行政院國家科學委員會專題研究計畫 成果報告
以抗體-陽離子性樹枝型高分子載體為目標導向基因傳送之 研究(3/3)
研究成果報告(完整版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2221-E-041-003-
執 行 期 間 : 95 年 08 月 01 日至 96 年 07 月 31 日 執 行 單 位 : 嘉南藥理科技大學生物科技系(所)
計 畫 主 持 人 : 郭榮華
計畫參與人員: 碩士班研究生-兼任助理:林宜陵、曾若雯
報 告 附 件 : 出席國際會議研究心得報告及發表論文
處 理 方 式 : 本計畫可公開查詢
中 華 民 國 96 年 10 月 02 日
Interactions between U-937 human macrophages and poly(propyleneimine) dendrimers
☆Jung-hua Steven Kuo
a,⁎ , Ming-shiou Jan
b, Yi-Lin Lin
aaGraduate Institute of Pharmaceutical Science, Chia Nan University of Pharmacy and Science, 60 Erh-Jen Road, Sec. 1, Jen-Te, Tainan 717, Taiwan
bDepartment of Microbiology and Immunology, Medical College of Chung Shan Medical University, 110 Jianguo Road, Sec. 1, Taichung, Taiwan Received 4 October 2006; accepted 25 March 2007
Available online 1 April 2007
Abstract
Interest in using poly(propyleneimine) (PPI) dendrimers for biomedical applications is increasing. Before using PPI dendrimers in vivo, their interactions with macrophages must be fully understood because they are primarily removed from circulation by the macrophages of the mononuclear phagocyte system. However, few investigators have studied in detail the intracellular responses that cationic dendrimers induce in macrophages. Here we examined the intracellular responses—reactive oxygen species (ROS) content, mitochondria membrane potential, cell size and complexity, and cell cycle profiles—in U-937 human macrophages treated with poly(propyleneimine) dendrimers generation 2 (DAB 2.0) and 3 (DAB 3.0). Our study focused on the concentration ranges within which cell viability was greater than 90% after PPI dendrimers had been incubated for 16 h. For spontaneous ROS generation, DAB 2.0 did not consistently generate hydrogen peroxide production with increasing dosages over the entire culture period while it was capable of generating superoxide content except during the 12 h of incubation. In contrast, DAB 3.0 did not induce any hydrogen peroxide and superoxide production except for an abrupt increase of superoxide content at 60μg/mL after 6 h of incubation. Our results showed that ROS responses in macrophages were strongly influenced by the nature of the dendrimer surface. Except at 3 h, DAB 2.0 increased mitochondrial membrane potential for every dose and culture period. In contrast, DAB 3.0 caused a significant fluctuation in mitochondrial membrane potential only at 6 h, compared with other incubation times. Exposing macrophages to PPI dendrimers caused dramatic and significant changes in macrophage cell size and complexity, and DAB 3.0 caused greater changes than DAB 2.0 did. For incubation times longer than 1 h, propidium iodide staining showed that cells treated with DAB 2.0 and 3.0 had a higher subG1 phase (indicative of apoptosis) than did untreated cells. PPI dendrimers induced different activated patterns in ROS generation and changes of mitochondrial membrane potential than did other carriers such as cationic liposomes and polyalkylcyanoacrylate. The nature of interactions between macrophages and PPI dendrimers is crucial for the design of safer and more effective delivery systems for macrophages. Our findings provide a novel insight into the cytotoxic effects at the molecular level that dendrimers cause in macrophages.
© 2007 Elsevier B.V. All rights reserved.
Keywords: Macrophages; Poly(propyleneimine) dendrimers; Reactive oxygen species; Mitochondrial membrane potential; Cell cycle; Flow cytometry
1. Introduction
Cationic dendrimers such as poly(propyleneimine) (PPI) have attractive nano-characteristics for the delivery of nucleic acid and various biomedical applications[1–3]. Similar to par- ticulate foreign bodies, dendrimers are primarily captured by
the mononuclear phagocyte system (MPS) after an intravenous injection. The interactions between dendrimers and macro- phages are important because macrophages are central in the host defense system and provide opportunities for targeting drug delivery. When peritoneal macrophages uptake nano-de- livery carriers such as polyalkylcyanoacrylate, they might cause an increase in respiratory bursts that release active substances into surrounding environments[4]. Our recent study[5]showed that, because of their high cationic density, cationic dendrimers were cytotoxic to RAW 264.7 macrophages and caused apo- ptosis. Few investigators, however, have studied in detail
Journal of Controlled Release 120 (2007) 51–59
www.elsevier.com/locate/jconrel
☆Grant sponsor: National Science Council, Taiwan.
⁎ Corresponding author. Tel.: +886 6 266 4911x212; fax: +886 6 266 6411.
E-mail address:[email protected](J.S. Kuo).
0168-3659/$ - see front matter © 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.jconrel.2007.03.019
the intracellular responses that cationic dendrimers induce in macrophages.
It has long been recognized that macrophages produce oxygen and nitrogen-reactive metabolites during phagocytosis or when stimulated by a variety of agents[6]. Reactive oxygen species (ROS) such as superoxide anion (O2−) and hydrogen peroxide (H2O2) are widely investigated as signaling mediators of both protection and destruction in macrophages [7]. ROS are important in apoptosis because they modulate multiple signaling pathways and transcriptional activation. ROS also regulate the cellular redox state using antioxidant enzymes[8].
Mitochondria are both the source and target of ROS and also responsible for producing cellular energy. Apoptotic activa- tion of mitochondria leads to a disturbance of mitochondrial membrane potential (ΔΨm), permeability transition, the gen- eration of ROS, and the release of apoptotic proteins[9].
The aim of this study was to evaluate, using flow cytometry, the intracellular events in U-937 human macrophages treated with generation-2 and -3 PPI dendrimers. We investigated the ROS content of O2−and H2O2, and the changes in mitochon- drial membrane potential after exposing macrophages to PPI dendrimers. We also analyzed cell size and the complexity of macrophages with PPI dendrimer treatment using flow cy- tometry. We did a cell cycle analysis to measure cell prolifer- ation and the apoptotic subG1 cell populations in macrophages treated with dendrimers. Our findings provide a novel insight into the cytotoxic effects, at the molecular level, that dendrimers cause in macrophages.
