Glutamine supplementation enhances mucosal immunity in rats with gut- derived sepsis
Yu-Ni Lai*, Sung-Ling Yeh
*, Ming-Tsan Lin
+, Chiu-Li Yeh,
*Wei-Jao Chen
+*
Institute of Nutrition and Health Sciences, Taipei Medical University, Taipei, Taiwan
+
Department of Surgery, College of Medicine, National Taiwan University, Taipei, Taiwan, ROC
Running title: glutamine on mucosal immunity in sepsis
Reprint requests:
Sung-Ling Yeh, PhD
Institute of Nutrition and Health Sciences, Taipei Medical University 250 Wu-Hsing Street, Taipei, Taiwan 110, Republic of China
Tel: 8862-27361661 ext. 6551-115 Fax: 8862-27373112
E-mail: [email protected]
Abstract
Objective: Supplemental glutamine (Gln) has been demonstrated to improve the
immunologic response and reduce mortality in rodents with sepsis. However, the effects of Gln on gut-associated lymphoid tissue function after infection and sepsis are not clear. The aim of this study was to study the effect of Gln-supplemented diets before and Gln-enriched total parenteral nutrition (TPN) after sepsis or both on the intestinal immunity in rats with gut-derived sepsis.
Methods: Male Wistar rats were assigned to control and 4 experimental groups. The
control and the experimental groups 1 and 2 were fed a semipurified diet, while in the diets of groups 3 and 4, part of the casein was replaced with Gln. After feeding rats the respective diets for 10 d, sepsis in the experimental groups was induced by cecal ligation and puncture (CLP), whereas the control group underwent sham operation; at the same time, the internal jugular vein was cannulated. All rats were maintained on TPN for 3 d. The control group and groups 1 and 3 were infused with conventional TPN, while groups 2 and 4 were given a TPN solution supplemented with Gln, which replaced 25% of total amino acids nitrogen. All rats were sacrificed 3 d after sham operation or CLP. Intestinal immunoglobin (Ig) A levels, total lymphocyte yields and lymphocyte subpopulations in Peyer’s patches were analyzed.
Results: Total Peyer’s patch lymphocyte numbers were significantly higher in the Gln
supplemented groups than the control group. The distributions of CD3+, CD4+ in the group 1 were significantly lower than the control group, whereas no differences were observed among the control and Gln supplemented groups. Plasma IgA levels were higher in the Gln supplemented groups than the control group and group 1. Intestinal IgA levels were significantly higher in the groups 2 and 4 than in the control group and group 1.
Conclusions: preventive use of a Gln -supplemented enteral diet or Gln started
intravenously after CLP have similar effects on promoting proliferation of total lymphocyte numbers in GALT, enhancing IgA secretion and maintained payer’s patch T lymphocyte populations. Gln administered both before and after CLP did not seem to have a synergistic effect on enhancing mucosal immunity in rats with gut- derived sepsis.
Key words: glutamine; Peyer’s patches; immunoglobin A; gut-derived sepsis
Introduction
Sepsis is a frequent clinical problem in patients undergoing major surgery. Under
a condition of sepsis, bacterial toxins insult the body, and profound tissue breakdown
and alterations in substrate metabolism may occur (1,2). The development of total
parenteral nutrition (TPN) has afforded a means to provide adequate calories and
protein to patients whose gastrointestinal tract is unusable. However, TPN has been shown to cause atrophy of the gut and impairment of gut-associated lymphoid tissue (GALT) is often observed (3-5). Small intestinal GALT is a major contributor to primary immunologic protection at all mucosal sites (6). Peyer’s patches have attracted special attention, because the genesis of generalized mucosal immunity resides within the Peyer’s patches in the GALT that process intraluminal antigen and stimulate naïve B and T cells to those antigens (7). Studies have shown that B and T cell populations in Peyer’s patches were significantly reduced when TPN was administered (5,8). Also, intestinal interleukin (IL)-4 and IL-10 accompanied by immunoglobulin (Ig) A levels decreased in parenterally fed animals (9). Clinically, enteral nutrition is considered to be a better delivery route for critically ill patients compared with those who were fed parenterally (10,11). However, sepsis has been shown to reduce mesenteric blood flow, and adversely affect the barrier and metabolic functions of the small intestine (12,13). Gardiner et al. (14) suggested that under the condition of gut-derived sepsis, the parenteral rather than the enteral route had
benefits of improving survival in rats.
