行政院國家科學委員會專題研究計畫 成果報告
高硒青花椰菜萃取液對砷去毒機制之探討(第 2 年) 研究成果報告(完整版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 95-2313-B-468-002-MY2
執 行 期 間 : 96 年 08 月 01 日至 97 年 07 月 31 日 執 行 單 位 : 亞洲大學生物科技學系
計 畫 主 持 人 : 葉貞吟 共 同 主 持 人 : 歐柏榮
計畫參與人員: 碩士班研究生-兼任助理人員:林良翰
處 理 方 式 : 本計畫可公開查詢
中 華 民 國 97 年 11 月 01 日
目錄
Introduction.................................1 Objectives..................................5 Materials & Methods..............................7 Results....................................9 Summary..................................10 Reference..................................11 Self-evaluation................................20 Table....................................21 Figures...................................22
INTRODUCTION
As toxicity and its detoxification mechanisms
Some heavy metals are industrial and environmental contaminants to which human beings could be chronically exposed (Goyer, 1986). Arsenic (As) is a ubiquitous metal in the environment and the accumulation of arsenic in ground water and plants poses a health risk to both humans and animals (Abernathy et al., 1999). Inorganic As compounds, especially the trivalent compounds, appear to be more toxic than the organic As compounds. Arsenic trioxide (As2O3) is used as a therapeutic drug in treating acute promyelocytic leukemia patients, therefore most arsenic studies have focused on the anti-leukemic effect of As2O3 in cancer cells (Gianni et al., 1998; Mccabe et al., 2000; Roboz et al., 2000; Ling et al., 2002). The toxic effects of As on animals and humans are important issues for investigation since it is a potential carcinogen (Salnikow and Cohen, 2002). Exposure to As compounds in animals and humans is reported to cause various health effects, such as Blackfoot disease, ischemic heart disease, vascular diseases, hypertension, diabetes, hepatic damages and cancers (Smith et al., 1992; Chen et al., 1995; Chiou et al., 1997; Hopenhayn et al., 1998; Santra et al., 2000; Tseng et al., 2000).
The As detoxification mechanistic paths involve antioxidative defense system and multidrug resistance transporters. Glutathione (L-r-glutamyl-L-cysteinylglycine; GSH) released continuousely from liver and heart of animals is a biologically active thiol tripeptide and the most abundant thiol in living cells that plays an important role in detoxification of exogenous compounds and oxidative insults (Reed, 1990). It is an important antioxidant in various tissues to protect the cells from oxidative injury (Walker et al., 1995). Glutathione exists as two forms in the cells, reduced form (GSH) and oxidized form (GSSG). Several antioxidative enzymes are involved in the detoxification of As compounds, such as glutathione peroxidase (GPX) family, superoxide dismutases (SODs), GSH reductase (GR) and glutathione S-transferase (GST). The GPX family is a group of selenoproteins that incorporate selenium (Se) as selenocysteine (SeCys) into the amino acid sequence. It prevents the formation of reactive oxygen species by catalyzing H2O2decomposition with the concomitant oxidation of GSH to GSSG (Wendel, 1980). GPXs are antioxidative-related enzymes with different substrate specificities and they collaborate to provide the main defense system against oxidative stress in mammalian cells for detoxification of intracellular oxidative stress (Gladyshev and Hatfield, 1999). SODs are ubiquitous metalloproteins that play a major protective role in living organisms by catalyzing the dismutation of superoxide into hydrogen peroxide and molecular oxygen. GSH reductase is the important enzyme responsible for the availability of GSH in GSH redox cycle by catalyzing the reduction of GSSG to GSH utilizing nicotinamide adenine dinucleotide phosphate (NADPH) (Meister, 1994). GSTs involve in metabolic detoxification by catalyzing the reaction between GSH and electrophilic substrates and prevent toxic damage in several tissues (Keen et al., 1976;
Habig and Jakoby, 1981).
Drug pumps are an important part of cell defense system against toxin or carcinostatic drug.
Several drug pumps are found in both mammalian cells and bacteria and these multidrug
resistance transporters are members of adenosine triphosphate (ATP)-binding cassette superfamily (Ambudkar et al., 1999; Putman et al., 2000). Two kinds of multidrug resistance pumps have been identified, P-glycoprotein transporter and Multidrug resistance protein (MRP) family. P-glycoprotein transporter is a phosphoglycoprotein encoded by a gene family that comprises MDR genes (MDR1 and MDR 3) in humans and mdr genes (mdr 1a, mdr 1b and mdr 2) in rodents, and functions as an ATP-dependent efflux pump on the cell membrane (Gottesman and Pastnan, 1993; Shapiro and Ling, 1995). The expression of human MDR1 as well as rodent mdr1a and mdr1b appears to confer multidrug resistance selectively. Active P-glycoprotein transporter is present in tumor and non-tumor tissues (Sleeman et al., 2000).
Multidrug resistance protein (MRP) family is a superfamily of transmembrane glycoproteins that mediate extracellular efflux of variety of xenobiotics and heavy metals (Keppler et al., 2000). The MRPs are organic anion transporters and they transport anionic drug and neutral drugs conjugated to acidic ligands, such as GSH, glucuronate or sulfate (Borst et al., 2000). The MRPs express in human tissues and cells with differential amounts (Cole et al., 1992;
Zaman et al., 1993). The MRP family is composed of seven members (MRP 1-7) at least (Borst et al., 2000). MRPs 1-3 share overlapping and similar substrate specificities, but their kinetic properties differ markedly (Keppler et al., 2000). MRP1 is the ATP-dependent transporter responsible for the efflux of GSH-conjugating complex out of the cell during cellular detoxification process and mediates the transport of negatively charged conjugated hydrophilic compounds with a large hydrophobic moiety such as glucuronide, sulfate and glutathione S-conjugates of drugs. MRP1 is expressed ubiquitously with a high level in kidney, lung, intestine and brain while low expression in liver (Cherrington et al., 2002). MRP2, expressed in liver, kidney and intestine, is the ATP-binding cassette protein that also known as the canalicular multispecific organic anion transporter. Treatment of As was reported to up-regulate MRP2 gene expression transcriptionally in primary rat and human hepatocytes and this regulation was related to cellular GSH level (Vernhet et al., 2001). MRP3 is also an organic anion transporter with the preference of glucuronate conjugates as substrates over GSH conjugates (Hirohashi et al., 1999).
MRP3 is expressed in pancreas and kidney with a high expression in intestines and adrenal cortex, but very low expression in liver (Cherrington et al., 2002). With the location in the basolateral membrane of hepatocyte, MRP3 may allow efflux of organic anions from the liver into the blood when secretion into bile is blocked (Kool et al., 1999a). MRP4 is suggested to be an organic anion transporter specific for phosphate conjugates. It functions as a cellular efflux pump for nucleotide analogue (Schuetz et al., 1999). MRP4 is expressed in muscle, prostate, lung, pancreas, testis, ovary, bladder and gallbladder (Borst et al., 2000). MRP5, expressed ubiquitously, is located on the basolateral membrane and is a multispecific organic anion transporter for nucleotide analogs (Wijnholds, et al., 2000). MRP6 gene is located on chromosome 16 which immediately next to MRP1. It is highly expressed in liver and kidney and to a low or very low extent in a few other tissues (Kool et al., 1999b). MRP7 is a recent addition to the MRP family (Borst et al., 2000).
