多功能性樹枝型高分子基因傳送系統之設計與開發 研究成果報告(精簡版)
計 畫 類 別 : 個別型
計 畫 編 號 : NSC 98-2221-E-041-002-
執 行 期 間 : 98 年 08 月 01 日至 99 年 07 月 31 日 執 行 單 位 : 嘉南藥理科技大學藥物科技研究所
計 畫 主 持 人 : 郭榮華
計畫參與人員: 碩士班研究生-兼任助理人員:丘學承 碩士班研究生-兼任助理人員:劉孟捷
報 告 附 件 : 出席國際會議研究心得報告及發表論文
處 理 方 式 : 本計畫可公開查詢
中 華 民 國 99 年 10 月 01 日
Evaluating the Gene-Expression Profiles of HeLa Cancer Cells Treated with Activated and Nonactivated Poly(amidoamine) Dendrimers, and Their DNA
Complexes
Jung-hua Steven Kuo,* Meng-jie Liou, and Hsueh-chen Chiu Graduate Institute of Pharmaceutical Science, Chia Nan UniVersity of Pharmacy and
Science, 60 Erh-Jen Road, Sec. 1, Jen-Te, Tainan 717, Taiwan
Received December 8, 2009; Revised Manuscript Received March 2, 2010; Accepted April 15, 2010
Abstract:Using dendrimers in cancer therapy as nonviral vectors for gene delivery seems promising. The biological performance of a dendrimer-based gene delivery system depends heavily on its molecular architecture. The transfection activity of dendrimers is significantly improved by processes activated in the heat degradation treatment of solvolysis. However, very little is known about the molecular mechanisms that dendrimers produce in cancer cells. We studied the changes in global gene-expression profiles in human cervical cancer HeLa cells exposed to nonactivated and activated poly(amidoamine) (PAMAM) dendrimers, alone or in complexes with plasmid DNA (dendriplexes). Real-time quantitative reverse transcriptase- polymerase chain reaction was used to confirm four regulated genes (PHF5A, ARNTL2, CHD4, and P2RX7) affected by activated dendrimers and dendriplexes. Activated and nonactivated dendrimers and dendriplexes alike induced multiple gene expression changes, some of which overlapped with their dendriplexes. Dendrimer activation improved transfection efficiency and induced additional gene expression changes in HeLa cells. Dendrimers and dendriplexes principally affect genes with the molecular functions of nucleic acid binding and transcription activity, metal-ion binding, enzyme activity, receptor activity, and protein binding. Our findings provide a deeper insight into the changes in gene expression patterns caused by the molecular structure of PAMAM dendrimers for gene-based cancer therapy.
Keywords: Poly(amidoamine) (PAMAM) dendrimers; nonviral vectors; microarrays; human cervical cancer HeLa cells; activation
Introduction
The lack of a safe and efficient vector system for gene delivery is a major limiting factor for gene therapy.1,2 Nonviral vectors are safer and more versatile than their more
efficient viral counterparts, which may induce adverse immunological responses in ViVo.3Generally, nonviral vec- tors interact with anionic DNA to form highly condensed complexes for increased binding to the cell surface, enter the cell via a vesicular transport mechanism, protect DNA from endosomal degradation, and are subsequently trans- ported into the nucleus for delivered gene expression.4 Among nonviral vectors, cationic dendrimers such as poly(pro- pylene imine) (PPI) and poly(amidoamine) (PAMAM) have unique nanocharacteristics and deliver nucleic acid in Vitro
* To whom correspondence should be addressed: Chia Nan University of Pharmacy and Science, Graduate Institute of Pharmaceutical Science, 60 Erh-Jen Road, Sec. 1, Jen-Te, Tainan 717, Taiwan. Tel: 886-6-266-4911ext 2104. Fax: 886- 6-266-6411. E-mail: [email protected].
(1) Seow, Y.; Wood, M. J. Biological gene delivery vehicles: beyond viral vectors. Mol. Ther. 2009, 17, 767–777.
(2) Kawakami, S.; Higuchi, Y.; Hashida, M. Nonviral approaches for targeted delivery of plasmid DNA and oligonucleotide. J. Pharm.
Sci. 2008, 97, 726–745.
(3) Gao, X.; Kim, K. S.; Liu, D. Nonviral gene delivery: what we know and what is next. AAPS J. 2007, 9, E92–104.
(4) Louise, C. Nonviral vectors. Methods Mol. Biol. 2006, 333, 201–
226.
10.1021/mp900303s 2010 American Chemical Society VOL. 7, NO. 3, 805–814 MOLECULAR PHARMACEUTICS 805
and in ViVo.5 The biological performances of dendrimer- based gene delivery systems depend heavily on their mo- lecular architecture. For example, processes activated in solvolytic heat-degradation treatment significantly improve dendrimer transfection activity.6-8Activated dendrimers are more flexible structures that are compact in a DNA complex and swell when released from the DNA complex.7
Cationic dendrimers induce multiple biological effects in Vitro and in ViVo. The intrinsic antitumor property of cationic PPI dendrimers synergistically improves the therapeutic effects of anticancer gene therapy.9,10Cationic dendrimers, when delivering plasmid and small interfering RNA (siRNA), cause marked global gene expression changes in human cells.11-13 The gene expression altered by dendrimers includes defense responses, cell proliferation, and apoptosis.
Cationic PPI dendrimers in DNA complexes (dendriplexes) altered gene expression profiles.11 Also, differences in the structural architecture of the dendrimers have opposing effects on targeted gene expression.12 The extent of den- drimer-induced gene expression changes depends on the dendrimer’s architecture and on the cell type.11Understand- ing the influences and consequences of dendrimer-induced gene changes should help us design and develop gene-based delivery systems.
The primary objective of this study was to explore, using high-density microarray analysis, the alteration of whole
genomic responses in human cervical HeLa cancer cells treated with nonactivated (intact) and activated PAMAM dendrimers alone or with their dendriplexes.
Materials and Methods
Materials. Starburst PAMAM dendrimer (Generation 5 (PAMAM 5.0; MW 28826 Da; 128 N-terminal amines)) was purchased from Sigma-Aldrich Chemicals (St. Louis, MO).
PAMAM dendrimers were supplied in methanol, which was removed using vacuum evaporation, and stored with phos- phate buffered saline (PBS) at 4°C. The activated dendrimers were prepared at a concentration of 2 mg/mL in a 50 °C water bath for the times indicated. pDNA (pSG5lacZ, 8 Kb), which encodes the lacZ gene for β-galactosidase, was driven by a SV40 promoter to assess gene expression. pSG5lacZ was amplified in Escherichia coli and purified using column chromatography (NucleoBond PC 2000; Macherey-Nagel GmbH & Co., Du¨ren, Germany). The purity of pSG5lacZ was established using ultraviolet (UV) spectroscopy (E260 nm/E280 nm ratio; range: 1.80-1.90). All other chemicals were of the highest grade available and were purchased locally.
Cell Culture. A human cervical cancer cell line (HeLa) was maintained in Dulbecco’s modified Eagle medium (Gibco DMEM; Invitrogen, Carlsbad, CA) supplemented with 10% heat-inactivated fetal bovine serum (Gibco FBS;
Invitrogen) and 100 U/mL of penicillin/100 µg/mL strepto- mycin (Sigma-Aldrich) in a humidified atmosphere of 5%
CO2at 37°C.
Cytotoxicity Assay. The activity of dehydrogenases (an indicator of cell viability) in the cells (treated or untreated with dendrimers, alone, or their DNA complexes) was simultaneously assessed. Negative control cells contained no PAMAM or their DNA complexes. The incubation condi- tions of cells were the same as the incubation conditions of the transfection assay described in the next section. To measure cell viability, 10 µL of a cell-counting kit solution, a tetrazolium salt that produces a highly water-soluble formazan dye upon biochemical reduction in the presence of an electron carrier (1-methoxy PMS) (Cell Counting Kit- 8; Dojindo Laboratories, Kumamoto, Japan), was added to a 100 µL culture medium and incubated for 4 h. The amount of yellow formazan dye generated by dehydrogenases in cells is directly proportional to the number of viable cells in a culture medium. The absorbance at 450 nm was obtained using an ELISA reader with a reference wavelength of 595 nm. Results are reported as the cell viability percentage (average optical density (OD)/average negative-control OD) ( standard deviation (SD).
