國立臺灣大學生命科學院植物科學研究所 博士論文
Institute of Plant Biology College of Life Science National Taiwan University
Doctoral Dissertation
阿拉伯芥麩胺基硫轉移酶 AtGSTU17 突變株耐旱及耐鹽性狀為麩 胺基與離層酸共同作用之結果
Drought and Salt Stress Tolerance of Arabidopsis Glutathione S-Transferase U17 Knockout Mutant are Attributed to the
Combined Effect of Glutathione and Abscisic Acid
陳瑞宏 Jui-Hung Chen
指導教授:林讚標 博士
Advisor: Tsan-Piao Lin, Ph.D.
中華民國一百年十二月
December, 2011
中文摘要
麩胺基硫轉移酶 (glutathione s-transferases, GSTs) 在氧化逆境代謝中扮 演一個相當重要的角色,但對於這類基因在植物體內所扮演的個別功能卻 所知有限。在整個 GST 基因家族中 GLUTATHIONE S-TRANSFERASE U17
(AtGSTU17
, At1g10370) 曾有被報導其參與了光的傳導訊息調控;經由與 phyA 的 交互作用,影響了植物體內的 GSH 含量,進一步影響了植物的生長發育。本篇論文提供進一步的研究證據顯示
AtGSTU17
在乾旱及鹽分逆境下扮演一個重要的負調控角色。阿拉伯芥的
atgstu17
突變株比野生型更為耐旱及耐鹽。生理分析顯示
atgstu17
突變株的植物體累積了較高含量的 GSH 與 ABA,同時在發芽時期對於 ABA 較不敏感,葉片的氣孔孔徑較小,較低的水分蒸發速率,根系 發育更為茂盛以及較長的營養生長期等生理性狀。
為了釐清
atgstu17
突變株累積的 ABA 是否是因為 GSH 含量升高引起,我們 對野生型澆灌 GSH 溶液進行研究。結果發現澆灌 GSH 溶液的野生型,其 ABA含量較未澆灌 GSH 的植株高,同時顯現
atgstu17
突變株的生理性狀,如開花延遲,根部發育,較為耐鹽與耐旱等。進一步研究
atgstu17
突變株是否是因為 GSH 與ABA 累 積 影 響 而 產 生 了 上 述 的 生 理 性 狀 。 我 們 將
atgstu17
突 變 株 種 植 在 L-buthionine-(S
,R
)-sulfoximine (BSO)溶液中。BSO 是一種可以抑制植物的 GSH 生合成的專一性藥劑。當
atgstu17
突變株的 GSH 含量受到 BSO 抑制減少到與野生型相同的含量時,觀察其性狀與生理反應,如根系,開花時間與對於鹽分與乾旱
的耐受性等,顯示與野生型類似。由以上實驗結果可以得到一個結論,
atgstu17
突變株的外表性狀是由於其植物體內含有較高的 GSH 與 ABA 所共同作用而產生 的結果。同時經由 DNA 微陣列 (microarray) 的資料顯示許多與生長或逆境相關 的轉錄調控基因受到 AtGSTU17 的影響而改變表現。綜合以上實驗資料結果,我 們推測 AtGSTU17 扮演了植物逆境訊息傳導反應的負調控功能。
ABSTRACT
Although glutathione S-transferases (GSTs) are thought to play major roles in oxidative stress metabolism, little is known about the regulatory functions of GSTs.
We have reported that GLUTATHIONE S-TRANSFERASE U17 (AtGSTU17, At1g10370) participates in light signaling and might modulate various aspects of development by affecting glutathione (GSH) pools via a coordinated regulation with phyA. Here we provided further evidence to support a negative role of AtGSTU17 in drought and salt stress tolerance.
When AtGSTU17 was mutated, plants were more tolerant to drought and salt stresses compared to wild-type (WT, Col-0) plants. In addition, atgstu17 accumulated higher level of GSH and abscisic acid (ABA), and exhibited hyposensitivity to ABA during seed germination, smaller stomatal apertures, a lower transpiration rate, better development of primary and lateral root systems, and longer vegetative growth.