Hydrogen peroxide (H2O2) production is associated with arsenic toxicity (Jing et al., 1999) and can damage vascular endothelium (Li amd Lau, 1993). Research data indicated that
As-induced oxidative stress, such as H2O2formation and lipid peroxidation, is associated with the change in intracellular GSH level and cellular regulations of antioxidative enzyme system (Han et al., 1996; Dai et al., 1999; Yeh et al., 2002). The elevations of cellular GSH level and antioxidative enzymes exert the protective effect in counteracting oxidative stress (Meister, 1991).
As was also reported to cause growth inhibition and affect cell cycle by inducing G2/M phase arrest and apoptosis through regulation of cell cycle-related proteins in a variety of human tumor cell lines (Smith and Fornace, 1996; Akao et al., 1999; Huang et al., 2000b; Park et al., 2001;
Vega et al., 2001; Vogt and Rossman, 2001; Yih and Lee, 2001; Cumming et al., 2002; Ling et al., 2002; Yeh et al., 2003). Some metals or toxins are taken up or removed from cells by GSH-mediated pathways (Ballatori, 1994). GSH and As form complex and are exported outside the cells to reduce metal toxicity (Rosen, 1999). The trivalent As compounds was reported to increase cellular GST activity (Yeh et al., 2002). GSTs are involved in the conjugation of GSH with As, thus increased GST activity by trivalent arsenic compounds suggests that the elevated intracellular GSH level in response to the oxidative stress may be used to conjugate arsenic in PAECs and facilitate the efflux of arsenic (Yeh et al., 2002). Research results from embryonic stem cells indicated that the activity of multidrug resistance transporters was associated with the change in intracellular GSH level and involved in the GSH efflux system (Rappa et al., 1997).
Thus, the As detoxification mechanism may also be mediated in part by multidrug resistance transporters. The living organisms are very complicated and the As-induced cellular responses observed during As toxicity are expected to be the combination effects for different detoxification mechanisms.
Protective effect of selenium on As-induced toxicity
Selenium (Se) is an essential trace element for humans and animals. Se deficiency results in skeletal muscle dystrophy in livestock and Keshan Disease in humans (Schubert et al., 1961;
Chen et al., 1980). Se acts as an antioxidant with anticancer effect and is a key component in selenoproteins that incorporate Se as selenocysteine translationally into the amino acid sequence, such as glutathione peroxidase (GPX) family, selenoprotein W, selenoprotein P, thioredoxin reductase, iodothyronine deiodinase and other selenoproteins (Behne and Kyriakopoulos, 2001;
Whanger, 2004). Se is known to suppress carcinogenesis nonspecifically in animals (Whanger, 2004). After oral supplementation with 200 ug Se/day as enriched yeast for 4.5 years, the cancer incidence of lung, colon and prostate cancers was significantly reduced (Clark et al., 1996; 1998).
This is consistent with other human trials that indicated that Se supplementation reduced the incidence of certain cancers (Whanger, 2004). Inhibition of cell proliferation, induction of apoptosis and elevation of antioxidative ability are important cellular responses that may be involved in the anticarcinogenic effect of Se (Sinha and El-Bayoumy, 2004). Se exerts many of its effects through modulating the functions of intracellular proteins related to cell proliferation to regulate cell cycle events in several human tumor cell lines (Ip, 1998; Venkateswaran et al., 2002).
Se as selenomethionine was reported to protect human fibroblasts and keratinocytes from DNA damage (Seo et al., 2002; Rafferty et al., 2003). Pretreatment of Se as selenite suppresses apoptosis in UVB-exposed human embryonic kidney cells (Park et al., 2000a).
Arsenic-induced toxicity is associated with increased oxidative stress through regulation of antioxidative-related enzymes and cell cycle progression (Barchowsky et al., 1996; Jing et al., 1999; Yeh et al., 2002). Se may exert its protective effect on As-induced toxicity through modulations of cell cycle-related protein and antioxidative-related selenoproteins. Recent reports indicated that Se exerts its protective effect against oxidative stress and vascular damage through regulation of selenoproteins both in vitro and in vivo (Huang et al., 2002a; Lewin et al., 2002). Se pretreatment as selenite reduced As2O3-induced cytotoxicity through GPX modulation (Yeh et al., 2003).
Potential effect of Se-enriched broccoli extract
Cancer is the leading cause for death in human health and may be prevented through improving physiological antioxidative system by consuming protective compounds or avoiding mutagen exposure (DeFlora, 1998). Natural plant extracts have gained much attention recently due to the antioxidative and antimutagenic abilities to defense the dangerous effects caused by toxins or mutagens (Bronzetti, 1994). The form and the concentration of Se used are critical for cancer prevention (Sinha and El-Bayoumy, 2004). Various Se compounds influence cellular responses differently in various cell types (Tapiero et al., 2003). With the knowledge of Se as an anticarcinogenic agent, delivery of this protective element through the food systems is a natural and harmless method to provide enrichment for humans. Since plants with high sulfur content tend to take up high levels of Se (Terry et al., 2000), broccoli (Finley et al., 2001), garlic (Ip et al., 1992), onion (Cai et al., 1995) and ramps (Whanger et al., 2000) have been enriched with Se for possible oral supplementation of Se through dietary consumption. The addition of Se-enriched broccoli (SeB) to the rat diets significantly reduced the mammary tumor incidence from 90% to 37% (Finley et al., 2001). In addition, SeB florets (Finley et al., 2000; Finley and Davis, 2001) and sprouts (Finley et al., 2001) significantly reduced colon tumors in rats.
A study investigating the differential effects of SeB extract on cell proliferation by using C6 rat glial cells indicated that the proliferation inhibition by 1000 nM Se as SeB extract was apparently related to the H2O2-dependent mechanism with elevated cellular glutathione peroxidase (cGPX) activity (Yeh et al., 2005). SeB extract also involved in the modulation of cell cycle progression and apoptosis (Yeh et al., 2005).
Effect of Se on multidrug resistance
Se was shown to inhibit the growth of drug-resistance cell lines (Caffrey and Frenkel, 1997;
Bjorkhem-Bergman et al., 2002) and to prevent the development of drug resistance (Caffrey et al., 1998). However, no available information can be found on the interaction of selenium with multidrug resistance transporters.
OBJECTIVES
Arsenic is a known endothelial toxin and has a direct effect on endothelial cells causing vascular disorders and endothelial cell dysfunction (Chang et al., 1991; Tsai et al., 2001). The vascular endothelial cells are considered to be one of the primary targets for As toxicity and are used extensively as the experimental model to examine cellular responses induced by As (Engel and Smith, 1994; Barchowsky et al., 1996). Previous in vitro studies indicated that P-glycoprotein is present and functionally active in primary cultures of rat, porcine and bovine cerebral endothelial cells (Greenwood, 1992; Huwyler et al., 1996; Fenart et al., 1998). Whereas, there were evidences indicated that the MRP expressed in primary cultures of bovine brain microvessel endothelial cells (Huai-Yun et al., 1998). It was reported that P-glycoprotein transporter and the MRP family expressed in porcine oocyte and the high homology of these multidrug resistance transporters between porcine and human genes was observed (Yashi et al., 2001). We propose to use the primary culture of porcine aortic endothelial cells (PAECs) continuously as the experimental materials in this proposed study not only for the high homology of multidrug resistance transporters observed between porcine and human gene (Yashi et al., 2001), but also for the possibly differential responses between normal and tumor cells.