2. Materials and methods 2.1. Materials
Generation-2 and -3 PPI dendrimers (DAB 2.0 and 3.0) were purchased from Sigma-Aldrich Chemicals (St. Louis, MO, USA). DAB 2.0 (DAB-8) and DAB 3.0 (DAB-16) have mo- lecular weights of 773 and 1686 g/mole and 8 and 16 primary surface amine groups, respectively. Propidium iodide (PI) was obtained from Sigma-Aldrich. All other chemicals were pur- chased locally and were of the highest grade available.
2.2. Cell culture and incubation protocol
A human macrophage-like U-937 large-cell lymphoma cell line was maintained in RPMI 1640 medium (Gibco, Grand Island, NY, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco) and 100 U/mL penicillin/
100 μg/mL streptomycin (Sigma) in a humidified atmosphere of 5% CO2 at 37 °C. The U-937 cells were treated with PPI dendrimers for the entire corresponding time period. After in- cubation, PPI dendrimers were removed from the medium and the cells were washed with PBS for further analysis.
2.3. Cytotoxicity assay
The activity of dehydrogenases (an indicator of cell viability) in the cells (1 × 106, treated or untreated with PPI dendrimers)
was simultaneously assessed[10]. Negative control cells con- tained no PPI dendrimers. To measure cell viability, 10μL of a cell-counting kit solution, a tetrazolium salt that produces a highly water-soluble formazan dye upon biochemical reduction in the presence of an electron carrier (1-Methoxy PMS) (Cell Counting Kit-8; Dojindo Laboratories, Tokyo, Japan), was added to a 100μL culture medium and incubated for 1–4 h. The amount of yellow formazan dye generated by dehydrogenases in cells is directly proportional to the number of viable cells in a culture medium. The absorbance at 450 nm was obtained using an ELISA reader with a reference wavelength of 595 nm.
Results are reported as the cell-viability percentage (average OD/average negative-control OD) ± standard deviation (SD).
2.4. Intracellular ROS contents
To detect intracellular H2O2 content, cell suspensions (1 × 106/mL) with or without dendrimer treatment were incubated with 10 μM membrane-permeable probe dichloro- fluorescin diacetate (DCFH-DA) (Molecular Probes, Eugene, OR, USA) for 30 min at 37 °C. Inside cells, acetate moieties of the probe were cleaved and oxidized, primarily by H2O2, to a green fluorescent 2′-7′-dichlorofluorescein (DCF). For intracel- lular O2−content, 10μM of hydroethidine (HE) incubated for 15 min was used. HE is oxidized primarily by O2− and forms ethidium bromide (EB), which emits red fluorescence[11]. Flow cytometry studies were done on a flow cytometer (FACScan;
Becton Dickinson, Mountain View, CA, USA). A 15-mM air- cooled argon-ion laser was used to excite fluorescent DCF at 488 nm, and the emitted fluorescence was measured using a 530/30-nm band-pass optical filter. Ethidium fluorescence was excited at 488 nm and collected using a 585/21-nm band- pass optical filter. Samples were run using 10,000 cells per test sample. Data were analyzed using the CELL QUEST programs.
2.5. Measuring mitochondria membrane potential (ΔΨm) Mitochondrial membrane potential was determined using a fluorescent probe, rhodamine-123[12], a lipophilic cation that accumulates in the mitochondrial matrix in proportion to mito- chondrial membrane potential. Cell suspensions (1 × 106/mL) were incubated with 10μM rhodamine-123 (R-123, Sigma) for 30 min at 37 °C and then thoroughly washed three times with PBS buffer. The fluorescence was excited at 488 nm and analyzed using a 530/30-nm band-pass optical filter.
2.6. Cell size and complexity
Flow cytometry can discriminate a variety of cell parameters, including cell size and complexity[13]. The typical indicators of cell size and complexity using flow cytometry are the light intensities of forward scatter (FSC) and 90 °C side scatter (SSC). The FSC signal is related to the size, volume, and other factors, such as the refractive index of the cell. In addition to size and volume, the SSC signal is related to the surface texture and internal structure of the cell.
52 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
2.7. Cell cycle analysis
Untreated and dendrimer-treated cells (1 × 106) were fixed using a solution containing 70% ethanol and 30% PBS for 12 h at 4 °C. The cells were then centrifuged at 1200 rpm for 10 min to remove the fixation solution. The cell pellets were incubated with DNA staining solution (40 μg/mL propidium iodide and 100 μg/mL RNase A in PBS) for 30 min in the dark. Ten thousand cells per sample were analyzed using flow cytometry.
2.8. Statistical analysis
Data are expressed as means ± standard deviations of three experiments carried out in duplicate. Statistical analyses were performed using a one-way analysis of variance (ANOVA) with a significance level of 0.05. The data with dendrimer-treated cells at different dosages were compared with data from untreated cells at each corresponding incubation time.
3. Results
3.1. Cytotoxicity assay
An ideal carrier for biomedical applications should be non- toxic to cells. DAB 2.0 and 3.0 were chosen in our study because they are less cytotoxic than other cationic dendrimers such as poly(amidoamine) [14]. Also, our study focused on the concentration ranges within which PPI dendrimers did not elicit toxic effects to cells. For the concentration ranges of PPI dendrimers used in the following experiments (DAB 2.0: 40–
300 μg/mL; DAB 3.0: 20–100 μg/mL), cell viability was greater than 90% after 16 h of incubation (confirmed using a trypan blue exclusion assay; data not shown). For higher con- centrations and longer incubation times, cell viability was less than 70% and excluded in the following experiments.