Glutamine (Gln) is a semi-essential amino acid that has been shown to posses
numerous useful physiologic properties (15). A study by Inoue et al.16 demonstrated
that bowel weight was greater and disaccharidase activity of the intestinal mucosa was
higher in an intravenous Gln-treated group than the control group in a peritonitis model. Naka et al.17 examined the effect of an intravenous Gln dipeptide on septic mortality. They found that mortality was significantly lower in the Gln-TPN group than in animals receiving conventional TPN. The beneficial effects of Gln were thought to result from its protective effect on increasing intestinal blood flow and in maintaining a functioning bowel barrier.
18,19Human studies also reported that the Gln-treated group experienced fewer clinical infections and shorter hospital stays.
20,21Some investigators suggest that Gln may be useful in the treatment of established infections or inflammation.
15To our knowledge, there is no study, so far, investigating the effects of Gln on GALT function after infection and sepsis.
Therefore, the aim of this study was to study whether Gln-supplemented diets before and Gln-enriched TPN after sepsis or both enhanced the function of Peyer’s patches and their ability to secrete IgA. In this study, we used cecal ligation and puncture (CLP) as a sepsis model, because CLP is more clinically relevant and is considered to be a simple and reproducible model of gut-derived sepsis in rats (22).
Materials and methods
Animals. Male Wistar rats weighing 230-250 g were housed in stainless steel cages
maintained in a temperature- and humidity-controlled room with a 12:12-h light-dark
cycle. Animals were allowed free access to a standard rat chow for 3 d prior to the
experiment. All procedures conducted in this study were approved by National
Taiwan University Animal Care Committee.
Surgical procedure and grouping. All rats were divided into the control and 4 experimental groups. The control group and experimental groups 1 and 2 were fed a common semipurified diet. Rats in the experimental groups 3 and 4 were fed an identical diet except that part of the casein was replaced by Gln, which provided 25%
of the total amino nitrogen (Table 1). After feeding rats the respective diets for 10 days, sepsis in the experimental groups was induced by CLP, whereas the control group underwent sham operation. CLP was performed according to the method of Wichterman et al. (22) Briefly, rats were anesthetized with intraperitoneal
pentobarbital (50 mg/kg), and the abdomen was opened through a midline incision.
The cecum was isolated, and a 3-0 silk ligature was placed around it, ligating the
cecum just below the ileocecal valve. The cecum was then punctured twice with an
18-gauge needle and was replaced back into the abdomen. The abdominal wound was
closed in 2 layers. Immediately after sham or CLP operation, all rats underwent
placement of a catheter for TPN infusion. A silicon catheter (Dow Corning, Midland,
MI, USA) was inserted into the right internal jugular vein. The distal end of the
catheter was tunneled subcutaneously to the back of the neck, and exited through a
coiled spring, which was attached to a swivel allowing free mobility of animals inside
individual metabolic cages. Two milliliters per h of TPN was administered on the
first day. Full-strength TPN was given thereafter. The infusion speed was controlled
by a Terufusion pump (Model STC-503, Terumo, Tokyo, Japan). The TPN solution without fat was prepared in a laminar flow hood. Sterilized fat emulsions were added to the TPN solution daily just before use. The TPN solution was infused for the entire day at room temperature. All rats were maintained on TPN for 3 d. No enteral
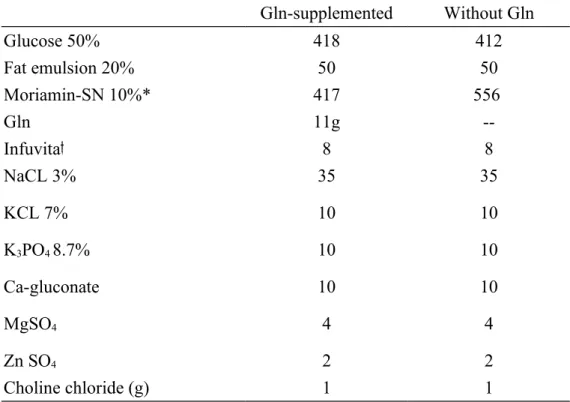
nutrition was administered during the period of TPN. The control group and the experimental groups 1 and 3 were infused with conventional TPN. Experimental groups 2 and 4 were supplemented with Gln, which replaced 25% of total amino acids nitrogen in the TPN solution. TPN provided 280 kcal/kg body weight, and the kcal/nitrogen ratio was 120:1. The TPN solutions were isonitrogenous and identical in nutrient composition except for the difference in the amino acid content (Table 2).