The primary objective of this proposed study was to investigate the possible mechanism responsible for the protective effect of SeB extract on As-induced toxicity. The activities of cGPX and GST as well as the cell cycle progression in primary culture of porcine aortic endothelial cells (PAECs) will be investigated. The regulation of specific multidrug resistance transporters expression by Se-enriched broccoli extract was also investigated.
MATERIALS AND METHODS
Cell culture
Porcine aortic endothelial cells (PAECs) were isolated and identified previously from our laboratory (Yeh et al., 2002). The primary culture of isolated PAECs was used as the experimental materials in this study. The cells were maintained routinely and passed in sterile basal medium (M199, 100 U penicillin/mL, 100 µg streptomycin/mL, 2.2 g sodium bicarbonate/L media, pH 7.4) supplemented with 10% FBS under a humidified atmosphere of 5% CO2and 95%
air at 37oC. The culture medium was changed every 2-3 days and the microscopic examination was used to observe the cell morphology and monitor cell growth. All experiments were performed when the cells reach 80-90% confluence.
In vitro digestion of Se-enriched broccoli (SeB) extract
Broccoli extracts containing low and high levels of Se was provided by Dr. John Finley (Human Nutrition Laboratory, USDA, Grand Forks, ND, USA). Briefly, sodium selenate solutions were added to the growth media for broccoli and the broccoli extracts were prepared by in vitro enzymatic digestion of broccoli powder. The mixture of broccoli powder with ascorbic and citric acids was adjusted to pH 2 and incubated with pepsin (40 mg/ml) in 0.1 HCl for 1 hr.
Then 2.5 ml of pancreatin (2 mg/ml) and bile (12 mg/ml) in 0.1 M sodium bicarbonate were added and incubated for another 2 hr. The digested mixture was then dialyzed against distilled water overnight using dialysis bags with 12,000 to 14,000 MW cut off. The high-Se (H-SeB) and low-Se (L-SeB) broccoli extracts were analyzed to contain 1.08 x 10-4M Se and 2.50 x 10-7M Se, respectively (Yeh et al., 2005).
MTT assay
MTT assay was used to measure cell proliferation and cytotoxicity. This assay is based on the cleavage of the yellow tetrazolium salt---MTT by mitochondrial dehydrogenases of metabolically active cells to purple formazan crystals which are then solubilized and spectrophotometrically quantified (Mosmann, 1983). PAECs were seeded in 96-well tissue culture microplates at a concentration of 1x104 cells per well. Various experiments were performed after the cells reach 80-90% confluence. Prior to investigate the protective effect of SeB on As2O3-induced cytotoxicity, the effect of various Se compounds and SeB extracts on cell proliferation in PAECs was determined. The different dilutions of H-SeB and L-SeB were applied to PAECs to determine the effect of SeB extracts alone and the possible contribution of broccoli extraction buffer on cell proliferation. The different concentrations of SeB (H-SeB or L-SeB) will be added to the culture media and the plates were incubated at 37oC up to 24 hr. An appropriate concentration of SeB extracts was determined for later pretreatment experiments To investigate the protective effect of SeB extract on As2O3-induced cytotoxicity, PAECs were pretreated with or without an appropriate concentration of SeB extract or Na2SeO3 (as positive control) at 37oC for 24 hr. The media from one set of the plates was then changed to the culture media containing 20 µM of As2O3, while the media from the other set of the plates was changed to the culture
media containing 20 µM of As2O3 with addition of SeB extract or Na2SeO3. The plates were incubated at 37oC for additional 24 hr. At the end of the experiments, 10 µL of MTT stock solution (final MTT concentration: 0.5 mg/mL) was added to each well (100 µL culture media/well) and the microplates were then incubated at 37oC for additional 4 hr. After adding 100 µL of solubilization solution (10% SDS in 0.01M HCl) to each well to dissolve the purple formazan crystals, the plates were read on a microplate reader at the wavelength of 590 nm.
Cell treatments for cell cycle progression, enzymatic assays and RT-PCR analysis
PAECs were cultured on 10-cm tissue culture plates, then were pretreated with or without an appropriate concentration of various selenocompounds or SeB extracts at 37oC for 24 hr. To investigate the protective effect of Se pretreatment on As2O3-induced cell cycle progression and enzyme activities, the media from one set of the plates was changed to the regular culture media (DMEM+10% FBS),while the media from the other set of the plates were changed to the culture media containing 20 µM of As2O3. The plates were then incubated at 37oC for additional 24 hr. At the end of the experiments, the plates were subjected to the following procedure for determination of cell cycle progression or frozen in -80oC after three washes with ice-cold PBS for further enzymatic analysis or RT-PCR analysis.
Cell cycle progression analysis
To determine the cell cycle phase-distribution, both detached and adherent cells were harvested from the plates at the end of experiments, washed with ice-cold PBS and resuspended in 8 mL of ice-cold 70% ethanol to fix overnight at -20oC. Centrifugation was performed at 300 x g for 5 min at 4oC the next day and 0.5 mL of 0.5% Triton X-100 and 2 µL of RNase A was added to resuspend the cell pellets. The mixture was incubated in a waterbath at 37oC for 30 min, and the supernatant was discarded following centrifugation at 300 x g for 5 min at 4oC. Then 0.5 mL of propidium iodide stock solution (20 µg/mL in PBS) was added to the pellet and this mixture was incubated for 10 min on ice. The mixture was then applied to the Flow Cytometer for analysis (approximately 10000 events).
Intracellular antioxidative enzyme activities
To prepare cell cytosols, an appropriate amount of homogenate buffer (20 mM Tris [pH 7.5], 0.25 M sucrose, 1mM EGTA, 5mM EDTA, 1mM PMSF, 50 mM beta-mercaptoethanol and 25 ug/ml leupeptin; pH 7.4) was added to each frozen plate and the cells were scraped into
centrifuge tubes by rubber policeman. After sonication for 10 sec on ice, the mixtures were centrifuged at 17000 x g for 10 min at 4oC. Protein concentration of cell cytosols was determined by the Bradford assay (Bradford, 1976) using bovine serum albumin as standards. The cell cytosols were then be used for measurement of cGPX and GST activities.
Cellular glutathione peroxidase activity was determined by an enzyme-coupled method with glutathione reductase, utilizing hydrogen peroxide as substrate (Paglia and Valentine, 1967) with
a DU Series 60 Spectrophotometer (Beckman Instruments, Fullerton, CA USA) at 30oC. The rate of decrease in the NADPH concentration was followed at 340 nm over the 3-min period at 30-s intervals. Glutathione-S-transferase (GST) activity was determined spectrophotometrically at 340 nm by monitoring the formation of the conjugated glutathione and CDNB at 25oC (Habig and Jakoby, 1981). The rate of increase in the absorbance at 340 nm was monitored over the 3-min period at 30-s intervals.
Total RNA extraction and RT-PCR analysis of multidrug resistance transporters
Due to the low expression level for multidrug resistance transporters, reverse transcription-polymerase chain reaction (RT-PCR) assay was used to identify the differential expression of multidrug resistance transporters in PAECs. The total RNA was extracted from PAECs using Trizol reagent (Invitrogen). The RNA was quantitated by spectrophotometer at a wavelength of 260/280 nm prior to RT-PCR analysis. RT reaction was performed using the commercial RT kit (TAKARA). Subsequent PCR reaction was performed using various specific primers for multidrug resistance transporters. The specific primers for various multidrug resistance transporters were designed from the published literatures and the sequences in GenBank (Liu et al., 2001; Vernhet et al., 2001; Yashi et al., 2001). The GAPDH primers will be used as the internal control.