3.2. Intracellular ROS content
Macrophages contribute a substantial portion of ROS pro- duced as part of the host defense function. When an imbalance occurs between ROS production and antioxidant defense, ROS- generated oxidative damage can affect cellular functions through a series of events and construct the molecular level in
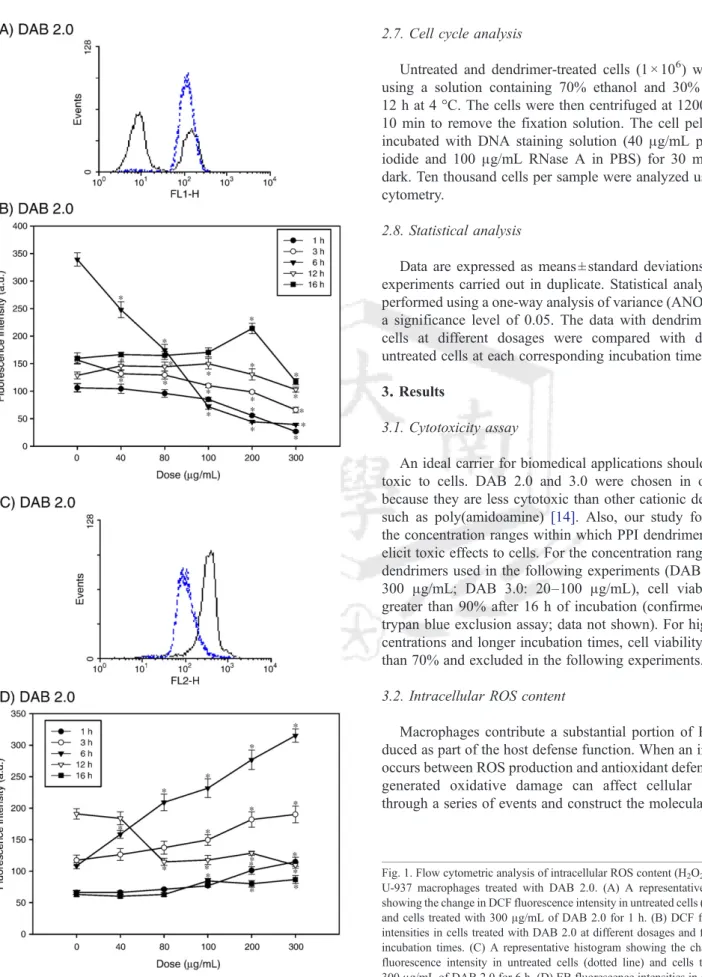
Fig. 1. Flow cytometric analysis of intracellular ROS content (H2O2and O−2) in U-937 macrophages treated with DAB 2.0. (A) A representative histogram showing the change in DCF fluorescence intensity in untreated cells (dotted line) and cells treated with 300μg/mL of DAB 2.0 for 1 h. (B) DCF fluorescence intensities in cells treated with DAB 2.0 at different dosages and for different incubation times. (C) A representative histogram showing the change in EB fluorescence intensity in untreated cells (dotted line) and cells treated with 300μg/mL of DAB 2.0 for 6 h. (D) EB fluorescence intensities in cells treated with DAB 2.0 at different dosages and for different incubation times. Untreated cells were controls for each corresponding incubation period. Data expressed as means ± standard deviations of three experiments carried out in duplicate.
⁎pb0.05, compared to control.
53 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
the development of various diseases [15]. Macrophages can produce ROS such as H2O2and O2−in response to interactions with nanoparticles[4,16,17].
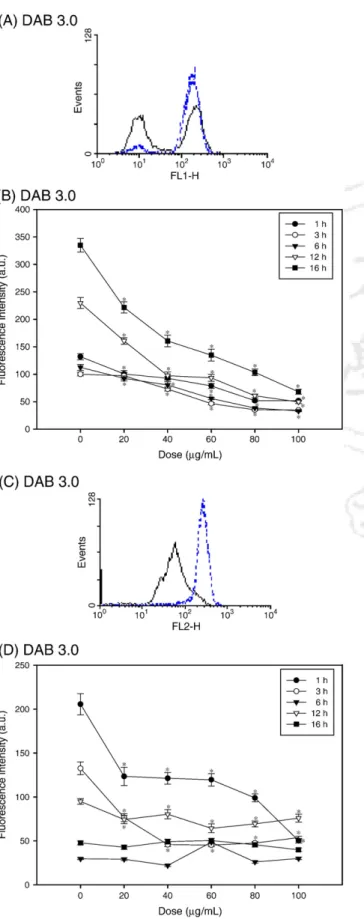
Although many biomedical applications using dendrimers are successful, intracellular mechanisms that stimulate mac- rophages remain an insufficiently studied area of obvious rele- vance to dendrimer-based delivery systems. H2O2production in DAB 2.0-treated macrophages was dose- and time-dependent, as indicated by the change in DCF fluorescence intensity levels (Fig. 1A and B). As DAB 2.0 concentrations increased from 40 to 300μg/mL, intracellular DCF fluorescence intensity decreased relative to fluorescence intensity in untreated con- trol cells over the first 6 h. However, after longer incubation times (12 and 16 h), DCF fluorescence intensity initially slow- ly increased at 12 h and was unchanged at 16 h, peaked at 100μg/mL at 12 h and at 200 μg/mL at 16 h, and decreased at 300μg/mL. In HE-treated cells, there was a significant rise in the mean fluorescence intensity relative to untreated control cells over the first 6 h, and then a drop in O2−production at 12 h, and, finally, a slow rise in cells treated with higher doses (100 to 300 μg/mL) at 16 h (Fig. 1C and D). DAB 3.0-treated macrophages showed dose-dependent decreases in DCF fluorescence intensity relative to untreated cells over the en- tire incubation period (Fig. 2A and B). Also, ethidium fluo- rescence intensity declined compared with untreated control cells after 1, 3, and 12 h (Fig. 2C and D). Except for an abrupt increase at 60μg/mL, ethidium fluorescence intensity maintained the same levels as did untreated control cells for all dosages at 6 h. We found no significant fluctuations in O2− content after 16 h of incubation. These results revealed that the effect of PPI dendrimers on the intracellular ROS content macrophages was dependent on the PPI dendrimer generation. DAB 3.0 has more primary surface amine groups than DAB 2.0 does and is more cytotoxic than DAB 2.0 is. Although cytotoxicity was not significantly different be- tween DAB 2.0 and 3.0, there was a considerable and generation-dependent difference in macrophage intracellular ROS content.