To explore how atgstu17 accumulated higher ABA content, we grew WT in the solution containing GSH and found that plants accumulated ABA to a higher extent than plants grown in the absence of GSH, and exhibited the atgstu17 phenotypes. WT plants treated with GSH also demonstrated more tolerant to drought and salt stresses.
Furthermore, the effect of GSH on root patterning and drought tolerance was confirmed by growing the atgstu17 in solution containing L-buthionine-(S,R)- sulfoximine (BSO), a specific inhibitor of GSH biosynthesis.
In conclusion, the atgstu17 phenotype can be explained by the combined effect of GSH and ABA. Microarray analysis provided evidence that expressions of many genes related to growth and stress inducible transcription factors altered in the
atgstu17 mutants. We propose a role of AtGSTU17 in adaptive responses to drought
and salt stresses, by functioning as a negative component of stress-mediated signaltransduction pathways.
CONTENTS
中文摘要 ... I ABSTRACT... II CONTENTS ... IV
INTRODUCTION ...1
MATERIALS AND METHODS ...5
Plant Material and Growth Conditions ...5
Stress-tolerance Tests, Water Loss Measurement and Feeding Experiment ...5
Seed Germination and Stomatal Aperture Measurements...6
Histochemical GUS Assay ...7
Subcellular Localization...7
Quantification of the GSH and ABA Content...8
RNA Isolation and Quantitative Real-time (q)RT-PCR Analysis...9
Microarray Analysis ...9
RESULTS...11
AtGSTU17 Gene in Arabidopsis thaliana...12
AtGSTU17 Affects Arabidopsis Developments...12
Tolerance to Drought and Salt Stresses of the atgstu17 Mutants...13
Effect of Abiotic Stresses on AtGSTU17 Gene Expression...13
Tissue-specific expression of the AtGSTU17 protein ...14
The atgstu17 Mutants Exhibit a Reduced Water Loss and Smaller Stomatal Aperture ...14
The atgstu17 Mutants Show Altered Physiological Responses Regulated by ABA...15
The atgstu17 Has Higher GSH and ABA Contents Compared to WT Plant ...16
Exogenous GSH Induced ABA Accumulation in Planta ...17
Effects of Exogenous GSH and ABA on Seed Germination, Stomata Aperture Size, Root architecture and Stress Tolerances ...17
atgstu17 Phenotypes were Abolished by GSH Synthesis Inhibitor...18
Effects of GSH and ABA on Stress Tolerances ...19
Global Gene Expression in AtGSTU17-Knockout Plants Identified by a GeneChip Analysis...20
Expression of Selected Genes from the Microarray Dataset in the
AtGSTU17 Mutant Lines and WT Plants ...22
AtGSTU17 Regulation of ABA-Downstream and ABA-independent
Gene Expressions under Dehydration Conditions...22
AtGSTU17 Induced by ABA-Dependent and -Independent Pathways under Drought Stress Treatment ...23
DISCUSSION ...25
atgstu17 plants accumulated higher level of GSH in shoot and root...26
AtGSTU17 Plays a Negative Role in Drought and Salt Stress Tolerance...26
The Role of GSH and ABA in Enhancing Drought and Salt Tolerance...27
GSH’s Effects on the Stomata Aperture Size and Root Patterning ...29
REFERENCE...31
TABLES...39
FIGURES...50
APPDENDIX ...83
INTRODUCTION
In both animals and plants, glutathione S-transferases (GSTs; EC 2.5.1.18) are induced by diverse environmental stimuli, with increased GST levels used to maintain cell redox homeostasis and protect organisms against oxidative stress. GSTs were proposed to afford protection under various stress conditions by detoxifying endogenous plant toxins that accumulate as a consequence of increased oxidative stress (Marrs, 1996). In plants, GST expression is induced by phytohormones, such as salicylic acid, ethylene, cytokinin, auxin, abscisic acid (ABA) (Marrs, 1996), methyl jasmonate (Moons, 2003) and brassinosteroid (Deng et al., 2010). It is obvious that GSTs are also stimulated by various stresses, such as pathogen infections, herbicide applications, hydrogen peroxide, ozone, 2,4-dichlorophenoxy-acetic acid (2,4-D), heavy metals, dehydration, senescence, wounding (Marrs, 1996), hypoxic stress, and salt (Moons, 2003), as well as different qualities of light (Loyall et al., 2000;
Tepperman et al., 2001; Chen et al., 2007). Glutathione (GSH) is an essential thiol antioxidant as well as a scavenger of reactive electrophilic compounds, functioning with GSTs to detoxify a range of herbicides (Marrs and Walbot, 1997; Edwards et al., 2000), by tagging electrophilic compounds for removal during oxidative stress.