Statistical analysis
Data were examined for equal variance and normal distribution prior to statistical analysis.
Mean values were compared by analysisofvariance(ANOVA)with Fisher’sleastsignificant difference (LSD) method for comparing groups (Steel and Torrie, 1980). A significance level of 5% was adopted for all comparisons.
RESULTS
1. Based on the MTT results, various selenocompounds as well as Se-enriched broccoli extracts exhibited higher protection in As2O3-induced toxicity at the concentration of 1 X 10-6M for 24 hr (Fig. 1) than at 1 X 10-7M (Fig 2). Thus, the concentration for Se pretreatment was determined to be 1 X 10-6M.
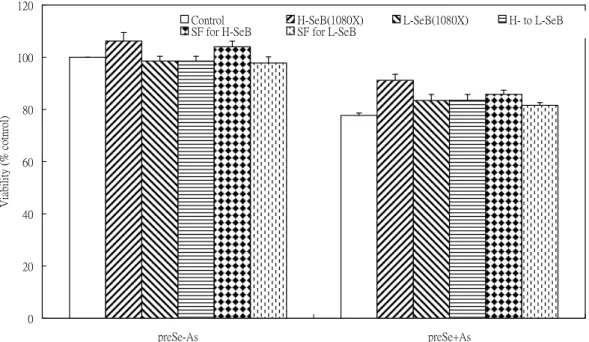
2. The protective effect observed with HSeB pretreatment for 24 hr in As2O3-induced toxicity is due to the high selenium content in HSeB and may be related to the sulforaphane (SF) content in HSeB (Figs. 3-4).
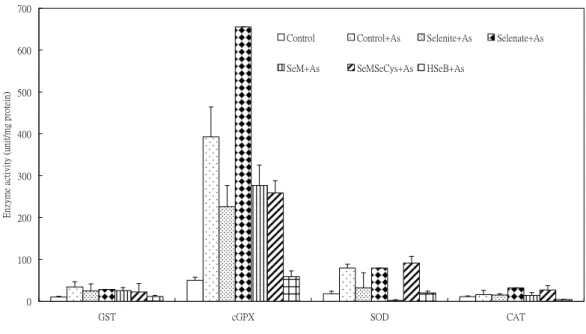
3. Pretreatment with various selenocompounds alone for 24 hr did not affect SOD (superoxide dismutase) and CAT (catalase) activities in PAECs. The GST activity in PAECs was increased by HSeB pretreatment for 24 hr and the cGPX activity was increased by selenate pretreatment (Fig. 5). Pretreatment of various selenocompounds did not affect the GST activity in As2O3-induced PAECs, whereas the As2O3-induced increase in cGPX activity was further increased by selenate pretreatment (Fig. 6).
4. The increased in GST activity in PAECs induced by HSeB pretreatment may be due to SF effect (Fig. 7).
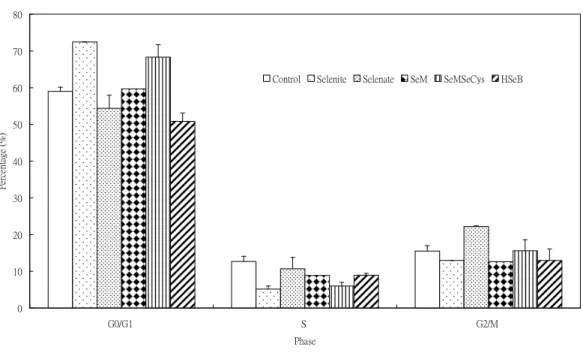
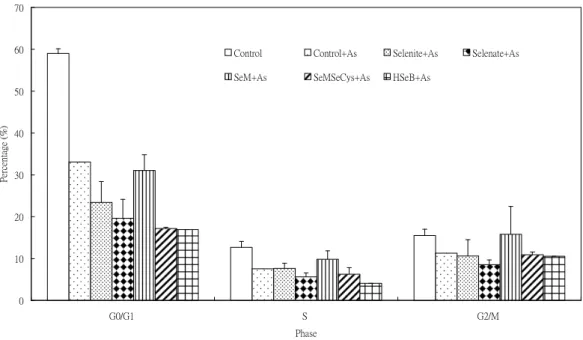
5. Pretreatment with selenite or SeM (selenomethionine) for 24 hr resulted in an increase in cell population in G0/G1phase (Fig. 8). There is no effect by Se pretreatment in cell population in S and G2/M phase. Se pretreatment did not influence the As2O3-induced cell cycle progression (Fig. 9).
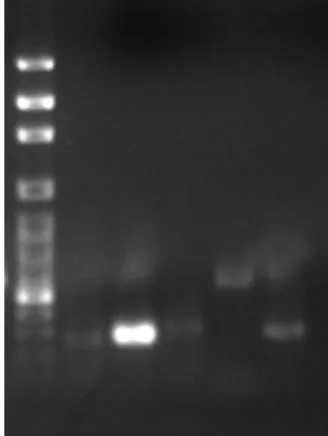
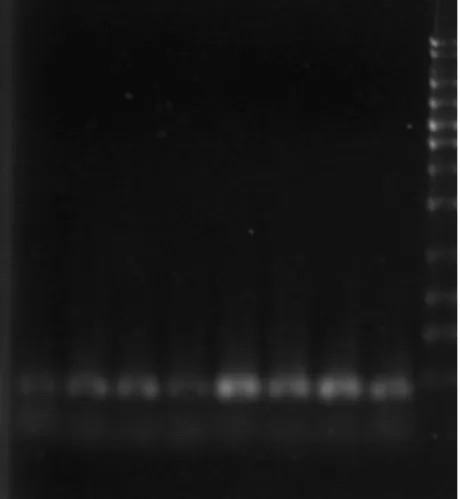
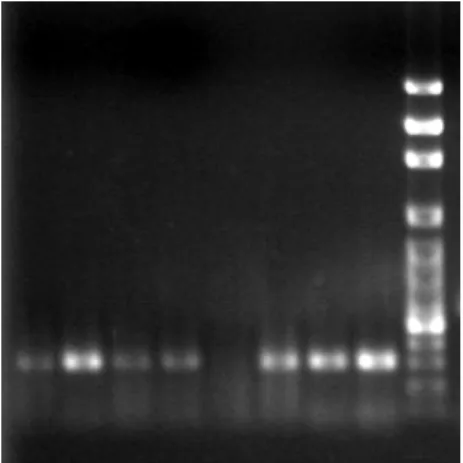
6. The specific primers used for multidrug resistance associated transporters in RT-PCR analysis are listed in Table 1. The mRNAs of GST2, MDR1, MRP1 and MRP5 were expressed in PAECs (Fig. 10). Using GAPDH mRNA as internal control (Fig. 11), the results from RT-PCR analysis indicated that there is no difference among pretreatment of Se-enriched broccoli extracts in As2O3-induced gene expression of multidrug resistance transporters (Figs.
12-15).