3.3. Responses of mitochondrial membrane potential (ΔΨm) Unstable mitochondrial membrane potential and redox tran- sitions have negatively affected cellular survival via mechan- isms involving ROS-induced ROS release [18]. A fall in mitochondrial membrane potential has usually been seen during apoptosis initiated by mitochondria releasing cytochrome c, whereas some studies have reported the reverse: respiring
Fig. 2. Flow cytometric analysis of intracellular ROS content (H2O2and O2−) in U-937 macrophages treated with DAB 3.0. (A) A representative histogram showing the change in DCF fluorescence intensity in untreated cells (dotted line) and cells treated with 100μg/mL of DAB 3.0 for 1 h. (B) DCF fluorescence intensities in cells treated DAB 3.0 at different dosages and for different incubation times. (C) A representative histogram showing the change in EB fluorescence intensity in untreated cells (dotted line) and cells treated with 100μg/mL of DAB 3.0 for 1 h. (D) EB fluorescence intensities in cells treated with DAB 3.0 at different dosages and for different incubation times. Untreated cells were controls for each corresponding incubation period. Data are expressed as means ± standard deviations of three experiments carried out in duplicate.
⁎pb0.05, compared to control.
54 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
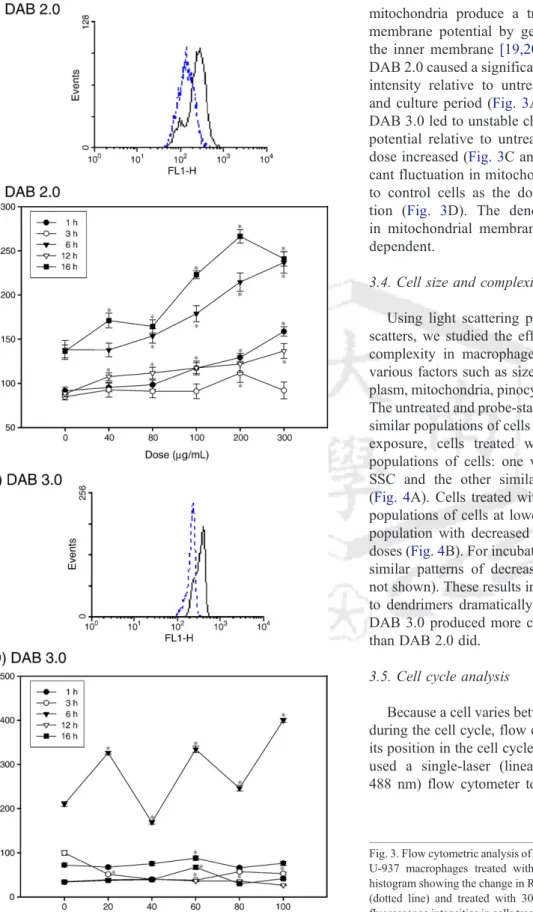
mitochondria produce a transient increase in mitochondrial membrane potential by generating a proton gradient across the inner membrane [19,20]. Except at 3 h incubation time, DAB 2.0 caused a significant rise in the mean red fluorescence intensity relative to untreated control cells for every dose and culture period (Fig. 3A and B). In contrast to DAB 2.0, DAB 3.0 led to unstable changes in mitochondrial membrane potential relative to untreated control cells as the DAB 3.0 dose increased (Fig. 3C and D). Notably, there was a signifi- cant fluctuation in mitochondrial membrane potential relative to control cells as the dose increased after 6 h of incuba- tion (Fig. 3D). The dendrimer-treatment-induced changes in mitochondrial membrane potential were also generation- dependent.
3.4. Cell size and complexity analysis
Using light scattering properties measuring FSC and SSC scatters, we studied the effect of dendrimers on cell size and complexity in macrophages. These properties are related to various factors such as size, volume, plasma membrane, cyto- plasm, mitochondria, pinocytic vesicles, and lysosomes[13,21].
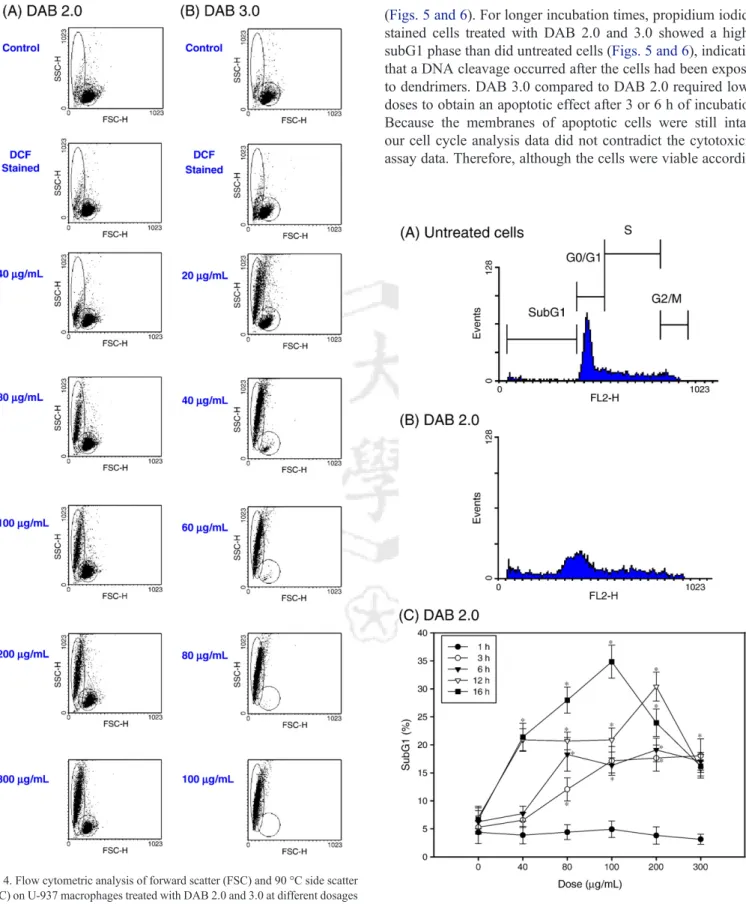
The untreated and probe-stained cells after 1 h of incubation had similar populations of cells (Fig. 4). With increasing dendrimer exposure, cells treated with DAB 2.0 had two different populations of cells: one with decreased FSC and increased SSC and the other similar to untreated or unstained cells (Fig. 4A). Cells treated with DAB 3.0 also had two different populations of cells at lower doses and then one uniform cell population with decreased FSC and increased SSC at higher doses (Fig. 4B). For incubation times longer than 1 h, we found similar patterns of decreased FSC and increased SSC (data not shown). These results indicated that exposing macrophages to dendrimers dramatically affected cell size and complexity.
