Dpse GA27452 Dper GL22317 Dana GF18504
Dyak GE21776
Dgri GH18426 Dwil GK13932
Dper GL24461
Dsec GM24094 Dsim GD18893 Dsim GD17217 Dsec GM12168 Dmel CG9743 Dere GG11952 Dyak GE23401 Dana GF16192 Dpse GA22002 Dper GL14059
Dmoj GI24323 Dvir GJ10408
Dvir GJ10410 Dgri GH18424 Dmoj GI24325 Dwil GK13143 Dana GF16194 Dper GL14061 Dpse GA22005 Dyak GE23403 Dere GG11954 Dsec GM12172 Dmel CG9747 Dsim GD17234
Dwil GK13142
Dana GF16193 Dere GG11953 Dyak GE23402 Dmel CG15531 Dsim GD17223 Dsec GM12171
Dmel ifc Dana GF17959
Dpse GA20691 Dper GL15669 Dana GF24026 Dana GF16174
Dere GG15465 Dmel CG7923 Dsim GD14271 Dsec GM25238 Dmoj GI24327 Dvir GJ10413
Dana GF17588 Dpse GA21221 Dere GG19533 Dyak GE26257 Dmel CG8630
Dgri GH18422 Dwil GK13141
Dgri GH18423 Dmoj GI24324 Dpse GA26941 Dper GL14060 100/100/*
100/50/*
Dvir GJ10409 Dper GL23117 Dgri GH21218
Dvir GJ23165
A
-/99/0.86
22
Fig 2.2 (continued)
0 1
Dwil GK17186 Dana GF24026 Dana GF16174
Dana GF18504 Dyak GE21776
Dere GG15465 Dmel CG7923 Dsec GM25238 Dsim GD14271 Dper GL15669 Dpse GA20691
Dper GL23117 Dpse GA27148 Dper GL22317 Dpse GA27452
Dwil GK14407 Dana GF17961
Dsec GM24036 Dsim GD18837 Dmel CG5887
Dyak GE21698 Dere GG18949 Dpse GA19204 Dper GL27316 Dmoj GI10485
Dvir GJ23166 Dgri GH21229
Dwil GK14406 Dyak GE26197
Dmel CG5925 Dsim GD18836 Dsec GM24035 Dana GF17959
Dgri GH21218 Dvir GJ23165 Dmoj GI10484
Dper GL27314 Dpse GA19234
Dana GF16192 Dyak GE23401 Dere GG11952 Dmel CG9743 Dsim GD17217 Dsec GM12168 Dwil GK13141
Dgri GH18422 Dvir GJ10408
Dmoj GI24323
Dper GL14059 Dpse GA22002
Dwil GK13142 Dana GF16193 Dere GG11953 Dyak GE23402 Dmel CG15531 Dsim GD17223 Dsec GM12171 Dpse GA26941 Dper GL14060
Dmoj GI24324 Dvir GJ10409
Dgri GH18423 Dwil GK13143
Dana GF16194 Dere GG11954 Dyak GE23403 Dmel CG9747 Dsim GD17234
Dsec GM12172 Dpse GA22005 Dper GL14061 Dgri GH18424
Dmoj GI24325 Dvir GJ10410 Dwil GK13932
Dvir GJ10413 Dmoj GI24327
Dgri GH18426 Dana GF17588 Dere GG19533
Dpse GA21221 Dper GL24461 Dyak GE26257
Dmel CG8630 Dsec GM24094 Dsim GD18893 0.1
0.93
Fad2 cluster
0.93 0.94 0.95
0.95 0.95 *
**
* *
*
*
0.99
* * 0.99
0.97
*
*
*
*
* 0.99
0.96
*
* *
*
*
desat1 cluster
desat2 cluster
0.94 0.95 0.96
* *
*
* 0.94
*
0.99 0.99
0.98
*
*
*
*
* *
*
0.90
0.92 0.98
* *
*
*
*
0.97
*
*
* *
* *
CG9743 cluster
CG15531 cluster
CG9747 cluster
CG8630 cluster
Dwil GK11373 Dmel ifc
B
23
Fig 2.3. The Dotplot of desaturase genes. A: The comparing of desat2 in D. melanogaster with D. erecta. B-L: The comparing of Fad2 in D.melanogaster with other 11 available Drosophila genome. The comparing included the 5’ and 3’ flanking genes. The black arrow points the residues of desat2 in D. erecta. 1: CG5844, 2: CG17207, 3: desat1, 4: klu, 5:
CG32079, 6: CG6327, a: desat2, b: Fad2.