Transfection Assay. Cells were seeded into 24-well cell- culture plates at a density of 3× 105cells/well and grown overnight (60% to 75% confluence). Immediately before transfection, the cells were rinsed with PBS and supple- mented with 1 mL of fresh DMEM per well. The pDNA (5 µg) and a range of concentrations of intact or activated PAMAM were each diluted into 500 µL of DMEM solution.
PAMAM dendrimers were added to the pDNA solution at
(5) Dufe`s, C.; Uchegbu, I. F.; Scha¨tzlein, A. G. Dendrimers in gene delivery. AdV. Drug DeliVery ReV. 2005, 57, 2177–2202.
(6) Dennig, J.; Duncan, E. Gene transfer into eukaryotic cells using activated polyamidoamine dendrimers. J. Biotechnol. 2002, 90, 339–347.
(7) Tang, M. X.; Redemann, C. T.; Szoka, F. C. In vitro gene delivery by degraded polyamidoamine dendrimers. Bioconjugate Chem.
1996, 7, 703–714.
(8) Navarro, G.; de ILarduya, C. T. Activated and non-activated PAMAM dendrimers for gene delivery in vitro and in vivo.
Nanomedicine 2009, 5, 287–297.
(9) Dufe`s, C.; Keith, W. N; Bilsland, A.; Proutski, I.; Uchegbu, I. F.;
Scha¨tzlein, A. G. Synthetic anticancer gene medicine exploits intrinsic antitumor activity of cationic vector to cure established tumors. Cancer Res. 2005, 65, 8079–8084.
(10) Chisholm, E. J.; Vassaux, G.; Martin-Duque, P.; Chevre, R.;
Lambert, O.; Pitard, B.; Merron, A.; Weeks, M.; Burnet, J.;
Peerlinck, I.; Dai, M. S.; Alusi, G.; Mather, S. J.; Bolton, K.;
Uchegbu, I. F.; Schatzlein, A. G.; Baril, P. Cancer-specific transgene expression mediated by systemic injection of nanopar- ticles. Cancer Res. 2009, 69, 2655–2662.
(11) Omidi, Y.; Hollins, A. J.; Drayton, R. M.; Akhtar, S. Polypro- pylenimine dendrimer-induced gene expression changes: the effect of complexation with DNA, dendrimer generation and cell type.
J. Drug Targeting 2005, 13, 431–443.
(12) Hollins, A. J.; Omidi, Y.; Benter, I. F.; Akhtar, S. Toxicogenomics of drug delivery systems: Exploiting delivery system-induced changes in target gene expression to enhance siRNA activity. J.
Drug Targeting 2007, 15, 83–88.
(13) Gras, R.; Almonacid, L.; Ortega, P.; Serramia, M. J.; Gomez, R.;
de la Mata, F. J.; Lopez-Fernandez, L. A.; Munoz-Fernandez, M. A. Changes in gene expression pattern of human primary macrophages induced by carbosilane dendrimer 2G-NN16. Pharm.
Res. 2009, 26, 577–586.
room temperature and allowed to sit for 15 min to allow dendrimer-DNA complexes to form. The cell medium was removed and the cells were then exposed to transfection mixtures for 4 h in the absence of serum, after which they were supplemented with 10% FBS and 1% antibiotics. After 24 h of incubation, β-galactosidase gene expression was analyzed using a β-Gal assay kit (Invitrogen) and a BCA protein assay reagent kit (Pierce, Rockford, IL). The trans- fection activity is expressed as nanomoles of o-nitrophenyl- β-D-galactopyranoside (ONPG)/min/mg protein.
Light Scattering and Zeta Potential Analysis. The particle size and zeta potential of dendriplexes in DNase/
RNase-free ultrapure water suspensions were measured using light scattering and capillary electrophoresis (Zetasizer 3000HS; Malvern Instruments Ltd., Malvern, Worcestershire, U.K.).
Purifying RNA, and Microarray Analysis. Under the same incubation conditions as in the transfection assay, total RNA was extracted from HeLa cells treated and untreated with intact and activated PAMAM alone or their dendri- plexes, using a reagent (Trizol; Invitrogen) and purified using a kit (RNeasy Mini Kit; Qiagen GmbH, Hilden, Germany).
To avoid the interaction between dendrimers and total RNA, the RNA pellets were washed in 2 M NaCl solution for 12 h.14 The purified RNA was quantified at an optical density (OD) of 260 nm using a spectrophotometer (ND- 1000; Nanodrop Technologies, Wilmington, DE) and qualitatively analyzed using a bioanalyzer (2100; Agilent Technologies, Inc., Santa Clara, CA). Microarray experi- ments were done following the manufacturer’s protocols.
Briefly, 0.5 µg of total RNA was amplified using a kit (Fluorescent Linear Amplification Kit; Agilent) and labeled with Cy3-CTP or Cy5-CTP (CyDye; PerkinElmer, Waltham, MA) during in Vitro transcription. RNA from cells treated with intact and activated PAMAM dendrimers or their dendriplexes was labeled using Cy5, and RNA from untreated cells was labeled using Cy3. Cy-labeled cRNA (0.825 µg) was fragmented to an average size of about 50-100 nucleotides (nt) using incubation with a fragmentation buffer (Agilent) at 60 °C for 30 min.
Correspondingly fragmented labeled cRNA was then pooled and hybridized to an oligo microarray kit (Whole Human Genome Microarray [4 × 44K] kit; Agilent) at 60 °C for 17 h. After they had been washed and dried with a nitrogen gun, the microarrays were scanned (microarray scanner; Agilent) at 535 nm for Cy3 and 625 nm for Cy5. Scanned images were analyzed (Feature Extraction Software 9.5.3; Agilent) using an image analysis and normalization software to quantify signal and background intensity for each feature. The data were substantially normalized using the rank-consistency-filter- ing LOWESS method. Genes whose expression increased
g2 times or decreased e0.5 time, and that had a p value
<0.05, were considered to be induced.
Real-Time Quantitative Reverse Transcriptase-Poly- merase Chain Reaction (qPCR). The same RNA isolated for the microarrays was used for qPCR. To prepare a cDNA pool from each RNA sample, total RNA (5 µg) was reverse- transcribed using reverse transcriptase (Moloney murine leukemia virus [M-MLV]; Promega, Madison, WI); the resulting samples were diluted 40 times by volume with nuclease-free water. Each cDNA pool was stored at -20
°C until it was subjected to additional real-time qPCR with specific oligonucleotide primer pairs from the Roche Uni- versal Probe Library (Roche Diagnostics Corporation, Roche Applied Science, Indianapolis, IN). Four genes (PHF5A, ARNTL2, CHD4, and P2RX7) were investigated using the following primer sequences: PHF5A, sense GGCACAC- CATATCGGAGAAA and antisense TCACAGTCAGTTC- CTCAAGCA; ARNTL2, sense TGGCACACTGTCCTCT- TGAA and antisense CACACCAATTCTCCCCATCT;
CHD4, sense TGTAAAGGAACAGCCCCAGT and anti- sense GCAGGAACCCACAACAGTTT; and P2RX7, sense CTGCCGTCCCAAATACAGTT and antisense GTGC- CAAAAACCAGGATGTC. Human PPIH (peptidylprolyl isomerase H (cyclophilin H)) was used as the reference gene.