DAB 3.0 produced more changes in cell size and complexity than DAB 2.0 did.
3.5. Cell cycle analysis
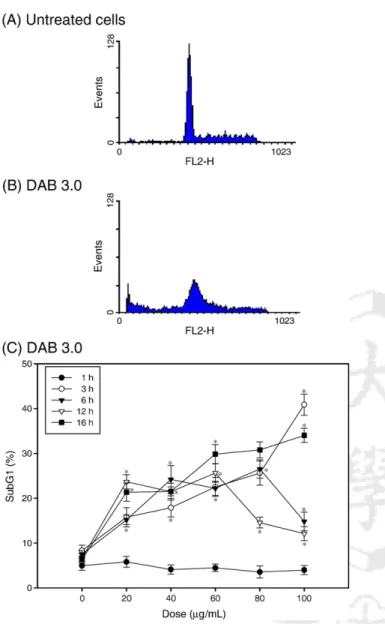
Because a cell varies between hypodiploid and diploid DNA during the cell cycle, flow cytometry can be used to determine its position in the cell cycle based on its DNA content[22].We used a single-laser (linear propidium iodide fluorescence, 488 nm) flow cytometer to determine DNA strand breaks in
Fig. 3. Flow cytometric analysis of mitochondria membrane potential (ΔΨm) on U-937 macrophages treated with DAB 2.0 and 3.0. (A) A representative histogram showing the change in R-123 fluorescence intensity in untreated cells (dotted line) and treated with 300μg/mL of DAB 2.0 for 6 h. (B) R-123 fluorescence intensities in cells treated with DAB 2.0 at different dosages and for different incubation times. (C) A representative histogram showing the change in R-123 fluorescence intensity in untreated cells (dotted line) and cells treated with 60μg/mL of DAB 3.0 for 6 h. (D) R-123 fluorescence intensities in cells treated with DAB 3.0 at different dosages and for different incubation times.
Untreated cells were controls for each corresponding incubation period. Data are expressed as means ± standard deviations of three experiments carried out in duplicate. ⁎pb0.05, compared to control.
55 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
cells treated with dendrimers. We found no significant difference in the subG1 phase (indicative of apoptotic cells) in cells exposed and not exposed to dendrimers after 1 h of incubation
(Figs. 5 and 6). For longer incubation times, propidium iodide- stained cells treated with DAB 2.0 and 3.0 showed a higher subG1 phase than did untreated cells (Figs. 5 and 6), indicating that a DNA cleavage occurred after the cells had been exposed to dendrimers. DAB 3.0 compared to DAB 2.0 required lower doses to obtain an apoptotic effect after 3 or 6 h of incubation.
Because the membranes of apoptotic cells were still intact, our cell cycle analysis data did not contradict the cytotoxicity assay data. Therefore, although the cells were viable according
Fig. 4. Flow cytometric analysis of forward scatter (FSC) and 90 °C side scatter (SSC) on U-937 macrophages treated with DAB 2.0 and 3.0 at different dosages for 1 h of incubation. The un-modified and modified cell populations are marked
by lower and upper circles, respectively. Fig. 5. Cell cycle analysis of U-937 macrophages treated with DAB 2.0.
Representative histograms showing the DNA content in (A) untreated cells and (B) cells treated with 300μg/mL of DAB 2.0 for 6 h. (C) The percentages of subG1 populations (subG1 %) in cells treated DAB 2.0 at different dosages and for different incubation times. Untreated cells were controls for each corresponding incubation period. Data are expressed as means ± standard deviations of three experiments carried out in duplicate. ⁎pb0.05, compared to control. subG1 % = subG1/M1; M1 = subG1 + G0 / G1 + S + G2 / M.
56 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
to the cytotoxicity assay, apoptotic progress had occurred inside the cells.
4. Discussion
In this study, we looked at the intracellular events in macro- phages that dendrimers will encounter in the body during drug delivery. There is an increasing body of evidence that nanoparticles can lead to considerable cytotoxicity [23]. ROS generation and oxidative stress measurement have been de- veloped to mechanistically evaluate cytotoxic responses to nanoparticles. Although biocompatibility and cytotoxicity of dendrimers have been widely documented, relatively few studies have focused on the field of in vitro ROS generation
and oxidative stress induced by dendrimers[24]. In the present study, the measurement of ROS generation and oxidative stress to study the cytotoxic effects of dendrimers lead to a number of interesting findings.
We showed that PPI dendrimers induce different kinetic responses of the cellular ROS production than other carriers and that they are also dependent on the generation of the dendrimer.
Cationic liposomes and polyalkylcyanoacrylate nano-carriers are reported to induce increased H2O2 production in macro- phages [4,25]. For the lower-generation dendrimer DAB 2.0, kinetic H2O2 production tended to decline with increasing dosages, but O2− generation tended to increase, except for the 12-h incubation (Fig. 1). In contrast, with the higher generation dendrimer DAB 3.0, H2O2content decreased dose-dependently after all incubation periods, as did O2−content, except for the longer culture times of 12 and 16 h (Fig. 2). Our results showed that ROS responses in macrophages were strongly influenced by the nature of the dendrimer surface. These ROS responses may be a direct effect of the different surface-positive amine groups of DAB 2.0 and DAB 3.0. Because DAB 3.0 is more cytotoxic than DAB2.0 is, decreases in H2O2and O2−contents reflect the fact that DAB 3.0 disturbed the delicate balance between detoxification and ROS generation than DAB 2.0 did[7].