A B
C D
24
Fig 2.3 (continued)
E F
G H
25
Fig 2.3 (continued)
I J
K L
26
Fig 2.4. Mapping of Fad2 homologs in Drosophila. The homologs genes were searched by FlyBase BLAST tool. Multiple homologs are found in D. ananassae, D. pseudoobscura, D. persimilis and D. willistoni. All symbols are from D. melanogaster homologs.
D. melanogaster D. simulans D. sechellia D. yakuba D. erecta D. ananassae
D. pseudoobscura
D. persimilis
D. willistoni
Fad2 GD14271 GM25238 GE21776 GG15465 GF16174 GF24026 GF18504 GA20691 GA27148 GA27452 GL15669 GL23117 GL22317 GK17186 GK11373
CG31523 CG31522
CG14899 kek4 CG14898
CG13917 CG12004
CG12104 CG33791
CG13672 CG33274 CG7207
CG7213 eIF4E-3
Sodh-2 CG4596 CG14695
CG6608
Ib1 Ibe CG7922 C15
CG14329 s11 InR
CG32152 CG5157 CG16940 pyx
Dhfr
CG5155 CG16838
klu Fad2 CG32079 CG32081 CG6327
27
Fad2 in Drosophila
The analysis of Fad2 homologs showed there were no degeneration occurred except in GE21776 (D. yakuba). There was a 10 bp deletion in the coding sequence and an annotated 32 bp intron that would result in a shorter transcript (1020 bp) in D.
yakuba. The first 300 bp of coding regions were the most divergent in D.
pseudoobscura, D. persimilis, and D. willistoni. The Fad2 homologs in D.
pseudoobscura, D. persimilis, and D. willistoni were longer than other species (1089 bp in GA27148 of D. pseudoobscura and GL23117 of D. persimilis; 1086 bp in GA27452 of D. pseudoobscura and GL22317 of D. persimilis; 1092 bp in GK17186 and GK11373 of D. willistoni) because their 5’ coding regions had additional sequences. The length of other Fad2 in other homologs except GE21776 in D. yakuba was 1065 bp. The
annotated four transmembrane domains and three histidine boxes were highly conserved among sixteen Fad2 homologs (Fig 2.5). The integrity of hisitidine box was very
important because it was essential for desaturase catalytic activity (Knipple et al. 2002).
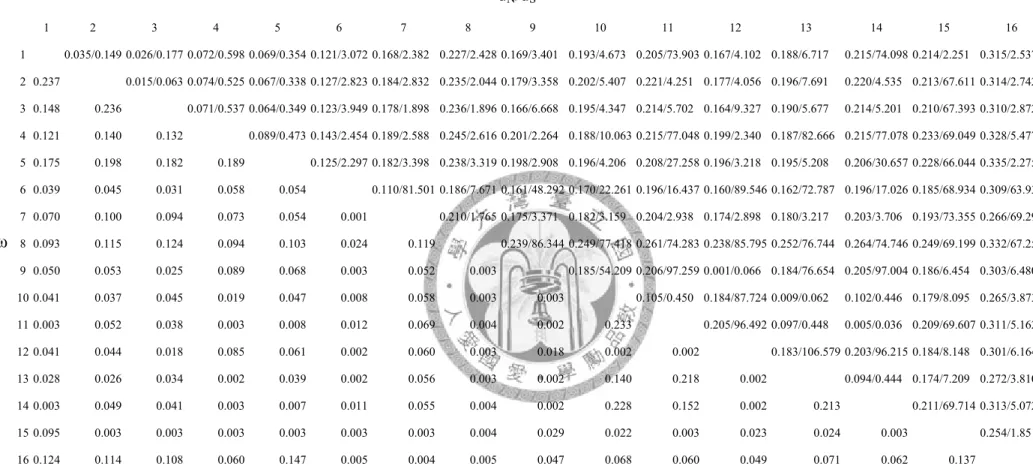
The pairwise dN/dS ratios of all Fad2 homologs were less than one (Fig 2.6).