The specificity of each primer pair was tested using rat common reference RNA (Stratagene, La Jolla, CA) as a template for real-time PCR, and then using a DNA 500 chip run on the Bioanalyzer 2100 to check the size of the PCR product. Primer pairs that yielded the predicted product at the predicted size, and no other primary or secondary products, were chosen for the real-time PCR reaction (LightCycler Instrument 1.5 using LightCycler FastStart DNA MasterPLUS SYBR Green I kit (Cat. 03 515 885 001);
Roche, Castle Hill, Australia). Briefly, 10 µL reactions contained 2 µL of 5× Master Mix, 2 µL of each 3.75 µM forward and reverse primer mixture, and 6 µL of cDNA (40:1 dilution). Each sample was run in triplicate. The real-time PCR program was 95°C for 10 min, 50 cycles at 95 °C for 10 s, 60°C for 15 s, and 72 °C for 10 s. At the end of the program, a melt curve analysis was done. For each real-time PCR run, the data were automatically analyzed by the system and an amplification plot was generated for each cDNA sample. From each of these plots, the system automatically calculated the CP value (crossing point; the turning point corresponds to the first maximum of the second derivative curve), which we infer to be the beginning of exponential amplification.
Statistical Analysis. Data were analyzed using one-way analysis of variance (ANOVA). Statistical significance was set at p< 0.05.
Results
Cytotoxicity Assay. To assess the cytotoxic effect of nonactivated PAMAM dendrimers and their dendriplexes, we incubated HeLa cells for 24 h with various concentrations of PAMAM dendrimers (10, 15, 30, and 50 µg/mL) and their
(14) Kuo, J. H.; Lin, Y. L. Remnant cationic dendrimers block RNA migration in electrophoresis after monophasic lysis. J. Biotechnol.
2007, 129, 383–390.
DNA (5 µg/mL) complexes (Figure 1). Cell viability was above 80% at the intact PAMAM dendrimer concentrations of 10 and 15 µg/mL but below 80% at 30 and 50 µg/mL.
Cell viability was above 80% until 50 µg/mL for intact dendriplexes (Figure 1). The negative charge of DNA neutralized the positive charge of the PAMAM dendrimers and increased cell viability. The intact PAMAM dendrimers were then activated for 8, 16, and 24 h, after which they were analyzed using cytotoxic assays (Figure 1). The cell viability of activated PAMAM dendrimers under all incuba- tion conditions was at best comparable and slightly more cytotoxic than that of their nonactivated counterparts.
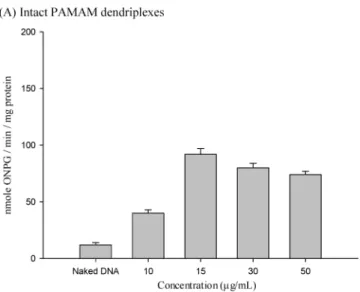
Transfection Assay. We examined the in Vitro gene expression of intact dendriplexes (5 µg/mL of DNA plus 10, 15, 30, or 50 µg/mL of nonactivated PAMAM) incubated for 24 h (Figure 2A). Gene expression using intact dendri- plexes was more efficient than using naked DNA alone.
Maximum transfection activity occurred at 15 µg/mL (Figure 2A). Taking into account transfection efficiency and cyto- toxicity, 15 µg/mL of intact PAMAM dendrimers were selected and heated for different times, after which they were combined in complexes with 5 µg/mL of DNA, and then transfection activity was assessed (Figure 2B). Heating intact PAMAM dendrimers for 16 h yielded the maximum levels of gene expression (cell viability ) 84 ( 3%), whereas transfection activity remained unchanged after 8 h of heating, and gene expression decreased after 24 h of heating (Figure 2B). Therefore, we chose 15 µg/mL PAMAM dendrimers (intact and activated for 16 h) and dendriplexes for microar- ray analysis.
Light Scattering and Zeta Potential Analysis. The mean diameter of 15 µg/mL intact PAMAM/5 µg/mL DNA dendriplexes was 177 ( 24 nm with a narrow distribution (polydispersity index (PI) ) 0.16; n ) 3). We found a smaller mean particle size (108 ( 16 nm; n ) 3) and slightly broader distribution (PI ) 0.21) for 15 µg/mL activated PAMAM/5 µg/mL DNA dendriplexes. The zeta potentials were com- parable between 15 µg/mL intact PAMAM/5 µg/mL DNA dendriplexes (12 ( 2 mV; n ) 3) and 15 µg/mL activated PAMAM (16 h heating)/5 µg/mL DNA dendriplexes (11 ( 2 mV; n ) 3).
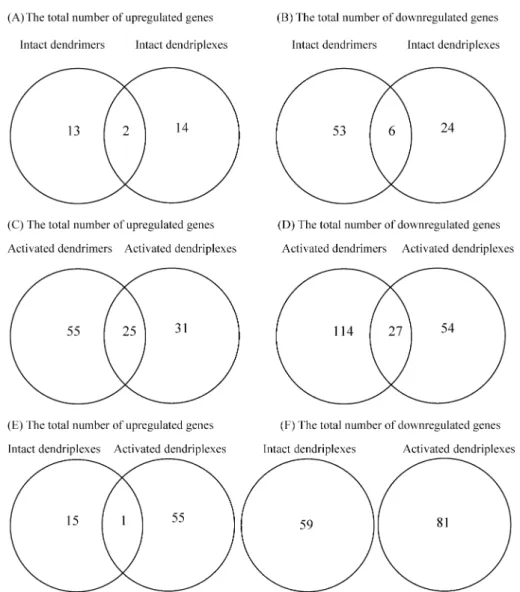
Global Gene Expression Profiles of HeLa Cells Treated with Intact and Activated Dendrimers Alone or with Their Dendriplexes. Activated PAMAM dendrimers alone and their dendriplexes produced more upregulated and downregulated genes than did intact dendrimers alone and their dendriplexes (Figure 3). Intact dendrimers and their dendriplexes produced similar numbers of upregulated genes, but intact dendrimers produced more downregulated genes Figure 1.Cytotoxicity assays of intact PAMAM dendrimers
and their dendriplexes and activated PAMAM dendrimers on HeLa cells by measuring generated dehydrogenases.
Negative control cells were grown without adding either intact dendrimers and their dendriplexes or activated dendrimers. Data are cell viability percentages (aver- age OD/average negative control OD) ( standard deviation (SD) (n ) 3).
Figure 2. Transfection of HeLa cells with intact dendriplexes (A) and activated PAMAM (15 µg/mL)/
DNA complexes (B). The data are means ( standard deviation (SD) (n ) 3).
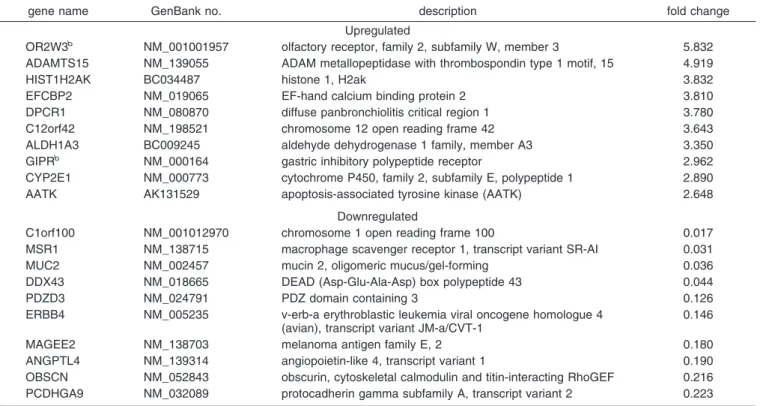
than did their dendriplexes (Figure 3A,B). The number of upregulated and downregulated genes was higher for acti- vated dendrimers than for activated dendriplexes (Figure 3C,D); in addition, there were more downregulated genes than upregulated genes. Moreover, some expressed genes were affected by being exposed to dendrimers as well as to dendriplexes (Figure 3A-D). Tables 1-4 provide a list of 10 most upregulated and downregulated genes in HeLa cells exposed to intact and activated dendrimers and their dend- riplexes. The GIPR and OR2W3 genes, which regulate membrane receptors, were both upregulated by intact PAM- AM dendrimers alone and their dendriplexes. In addition to the GIPR and OR2W3 genes, the intact PAMAM dendrimers alone upregulated 13 other genes, and the intact dendriplexes upregulated 14 genes. Both intact dendrimers and their dendriplexes downregulated 6 genes. More upregulated and downregulated genes were identified for activated dendrimers alone and their dendriplexes than for intact dendrimers and their dendriplexes. This indicated that activated dendrimers and their dendriplexes produced more profound changes than did intact dendrimers and their dendriplexes in transfection
efficiency and in gene upregulation and downregulation.