We showed that mitochondrial membrane potential in macrophages treated with dendrimers is also dependent on dendrimer generation (Fig. 3). DAB 2.0 mostly increased mitochondrial membrane potential for every dose and culture period, which indicated that mitochondrial swelling, but not rupture, occurred[19]. In contrast, DAB 3.0 caused a significant fluctuation in mitochondrial membrane potential at 6 h com- pared with other incubation times. These changes in mitochon- drial membrane potential, however, did not induce any cytotoxicity in macrophages, because the activity of dehydro- genases, produced primarily by mitochondria, was well main- tained, as evidenced by the cytotoxicity assay.
Typically, superoxide generation was closely related to the alterations in mitochondria before apoptosis[9]. DAB 2.0, but not DAB 3.0, caused superoxide generation and correlated well with the changes in mitochondrial membrane potential. We found no association between H2O2production and changes in mitochondrial membrane potential for either dendrimer.
Decreased mitochondrial membrane potential and increased H2O2production in macrophages have been reported for cat- ionic liposomes [26]. Our results indicated that cationic dendrimers and cationic liposomes had different effects on ROS generation and the changes in mitochondrial membrane potential in macrophages. Although we have no direct evidence that dendrimers interact with the mitochondrial membrane, a previous study [27] revealed that nanospheres with surface cationic amine groups caused mitochondria to disappear from macrophages. Studies on the subcellular trafficking of poly- amidoamine dendrimer also indicated that certain amounts of the dendrimer shuttled between lysosomes and mitochondria at later time points[28]. The buffering capacity of polycations has been reported to induce endosome rupture because of excessive water influx [29]. From there, polycations may interact with
Fig. 6. Cell cycle analysis of U-937 macrophages treated with DAB 3.0.
Representative histograms showing the DNA content in (A) untreated cells and (B) cells treated with 80μg/mL of DAB 3.0 for 3 h. (C) The percentages of subG1 populations (subG1 %) in cells treated with DAB 3.0 at different dosages and for different incubation times. Untreated cells were controls for each corresponding incubation period. Data are expressed as means ± standard deviations of three experiments carried out in duplicate. ⁎pb0.05, compared to control subG1 % = subG1 / M1; M1 = subG1 + G0 / G1 + S + G2 / M.
57 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
other membrane-protected targets such as mitochondria. The above observations imply a reasonable possibility that DAB dendrimers interact with the mitochondrial membrane in macrophages at later incubation times.
One hour after macrophages had been exposed to dendri- mers, their cell size and complexity dramatically changed (Fig. 4). DAB 3.0 produced more changes in cell size and complexity than did DAB 2.0, and these findings were con- sistent with the greater membrane disruption caused by other high-generation cationic dendrimers[30]. Cationic nanospheres formed particle-filled vacuoles after uptake into phagosomes in RAW 264.7 macrophages [27]. This finding may explain the increase of internal complexity (cytoplasmic granularity and cell density) in Fig. 4. Our findings of changed cell size and complexity were also supported by the changes of cell morphology seen using scanning electron microscopy the cells had been exposed to DAB dendrimers for 1 h[31].
Next, we investigated the cell cycle profiles of macrophages treated with dendrimers. The increase observed in the subG1 population of cells treated with dendrimers at very short incubation times (N3 h) indicated that apoptosis rapidly pro- gressed. Although the changes in intracellular ROS generation and mitochondrial membrane potential were different between DAB 2.0 and 3.0, these changes contributed to the increases in the subG1 population of cells treated with dendrimers, and this marked the point of no return for apoptotic cells. After we removed dendrimers from the culture medium and re-cultured the cells for another 24 h, viability significantly decreased in cells incubated with dendrimers for longer than 1 h (data not shown). These results indicated that the changes of ROS generation and mitochondrial membrane potential strongly influenced cell proliferation and caused apoptosis in cells incubated with dendrimers for longer than 1 h. However, viability was well maintained and reached confluence after 3 days in cells incubated with dendrimers for 1 h and 20 min (data not shown). This indicates that minor levels of oxidative stress produced by dendrimers induce adequately protective effects in cells.
Finally, we did not detect any TNF-α production in macrophages treated with dendrimers (unpublished data). We also found this in macrophages activated by cationic chitosan- DNA nanoparticles[32]. Because proinflammatory effects are regulated by the redox-sensitive MAP kinase and NF-κB cascades, our results demonstrated that the regulation of these pathways in dendrimer-activated macrophages did not lead to proinflammatory cytokine production[27].
5. Conclusion
Although the concentrations of PPI dendrimers used in our study were not cytotoxic, various intracellular responses in dendrimer-treated macrophages still occurred time-dependently.
These intracellular responses—ROS content, mitochondria membrane potential, cell size and complexity, and cell cycle profiles—are dependent on the dendrimer generation and stand apart from the other particle types. The nature of interactions between macrophages and PPI dendrimers is of greater
importance for the design of safer and more effective delivery systems for macrophages. Our findings contribute to a better understanding of the molecular action of PPI dendrimers in the cellular system.
Acknowledgment
This work was supported by grant NSC 95-2221-E-041-003 from the National Science Council, Taiwan.
References
[1] C. Dufes, C.I.F. Uchegbu, A.G. Schatzlein, Dendrimers in gene delivery, Adv. Drug Deliv. Rev. 57 (2005) 2177–2202.
[2] U. Boas, P.M. Heegaard, Dendrimers in drug research, Chem. Soc. Rev. 33 (2004) 43–46.
[3] R. Esfand, D.A. Tomalia, Poly(amidoamine) (PAMAM) dendrimers: from biomimicry to drug delivery and biomedical applications, Drug Discov.
Today 6 (2001) 427–436.
[4] T. Cruz, R. Gaspar, A. Donato, C. Lopes, Interaction between poly- alkylcyanoacrylate nanoparticles and peritoneal macrophages: MTT metab- olism NBT reduction and NO production, Pharm. Res. 14 (1997) 73–79.