There were 5 groups in this study: control group, without CLP, Gln was not
supplemented before and after sham operation; group 1, Gln was not supplemented before and after CLP (-/-); group 2, semipurified diet before and Gln-containing TPN after CLP (-/+); group 3, Gln-supplemented diet before and conventional TPN after CLP (+/-); and group 4, Gln-supplemented diet before and Gln-containing TPN after
CLP (+/+).
Plasma Gln level analysis. Continuous TPN infusion was maintained until the time
of sacrifice. All rats were anesthetized then sacrificed by drawing arterial blood from
the aorta of the abdomen. Blood samples were collected in tubes containing heparin
and immediately centrifuged. Plasma amino acid was analyzed by standard ninhydrin technology (Beckman Instrument, model 6300, Palo Alto, CA), after deprotienization
of the plasma with 5% salicylic acid (23).
Lymphocyte isolation from Peyer’s patches. On day 3 after sham operation or
CLP, all surviving rats (control: 9/9; -/-: 11/17; -/+: 9/13; +/-: 9/14 and +/+: 11/16)
were weighed and anesthetized with pentobarbital. The survival rates did not differ
among the 4 experimental groups. A middle abdominal incision was made, and the
intestine from each rat was carefully removed from the stomach-duodenum junction
to the ileum ascending colon junction. Total numbers of Peyer’s patches were
counted from each removed small intestine, and lymphocytes were isolated from
Peyer’s patches as previously described (24). Briefly, Peyer’s patches were excised
from the serosal side of the intestine and teased apart with 18-gauge needles. The
fragments were treated with 50 units/mL of type 1 collagenase (Gibco Life
Technologies, Rockville, MD, USA) in RPMI-1640 supplemented with 100 g/mL of
penicillin and streptomycin for 60 min at 37
oC with constant shaking. After
collagenase digestion, the cell suspensions were passed through nylon filters, and
washed 3 times with Hank’s balanced salt solution (HBSS). The cell numbers were
adjusted to 2 ×10
6cells/mL and were suspended in RPMI-1640 containing 10% fetal
calf serum, 2 mM Gln, and antibiotics.
Cytokine immunoassay. Phytohemagglutinin (PHA, 200 ng/mL; Sigma) was used
to stimulate cytokine production by Peyer’s patch lymphocytes in culture. Triplicate
wells of 96-well flat-bottomed microtiter plates (Falcon, Becton Dickinson, Oxford,
CA, USA) were seeded with lymphocytes (2 x 10
6cells/ml in RPMI-1640) and
mitogen. The control well contained cells plus an equal volume of medium. After
incubating PHA for 24 h at 37
oC in a CO
2incubator, supernatants were centrifuged
and stored at –70
oC until being analyzed for cytokine. Interleukin (IL)-2, IL-4, IL-
10, and interferon (IFN)- concentrations in payer’s patch lymphocyte supernatants
were determined by commercially available enzyme-linked immunosorbent assay
(ELISA) kits (Amersham Pharmacia Biotech, Buckinghamshire, UK).
Lymphocytes subpopulation distribution. To determine the phenotypes of
lymphocytes isolated from Peyer’s patches, 10
5cells were suspended in 100 l HBSS
containing fluorescein-conjugated mouse anti-rat CD3 (Serotec, Oxford, UK) and
phycoerythrin-conjugated mouse anti-rat CD45Ra (Serotec) to distinguish T cells and
B cells, respectively. Fluorescein-conjugated mouse anti-rat CD8 and phycoerythrin-
conjugated mouse anti-rat CD4 (Serotec) to identify T helper cells and cytotoxic T
lymphocyte cells, respectively. After staining for 15 min, 1 mL red blood cell (RBC)
lysing buffer (Serotec) was added to lyse the RBCs and to fix the stained
lymphocytes. Fluorescence data were collected on 5 x 10
4viable cells and analyzed
by flow cytometry (Coulter, Miami, FL, USA).