Also, the Venn diagram of activated vs nonactivated dend- riplexes demonstrated that the GIPR gene was the only upregulated gene affected by both activated and nonactivated dendriplexes, and that no downregulated genes were over- lapped by both activated and nonactivated dendriplexes (Figure 3E,F). This reflected that the differences in the structural architecture of the dendrimers induced almost totally different gene-expression profiles in cancer cells.
Figure 4 shows the percentage of principal molecular functions of these upregulated and downregulated annotated genes compared with the total number of annotated genes (analyzed using the Gene Ontology (GO) database). The 5 most principal molecular functions of regulated genes for intact and activated dendrimers and their dendriplexes were nucleic acid binding and transcription activity, metal-ion binding, enzyme activity, receptor activity, and protein binding. Activated dendrimers and their dendriplexes induced more molecular functions than did intact dendrimers and their dendriplexes.
Figure 3. A Venn diagram showing the number of regulated genes in HeLa cells treated with intact dendrimers and their dendriplexes (A, B), activated dendrimers and their dendriplexes (C, D), and activated and intact dendriplexes (E, F).
Confirming Microarray Results Using qPCR. To com- pare the gene expression results obtained from the microarray analysis with the results from qPCR, we tested four genessPHF5A, ARNTL2, CHD4, and P2RX7, responsible for nucleus gene regulation of transcription,
signal transduction, and DNA binding, and membrane gene regulation of ion transport, respectivelysfrom activated dendrimers and their dendriplexes (Figure 3). The qPCR were qualitatively consistent with those from the microar- ray (Figure 5).
Table 1. The 10 Most Upregulated and Downregulated Genes in Nonactivated Dendrimersa
gene name GenBank no. description fold change
Upregulated
OR2W3b NM_001001957 olfactory receptor, family 2, Subfamily W, member 3 5.350
TRIM49 NM_020358 tripartite motif-containing 49 4.834
FOLH1 NM_004476 folate hydrolase (prostate-specific membrane antigen) 1, transcript variant 1
4.747
ENST00000370419 ENST00000370419 regulating synaptic membrane exocytosis protein 1 (Rab3-interacting molecule 1) (RIM 1)
3.969
CHERP NM_006387 calcium homeostasis endoplasmic reticulum protein 3.060
GIPRb NM_000164 gastric inhibitory polypeptide receptor 3.031
ABCB1 NM_000927 ATP-binding cassette, subfamily B (MDR/TAP), member 1 2.781
RSU1 NM_012425 Ras suppressor protein 1, transcript variant 2.776
PTPRR NM_002849 protein tyrosine phosphatase, receptor type, R, transcript variant 1 2.659
C5orf23 NM_024563 chromosome 5 open reading frame 23 2.536
Downregulated
KCNJ10 NM_002241 potassium inwardly rectifying channel, subfamily J, member 10 0.010
PTGFR NM_001039585 prostaglandin F receptor (FP), transcript variant 2 0.028
SLC8A1 X91815 Na+/Ca2+exchanger isoform 0.046
USP29 NM_020903 Ubiquitin specific peptidase 29 0.068
PDE11A NM_016953 phosphodiesterase 11A, transcript variant 4 0.070
UGT8b NM_003360 UDP glycosyltransferase 8 (UDP-galactose ceramide galactosyltransferase)
0.076
ART4 NM_021071 ADP-ribosyltransferase 4 (Dombrock blood group) 0.077
RGS9BP NM_207391 regulator of G protein signaling 9 binding protein 0.085
MATN1 ENST00000373765 cartilage matrix protein precursor (Matrilin-1) 0.128
CXYorf3 BC028151 Homo sapiens chromosome X and Y open reading frame 3 0.137
aA full list is available in the Supporting Information.bGenes affected by both nonactivated dendrimers and dendriplexes.
Table 2. The 10 Most Upregulated and Downregulated Genes in Nonactivated Dendriplexesa
gene name GenBank no. description fold change
Upregulated
OR2W3b NM_001001957 olfactory receptor, family 2, subfamily W, member 3 5.832 ADAMTS15 NM_139055 ADAM metallopeptidase with thrombospondin type 1 motif, 15 4.919
HIST1H2AK BC034487 histone 1, H2ak 3.832
EFCBP2 NM_019065 EF-hand calcium binding protein 2 3.810
DPCR1 NM_080870 diffuse panbronchiolitis critical region 1 3.780
C12orf42 NM_198521 chromosome 12 open reading frame 42 3.643
ALDH1A3 BC009245 aldehyde dehydrogenase 1 family, member A3 3.350
GIPRb NM_000164 gastric inhibitory polypeptide receptor 2.962
CYP2E1 NM_000773 cytochrome P450, family 2, subfamily E, polypeptide 1 2.890
AATK AK131529 apoptosis-associated tyrosine kinase (AATK) 2.648
Downregulated
C1orf100 NM_001012970 chromosome 1 open reading frame 100 0.017
MSR1 NM_138715 macrophage scavenger receptor 1, transcript variant SR-AI 0.031
MUC2 NM_002457 mucin 2, oligomeric mucus/gel-forming 0.036
DDX43 NM_018665 DEAD (Asp-Glu-Ala-Asp) box polypeptide 43 0.044
PDZD3 NM_024791 PDZ domain containing 3 0.126
ERBB4 NM_005235 v-erb-a erythroblastic leukemia viral oncogene homologue 4 (avian), transcript variant JM-a/CVT-1
0.146
MAGEE2 NM_138703 melanoma antigen family E, 2 0.180
ANGPTL4 NM_139314 angiopoietin-like 4, transcript variant 1 0.190
OBSCN NM_052843 obscurin, cytoskeletal calmodulin and titin-interacting RhoGEF 0.216
PCDHGA9 NM_032089 protocadherin gamma subfamily A, transcript variant 2 0.223
aA full list is available in the Supporting Information.bGenes affected by both nonactivated dendrimers and dendriplexes.
Discussion
Using dendrimers in cancer therapy as nonviral vectors for gene delivery has been successful in ViVo;9,10they not only delivered genes to tumors but also had a synergistic
therapeutic effect.9,15 However, little is known about the molecular mechanism that dendrimers and their dendriplexes
(15) Akhtor, S. Beyond delivery. Gene Ther. 2006, 13, 739–740. Table 3. The 10 Most Upregulated and Downregulated Genes in Activated Dendrimersa
gene name GenBank no. description fold change
Upregulated
ENST00000379879 ENST00000379879 immunoglobulin heavy chain variable region (Fragment) 8.937
ZSCAN4 NM_152677 zinc finger and SCAN domain containing 4 8.871
NPTX1 NM_002522 neuronal pentraxin I 7.116
MYH8 NM_002472 myosin, heavy chain 8, skeletal muscle, perinatal 5.371
HMGCS1 NM_002130 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 1 (soluble) 4.988
IGF1 NM_000618 insulin-like growth factor 1 (somatomedin C) 4.712
ENST00000374458 ENST00000374458 gametogenetin-binding protein 1 4.681
TMEM16C NM_031418 transmembrane protein 16C 4.618
ANKRD19 NM_001010925 ankyrin repeat domain 19 4.522
SLC4A3 NM_005070 solute carrier family 4, anion exchanger, member 3, transcript variant 1
4.432
Downregulated
ENST00000319098 ENST00000319098 prosaposin-like protein 1 0.147
TEP1 NM_007110 telomerase-associated protein 1 0.154
ARNTL2 AF256215 cycle-like factor CLIF 0.171
C1QTNF5 NM_015645 C1q and tumor necrosis factor related protein 5 0.186
ZNF652 NM_014897 zinc finger protein 652 0.186
RNF125 NM_017831 ring finger protein 125 0.188
TEAD4 NM_003213 TEA domain family member 4, transcript variant 1 0.195
LRRC4 NM_022143 leucine rich repeat containing 4 0.205
DIRAS2 NM_017594 DIRAS family, GTP-binding RAS-like 2 0.205
ARHGAP26 NM_015071 Rho GTPase activating protein 26 0.206
aA full list is available in the Supporting Information.