28
Majority MPPNSE---DTTGVLYESDAETVDG--GLAKDLSTLKTTDGRKLELVWLNIVLFIILHISSLYGVWLLFTAAKWQTFVLFVPAVVVTILGISGG
Dmel_Fad2 ..R.TK---...V...--..STE..NR...S...V...V...S.T.T.V...W.T...V...
Dsim_GD14271 ....TK---.K.V...Q....--..STE..NP...MK...F...L...F..S.T.T..L.LW...V...
Dsec_GM25238 ....TK---...Q-..S--..STE..NT...F...L...S.T.T...S...V...
Dyak_GE21776 ....GK---...V..L..--..ST..GN...F...V...L...T.A.CL....I...
Dere_GG15465 ..S...---.I...T.VQ..G.--..SR..GR..A...T.M...V...L...ST.T..L..I...A..T....A.
Dana_GF16174 ...K.N---.I...L..--...GQ...S..Q...R...V...L...T.T.T..L..PV.L...A...
Dana_GF24026 ...N---E...Q..--..K....Q...S..Q...V...I..E.D.R.TAA.LVVGTISG...
Dana_GF18504 ...S---A...PETKLLEDSPAMD..IGLQN.SKDYE.KW..R..I..AV....AV...F..E.S.R.HLF.LVTQAIG.F...
Dpse_GA20691 ...---DPT...F.G.TA-AA---TVL.KIPDPN----H....L....T..T...V...V...PA.L.L...
Dpse_GA27148 ...VVEFGPKVS...M...-PE---.T...M...YV.TA.V..I...L..I...FS.ILFLFSTV...
Dpse_GA27452 ...DVIELGPSP-...D..C.T..G-PK---.T.S...E...I..I...YV.TA....I..ML.TT....D.FA.ILFSLAS...TA.
Dper_GL15669 ...---YPT...F.G.TA-AA---TVL.KIPDPN----H....L....T..T...V...V...PA.L.L...
Dper_GL23117 ...VVEFGPKVS...M...-PE---.T...M...YV.TA....I...L..I...FS.ILFLFSAV...
Dper_GL22317 ...DVIELGPSP-...D..C.T..G-PK---.T.S...S...I..I...YV.TA....I..ML.TT....D.FA.ILFSLAS...TA.
Dwil_GK17186 ....A.EQKISQPDMC...F.A....I..--...IKH..HAEK..I....F..IA.AY.YVA.T..I...V....S.FTLF.ILFSA...
Dwil_GK11373 ...Y.QTKS--IKKWE...F.K....I..--..N..ITK...AGK..Y.YI....F.LSYIYAL.G..L..F...MIFT.LG.IANYVASLA.
Majority AHRLWAHRTFKANTPLRLIFLFLNTLAFQDAVYYWARDHRVHHKYTETDADPYNSQRGWFFAHIGWLCCKKHPDVVAKGKQIDLSDLEADPLVMFQKKYY
Dmel_Fad2 ...K...R...E..E...I...
Dsim_GD14271 ...K...R...E...I...
Dsec_GM25238 ...K...K...R...E..E...I...
Dyak_GE21776 ...R.K...F...M...R...L...I...
Dere_GG15465 ...K..L...F...N...M...R...A..E...
Dana_GF16174 ...V...I...R...S...R...
Dana_GF24026 ...S...L..K...M....I...I...F...S...E.L...R..V...
Dana_GF18504 ...S..A....L..Q..L..C..F....S..V.V...AK...E...KR...I...H...
Dpse_GA20691 ...M...L....S...TI..RD...RN.K...H.
Dpse_GA27148 ...L...L..F..V...N.S....L....S...R...S.M...N.T....L...RR...H.
Dpse_GA27452 ...Y...VAV...L..F...L...C.D...K...W...T....L...QR...
Dper_GL15669 ...M...L....S...TI..RD...RN.K...H.
Dper_GL23117 ...Y...L...L..F...N.S....L....S...R...S...N.T....L...RR...H.
Dper_GL22317 ...Y...VAV...L..F...L...C.D...K...W...T....L...QR...
Dwil_GK17186 ...T...L..Q..L...M...I...S...S...I....G...RN...I...
Dwil_GK11373 P...YS..S....M..QF.I.L.HST....SAFD...S.V....T....E.L....E..V...TSN..L...HI
Fig 2.5. The alignment of Fad2 homolog amino acid sequences in Drosophila. Three histidine boxes are shown in red and four annotated transmembrane domains are labelled in green.