Theoretically, GST activities catalyze the conjugation of electrophilic compounds to GSH and target them for storage in vacuoles or apoplast (Marrs, 1996). Some plant GSTs play direct roles in reducing oxidative damage (Cummins et al., 1999; Roxas et al., 2000) and enhancing tolerance to stresses (Edwards and Dixon, 2005).
The GST family of Arabidopsis thaliana contains 54 members belonging to seven distinct classes (Fig. S1) (Dixon et al., 2009). The plant-specific phi (GSTF) and tau (GSTU) classes are the largest, with 13 and 28 members, respectively. By applying a range of stress stimuli, with a focus on early changes in gene expressions, Sappl et al.
(2009) indicated that individual GST genes have highly specific induction patterns, and they linked individual GSTs to particular stress stimuli. However, analysis of
metabolite pools of lines in which GSTF genes were silenced showed involvement in protecting plants against oxidation of the primary metabolism. There appears to be a high degree of functional redundancy within the GST family for protecting against oxidative stresses. So far no known specific functions were identified in plant development for most of the members in the large GST gene family indicates a challenge when studying the function of individual genes in response to a stress.
In addition to this well-documented catalytic function, GSTs also function as non-catalytic carrier proteins (Sheehan et al., 2001). Only limited reports revealed the endogenous function of GSTs. For example, At5g17200 (AtGSTF12, also referred to as TT19) is required for the vacuolar uptake of anthocyanins (Kitamura et al., 2004).
Also, GSTs can serve as signaling molecules, and are involved in regulating chalcone synthase following exposure to UV light (Loyall et al., 2000), possibly due to redox-modulated mechanisms. Recently, At1g78730 (AtGSTU20) was demonstrated to physically interact with far-red insensitive 219 (FIN219) in response to light and to play a crucial signaling role in cell elongation and plant development (Chen et al., 2007).
Very little information is available on the involvement of GSTs in response to drought and salt stresses, although changes in the GSH pool, glutathione reductase and glutathione peroxidase activities in dehydrated plants were described (Loggini et al., 1999; Galle et al., 2009). Tobacco seedlings but not mature plants over-expressing a tobacco tau GST gene were more tolerant to low- and high-temperature stresses, and salt stress (Roxas et al., 2000). Tobacco plants overexpressing a tau class of the GST gene, GsGST from Glycine soja exhibited enhanced dehydration tolerance (Ji et al., 2010). However, no further study was provided to support a hypothesis of the regulatory role of this gene in drought-exposed plants.
Recent studies have shown that AtGSTU17 transcripts were induced by FR light
irradiation and regulated by different photoreceptors, especially phyA. Its loss-of-function mutants resulted in a long-hypocotyl phenotype under FR light and delayed flowering under long-day conditions (Jiang et al., 2010). AtGSTU17 came to our attention because it was regulated by FIN219/JAR1 and rapidly increased with far-red (FR) light irradiation but was inhibited by a phyA mutation (Tepperman et al., 2001; Jiang et al., 2010). The full-length complementary DNA (cDNA) of AtGSTU17 encodes a 227 amino acid protein. The recombinant proteins generated from an
Escherichia coli expression system showed enzymatic activities to the substrates GSH
and 1-chloro-2,4-dinitrobenzene (CDNB), which indicates that AtGSTU17 has high affinity to both substrates (Dixon et al., 2009; Jiang et al., 2010). Moreover, we found that AtGSTU17 participates in phyA-mediated photomorphogenesis, and integrates with various phytohormones to modulate GSH homeostasis in regulating Arabidopsis development (Jiang et al., 2010). Surprisingly, atgstu17 plants exhibited robust root system development especially under stress conditions and lack of sensitivity to ABA-mediated inhibition of lateral root elongation. Our present data clearly elucidate AtGSTU17 functions in an undiscovered negative role of adaptation to drought and salt stresses, and the underlying mechanism of the atgstu17 phenotypes can be explained by the synergic action of GSH and ABA which accumulated to a much higher levels than in WT plants. GSH in addition to ABA in protection of plant under drought and salt stress therefore are important for the survival and growth of eukaryotic organisms.MATERIALS AND METHODS