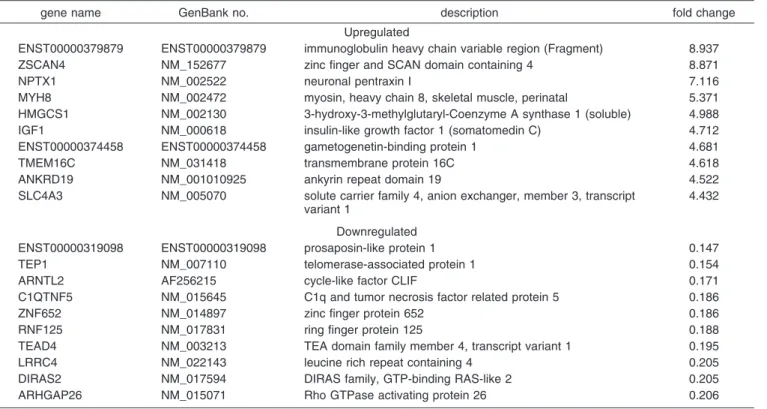
Table 4. The 10 Most Upregulated and Downregulated Genes in Activated Dendriplexesa
gene name GenBank no. description fold change
Upregulated
HS6ST3 NM_153456 heparan sulfate 6-O-sulfotransferase 3 11.019
GATS NM_178831 opposite strand transcription unit to STAG3 8.069
KLF17 NM_173484 Kruppel-like factor 17 7.890
VNN3 NM_078625 vanin 3 (VNN3), transcript variant 2 5.334
HMGCS1b NM_002130 3-hydroxy-3-methylglutaryl-Coenzyme A synthase 1 (soluble) 5.118 B4GALT5 NM_004776 UDP-Gal:betaGlcNAc beta 1,4- galactosyltransferase, polypeptide 5 3.774
EGR1 NM_001964 early growth response 1 3.764
PLA2G2E NM_014589 phospholipase A2, group IIE 3.556
UBQLN1b NM_013438 ubiquilin 1, transcript variant 1 3.208
SYNPO ENST00000307662 synaptopodin 3.019
Downregulated
IL31 NM_001014336 interleukin 31 0.064
MED18 NM_017638 mediator of RNA polymerase II transcription, subunit 18 homologue (S. cerevisiae)
0.075
TEP1 NM_007110 telomerase-associated protein 1 0.125
DNAJC12 NM_021800 DnaJ (Hsp40) homologue, subfamily C, member 12, transcript variant 1
0.150
ARNTL2 AF256215 cycle-like factor CLIF 0.184
RNF125 NM_017831 ring finger protein 125 0.196
ZNF652 NM_014897 zinc finger protein 652 0.204
HKDC1 NM_025130 hexokinase domain containing 1 0.213
C1QTNF5 NM_015645 C1q and tumor necrosis factor related protein 5 0.228
CD74 NM_001025158 CD74 molecule, major histocompatibility complex, class II invariant chain, transcript variant 3
0.243
aA full list is available in the Supporting Information.bGenes affected by both activated dendrimers and dendriplexes.
use in cancer cells.9One microarray analysis11showed that, in human A431 cells, cationic PPI dendrimers induced changes in the expression of a variety of gene ontologies, including those involved in defense responses, cell prolifera-
tion, and apoptosis, which might have been related to their antitumor effects. Furthermore, the patterns of global gene expression were dramatically changed when PPI dendrimers were bound in complexes with plasmid DNA. It is, therefore, Figure 4. Graphical categories by principal molecular functions of regulated genes in HeLa cells treated with intact dendrimers (A, B), intact dendriplexes (C, D), activated dendrimers (E, F), and activated dendriplexes (G, H). The percentage of each category compared to the total number of annotated genes is shown.
important to understand the influence of the molecular architecture of dendrimers on gene expression profiling.
In this microarray study, we investigated 44,000 high- density genes instead of 161 low-density genes.11We found that the extent of gene changes was dependent on the molecular architecture of intact and activated dendrimers,11 and that the patterns of gene expression were different between dendrimers and their dendriplexes. However, the type of genes significantly upregulated and downregulated is different from those in the only other study on this topic.11 These differences may depend upon on the type of dendrimer (PPI vs PAMAM), cell (A431 vs HeLa), or gene chips (low- density vs high-density) used. Fewer upregulated and down- regulated genes were involved in apoptosis and immune responses than in the previous study.11
The activated PAMAM dendrimers and their dendriplexes used in our study showed not only higher transfection efficiencies but also more significant upregulated and down- regulated genes than intact dendrimers and their dendriplexes.
Our transfection results were consistent with those of other studies7,8reporting that a highly flexible activated structure was responsible for this effect.7,8We showed that dendrimer activation induced more gene expression changes in cancer cells. Genes that regulate transcription and proliferation have also been identified in macrophages treated with carbosilane dendrimers.13
In our study, the criteria for selecting genes measured using qPCR were based both on significant expression changes and on their pharmacological functions that are closely related to antitumor actions. Of the four genes selected, three (PHF5A, ARNTL2, and CHD4) are in- volved in survival signaling and the proliferation of cancer cells, and one (P2RX7) is related to signaling between immune cells and target cells. PHF5A is a highly
conserved chromatin-associated protein of various species and required for cell viability.16 Our results indicate that PHF5A upregulation is the survival signal in HeLa cells treated with activated dendrimers and their dendriplexes.
ARNTL2 downregulation increased cell proliferation in hepatocellular carcinoma.17 In contrast, downregulation of CHD4, involved in the regulation of the early growth response, inhibits cell proliferation in various cells.18,19 P2RX7 is a membrane receptor of ligand-gated cation channels and associated with diverse physiological roles including cancer, pain, inflammation, and unusual re- sponses to infection.20,21In the presence of ATP and when divalent cation levels are low, P2RX7 converts to an intrinsic pore that initiates binding to ATP, which allows ions to flow.20 P2X7 receptor is involved in signaling between macrophages or other cells involved in the immune response and target cells, and may cause cell swelling and cell death after prolonged activation.20The consequences of downregulating P2X7 in HeLa cells have not been reported before, and the detailed molecular mechanism remains to be clarified. In addition, some of the gene expression changes from qPCR may have been due to the presence of the transgene rather than just the delivery process. Previous studies9,10 reported that the dendrimers not only had delivered the encoded gene for tumor necrosis factor-alpha but also had an intrinsic antitumor effect in tumor-bearing mice, and that these data demonstrated the possibilities for enhanced cancer gene therapy using dendrimers as the delivery vectors. However, the exact mechanism of antitumor activity from dendrimers remains largely unknown. Our study of these four regulated genes explained that the engagement of cell survival and cell death mechanisms, as well as immune stimulation by activated PAMAM dendrimers and dend- riplexes, may be important for cancer gene therapy because of their antitumor effects. Taken together, our results suggest that activated dendrimers and dendriplexes have diverse roles in controlling gene expression changes in HeLa cells.