..R.TK---...V...--..STE..NR...S...V...V...S.T.T.V...W.T...V...
...K...R...E..E...I...
29
Majority LILMPIICFVLPTVVPMYLWGESLNVSWHVMALLRWCLSLNLVWLVNSSAHMHGKRPYDKNISPTDESFLIWLRVGEGYHNYHHVFPWDYKSAELGKYSQ Dmel_Fad2 .L...L...I.T...M...C.V.QG...FF...
Dsim_GD14271 .L...T...I....I.T...M...C.V.Q....FFH...
Dsec_GM25238 .L...T...I....I.T...M...C.V.Q....FFH...
Dyak_GE21776 .L...A.L...CP---H---.RHEAL.---G-.V.Q....FFH...M...
Dere_GG15465 .L...V...F...F...F...I..HF...M...C.VNL....FF.F...
Dana_GF16174 .W...P...L....L...CA...TT...I...F.M...VG..F...P..K Dana_GF24026 ...LL..I....L...KC...IV...VT...I..L...VF...S...S...Y...A..N Dana_GF18504 .Y...S...AI...F...C...SMF....TM.II.M...KF.MK...G..N.NV...M.L...T...AS...P..W Dpse_GA20691 ...L...Y...LC..SCFT...A...I...Y...A....F...R Dpse_GA27148 ...C..L...L..V.F...S.A...P...V..G...L...LY...QS....NQT...M...M.HF..
Dpse_GA27452 ...F..I...L..V.F...S.A...P..T....M..V...L...G...RS....SLV...Y...G..M.R...
Dper_GL15669 ...L...Y...IC..SCFT...A...I...Y...A....F...R Dper_GL23117 ...C..L...L..V.F...S.A...P...V..G...L...LY...QS....NQ...M...M.HF..
Dper_GL22317 ...F..I...L..V.F...S.A...P..T....M..V...L...G...RS....SLV...Y...G..M.R...
Dwil_GK17186 M...C..L.T.AI...C.N..F.NA...PT.F....T...I...Y...T...H..V...F...R Dwil_GK11373 S.I..TFA...LM...F.N.G....F..ATI..LF.E..FTF.I...IY.N...T.N..N.PI....HL...T...N..Q...AY
Majority DVTTKFIDFFARLGWAYDLKSVSLDMVRQRVQRTGDGSHPVWGWGDKDQPQEDIDVTTITHQRKVK---
Dmel_Fad2 ...E.M.Y...L.K...S...LK..VG...NE..
Dsim_GD14271 ...E.M.Y...S.K..A...LK..VG....S...NG..
Dsec_GM25238 ...E.M.Y...S.K..A...LK..VG....S.H.NG..
Dyak_GE21776 ...L...M.Y...L.K..A...RE..V...S....E.
Dere_GG15465 ...E.M.Y...L.K..A...LK..VG...RS...SG.
Dana_GF16174 ....G...I...C.L..K...QR...VG..V...DD.
Dana_GF24026 .L..G..N....I...PEL.QK..M...H.C...RE...K...Q.
Dana_GF18504 ELI.WL..C..K...A.E.LI...L...LM...VG...SN.
Dpse_GA20691 .A..Y...I...TE.L...IK...AD...RL..NAS.
Dpse_GA27148 .L..N..KT...QK..L...T..I...H..Q...S.I...KK.I..
Dpse_GA27452 .LP.IL.R...V...M....K..L...T...EH...S....YKK....
Dper_GL15669 .A..Y...I...TE.L...IK...AD...RL..NAS.
Dper_GL23117 .L..N..KT...QK..L...T..I...H..Q...S...KK.I..
Dper_GL22317 .LP.IL.RI...M....K..L...T...EH...S....YKK....
Dwil_GK17186 .F..Y..Q...SI...T...KR..I...N...AK..MEE...KDE.
Dwil_GK11373 .F..Y..K..SHI...T...T..K....K..K...T..I...TN..RKEAI.IN.
Fig 2.5 (continued)
.L...L...I.T...M...C.V.QG...FF...
...E.M.Y...L.K...S...LK..VG...NE..