Antibody quantitative analysis. IgA was measured in plasma and intestinal washings.
The collection of intestinal washings was modified from a previously described procedure (25). After excising Peyer’s patches, a 12-cm small intestine fragment proximal to the cecum was removed; the intestinal was washed intraluminally with 5 mL PBS with 1% protease inhibitor (Sigma, St Louis, MO, USA). Debris was removed by centrifugation for 10 min at 3000 rpm to harvest the supernatant for analysis of intestinal IgA after an appropriate dilution. The plasma and intestinal IgA level was determined by sandwich-type ELISA, in which anti-rat IgA capture antibody (clone A93-3; PharMingen, San Diego, CA, USA) was coated on the plate and detected with peroxidase-labeled anti-human IgA detecting antibody (clone A93-
2; PharMingen). IgA in the intestinal washing was expressed as mg/mL.
Statistical analysis. Data are expressed as the mean SD. Differences among groups were analyzed by ANOVA using Duncan’s test. A p value of < 0.05 was considered statistically significant.
Results
Body weight and plasma Gln levels. There were no differences in initial body
weights and body weights after feeding the experimental diets for 10 days, or after
TPN administration for 3 days among the 5 groups (data not shown). The Gln-
supplemented groups (groups 2, 3, and 4) had significantly higher plasma Gln levels
than did the group 1 and had no difference from the control group (Fig. 1).
Total Peyer’s patch lymphocyte yields. The total number of Peyer’s patches on the small intestine were significantly higher in group 4 (+/+) than in the control and group 1 (-/-), and had no difference from the groups 2 and 3 (-/+, +/-) (Fig 2A). The total Peyer’s patch lymphocyte yields were significantly higher in the 3 Gln supplemented groups than the control group, whereas no difference was observed
between the control group and group 1 (Fig 2B).
Lymphocyte subpopulations. There were no differences in the distributions of CD45Ra+and CD8+ cells in Peyer’s patches among the 5 groups. The distributions of CD3+, CD4+ in the group 1 (-/-) were significantly lower than the control group, whereas no differences were observed among the control and Gln supplemented
groups (Fig. 3).
Plasma and intestinal IgA levels. Plasma IgA levels were significantly higher in the Gln supplemented groups than the control and group 1. IgA levels in intestinal lavage fluid were significantly higher in the group 2 and 4 (-/+, +/+) than in the
control and group 1 (-/-). (Fig. 4).
In vitro cytokine production. The IL-10 secreted by peyer’s patch lymphocytes
after stimulation were significantly lower in the experimental groups than the control
group. IFN- levels did not differ among all groups, while the IL-2 and IL-4 levels
were not detectable (Fig 5A, B).
Discussion
Supplemental Gln has been demonstrated to improve the immunologic response in both in vivo and in vitro studies. The clinical relevance of these effects has been documented in several animal and human experiments (26-30). In this study, 25% of total amino nitorgen was supplied by Gln, because this amount of Gln was found to enhances murine immune responses (29,30). We administered TPN for 3 d after CLP because in a preliminary study, we found that the total numbers of Peyer’s patches on the serosal side of the intestine were much greater on the 3
rdday than on any other day after CLP. In this study, we did not observe a survival benefit on Gln
supplementation after CLP. Since survival was only noted for 3 days, whether Gln supplementation may improve survival over a longer period required further
investigation.