Plant Material and Growth Conditions
Arabidopsis thaliana WT, transgenic plants, and T-DNA-tagged mutants used in
this work were of the Col-0 ecotype. The mutant seeds with a T-DNA insertion in theAtGSTU17 gene (Salk_139615 line for atgstu17-1 and Salk_025503 for atgstu17-2)
were obtained from the Arabidopsis Biological Resource Center. Vector construction and plant transformation for generating AtGSTU17-overexpressing lines were previously described (Jiang et al., 2010). For creating complementary lines,35S:GSTU17OE-5/atgstu17-1 and 35S:GSTU17OE-3/atgstu17-2, the plasmid
originally used in the paper of Jiang et al. (2010) was applied.Seeds were sown in a 2:2:1 mixture of vermiculite: perlite: and peat moss. Plants were placed at 4 oC for 3 days in the dark for stratification and then transferred to normal growth conditions. Plants were grown at 22 oC under long-day conditions (a 16-h light/8-h dark cycle). For in vitro culture, seeds were surface-sterilized by treatment with 70% ethanol for 5 min, followed by commercial bleach (0.5% sodium hypochlorite) containing 0.05% Triton X-100 for 20 min, followed by four washes with sterile distilled water. Seeds were stratified in the dark at 4°C for 3 d. Then, seeds were sown on half-strength MS medium composed of MS basal salts, 1% agar, and 1% sucrose. The pH was adjusted to 5.7 with potassium hydroxide before autoclaving. Plates were sealed and incubated in a growth chamber at 22 °C under a 16-h light, 8-h dark photoperiod.
Stress-tolerance Tests, Water Loss Measurement and Feeding Experiment
For the drought-tolerance test, plants were initially grown in soil under a normal watering regime for 3 weeks. Watering was then halted and observations were made
after a further 10~12 days without water. When WT plants exhibited lethal effects of dehydration, watering was resumed and the plants were allowed to grow for a subsequent 5 days. For the salt-tolerance test, 3-week-old plants were watered for 12 days at 4-day intervals with increasing concentrations of NaCl of 100, 200, and 300 mM. The survivor was recognized by examining the inflorescence base if it still remains green. For freezing tolerance test, 3-week-old plants (WT and atgstu17mutant lines) were grown in a single pot at 22 oC under 16-h light/ 8-h dark conditions.
Three-week-old plants were cold-acclimated (2 oC) for 12 hours. The samples were transferred into freezer at -6 oC for 18 hours. After freezing treatment, the plants were grown in normal condition for 10 days and calculated survival rate.
For transpiration (water loss) measurements, detached leaves from 5-week-old plants were exposed to room temperature (25°C). Leaves were weighed at various time intervals, and the loss of fresh weight (%) was used to indicate water loss.
Exogenous GSH has been used in feeding experiment because it could be taken up by the plant root system (Lappartient & Touraine, 1997; Tausz et al., 2004). For feeding experiment, we germinated the Arabidopsis seeds in Petri dish for 7 days and transferred the seedlings into 1/2 MS agar plate supplemented with GSH or ABA in regular growth condition (22 oC under 16-h light/ 8-h dark cycles) for two weeks. We analyzed the stability of GSH in the growth medium and found the GSH is stable for 2 weeks (Fig. S2). Or the seedlings were transferred into soil medium in regular growth condition supplemented with GSH or ABA or combinations of GSH and ABA for another 2 weeks. To prevent degradation or oxidation of GSH and ABA, the water solution was replaced every two days with newly prepared chemicals.