30
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1 0.035/0.149 0.026/0.177 0.072/0.598 0.069/0.354 0.121/3.072 0.168/2.382 0.227/2.428 0.169/3.401 0.193/4.673 0.205/73.903 0.167/4.102 0.188/6.717 0.215/74.098 0.214/2.251 0.315/2.537 2 0.237 0.015/0.063 0.074/0.525 0.067/0.338 0.127/2.823 0.184/2.832 0.235/2.044 0.179/3.358 0.202/5.407 0.221/4.251 0.177/4.056 0.196/7.691 0.220/4.535 0.213/67.611 0.314/2.742 3 0.148 0.236 0.071/0.537 0.064/0.349 0.123/3.949 0.178/1.898 0.236/1.896 0.166/6.668 0.195/4.347 0.214/5.702 0.164/9.327 0.190/5.677 0.214/5.201 0.210/67.393 0.310/2.872 4 0.121 0.140 0.132 0.089/0.473 0.143/2.454 0.189/2.588 0.245/2.616 0.201/2.264 0.188/10.063 0.215/77.048 0.199/2.340 0.187/82.666 0.215/77.078 0.233/69.049 0.328/5.477 5 0.175 0.198 0.182 0.189 0.125/2.297 0.182/3.398 0.238/3.319 0.198/2.908 0.196/4.206 0.208/27.258 0.196/3.218 0.195/5.208 0.206/30.657 0.228/66.044 0.335/2.275 6 0.039 0.045 0.031 0.058 0.054 0.110/81.501 0.186/7.671 0.161/48.292 0.170/22.261 0.196/16.437 0.160/89.546 0.162/72.787 0.196/17.026 0.185/68.934 0.309/63.937 7 0.070 0.100 0.094 0.073 0.054 0.001 0.210/1.765 0.175/3.371 0.182/3.159 0.204/2.938 0.174/2.898 0.180/3.217 0.203/3.706 0.193/73.355 0.266/69.291 8 0.093 0.115 0.124 0.094 0.103 0.024 0.119 0.239/86.344 0.249/77.418 0.261/74.283 0.238/85.795 0.252/76.744 0.264/74.746 0.249/69.199 0.332/67.251 9 0.050 0.053 0.025 0.089 0.068 0.003 0.052 0.003 0.185/54.209 0.206/97.259 0.001/0.066 0.184/76.654 0.205/97.004 0.186/6.454 0.303/6.480 10 0.041 0.037 0.045 0.019 0.047 0.008 0.058 0.003 0.003 0.105/0.450 0.184/87.724 0.009/0.062 0.102/0.446 0.179/8.095 0.265/3.873 11 0.003 0.052 0.038 0.003 0.008 0.012 0.069 0.004 0.002 0.233 0.205/96.492 0.097/0.448 0.005/0.036 0.209/69.607 0.311/5.162 12 0.041 0.044 0.018 0.085 0.061 0.002 0.060 0.003 0.018 0.002 0.002 0.183/106.579 0.203/96.215 0.184/8.148 0.301/6.164 13 0.028 0.026 0.034 0.002 0.039 0.002 0.056 0.003 0.002 0.140 0.218 0.002 0.094/0.444 0.174/7.209 0.272/3.810 14 0.003 0.049 0.041 0.003 0.007 0.011 0.055 0.004 0.002 0.228 0.152 0.002 0.213 0.211/69.714 0.313/5.072 15 0.095 0.003 0.003 0.003 0.003 0.003 0.003 0.004 0.029 0.022 0.003 0.023 0.024 0.003 0.254/1.851 16 0.124 0.114 0.108 0.060 0.147 0.005 0.004 0.005 0.047 0.068 0.060 0.049 0.071 0.062 0.137
Fig 2.6. The pairwised dN/dS of DNA sequences among 16 Fad2 homologs. 1: Dmel_Fad2, 2: Dsim_GD14271, 3: Dsec_GM25238, 4:
Dyak_GE21776, 5: Dere_GG15465, 6: Dana_GF16174, 7: Dana_GF24026, 8: Dana_GF18504, 9: Dpse_GA20691, 10: Dpse_GA27148, 11:
Dpse_GA27452, 12: Dper_GL15669, 13: Dper_GL23117, 14: Dper_GL22317, 15: Dwil_GK17186, 16: Dwil_GK11373.
dN/dS
ω