SUMMARY
The results in this study indicated that Se-enriched broccoli extracts protected porcine aortic endothelial cells (PAECs) from arsenic-induced toxicity mainly through modulation of glutathione S-transferase (GST) activity, but not mRNA expression. Multidrug resistance associated transporters, MDR1, MRP1 and MRP5 were not involved in selenium protective mechanism for arsenic toxicity. However, other multidrug resistnace associated transporters may be involved in the possible mechanism. Therefore, further investigation of expression of other multidrug resistance associated transporters in As-induced toxicity is needed.
REFERENCE
Abernathy, C. O., Liu, Y. P., Longfellow, D., Aposhian, H. V., Beck, B., Fowler, B., Goyer, R., Menzer, R., Rossman, T., Thompson, C. and Waalkes, M. (1999) Arsenic: health effects, mechanisms of actions, and research issues. Environ. Health Perspect. 107: 593-597.
Akao, Y., Nakagawa, Y. and Akiyama, K. (1999) Arsenic trioxide induces apoptosis in neuroblastoma cell lines through the activation of caspase 3 in vitro. FEBS Lett. 455: 59-62.
Ambudkar, S. V., Dey, S., Hrycyna, C. A., Ramachandra, M., Pastan, I. and Gottesman, M. M.
(1999) Biochemical, cellular and pharmacological aspects of the multidrug transporter. Annu. Rev.
Pharmacol. Toxicol. 39: 361-398.
Barchowsky, A., Dudek, E. J., Treadwell, M. D. and Wetterhahn, K. E. (1996) Arsenic induces oxidant stress and NF-B activation on cultured aortic endothelial cells. Free Radical Biology &
Medicine 21:783-790.
Barchowsky, A., Klei, L. R., Dudek, E. J., Swartz, H. M. and James, P. E. (1999) Stimulation of reactive oxygen, but not reactive nitrogen species, in vascular endothelial cells exposed to low levels of arsenite. Free Radical Biol. & Med. 27: 1405-1412.
Behne, D. and Kyriakopoulos, A. (2001) Mammalian selenium-containing proteins. Annu. Rev.
Nutr. 21:453-473.
Bjorkhem-Bergman, L., Jonsson, K., Eriksson, L. C., Olsson, J. M., Lehmann, S., Paul, C. and Bjornstedt, M. (2002) Drug-resistant human lung cancer cells are more sensitive to effects on thioredoxin reductase and glutathione reductase. Biochem. Pharmacol. 63:1875-1884.
Bradford, M. M. (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
Borst, P., Evers, R., Kool, M. and Wijnholds, J. (2000) A family of drug transporters: the multidrug resistance-associated proteins. J. Natl. Cancer Inst. 92: 1295-1302.
Bronzetti, G. (1994) Antimutagens in food. Trends in Food Science and Techno. 5: 390-395.
Caffrey, P. B. and Frenkel, G. D. (1997) Sensitivity of melphalan-resistant tumors to selenite in vivo. Cancer Lett. 121:177-180.
Caffrey, P. B., Zhu, M. and Frenkel, G. D. (1998) Prevention of the development of melphalan resistance in vitro by selenite. Biol. Trace Elem. Res. 65:187-195.
Cai, X.-Y., Block, E., Uden, P. C., Zhang, X., Quimby, B. D., Sullivan, J. J. (1995) Allium chemistry: identification of selenoamino acids in ordinary and selenium enriched garlic, onions, and broccoli using gas chromatography with atomic emission detection. J. Agric. Food Chem. 43:
1754-1760.
Chang, W. C., Chen, S. H., Wu, H. L., Shi, G. Y., Murota, S. I. and Morita, I. (1991) Cytoprotective effect of reduced glutathione in arsenical-induced endothelial injury. Toxicology 69: 101-110.
Chen, C. J., Hsueh,, Y. M., Lai, M. S., Shyu, M. P., Chen, S. Y., Wu, M. M. et al. (1995) Increased prevalence of hypertension and long-term exposure. Hypertension 25: 53-60.
Chen, X. S., Yang, G. Q., Chen, J. S., Chen, X. C., Wen, G. M. and Ge, K. Y. (1980) Studies in the relations of selenium and Keshan disease. Biol. Trace Elem. Res. 2:91-107.
Cherrington, N. J., Hartley, D. P., Johnson, N. L. D. R. and Klaassen, C. D. (2002) Organ distribution of multidrug resistance proteins 1, 2, and 3 (Mrp 1, 2, and 2) mRNA and hepatic induction of Mrp3 by constitutive androstance receptor activators in rats. J. Pharmacol. Exper.
Ther. 300: 97-104.
Chiou, H. Y., Huang, W. I., Su, C. L., Chang, S. F., Hsu, Y. H. and Chen, C. J. (1997) Dose-response relationship between prevalence of cerebrovascular disease and ingested inorganic arsenic. Stroke 28: 1717-1723.
Clark, L. C, Combs, G. F. Jr, Turnbull, B. W., et al. (1996) Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial.
Nutritional Prevention of Cancer Study Group. J. Am. Med. Assoc. 276: 1957-1963.
Clark, L. C., Dalkin, B., Krongrad, A., et al. (1998) Decreased incidence of prostate cancer with selenium supplementation: results of a double-blind cancer prevention trial. Brit. J. Urol. 81:
730-734.
Cole, S.P.C., Bhardwaj, G., Gerlach, J. H., Mackie, J. E., Grant, C. E., Almquist, K. C., Stewart, A.
J., Kurz, E. U., Duncan, A. M. V. and Deeley, R. G. (1992) Overexpression of a transporter gene in a multidrug-resistant human lung cancer cell line. Science 258: 1650-1654.
Cummings, M., Siitonen, T., Higginbottom, K., Newland, A. C. and Allen, P. D. (2002) p53-mediated downregulation of chk1 abrogates the DNA damage-induced G2M checkpoint in K562 cells, resulting in increased apoptosis. Br. J. Haematol. 116: 421-428.
Dai J., Weinberg R.S., Waxman S. and Jing Y. (1999) Malignant cells can be sentized to undergo growth inhibition and apoptosis by arsenic trioxide through modulation of the glutathione redox system. Blood 93:268-277.
DeFlora, S. (1998) Mechanisms of inhibitors of mutagenesis and carcinogenesis. Mutat. Res. 402:
151-158.
Engel, R. R. and Smith, A. H. (1994) Arsenic in drinking water and mortality from vascular disease: An ecologic analysis in 30 countries in the United States. Arch. Environ. Health 49:
418-427.
Fenart. L., Buee-Scherrer, V., Descamps, L., Duhem, C., Poullain, M. G., Cecchelli, R. and Dehouck, M. P. (1998) Inhibition of P-glycoprotein: rapid assessment of its implication in blood-brain barrier integrity and drug transport to the brain by an in vitro model of the blood-brain barrier. Pharm. Res. 15: 993-1000.
Finley, J. W, Davis, C. D. (2001) Selenium (Se) from high-selenium broccoli is utilized differently than selenite, selenate and selenomethionine, but is more effective in inhibiting colon carcinogenesis. Biofactors 14: 191-196.
Finley JW, Davis CD, Feng Y. (2000) Selenium from high selenium broccoli protects rats from colon cancer. J Nutr 130: 2384-2389.
Finley JW, Ip C, Lisk DJ, Davis CD, Hintze K, Whanger PD. (2001) Cancer-protective properties of high-selenium broccoli. J Agric Food Chem 49: 2679-2685.
Gianni M., Koken M. H. M., Chelbi-Alix M. K., Benoit G., Lanotte M., Chen Z. and de The H.