Seed Germination and Stomatal Aperture Measurements
Imbibed seeds were cold-treated at 4 oC in the dark for 3 days, and moved to 22
°C with a 16/8-h light/dark photoperiod. Germination was defined as 1-mm protrusion of the radicle. Epidermal peels were stripped from fully expanded leaves of 5-week-old plants, and were floated in a solution of 30 mM KCl and 10 mM MES-KOH, pH 6.15 in Petri dishes. After incubation for 2.5 h under white light at 22
°C to induce stomatal opening, different concentrations of ABA were added. Stomatal apertures were recorded under an Olympus BX51 system microscope, and were analyzed using DP-PSW software. Measurements were performed using the free software IMAGEJ 1.36b (Broken Symmetry Software; http://brokensymmetry.com).
Histochemical GUS Assay
To investigate AtGSTU17 gene expression, approximately 1.5 kb of the promoter (-1376 to -1 from the translation initiation codon) was amplified by PCR from genomic DNA. The PCR product was inserted into the pCAMBIA 1391Z vector at the PstI and BamHI sites upstream from the GUS gene. Twenty-five hygromycin-resistant transgenic (T1) plants were obtained. Four single-copy insertion lines were identified by Southern blotting (data not shown). Histochemical assays for GUS activity in transgenic plants were performed as described by Jefferson et al. (1987). Tissues were visualized using an Axiophot microscope (Olympus BX51 system) coupled to a CCD camera.
Subcellular Localization
For subcellular localization, the cDNA fragment containing the AtGSTU17 coding region without stop codon was amplified by PCR. The PCR product was then inserted downstream from the CaMV 35S promoter and in frame with the 5’ terminus of the GFP gene in the pEarlyGate 103 vector (obtained from ABRC) using the Gateway (Invitrogen) system, according to the manufacturer’s instructions. The
construct was subsequently delivered into onion epidermal cells by microprojectile bombardment using a PDS-1000/He biolistic particle 18 delivery system (Dupont), essentially according to the manufacturer’s instructions. Onion epidermal cell layers were placed on 1% agar plates with half-strength MS salts and bombarded using a rupture disk of 900 Pascal per square inch at a target distance of;10 cm. At 24 h after bombardment, GFP fluorescence was analyzed with the 488-nm argon laser 19 using Olympus BX51 system.
Quantification of the GSH and ABA Content
Leaf tissues of 200 mg were ground with mortar and pestle in the liquid nitrogen.
Subsequently, 2 ml of 1 mM EDTA and 6% (v/v) metaphosphoric acid, pH2.8, were added and mixed, then centrifuged at 15000 g for 20 min. Supernatant was neutralized with 0.2 M NaOH. The final pH of the neutralized acid extracts was between 5 and 6.
The methods used to measure the total level of glutathione (GSH + GSSG) were as described by Griffith (1980). The oxidized glutathione in the supernatant was reduced to glutathione by glutathione reductass. Glutathione was determined in a kinetic assay in which the reduction of 5,5-dithiobis (2-nitrobenzoic acid) (DTNB) to yellow TNB was spectrophotometrically measured at 412 nm.
For determining endogenous ABA contents in aerial parts, leaf tissues of 200mg
harvested at appropriate stages were treated with extraction buffer (80% methanol and
2% glacial acetic acid) for 24 h under darkness, followed by centrifugation for 10 min
at 2,000g. Supernatants were taken up and dried in a speedvac, then resuspended in
100% methanol plus 0.2
MNH
4H
2PO
4(pH 6.8) for 10 min. To avoid plant pigment
and other nonpolar compound effects on the immunoassay, the extracts were first
passed through a polyvinylpolypyrrolidone column and then C18 cartridges. Elutes
were concentrated to dryness in a speedvac and resuspended in Tris-buffered saline
for immunoassay (Hsu and Kao, 2003 ). For ABA determination, ABA was quantified by ELISA (Phytodetek ABA kit; Agdia) according to the manufacturer's protocol.