Payer’s patches are specialized lymphoid aggregates within the gastrointestinal tract (6). Naïve lymphocytes are sensitized within Peyer’s patches, proliferate within the mesenteric lymph nodes, and migrate via the thoracic duct to the lamina propria of various mucosal sites where they produce secretory IgA (6). In this study, we found that the total peyer’s patch lymphocyte yields were higher in the Gln-
supplemented groups than in the control group, whereas no difference were observed
between the control group and group 1. This result indicated that Gln
supplementation enhance the proliferation of total lymphocyte number in GALT in
gut-derived peritonitis. In order to understand the effect of Gln on the distribution of
total B lymphocytes (CD45Ra+), total T cells (CD3+), helper T cells (CD4+), and
cytotoxic T cells (CD8+) within Peyer’s patches, the subpopulations of lymphocytes
were evaluated. The results showed that the distributions of B and cytotoxic T cells
in Peyer’s patches did not differ among the groups. However, the distributions of
CD3+, CD4+ in the group 1 (-/-) were significantly lower than the control group,
whereas no differences were observed among the control and Gln supplemented
groups. This finding indicated that Gln supplementation before or/and after CLP
maintained total T and helper T lymphocyte populations in peyer’s patches in septic
condition. This result was similar to the report by Alverdy et al. (31) that addition of
Gln to the standard TPN solution maintained CD4+ cell in gut laminar propria at
levels similar to chow-fed animals. Secretory IgA is the principle effector of specific
immunity against invading pathogens (32). In this study, we found that plasma and
intestinal IgA levels were significantly higher in the groups 2 and 4 (-/+, +/+) than the
groups without Gln. Also, plasma IgA concentrations were higher in group 3 than the
control and group 1. These finding indicated that Gln supplementation before or/and
after CLP enhanced the secretion of IgA. Since there were no differences in peyer’s
patch lymphocyte yields, lymphocyte distribution and IgA production among the Gln supplemented groups. Gln administered both before and after CLP seemed to have no synergistic effect on enhancing mucosal immunity.
Cytokines are peptides produced by cells of the immune system that act as mediator of immune response and the response of tissues to injury. In order to understand the possible roles of cytokine secretions on intestinal immunity, IL-2, IFN-, IL-4, and IL-10 productions in mitogen-stimulated Peyer’s patch lymphocyte cultures were analyzed. IL-2 and IFN- are produced by Th1 lymphocytes. Th1 cytokines enhance cell-mediated immunity. A predominant Th1 effect results in activation of T lymphocytes. Th2 cytokines, including IL-4 and IL-10, enhance humoral immunity. A predominant Th2 effect results in activation of B lymphocytes and up-regulation of antibody production (33). The effects of Th1 or Th2
lymphocytes are counter-regulatory (34). Study by Kudsk et al.9 reported that Gln- enriched TPN had a normalizing effect of intestinal IL-4 and IL-10 production, which may consequently preserve intestinal IgA levels. We analyzed in vitro cytokine production by Payer’s patches lymphocytes, and found that the IL-2, IL-4 and IFN-
were either undetectable, or no differences among the 5 groups after mitogen
stimulation. IL-10 production was significantly lower in the 4 experimental groups
than in the control group, and no difference was observed among the experimental
groups. In this study, we did not observe a comparable changes between intestinal IgA and in vitro peyer’s patches IL-4 or IL-10 production. It is possible that the peak acting time of IL-4 or IL-10 precede the time we took the measurement, or these cytokines are bioactive at levels well below the range of detectability by current immunoassays. Quantitation of cytokine mRNA expression in spleen and Peyer’s patches is now under investigation in our laboratory.
In conclusion, the results of this study show that preventive use of a Gln -supplemented enteral diet or Gln started intravenously after CLP have similar effects on promoting proliferation of total lymphocyte numbers in GALT, enhancing IgA secretion, maintained Payer’s patch T lymphocyte populations and plasma Gln levels in septic rats. Gln administered both before and after CLP did not seem to have a synergistic effect on enhancing mucosal immunity in rats with gut-derived sepsis.
Acknowledgements
This study was supported by research grant NSC 91-2320-B-038-017 from National Science Council, ROC.