(1998) Combined arsenic and retinoic acid treatment enhances differentiation and apoptosis in arsenic-resistant NB4 cells. Blood, 91, 4300-4310.
Gladyshev V. N. and Hatfield D. L. (1999) Selenocysteine-containing proteins in mammals.
Journal of Biomedical Science 6: 151-160.
Gottesman, M. M. and Pastan, I. (1993) Biochemistry of multidrug resistance mediated by the multidrug transporter. Annu. Rev. Biochem. 62: 385-427.
Goyer, R. A. (1986) Toxic effects of metals. In: Toxicology: the basic science of poisons (Klaassen, C. D., Amdur, M. O. and Doull, J., eds), Pp. 582-635., Macmillan Publishing Co., New York. USA.
Greenwood, J. (1992) Characterisation of a rat endothelial cell culture and the expression of P-glycoprotein in brain and retinal endothelium in vitro. J. Neuroimmunol. 39: 123-132.
Habig W. H. and Jakoby W. B. (1981) Assays for differentiation of glutathione S-transferases.
Methods Enzymol. 77: 398-405.
Han S. K., Mytilineou C. and Cohen G. (1996) L-DOPA up-regulates glutathione and protects mesencephalic cultures against oxidative stress. J. Neurochem. 66:501-510.
Hirohashi, T., Suzuki, H. and Sugiyama, Y. (1999) Characterization of the transport properties of cloned rat multidrug resistance-associated protein 3 (MRP3). J. Biol. Chem. 274: 15181-15185.
Hong, D. H., Huan, J., Ou. B. R., Yeh, J. Y., Saido, T. C., Cheeke, P. R. and Forsberg, N. E. (1995) Protein kinase C isoforms in muscle cells and their regulation by phorbol ester and calpain.
Biochim. Biophys. Acta 1267: 45-54.
Hopenhayn-Rich C., Biggs M. L. and Smith A. H. (1998) Lung and kidney cancer mortality associated with arsenic in drinking water in Cordoba, Argentina. Intl. J. Epidemiol. 27: 561-569.
Huai-Yun, H., Secrest, D. T., Mark, K. S., Carney, D., Brandquist, C., Elmquist, W. F. and Miller, D. M. (1998) Expression of multidrug resistance-associated protein (MRP) in brain microvessel endothelial cells. Biochem. Biophys. Res. Commun. 243: 816-820.
Huang K., Liu H., Chen Z. and Xu H. (2002a) Role of selenium in cytoprotection against cholesterol oxide-induced vascular damage in rats. Atherosclerosis 162: 137-144.
Huang S. C., Huang C. Y. F. and Lee T. C. (2000b) Induction of mitosis-mediated apoptosis by sodium arsenite in HeLa S3 cells. Biochemical Pharmacology 60: 771-780.
Huwyler, J., Drewe, J., Klusemann, C. and Fricker, G. (1996) Evidence for P-glycoprotein-modulated penetration of morphine-6-glucuronide into brain capillary endothelium. Br. J. Pharmacol. 118: 1879-1885.
Ip C. (1998) Lessons from basic research in selenium and cancer prevention. J. Nutr. 128:
1845-1854.
Ip C, Lisk DJ, Stoewsand GS. (1992) Mammary cancer prevention by regular and selenium-enriched garlic. Nutr Cancer 17: 279-284.
Jing Y., Dai J., Chalmers-Redman R. M. E., Tatton W. G. and Waxman S. (1999) Arsenic trioxide selectively induced acute promyelocytic leukemia cell apoptosis via hydrogen peroxidase-dependent pathway. Blood 94: 2101-2111.
Keen J. H., Habig W. H. and Jakoby W. B. (1976) Mechanism for the several activities of the glutathione S-transferases. J. Biol. Chem. 251: 6183-6188.
Keppler, D., Kamisako, T., Leuer, I., Cui, Y., Nies, A. T., Tsujii, H. and Konig, J. (2000) Localization, substrate specificity, and drug resistance conferred by conjugate export pumps of the MRP family. Advan. Enzyme Regul. 40: 339-349.
Kool, M., van der Linden, M., de Haas, M., Scheffer, G. L., de Vree, J. M., Smith, A. J., et al.
(1999a) MRP3, an organic anion transporter able to transport anti-cancer drugs. Proc. Natl. Acad.
Sci. USA 96: 6914-6919.
Kool, M., van der Linden, M., de Haas, M., Baas, F. and Borst, P. (1999b) Expression of human MRP6, a homologue of the multidrug resistance protein gene MRP1, in tissues and cancer cells.
Cancer Res. 59: 175-182.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
Lewin M. H., Arthur J. R., Riemersma R. A., Nicol F., Walker S. W., Millar E. M., Howie A. F.
and Beckett G. J. (2002) Selenium supplementation acting throught the induction of thioredoxin reductase and glutathione peroxidase protects the human endothelial cell line Eahy926 from damage by lipid hydroperoxides. Biochimica et Biophysica Acta 1593: 85-92.
Li L. and Lau B. J. (1993) A simplified in vitro model of oxidant injury using vascular endothelial cells. In Vitro Cell. Dev. Biol. 29A: 531.
Ling Y. E., Jiang J. D., Holland J. F. and Perez-Soler R. (2002) Arsenic trioxide produces polymerization of microtubules and mitotic arrest before apoptosis in human tumor cell lines.
Molecular Pharmacology 62: 529-538.
Liu, J., Chen, H., Miller, D. S., Saavedra, J. E., Keefer, L. K., Johnson, D. R., Klaassen, C. D. and Waalkes, M. P. (2001) Overexpression of glutathione S-transferase II and multidrug resistance transport proteins is associated with acquired tolerance to inorganic arsenic. Mol. Pharmacol. 60:
302-309.
Mccabe M. J. Jr., Singh K. P., Reddy S. A., Chelladurai B., Pounds J. G., Reiners J. J. Jr. and States J. (2000) Sensitivity of myelomonocytic leukemia cells to arsenite-induced cell cycle disruption, apoptosis, and enhanced differentiation is dependent on the inter-relationship between arsenic concentration, duration of treatment, and cell cycle phase. Journal of Pharmacology &
Experimental Therapeutics 295: 724-733.
Meister A. (1991) Glutathione deficiency produced by inhibition of its synthesis, and its reversal:
applications in research and therapy. Pharmacol. Ther. 51:155-194.
Meister A. (1994) Glutathione, ascorbate and cellular protection. Cancer Res. 54: 1969s-1975s.
Mosmann, T. (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunological Methods 65:55-63.
Paglia D. E. and Valentine W. N. (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 70: 158-169.
Park J. W., Choi Y. J., Jang M. A., Baek S. H., Lim J. H., Passaniti T. and Kwon T. K. (2001) Arsenic trioxide induces G2/M growth arrest and apoptosis after caspase-3 activation and bcl-2 phosphorylation in promonocytic U937 cells. Biochemical & Biophysical Research Communications 286: 726-734.
Putman, M., van Veen, H. W. and Koning, W. N. (2000) Molecular properties of bacterial multidrug transporters. Microbiol. Mol. Biol. Rev. 64: 672-693.
Rafferty TS, Green MH, Lowe JE et al. (2003) Effects of selenium compounds on induction of DNA damage by broadband ultraviolet radiation in human keratinocytes. Br J Dermatol 148:
1001-1009.