[5] J.H. Kuo, M.S. Jan, H.W. Chiu, Mechanism of cell death induced by cationic dendrimers in RAW 264.7 murine macrophage-like cells, J. Pharm. Pharmacol. 57 (2005) 489–495.
[6] K.E. Iles, H.J. Forman, Macrophage signaling and respiratory burst, Immunol. Res 26 (2002) 95–105.
[7] M. Nagata, Inflammatory cells and oxygen radicals, Curr. Drug Targets Inflamm. Allergy 4 (2005) 503–504.
[8] C. Napoli, F. de Nigris, W. Palinski, Multiple role of reactive oxygen species in the arterial wall, J. Cell. Biochem. 82 (2001) 674–682.
[9] H.U. Simon, A. Haj-Yehia, F. Levi-Schaffer, Role of reactive oxygen species (ROS) in apoptosis induction, Apoptosis 5 (2000) 415–418.
[10] M. Ishiyama, H. Tominaga, M. Shiga, K. Sasamoto, Y. Ohkura, K. Ueno, A combined assay of cell viability and in vitro cytotoxicity with a highly water-soluble tetrazolium salt neutral red and crystal violet, Biol. Pharm.
Bull. 19 (1996) 1518–1520.
[11] G. Rothe, G. Valet, Flow cytometric analysis of respiratory burst activity in phagocytes with hydroethidine and 2′,7′-dichlorofluorescin, J. Leukoc.
Biol. 47 (1990) 440–448.
[12] R.K. Emaus, R. Grunwald, J.J. Lemasters, Rhodamine 123 as a probe of transmembrane potential in isolated rat-liver mitochondria: spectral and metabolic properties, Biochim. Biophys Acta. 850 (1986) 436–448.
[13] M.C. Serrano, R. Pagani, M. Manzano, J.V. Comas, M.T. Portoles, Mitochondrial membrane potential and reactive oxygen species content of endothelial and smooth muscle cells cultured on poly(epsilon-caprolac- tone) films, Biomaterials 27 (2006) 4706–4714.
[14] B.H. Zinselmeyer, S.P. Mackay, A.G. Schatzlein, I.F. Uchegbu, The lower- generation polypropylenimine dendrimers are effective gene-transfer agents, Pharm. Res. 19 (2002) 960–967.
[15] B.A. Freeman, J.D. Crapo, Biology of disease: free radicals and tissue injury, Lab. Invest. 47 (1982) 412–426.
[16] P.G. Barlow, K.J. Donaldson, MacCallum, A. Clouter, V. Stone, Serum exposed to nanoparticle carbon black displays increased potential to induce macrophage migration, Toxicol. Lett. 155 (2005) 397–401.
[17] R. Fernandez-Urrusuno, E. Fattal, J. Feger, P. Couvreur, P. Therond, Evaluation of hepatic antioxidant systems after intravenous administration of polymeric nanoparticles, Biomaterials 18 (1997) 511–517.
[18] D.B. Zorov, M. Juhaszova, S.J. Sollott, Mitochondrial ROS-induced ROS release: an update and review, Biochim. Biophys. Acta. 1757 (2006) 509–517.
[19] A.R. Khaled, D.A. Reynolds, H.A. Young, C.B. Thompson, K. Muegge, S.K.
Durum, Interleukin-3 withdrawal induces an early increase in mito- chondrial membrane potential unrelated to the Bcl-2 family. Roles of intracellular pH, ADP transport and F(0)F(1)-ATPase, J. Biol. Chem. 276 (2001) 6453–6462.
58 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
[20] N. Zamzami, P. Marchetti, M. Castedo, C. Zanin, J.L. Vayssiere, P.X. Petit, G. Kroemer, Reduction in mitochondrial potential constitutes an early irreversible step of programmed lymphocyte death in vivo, J. Exp. Med.
181 (1995) 1661–1672.
[21] J.N. Udall, R.A. Moscicki, F.I. Preffer, P.D. Ariniello, E.A. Carter, A.K.
Bhan, K.J. Bloch, Flow cytometry: a new approach to the isolation and characterization of Kupffer cells, Adv. Exp. Med. Biol. 216A (1987) 821–827.
[22] P. Smolewski, J. Grabarek, D. Halicka, Z. Darzynkiewicz, Assay of caspase activation in situ combined with probing plasma membrane integrity to detect three distinct stages of apoptosis, J. Immunol. Methods.
265 (2002) 111–121.
[23] A. Nel, T. Xia, L. Madler, N. Li, Toxic potential of materials at the nanolevel, Science 311 (2006) 622–627.
[24] R. Duncan, L. Izzo, Dendrimer biocompatibility and toxicity, Adv. Drug Deliv. Rev. 57 (2005) 2215–2237.
[25] Y. Aramaki, S. Takano, S. Tsuchiya, Induction of apoptosis in ma- crophages by cationic liposomes, FEBS Lett. 460 (1999) 472–476.
[26] Y. Aramaki, S. Takano, S. Tsuchiya, Cationic liposomes induce macrophage apoptosis through mitochondrial pathway, Arch. Biochem.
Biophys. 392 (2001) 245–250.
[27] T. Xia, M. Kovochich, J. Brant, M. Hotze, J. Sempf, T. Oberley, C. Sioutas, J. Yeh, M. Wiesner, A. Nel, Comparison of the abilities of ambient and
manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm, Nano Lett. 6 (2006) 1794–1807.
[28] P.S. Lai, M.J. Shieh, C.L. Pai, C.Y. Wang, P.J. Lou, Studies on the Subcellular Trafficking of PAMAM Dendrimer, Nanotech 2005 Confer- ence Abstract, 2005.
[29] O. Boussif, F. Lezoualc’h, M.A. Zanta, M.D. Mergny, D. Scherman, B.D.
Demeneix, J.P. Behr, A versatile vector for gene and oligonucleotide transfer into cells in culture and in vivo: polyethylenimine, Proc. Natl.
Acad. Sci. U. S. A. 92 (1995) 7297–7301.
[30] Z.Y. Zhang, B.D. Smith, High-generation polycationic dendrimers are usually effective at disrupting anionic vesicles: membrane bending model, Bioconjug. Chem. 11 (2000) 805–814.