(16) Trappe, R.; Ahmed, M.; Glaser, B.; Vogel, C.; Tascou, S.;
Burfeind, P.; Engela, W. Identification and characterization of a novel murine multigene family containing a PHD-finger-like motif.
Biochem. Biophys. Res. Commun. 2002, 293, 816–826.
(17) Yeh, C. T.; Lu, S. C.; Tseng, I. C.; Lai, H. Y.; Tsao, M. L.; Huang, S. F.; Liaw, Y. F. Antisense overexpression of BMAL2 enhances cell proliferation. Oncogene 2003, 22, 5306–5314.
(18) Osman, A. M.; van Kol, S.; Peijnenburg, A.; Blokland, M.;
Pennings, J. L.; Kleinjans, J. C.; van Loveren, H. Proteomic analysis of mouse thymoma EL4 cells treated with bis(tri-n- butyltin)oxide (TBTO). J. Immunotoxicol. 2009, 6, 174–183.
(19) Srinivasan, R.; Mager, G. M.; Ward, R. M.; Mayer, J.; Svaren, J.
NAB2 represses transcription by interacting with the CHD4 subunit of the nucleosome remodeling and deacetylase (NuRD) complex. J. Biol. Chem. 2006, 281, 15129–15137.
(20) Ralevic, V.; Burnstock, G. Receptors for purines and pyrimidines.
Pharmacol. ReV. 1998, 50, 413–492.
(21) Khakh, B. S.; North, R. A. P2X receptors as cell-surface ATP sensors in health and disease. Nature 2006, 442, 527–532. Figure 5.Confirmation of microarray results with real-time
quantitative RT-PCR (qPCR). Relative expression levels of 4 genes (PHF5A, ARNTL2, CHD4, and P2RX7) in HeLa cells incubated with activated dendrimers and activated dendriplexes are shown. Data are means ( standard devia- tion (SD) (n ) 3).
Using dendrimers in ViVo has significant potential in gene-delivery cancer therapy. Two early in ViVo studies22,23 reported that activated PAMAM dendrimer-mediated angiostatin and metalloproteinase (TMP)-2 inhibited tumor growth and angiogenesis, and that gene delivery of Fas ligand by activated PAMAM dendrimers induced signifi- cant growth suppression and apoptotic tumor cell death in prostate cancer. Interestingly, like polypropylenimine dendrimers, activated PAMAM dendrimers alone also delayed tumor growth, but only polypropylenimine den- driplexes lead to a significant regression of tumors.9Our results from exploring the gene-expression profiles of dendrimers and dendriplexes might allow a more detailed insight for understanding their molecular mechanisms in cancer cells. Our results showed that activated dendrimers and dendriplexes induced marked global gene-expression changes in cancer cells. We found that genes involved in cell proliferation and immune responses were altered by activated dendrimers and dendriplexes, a finding consistent with their direct antitumor effect. Besides that, genes involved in various cellular processes, such as nucleic acid binding and transcription activity, metal-ion binding, enzyme activity, receptor activity, and protein binding, were also altered. Furthermore, the extent of gene changes was different between activated dendrimers and activated dendriplexes, which explains their variable antitumor effects in cancer cells. Our findings suggested that a successful vector for cancer gene therapy might be designed to have desirable molecular functions, such as an antitumor activity, optimal transfection efficiency, and minimal cytotoxicity.
Finally, the transfection efficiency results of nonviral gene delivery systems depend upon overcoming extra- and intra- cellular barriers involved in gene delivery. Once the gene delivery systems are at the targeted tumor cells, there are still several intracellular barriers: internalization, endosomal escape, cytoplasm trafficking, nuclear translocation, and transcription. We found that activated and nonactivated dendriplexes principally affected genes with the molecular functions of nucleic acid binding and transcription activity, metal-ion binding, enzyme activity, receptor activity, and protein binding. We hypothesize that the genes involved in
nucleic acid binding and transcription activity are a major factor that directly leads to the transfection efficiency of dendriplexes. Because we used in our study the same plasmid that was used in previous studies,9,10,22,23we conclude that dendrimer architecture is important for regulating transcrip- tion processes. Nonviral vectors compact DNA so efficiently that transcription processes are hindered.24 It has been reported25that PAMAM dendrimers inhibited the initiation of transcription in Vitro but did not affect the elongation of the RNA transcript. However, activated dendrimers are reported7 to undergo DNA decondensation after swelling, to be more flexible structures than intact dendrimers, and, therefore, to increase transfection efficiency. We found that activated dendriplexes have more regulated genes involved in transcription processes than intact ones, and that this may assist the activities of transcription factors and reach the ultimate goal of transgene expression.
Conclusion
In summary, our results illustrate the importance of the molecular architecture of the dendrimers on the effect of global gene expression profiles in HeLa cells. Dendrimers and dendriplexes induced gene expression changes; some of the dendrimer-induced expression changes were the same as dendriplex-induced expression changes. Activating den- drimers led to higher transfection efficiencies and induced more gene expression changes in cancer cells. Dendrimers and dendriplexes principally affect genes via the molecular functions of nucleic acid binding and transcription activity, metal-ion binding, enzyme activity, receptor activity, and protein binding. We hypothesize that a successful vector for cancer gene therapy may be designed to have desirable molecular functions, such as antitumor activity, optimal transfection efficiency, and minimal cytotoxicity.
Acknowledgment. This work was supported by Grant NSC 98-2221-E-041-002 from the National Science Council, Taiwan.
Supporting Information Available:Tables of intact and activated dendrimers and dendriplexes. This material is available free of charge via the Internet at http://pubs.acs.org.
MP900303S (22) Nakanishi, H.; Mazda, O.; Satoh, E.; Asada, H.; Morioka, H.;
Kishida, T. Nonviral genetic transfer of Fas ligand induced significant growth suppression and apoptotic tumor cell death in prostate can in ViVo. Gene Ther. 2003, 10, 434–442.
(23) Vincent, L.; Varet, J.; Pille, J. Y.; Bompais, H.; Opolon, P.;
Makshimenko, A. Efficacy of dendrimer-mediated Angiostatin and TMP-2 gene delivery on inhibition of tumor growth and angio- genesis: In Vitro and in ViVo studies. Int. J. Cancer 2003, 105, 419–429.
(24) Douglas, K. L. Toward development of artificial viruses for gene therapy: A comparative evaluation of viral and non-viral trans- fection. Biotechnol. Prog. 2008, 24, 871–883.
(25) Bielinska, U.; Kukowska-Latallo, F.; Baker, J. R. The interaction of plasmid DNA with polyamidoamine dendrimers: mechanism of complex formation and analysis of alterations induced in nuclease sensitivity and transcriptional activity of the complexed DNA. Biochim. Biophys. Acta 1997, 1353, 180–190.
99 年 7 月 20 日 報告人姓名 郭榮華
嘉南藥理科技大學藥物科技 研究所
時間 會議地點
07/10/2010-07/14/2010 Oregon Convention Center, Portland, Oregon, USA
會議名稱
(中文) 37 屆控制釋放協會年會暨展覽會
(英文) 37th
Annual Meeting & Exposition of the Controlled Release Society
發表論文 題目
(中文) 以細胞穿透性胜肽 Octaarginine 處理後人類免疫細胞株 U-937 之基因表現
(英文) Global Gene Expression Profiles in U-937 Human Macrophages Treated with
Octaarginine
一、參加會議經過
本屆 2010 年的控制釋放協會年會暨展覽會 (37th
Annual Meeting & Exposition of the Controlled Release Society)為期五天(07/10/2010-07/14/2010),在美國波特蘭奧勒岡會議中心舉行。此次會議
為藥學及控制釋放領域中最為重要的會議之一,今年有超過一千位的相關研究領域學業界參與這 場盛會。來自全世界各大製藥公司的研發人員都聚集於此,一方面發表最新的研究成果,另一方 面可以了解未來控制釋放與創新應用方向。本次 plenary session 所邀請的是來自 Harvard University 之 Prof. R. Kucherlapati 討論 personalized medicines、NovaBay Pharmaceuticals 之 R. Najafi 以業 界的角度討論如何在研究、製程、財務上能創新發展、University of California-San Francisco 之 Prof.F. Szoka 討論 Polymer architecture in tumor drug delivery、Sojo University (Japan)之 Prof. H. Maeda
討論 Enhanced Permeability and Retention Effect 之發展、University of Texas-Houston 之 Prof. M.Ferrari 討論 Nanotechnologies for personalized medicines、University of Texas-Austin 之 Prof. N.