References
1. Clowes GHA, O’Donnell TF, Ryan NT, et al. Energy metabolism in sepsis:
treatment based on different patterns in shock and high output stage. Ann Surg 1974;179:684
2. O’Donnell TF, Clowes GHA, Blackburn GL, et al. Proteolysis associated with a deficit of peripheral energy fuel substrate in septic men. Surgery 1976;80:192 3. Alverdy JC, Aoys E, Moss GS. Total parenteral nutrition promotes bacterial
translocation from the gut. Surgery 1988; 104: 185-190
4. Purandare S, Offenbartl K, Westerom B, Bengmark S. Increased gut permeability to fluorescein isothiocyanate-dextran after total parenteral nutrition in the rat. J Gastroenterol 1989; 24: 678-682
5. Li J, Kudsk KA. Gocinski B. Dent D. Glezer J. Langkamp-Henken B. Effects of parenteral nutrition on gut-associated lymphoid tissue. J Trauma 1995; 39: 44-52 6. Salmi M, Jalkanen S. Regulation of lymphocyte traffic to mucosa-associated
lymphatic tissues. Gastroenterol Clin North Am 1991; 20: 495-510
7. McGhee JR, Mestecky J, Dertzbaugh MT, et al. The mucosal immune system:
from fundamental concepts to vaccine development. Vaccine 10:75-88, 1992 8. King BK, Li J, Kudsk KA. A temporal study of TPN-induced changes in gut-
associated lymphoid tissue and mucosal immunity. Arch Surg 11997; 32: 1303- 1309
9. Kudsk KA, Wu Y, Fukatsu K, Zarzaur BL. Johnson CD. Wang R. et al.
Glutamine-enriched total parenteral nutrition maintains intestinal interleukin-4 and mucosal imunoglobulin A levels. J Parenter Enter Nutr 2000; 24: 270-275 10. Kudsk KA, Croce MA, Fabian TC, Minard G. Tolley EA. Poret HA. et al.
Enteral versus parenteral feeding. Effects on septic morbidity after blunt and
penetrating abdominal trauma. Ann Surg 1992; 215: 503-513
11. Moore FA, Noore EE, Jones TN. TEN vs. TPN following major abdominal
trauma: reduced septic morbidity. J Trauma 1989; 29: 916-923
12. Navaratnam RL, Morris SE, Traber DL, Flynn J. Woodson L. Linares H. et al.
Endotoxin (LPS) increases mesenteric vascular resistance (MVR) and bacterial translocation (BT). J Trauma 1990; 30: 1104-1113
13. Gardiner K, Barbul A. Intestinal amino acid absorption during sepsis. J Parenter
Enter Nutr 1993; 17: 277-283
14. Gardiner KR, Gardiner RE, Barbul A. Reduced intestinal absorption of arginine
during sepsis. Crit Care Med 1995; 23: 1227-1232
15. Wilmore DW, Shabert JK. Role of glutamine in immunologic response.
Nutrition 1998;14:618-626
16. Inoue Y, Grant JP, Snyder PJ: Effect of glutamine-supplemeted intravenous nutrition on survival after Escherichia coli-induced peritonitis. JPEN 17:41-46, 1993
17. Naka S, Saito H, Hashiguchi Y, et al: Alanyl-glutamine-supplemented total parenteral nutrition improves survival and protein metabolism in rat protracted bacterial peritonitis model. JPEN 20:417-423, 1996
18. Houdijk APJ, van Leeuwen PAM, Boermeester MA, et al: Glutamine-enriched enteral diet increases splenic blood flow in the rat. Am J Physiol 267:G1035- 1040, 1994
19. Yoshida S, Leskiw MJ, Schluter MD, et al: Effect of total parenteral nutrition, systemic sepsis, and glutamineon gut mucosa in rats. Am J Physiol 263:E368- 373, 1992
20. Morlion BJ, Stehle P, Wachtler P, et al. Total parenteral nutrition with glutamine
dipeptide after major abdominal surgery. Ann Surg 1998;227:302-308
21. Houdijk APJ, Rijnsburger ER, Jansen J, et al. Randomized trial of glutamine- enriched enteral nutrition on infectious morbidity in patients with multiple trauma. Lancet 1998;352:772-776
22. Wichterman KA, Baue AE, Chaudry IH. Sepsis and septic shock-a review of
laboratory models and a proposal. J Surg Res 1980; 29: 189-201
23. Smith RJ, Panico K. Automated analysis of o-phthalaldehyde derivatives of amino acids in physiological fluids of reverse phase high performance liquid chromatography. J Liq Chromatogr 1985; 8: 1783-1795