[31] N. Malik, R. Wiwattanapatapee, R. Klopsch, K. Lorenz, H. Frey, J.W.
Weener, E.W. Meijer, W. Paulus, R. Duncan, Dendrimers: relationship between structure and biocompatibility in vitro, and preliminary studies on the biodistribution of125I-labelled polyamidoamine dendrimers in vivo, J. Control. Release 65 (2000) 133–148.
[32] F. Chellat, A. Grandjean-Laquerriere, R. Le Naour, J. Fernandes, L. Yahia, M. Guenounou, D. Laurent-Maquin, Metalloproteinase and cytokine production by THP-1 macrophages following exposure to chitosan-DNA nanoparticles, Biomaterials 26 (2005) 961–970.
59 J.S. Kuo et al. / Journal of Controlled Release 120 (2007) 51–59
表 Y04
行政院國家科學委員會補助參與國際學術學會會議報告
96 年 7 月 16 日
報告人姓名 郭 榮 華 服務機構
及職稱
嘉南藥理科技大學藥物科技研究所 教授
時間 地點
07/07/2007-07/11/2007;
Long Beach Convention Center, Long Beach, California U.S.A.
本會核定 補助文號 會議
名稱
(中文)第三十四屆控制釋放協會年會暨展覽會
(英文)34thAnnual Meeting & Exposition of the Controlled Release Society 發表
論文 題目
The effect of remnant cationic dendrimers on RNA purification.
附件三
表 Y04
1.參加會議經過:
第三十四屆控制釋放協會年會暨展覽會為 國 際 上 研 討 controlled release technology 之 最 大 型 國 際 會 議 。 該 會 議 每 年 召 開 一 次 , 歷 屆 會 議 均 有 眾 多 先 進 國 家 之 專 家 學 者 與 會 , 論 文 涵 蓋 理 論 與 實 務 應 用 , 層 面 廣 泛 而 多 樣 , 一 向 為 各 國 專 家 視 為 重 要 且 前 瞻 性 之 國 際 學 術 會 議 。 該 會 議 於 7 月 07 日至 11 日在 Long Beach Convention Center (Long Beach, California U.S.A.) 舉行,總共有超過 1000 人,來自四十幾個國家的專家 學者與會參加。本次會議包括 general conference; symposiums; keynote address; posters
exposition; Soapbox Session 及 workshops
。
symposiums 分別是 Advances in Peptide and Protein Delivery ; Advances in Polymer Micelles; Advances in Process Technology;Biodegradable/Biocompatible Polymers; Cellular Barriers; Industrial Session;
Gastroretention-Animal vs. Human; Advances in Vaccine Delivery; Controlled Release in Food;
Eurand Special Session; Delivery for Bioimaging; Nanotechnology; New Polymers for Drug Delivery; Advances in Colloidal Drug Delivery ; Emerging Role of Alternative delivery in Veterinary Medicine; Recent Delivery in Diabetes; Nonviral Gene Delivery; Oral Delivery;
Polymer Conjugates; Biological responses to Nanomaterials; Biomaterials and Biointerfaces;
Capsugel Special Session; Innovative Materials and Release Mechanisms; Liposomes: Alive &
Kicking; Pulmonary Delivery; Industrial Advances in Oral Delivery; Injectable Depot System;
Stimuli Responsive Nanosystems; Nanocapsulation; Personal Care; Tumor Targeting; Advances in Ocular Delivery; Advances in Transdermal Delivery; Controlled Release in Health and Wellness;
Imaging and Delivery in Clinical Settings; Targeted Delivery
。
邀 請 世 界 知 名 的 學 者 專 家 授 課 workshops 為 Transdermal Drug Delivery through Microporated Skin; Oral Extended Drug Release Based on Hydrophilic Polymers: Single and Multi-Unit Systems; Hyperspectral Chemical Imaging for the Physical and Chemical Characterization of Drugs in Polymers; Microdialysis:Continuous In Vivo or In vitro Drug & Target Collection; LipoBridge: A Molecular Queen Mary for Crossing the BBB; Liquid Filled Two-Piece Capsules for Challenging Molecules
。
本次會議中安 排了 keynote address 分別為: non-Canonical Amino Acids in Protein Design, Evolution and Analysis (D.A. Tirrell, California Institute of Technology, USA); Improved Childhood Vaccines(S.Buchsbaum, Bill and Melinda Gates Foundation, USA); Cell Sheet Tissue Engineering and Their Clinical Applications (T. Okano, Women’s Medical University, Japan); Organic Delivery Vehicles for Probing and Targeting Biological Systems (J. DeSimone, University of North Carolina,
USA);The Role of Biomarkers and Biological Interactive Delivery Systems (receptor mediated transcytosis) in the Future of Chemotherapy (P. Soon-Shiong, AbraxisBioscience, USA)
。
投 稿 本 次 會 議 之 poster 論 文 非 常 踴 躍,本次參加 poster 超過 1000 篇,各家展覽藥廠超過 100 家。
2.與會心得:
在會場有許多專家學者參加此次會議,與這些專家學者做了一些深入的討論,學術交流並汲 取新知,並且瞭解這些專家學者所做的研究。在 Symposiums 中也瞭解了其他專家學者所做的 研究,特別是在 gene delivery, nanomedicines, vaccines, polymer genomics 等方面,收穫豐富。
各家藥廠( 3M Drug Delivery Systems, Genzyme Pharmaceuticals, BASF Pharmaceutical
Systems,etc.)亦展覽最新控制釋放研發劑型,從研討會中發現應用生物技術如分子生物操作,
細胞培養,免疫測試及 DNA 轉染於控制釋放之各方面應用領域均蓬勃發展,亦為未來之主流
趨勢,對未來教學及研究非常的有幫助
。
3. 攜回資料名稱及內容:
1. 33rdAnnual Meeting and Exposition for the Controlled Release Society 論文 CD-ROM,光碟片
中含有所有在會議中發表的文章全文
。
2. 各大藥廠最新產品及精密儀器目錄
。
表 Y04