Peppas 討論 Responsive Intelligence Delivery,因此對於學界在研究成果如何轉換成兼具實用與價
值的產品,都可以在這個會議中獲益無窮。而本次會議涵蓋的主題非常廣泛及實用,如Theranostics、New chemistriesfor drug delivery、Non-Parenteral delivery of biologics、Novel materials
& release systems、Opthalmic medicines、 prediction & application of IVIVC for veterinary species、
Transdermal drug delivery、Biomarkers、Nanoparticles & fiber for controlled release systems、Peptide
& protein medicines、Polymer therapeutics、pulmonary drug delivery、Translational medicine、DNA delivery、Encapsulation for environmental protection、Medical devices、siRNA/Micro RNA、Oral delivery、PEGylated technologies、Vaccines、Encapsulation of cell & microorganisms、Intracellular trafficking、Liposomes、Tumor targeting、Environmental friendly and biodegradable controlled release systems、Biomedical photonics、Stem cell、Nanomedicines、Hydrogel、Biomaterials 、Biomedical imaging、Vaginal drug delivery、Tissue Engineering 等。
二、與會心得
這次我報告的壁報論文,被大會安排在第三天的 Peptide & protein medicines 壁報論文場次中張貼 並與其他有興趣的研究學者一同討論,在壁報張貼期間許多相關領域的專家都前來參觀壁報,也 提出許多有用的意見及討論,使我能知道自己研究的盲點何在及本實驗在生技製藥界的價值及重 要性,因此受益匪淺。此外在會議進行期間也有許多出版社、廠商設攤展示,進行一連串示範及 演講活動,對於往後在研究中所使用相關材料及儀器亦十分有所幫助。
三、攜回資料名稱及內容 大會相關論文手冊、廠商 DM
Jung-hua Kuo, Meng-jie Liou, and Hsueh-chen Chiu
1 Graduate Institute of Pharmaceutical Science, Chia Nan University of Pharmacy and Science, 60 Erh-Jen Road, Sec. 1, Jen-Te, Tainan 717, Taiwan
[email protected] ABSTRACT SUMMARY
Cell penetrating peptides such as octaarginine (R8) have been widely used as intracellular delivery vectors to import biologically active membrane-impermeable molecules. However, before using these peptides clinically, human immune responses to them must be fully understood. Because macrophages are important for immune responses, we evaluated genome-wide profiling of gene expression after U-937 cells had been incubated with 10 µM R8 for 0.5 h and 4 h. Microarray analysis identified 4386 genes time-dependently regulated; the majority of these genes were upregulated for each time point.
Thirty-five upregulated genes responded to the stimuli with immune functions, and, using real-time quantitative reverse transcriptase-polymerase chain reaction analysis, five genes —FOS, OSM, C1R, TNF, IL1R1—were confirmed. The results may offer a variety of valuable insights into the molecular mechanisms of R8-induced immune responses in macrophages.
INTRODUCTION
Interest in using cell penetrating peptides (CPPs) or protein transduction domains as carriers for intracellular delivery is increasing. 1-3 Of the cell penetrating peptides used, octaarginine (R8), a member of the synthetic peptide family of arginine-rich peptides, carries various cargos for intracellular delivery. 4, 5 Arginine-rich peptides share efficient cellular uptake and an abundance of basic arginine amino acids with the most notable cell penetrating peptide, human immunodeficiency virus (HIV)-1
TAT-derived peptide (48-60).
Despite the highly efficient intracellular delivery of arginine-rich peptides, their immune responses when injected into the human body must be fully understood because they are derived primarily from non-human proteins, and very few studies have focused on the activity of immune cells after the uptake of these peptides.
Immune cells, especially macrophages, are crucial for
directing the host's foreign-body reactions and producing various proinflammatory mediators. Most studies, however, have been limited to functional gene expression profiles in cellular responses induced by arginine-rich peptides.
Therefore, we used a whole-genome analysis by
microarray to explore global gene expression profiles from U-937 human macrophages treated with R8.
EXPERIMENTAL METHODS
Total RNA was extracted using a reagent (Trizol;
Invitrogen Corp., USA) and then a kit (RNeasy Mini;
Qiagen GmbH, Germany). 0.5 µg of total RNA was amplified using a kit (Fluorescent Linear Amplification Kit;
Agilent) and labeled with Cy3-CTP (CyDye, Perkin-Elmer, USA) during the in vitro transcription process. Two micrograms of Cy-labeled cRNA was fragmented to an average size of about 50-100 nucleotides by incubating it with fragmentation buffer at 60°C for 30 min.
Correspondingly fragmented labeled cRNA was then hybridized to a 4 × 44-k microarray (Whole Human Genome Oligo Microarray Kit; Agilent) at 60°C for 17 h, and then scanned at 535 nm (Microarray Scanner; Agilent) at 535 nm. The scanned images were analyzed using commercial software (Feature Extraction 9.5.3; Agilent), as were the microarray data (GeneSpring GX 7.3.1;
Agilent). Chip intensities of six microarray data from three duplicate time points were normalized to equal signal distribution using per-chip normalization at the 75th percentile. Per-gene normalization to time point 0 h was also used. The resulting 28,060 genes were selected for differential expression; 4,386 differentially expressed genes were selected using 1.5-fold changes at one of three time points and the Significance Analysis of Microarray (SAM) method with a false-discovery rate (FDR) threshold of 0.05.
The same RNA isolated for the microarray was used for real-time quantitative reverse transcriptase-polymerase chain reaction analysis (RT-PCR). Real-time PCR
Instrument 1.5; Roche Diagnostics Australia Pty. Ltd., Australia) using a kit (LightCycler FastStart DNA MasterPlus SYBR Green I, Cat. 03 515 885 001; Roche).
Each sample was run in triplicate. The real-time PCR program steps were 95°C for 10 min, 50 cycles of 95°C for 10 s, 60°C for 15 s, and 72°C for 10 s. Five genes (FOS, OSM, C1R, TNF, IL1R1) were investigated using the following primer sequences: FOS sense CTACCACTCACCCGCAGACT and antisense
AGGTCCGTGCAGAAGTCCT; OSM sense
GTGGATGAGAGGAACCATCG and antisense
GTAGCAGAGGGGAACAGGTTT; CIR sense
TGCTCAACTACGTGGACTGG and antisense GATTCGAACCTAGTGAATTCTGG; TNF sense CAGCCTCTTCTCCCCTTCGAT and antisense
GCCAGAGGGCTGATTAGAGA; IL1R1 sense
ATTGTGCTTTGGTACAGGGATTCC and antisense ATGCGTCATAGGTCTTTCCATCTG. Human TBP (TATA box binding protein) was used as the reference gene.
RESULTS AND DISCUSSION
We used 1.5-fold changes at one of three time points (0 h, 0.5 h, and 4 h) to select 4386 differentially expressed genes from 28,060 identified genes; cluster analysis was then used for additional analysis. We found 11 clusters
with significant gene-expression changes. The majority of these genes with significant gene expression changes at each time point (0.5 h and 4 h) were upregulated (Table 1); 35 upregulated genes responded to the stimuli with immune functions in all 11 clusters and were also selected (Table 2). These genes included mostly cytokines, cytokine binding proteins, cytokine receptors, receptors, histocompatibility molecules, chemokines, complement components, CD8 antigens, pore-forming
compare the gene expression results obtained from the microarray with the results from RT-PCR, five genes (FOS, OSM, C1R, TNF, IL1R1) at each time point (Table 2) were tested. We found that the regulation patterns of the five genes, as measured by RT-PCR, were consistent with those from the microarray.