24. Janu P, Li J, Renegar KB, Kudsk KA. Recovery of gut-associated lymphoid tissue and upper respiratory tract immunity after parenteral nutrition. Ann Surg 1992; 225: 707-717
25. Perdigon G, Alvarez S, Pesce de Ruiz Holgado A. Immunoadjuvant activity of oral Lactobacillus casei: influence of dose on the secretory immune response and protective capacity in intestinal infections. J Dairy Res 1991; 58: 485-496
26. Heberer M, Babst R, Juretic A, et al. Role of glutamine in the immune response in critical illness. Nutrition 1996;12(suppl):S71-S72
27. O’Riordain M, Fearon KC, Ross JA, et al: Glutamine supplemental parenteral nutrition enhances T-lymphocyte response in surgical patients undergoing colorectal resection. Ann Surg 220:212-221, 1994
28. Furukawa S, Saito H, Inoue T, et al. Supplemental glutamine augments
phagocytosis and reactive oxygen intermediate production by neutrophils and
monocytes from postoperative patients in vitro. Nutrition 2000;16:323-329
29. Kew S, Wells SM, Yaqoob P, et al. Dietary glutamine enhances murine T-
lymphocyte responsiveness. J Nutr 1999;129:1524-1531
30. Well AM, Kew S, Yaqoob P, et al. Dietary glutamine enhances cytokine
production by murine macrophages. Nutrition 1999;15:881-884
31. Alverdy JA, Aoys E, Weiss-Carrington P, et al. The effect of glutamine-enriched
TPN on gut immune cellularity. J Surg Res 1992;52:34-38
32. Svanborg C. Bacterial adherence and mucosal immunity. In: Handbook of Mucosal Immunology. Ogra PL, Lamm ME, McGhee JR, Mestecky J, Strober W, Bienenstock J (eds). Academic Press, New York, 1994, pp 71-8
33. DiPiro JT. Cytokine networks with infection: mycobacterial infections, leishmaniasis, human immunodeficiency virus infection, and sepsis.
Pharmacotherapy 1997; 17: 205-223
34. Parronchi P, DeCarli M, Manetti R, Simonelli C. Sampognaro S. Piccinni MP. et al. IL-4 and IFN (and) exert opposite regulatory effects on the development of cytolytic potential of Th1 or Th2 human T cell clones. J Immunol 1992; 149:
2977-2983
FIGURE LEGENDS
Fig. 1 Plasma Gln levels among the groups. Gln levels in group 1 were significantly lower than those of other groups. Different letters indicate a significant difference among the groups. (control, n = 9; group 1, n = 11; group 2, n = 9; group 3, n = 9;
group 4, n = 11).
Fig. 2 (A) Total number of Peyer’s patches on the small intestine and (B) the number of lymphocyte in Peyer’s patches of the control group and the experimental groups 3 d after CLP. Different letters indicate a significant difference among the groups.
(control, n = 9; group 1, n = 11; group 2, n = 9; group 3, n = 9; group 4, n = 11).
Fig. 3 The distribution of CD45Ra
+, CD3
+, CD4
+, and CD8
+lymphocytes in Peyer’s patches among the control and the experimental groups 3 d after CLP. Different letters indicate a significant difference among the groups. (control, n = 9; group 1, n =
11; group 2, n = 9; group 3, n = 9; group 4, n = 11).
Fig. 4 Plasma and intestinal immunoglobulin A (IgA) concentrations in the control and experimental groups 3 d after CLP. Different letters indicate a significant difference among the groups. (control, n = 9; group 1, n = 11; group 2, n = 9; group
3, n = 9; group 4, n = 11).
Fig. 5 Concentrations of A) IL-2, IL-4, IL-10 and B) IFN- released by peyer’s patches lymphocytes stimulated by PHA for 24 h. IL-2 and IL-4 were undetectable.
There were no differences in IFN- among the groups. (control, n = 9; group 1, n =
11; group 2, n = 9; group 3, n = 9; group 4, n = 11).
Table 1 Composition of the semipurified diet (g/kg)
Ingredients Gln-supplemented Without Gln
Casein 165 220
Gln 45 --
Total nitrogen 34.4 34.4
Corn starch 667 657
Soybean oil 44 44
Vitamin
*10 10
Salt mixture 35 35
Methyl-cellulose 30 30
Choline chloride 1 1
DL-methionine 3 3
*