Rappa, G., Lorico, A., Flavell, R. A. and Sartorelli, A. C. (1997) Evidence that the multidrug resistance protein (MRP) functions as a co-transporter of glutathione and natural product toxins.
Cancer Res. 57: 5232-5237.
Reed D. J. (1990) Glutathione: toxicological implication. Annu. Rev. Pharmacol. Toxicol. 30:
603-631.
Roboz G. J., Dias S., Lam G., Lane W. J., Soignet S. L., Warrell R. P. J. and Rafii S. (2000) Arsenic trioxide induces dose- and time-dependent apoptosis of endothelium and may exert an antileukemic effect via inhibition of angiogenesis. Blood 96: 1525-1530.
Salnikow K. and Cohen M. (2002) Backing into cancer: effect of arsenic on cell differentiation.
Toxicological Science 65: 161-163.
Santra A., Maiti A., Das S., Lahiti S., Charkaborty S. K. and Mazumder D. N. G. (2000) Hepatic damage caused by chronic arsenic toxicity in experimental animals. Clin. Toxicol. 38: 395-405.
Schubert, J. R., Muth, O. H., Oldfield, J. E. and Remmert, L. F. (1961) Experimental results with selenium in white muscle disease of lambs and calves. Fed. Proc. 10:689-692.
Schuetz, J. D., Connelly, M. C., Sun, D., Paibir, S. G., Flynn, P. M., Srinivas, R. V., et al. (1999) MRP4: a previously unidentified factor in resistance to nucleoside-based antiviral drugs. Nat.
Med. 5: 1048-1051.
Seo YR, Sweeney C, Smith ML. (2002) Selenomethionine induction of DNA repair response in human fibroblasts. Oncogene 21: 3663-3669.
Shapiro, A. B. and Ling, V. (1995) Reconstitution of drug transport by purified P-glycoprotein. J.
Biol. Chem. 270: 16167-16175.
Sinha R, El-Bayoumy K. (2004) Apoptosis is a critical cellular event in cancer chemoprevention and chemotherapy by selenium compounds. Curr Cancer Drug Targets 4: 13-28.
Sleeman, M. A., Watson, J. D. and Murison, J. G. (2000) Neonatal murine epidermal cells express a functional multidrug-resistance pump. J. Invest. Dermatol. 115: 19-23.
Smith A. H., Hopenhayn-Rich C., Bates M. N., Goeden H. M., Hertz-Picciotto I., Duggan H. M.
et al. (1992) Cancer risks from arsenic in drinking water. Environmental Health Perspectives 97:259-267.
Smith M. L. and Fornace A. J. Jr. (1996) Mammalian DNA damage-inducible gene associated with growth arrest and apoptosis. Mutation Research 340: 109-124.
Steel R. G. D. and Torrie L. H. (1980) Principles and Procedures of Statistics. McGraw-Hill, New York, New York USA.
Tapiero H, Townsend DM, Tew KD. (2003) The antioxidant role of selenium and seleno-compounds. Biomed Pharmacother 57: 134-144.
Terry N, Zayed AM, deSouza MP, Tarun AS. (2000) Selenium in higher plants. Annu Rev Plant Physiol 51: 401-420.
Towbin, H., Staehelin, T., and Gordon, J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedures and some applications. Proc. Natl. Acad.
Sci. USA, 76: 4350-4354.
Tsai S. H., Hsieh M. S., Chen L., Liang Y. C., Lin J. K. and Lin S. Y. (2001) Suppression of Fas ligand expression on endothelial cells by arsenite through reactive oxygen species. Toxicology Lett. 123: 11-19.
Tseng C. H., Tai T. Y., Chong C. K., Tseng C. P., Lai M. S., Lin B. J. et al. (2000) Long-term arsenic exposure and incidence of non-insulin-dependent diabetes mellitus: a cohort study in arseniasis-hyperendemic villages in Taiwan. Environ. Health Perspect. 108: 847-851.
Vega L., Styblo M., Pattersons R., Cullen W., Wang C. and Germolec, D. (2001) Differential effects of trivalent and pentavalent arsenicals on cell proliferation and cytokine secretion in normal human epidermal keratinocytes. Toxicology and Applied Pharmacology 172: 279-286.
Venkateswaran V, Klotz LH, Fleshner NE. (2002) Selenium modulation of cell proliferation and cell cycle biomarkers in human prostate carcinoma cell lines. Cancer Res. 62: 2540-2545.
Vernhet, L., Seite, M-P., Allain, N., Guillouzo, A. and Fardel, O. (2001) Arsenic induced expression of the multidrug resistance-associated protein 2 (MRP2) gene in primary rat and human hepatocytes. J. Pharmacol. Exper. Ther. 298: 234-239.
Vogt B. L. and Rossman T. G. (2001) Effects of arsenite on p53, p21 and cyclin D expression in normal human fibroblasts --- apossiblemechanism forarsenite’scomutagenicity.Mutat. Res.
487: 159-168.
Walker, M. W., Kinter, M. T., Roberts, R. J. and Spitz, D. R. (1995) Nitric oxide-induced xytotoxicity: involvement of cellular resistance to oxidative stress and the role of glutathione in protection. Pediatr. Res. 37:41.
Wendel A. (1980) Glutathione peroxidase. In: Enzymatic basis of detoxication, pp. 333-353, Academic Press, San Diego, California USA.
Whanger PD. (2004) Selenium and its relationship to cancer: an update. Br. J. Nutr. 91: 11-28.
Whanger PD, Ip C, Polan CE, Uden PC, Welbaum G. (2000) Tumorigenesis, metabolism, speciation, bioavailability, and tissue deposition of selenium in selenium-enriched ramps (Allium tricoccum). J. Agric. Food Chem. 48: 5723-5730.
Wijnholds, J., Mol, C. A. A. M., van Deemter, L., de Haas, M., Scheffer, G. L., Baas, F., Beijnen, J. H., Scheper, R. J., Hatse, S., de Clercq, E., Balzarini, J. and Borst, P. (2000) Multidrug-resistance protein 5 is a multispecific organic anion transporter able to transport nucleotide analogs. Proc. Natl. Acad. Sci. USA 97: 7476-7481.
Yashi, Y. T., Nakayama, K., Fujioka, T., Kanzaki, A., Mutho, M., Uchida, T., Miyazaki, K., Ito, M.
and Fukumoto, M. (2001) Expression of multidrug resistance associated transporters (MDR1, MRP1, LRP and BCRP) in porcine oocyte. International J. of Mol. Med. 7: 397-400.
Yeh, J. Y., Cheng, L. C., Liang, Y. C. and Ou, B. R. (2003) Modulation of the arsenic effects on cytotoxicity, cell viability and cell cycle in porcine endothelial cells by selenium. Endothelium 10: 127-139.
Yeh, J. Y., Cheng, L. C., Ou, B. R., Whanger, P. D. and Chang, L. W. (2002) Differential influences of various arsenic compounds on glutathione redox status and antioxidative enzymes in porcine endothelial cells. Cell. Mol. Life Sci. 59:1972-1982.
Yeh, J. Y., Ou, B. R., Liang, Y. C., Burchfiel, J., Butler, J. A., Forsberg, N. E. and Whanger, P. D.
(2005) Mechanism for proliferation inhibition by various selenium compounds and selenium-enriched broccoli extract in rat glial cells. Biometals (accepted)
Yih L. H. and Lee T. C. (2000) Arsenite induces p53 accumulation through an ATM-dependent pathway in human fibroblasts. Cancer Res. 60:6346-6352.