CONCLUSION
R8 induced various changes in gene expression in macrophages after it had bound with and was internalized by them. Our study may provide helpful information for the molecular mechanisms of R8-induced immune responses in macrophages
REFERENCES
1. Mae, M.; Langel, U. Curr. Opin. Pharmacol. 2006, 6, 509-514.
2. Vives, E. J. Control. Release. 2005, 109, 77-85.
3. Gupta, B.; Levchenko, T. S.; Torchilin V. P. Adv. Drug Deliv. Rev. 2005, 57, 637-651.
4. Futaki S. Biopolymers. 2006, 84, 241-249.
5. Futaki S. Adv. Drug Deliv. Rev. 2005, 57, 547-558.
ACKNOWLDEGEMENTS
This work was supported by grant NSC 97-2221-E-041-005 from the National Science Council, Taiwan.
Table 1. The number of genes with significant gene expression changes (fold change >1.5 )
Number of up-regulated genes
Number of down-regulated genes
0.5h 1276 25
4h 3967 29
Fold change GenBank Synonym 0.5 h 4h
NM_005252 FOS 41.821 1.964
NM_000594 TNF 6.918 1.965
NM_020530 OSM 2.901 1.439
NM_002983 CCL3 3.414 2.233
NM_002190 IL17 1.950 2.221
NM_000912 OPRK1 1.556 2.019
NM_001185 AZGP1 1.262 1.997
1C7 precursor 1.570 2.077
NM_003811 TNFSF9 1.918 2.313
BX640624 MGC27165 1.251 2.064
NM_002000 FCAR 1.336 2.291
NM_173044 IL18BP 1.654 2.132
NM_000594 TNF 2.752 1.920
NM_001733 C1R 2.053 2.223
NM_001242 TNFRSF7 1.971 2.044
NM_133280 FCAR 2.880 1.650
NM_003485 GPR68 1.129 2.070
BC034142 GKV1-5 1.865 3.592
NM_005515 HLXB9 1.546 2.073
cDNA FLJ39978 fis
2.761 3.294
NM_139011 HFE 2.636 2.031
NM_002258 KLRB1 1.349 2.008
NM_005118 TNFSF15 1.695 2.393
NM_001622 AHSG 1.762 2.123
NM_000877 IL1R1 2.065 1.456
NM_001768 CD8A 1.591 2.238
NM_001712 CEACAM1 1.953 3.026
NM_182549 HLA-DQB2 1.299 2.516
NM_000715 C4BPA 1.199 2.431
NM_006274 CCL19 1.696 2.319
NM_012400 PLA2G2D 1.550 2.308
NM_153758 IL19 1.759 1.995
NM_002121 HLA-DPB1 1.642 2.056
NM_005508 CCR4 1.918 2.064
J00231 IGHG3 1.334 2.195
NM_006610 MASP2 1.095 2.117
NM_130441 CLEC4C 1.248 2.010
NM_005041 PRF1 1.108 2.086
計畫主持人:郭榮華 計畫編號:98-2221-E-041-002- 計畫名稱:多功能性樹枝型高分子基因傳送系統之設計與開發
量化
成果項目 實際已達成
數(被接受 或已發表)
預期總達成 數(含實際已
達成數)
本計畫實 際貢獻百
分比
單位
備 註 ( 質 化 說 明:如 數 個 計 畫 共 同 成 果、成 果 列 為 該 期 刊 之 封 面 故 事 ...
等)
期刊論文 0 0 100%
研究報告/技術報告 0 0 100%
研討會論文 1 1 100%
論文著作 篇
專書 0 0 100%
申請中件數 0 0 100%
專利 已獲得件數 0 0 100% 件
件數 0 0 100% 件
技術移轉
權利金 0 0 100% 千元
碩士生 0 0 100%
博士生 0 0 100%
博士後研究員 0 0 100%
國內
參與計畫人力
(本國籍)
專任助理 0 0 100%
人次
期刊論文 1 1 100%
研究報告/技術報告 0 0 100%
研討會論文 0 0 100%
論文著作 篇
專書 0 0 100% 章/本
申請中件數 0 0 100%
專利 已獲得件數 0 0 100% 件
件數 0 0 100% 件
技術移轉
權利金 0 0 100% 千元
碩士生 2 2 100%
博士生 0 0 100%
博士後研究員 0 0 100%
國外
參與計畫人力
(外國籍)
專任助理 0 0 100%
人次
其他成果 (無法以量化表達之成 果如辦理學術活動、獲 得獎項、重要國際合 作、研究成果國際影響 力及其他協助產業技 術發展之具體效益事 項等,請以文字敘述填 列。)
成果項目 量化 名稱或內容性質簡述
測驗工具(含質性與量性) 0
課程/模組 0
電腦及網路系統或工具 0
教材 0
舉辦之活動/競賽 0
研討會/工作坊 0
電子報、網站 0
科 教 處 計 畫 加 填 項
目 計畫成果推廣之參與(閱聽)人數 0
請就研究內容與原計畫相符程度、達成預期目標情況、研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)、是否適 合在學術期刊發表或申請專利、主要發現或其他有關價值等,作一綜合評估。
1. 請就研究內容與原計畫相符程度、達成預期目標情況作一綜合評估
■達成目標
□未達成目標(請說明,以 100 字為限)
□實驗失敗
□因故實驗中斷
□其他原因 說明:
2. 研究成果在學術期刊發表或申請專利等情形:
論文:■已發表 □未發表之文稿 □撰寫中 □無 專利:□已獲得 □申請中 ■無
技轉:□已技轉 □洽談中 ■無 其他:(以 100 字為限)
3. 請依學術成就、技術創新、社會影響等方面,評估研究成果之學術或應用價 值(簡要敘述成果所代表之意義、價值、影響或進一步發展之可能性)(以 500 字為限)
瞳蚚皉闚準瓷馮俶徭闚酕價秪冞蚚黺骨痌笥?準都撿衄嶱追?薯﹝
皉闚價秪冞?闚眳汜昜魂俶桶政鷂煦赽磐?洘洘眈燊﹝瞳蚚⺌ 樓?皉闚窒煦蔥 賤眳魂趙庨最褫湮盟枑汔皉闚眳滖虴彆﹝ 奧ㄛ皉闚婓骨痌?婉眳煦赽滖帤坋煦 ラ奠﹝扂?旃噶魂趙摯帤魂趙 poly(amidoamine)(PAMAM)皉闚摯鷂 DNA 磐磁眳恚磁闚
黺?赽蟊骨 HeLa?婉眳價秪桶政耀宒?趙﹝扂?瞳蚚撈講趙毀滖?擄磁繪?毀復庲賸侐 意價秪桶政(PHF5A﹜ARNTL2﹜CHD4﹜P2RX7)掩皉闚垀荌?﹝帤魂趙鷂魂趙皉闚摯鷂 DNA 恚磁闚眳價秪蜊?耀宒衄湮窒爺笭葡﹝魂趙眳皉闚壺賸枑滖虴薹﹜砫謞俋竘追價 秪桶政眳蜊?﹝皉闚摯鷂 DNA 恚磁闚翋猁荌?狟蹈價秪煦赽髡夔ㄩ瞄呫磐磁摯滖?魂 俶﹜踢褩赽磐磁﹜談匼魂俶﹜諉忳闚魂俶﹜摯粥啞斮磐磁ㄛ扂?腔旃噶皉闚磐?眳 蜊?⺼睡荌?骨?婉眳價秪桶政蜊?ㄛ瞳蚚黺骨痌笥?奻枑鼎賸旮⻌眳罠﹝