Zaman, G. J. R., Versantvoort, C. H. M., Smit, J. J. M., Eijdems, E. W. H. M., de Haas, M., Smith, A. J., Broxterman, H. J., Mulder, N. H., de Vries, E. G. E., Baas, F. and Borst, P. (1993) Analysis of the expression of MRP, the gene for a new putative transmembrane drug transporter, in human multidrug resistant lung cancer cell lines. Cancer Res. 53: 1747-1750.
SELF-EVALUATION
The results from this study are promising. Further investigation of regulation of other specific multidrug resistance transporters by Se-enriched broccoli extract will help to understand the mechanism responsible for the protective effect of Se-enriched broccoli extracts on arsenic-induced toxicity.
Table 1. Specific primers used for RT-PCR analysis
Primer Sequence(5'→3') PCR product
MDR1-F GCCTGGCAGCTGGAAGACAAATACACAAAATT
283 bp
MDR1-R CAGACAGCAGCTGACAGTCCAAGAACAGGACT
MRP1-F CACACTGAATGGCATCACCTTC 317 bp
MRP1-R CCTTCTCGCCAATCTCTGTCC
MRP5-F ACATCACGTCCAGCATACAGG 594 bp
MRP5-R TGCAGCCTCCAGATAACTCC
GST 2-F GATGGGGTGGAGGACCTTCGATGC 272 bp
GST 2-R CTGAGGCGAGCCACATAGGCAGAG
GAPDH-F AGTCGGAGTCAACGGATTTG 305 bp
GAPDH-R TCTCCATGGTGGTGAAGACA
Figure 1. Pretreatment of various selenocompounds (1X10-6M Se) for 24 hr followed by 20 uM As2O3 treatment for 24 hr
0 20 40 60 80 100 120 140
preSe-As preSe+As
Treatment
Viability(%control)
Control Selenite Selenate SeMSeCys SeMet H-SeB(108X)
Figure 2. Pretreatment of various selenocompounds (1X10-7M Se) for 24 hr followed by 20 uM As2O3 treatment for 24 hr
0 20 40 60 80 100 120 140
preSe-As preSe+As
Treatment
Viability(%control)
Control Selenite Selenate SeMSeCys SeMet H-SeB(1080X)
Figure 3. Pretreatment of Se-enriched broccoli extracts (108X dilution) and sulforaphane for 24 hr followed by 20 uM As2O3 treatment for 24 hr
0 20 40 60 80 100 120
preSe-As preSe+As
Treatment
Viability(%control)
Control H-SeB(108X) L-SeB(108X) H- to L-SeB SF for H-SeB SF for L-SeB
Figure 4. Pretreatment of Se-enriched broccoli extracts (1080X dilution) and sulforaphane for 24 hr followed by 20 uM As2O3 treatment for 24 hr
0 20 40 60 80 100 120
preSe-As preSe+As
Treatment
Viability(%cotnrol)
Control H-SeB(1080X) L-SeB(1080X) H- to L-SeB
SF for H-SeB SF for L-SeB
Figure 5. Pretreatment effect of various Se compounds (1 X 10-6M) for 24 hr on enzyme activities in PAECs
0 20 40 60 80 100 120 140 160 180
GST cGPX SOD CAT
Enzymeactivity(unit/mgprotein)
Control Selenite Selenate SeM SeMSeCys HSeB
Figure 6. Pretreatment effect of various Se compounds (1 X 10-6M) for 24 hr on As2O3-induced enzyme activities in PAECs
0 100 200 300 400 500 600 700
GST cGPX SOD CAT
Enzymeactivity(unit/mgprotein)
Control Control+As Selenite+As Selenate+As
SeM+As SeMSeCys+As HSeB+As
Figure 7. Pretreatment effect of Se-enriched brocooli extracts and sulforaphane for 24 hr on enzyme activities in PAECs
0 100 200 300 400 500
GST cGPX SOD CAT
Enzymeactivity(unit/mgprotein)
Control HSeB LSeB SF
Control+As HSeB+As LSeB+As SF+As
Figure 8. Pretreatment effect of various Se compounds (1 X 10-6M) for 24 hr on cell cycle progression in PAECs
0 10 20 30 40 50 60 70 80
G0/G1 S G2/M
Phase
Percentage(%)
Control Selenite Selenate SeM SeMSeCys HSeB
Figure 9. Pretreatment effect of various Se compounds (1 X 10-6M) for 24 hr on As2O3-induced cell cycle progression in PAECs
0 10 20 30 40 50 60 70
G0/G1 S G2/M
Phase
Percentage(%)
Control Control+As Selenite+As Selenate+As
SeM+As SeMSeCys+As HSeB+As
1 2 3 4 5 6
Figure 10. The mRNA Expression of GST2, MDR1, MRP1, MRP5 and GAPDH in PAECs. Lane 1: 100bp marker; Lane 2: GST 2; Lane 3: MDR1; Lane 4: MRP1; Lane 5: MRP5 and Lane 6: GADPH.
1 2 3 4 5 6 7 8 9
Figure 11. GAPDH mRNA expression by Se-enriched broccoli extracts and sulforaphane (SF) pretreatments for 24 hr in As2O3-induced PAECs. Lane 1: HSeB; Lane 2: LSeB;
Lane 3: SF; Lane 4: control+As; Lane 5: HSeB+As; Lane 6: LSeB+As; Lane 7:
SF+As; Lane 8: control and Lane 9: 100 bp marker.
1 2 3 4 5 6 7 8 9
Figure 12. GST2 mRNA expression by Se-enriched broccoli extracts and sulforaphane (SF) pretreatments for 24 hr in As2O3-induced PAECs. Lane 1: control; Lane 2: HSeB;
Lane 3: LSeB; Lane 4: SF; Lane 5: control+As; Lane 6: HSeB+As; Lane 7:
LSeB+As; Lane 8: SF+As and Lane 9: 1 kb marker.
1 2 3 4 5 6 7 8 9
Figure 13. MDR1 mRNA expression by Se-enriched broccoli extracts and sulforaphane (SF) pretreatments for 24 hr in As2O3-induced PAECs. Lane 1: control; Lane 2: HSeB;
Lane 3: LSeB; Lane 4: SF; Lane 5: control+As; Lane 6: HSeB+As; Lane 7:
LSeB+As; Lane 8: SF+As and Lane 9: 100 bp marker.
1 2 3 4 5 6 7 8 9
Figure 14. MRP1 mRNA expression by Se-enriched broccoli extracts and sulforaphane (SF) pretreatments for 24 hr in As2O3-induced PAECs. Lane 1: control; Lane 2: HSeB;
Lane 3: LSeB; Lane 4: SF; Lane 5: control+As; Lane 6: HSeB+As; Lane 7:
LSeB+As; Lane 8: SF+As and Lane 9: 100 bp marker.
1 2 3 4 5 6 7 8 9
Figure 15. MRP5 mRNA expression by Se-enriched broccoli extracts and sulforaphane (SF) pretreatments for 24 hr in As2O3-induced PAECs. Lane 1: 100 bp marker; Lane 2:
control; Lane 3: HSeB; Lane 4: LSeB; Lane 5: SF; Lane 6: control+As; Lane 7:
HSeB+As; Lane 8: LSeB+As and Lane 9: SF+As.