RNA Isolation and Quantitative Real-time (q)RT-PCR Analysis
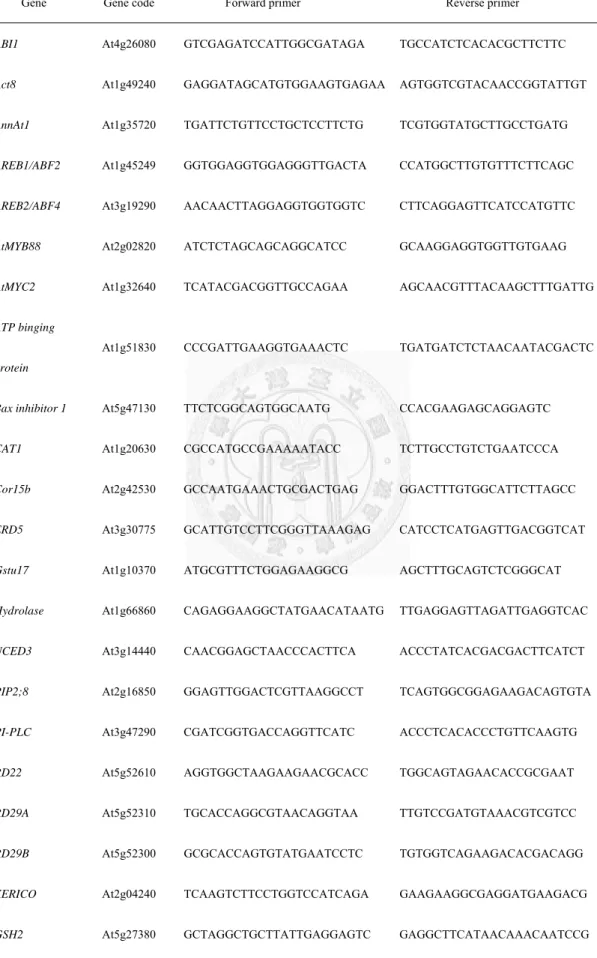
Total RNA samples were isolated from various plant tissues with the RezolTM C&T reagent (PROtech, Taiwan). For the RT-PCR, SuperScript III M-MLV Reverse Transcriptase (Invitrogen; http://www.invitrogen.com) was used, following the manufacturer’s instructions. The resulting single-stranded cDNA was then used as the template in a qRT-PCR. qRT-PCRs were carried out with gene-specific primers, designed using Vector NTI 9.0 Software. For the qRT-PCR experiments, KAPA SYBR Premix ExTaq was used according to the manufacturer’s instructions (KAPA Biosystems, USA) using the BIO-RAD MyiQTM (Hercules, CA, USA). qRT-PCR experiments were carried out in three separate biological replicates. Primers used for the qRT-PCRs are listed in Table 3.
Microarray Analysis
Three independent biological replicates of microarray experiments were performed using 4-week-old WT and AtGSTU17-knockout plants grown under normal conditions. Total RNA was isolated from the rosette leaves using RezolTM C&T reagent (PROtech, Taiwan). One μg of total RNA was amplified by a Quick-Amp Labeling kit (Agilent Technologies, USA) and labeled with Cy3 or Cy5 (CyDye, PerkinElmer, USA) during the in vitro transcription process. CyDye-labled cRNA (0.825 μg) was fragmented to an average size of about 50-100 nucleotides by incubation with fragmentation buffer at 60 oC for 30 minutes. Correspondingly fragmented labeled cRNA is then pooled and hybridized to Agilent Arabidopsis V4
Oligo 4×44K Microarray (Agilent Technologies, USA) at 60 °C for 17 h. After washing and drying by nitrogen gun blowing, microarrays are scanned with an Agilent microarray scanner (Agilent Technologies, USA) at 535 nm for Cy3. Scanned images are analyzed by Feature extraction 9.5.3 software (Agilent Technologies, USA), and normalization software is used to quantify signal and background intensity for each feature. Genes exhibiting more than a 2-fold enhanced or reduced transcription level in three independent experiments were considered to show significant alterations in expression, and P values for the Benjamini and Hochberg method (false discovery rates; FDR) were calculated by by Genespring 11 (Agilent Technologies, USA).
RESULTS
AtGSTU17 Gene in Arabidopsis thaliana
In Arabidopsis thaliana, the AtGSTU17 gene consists of two exon and one intron, and locates on the first chromosome. In TAIR database, AtGSTU17 gene has 1278 bp in total, and the coding sequence has 684 bp. This gene encodes a 227-amino-acid polypeptide (25.3 kD) with an isoelectric point of 6.54 according to sequence prediction. AtGSTU17 encodes a glutathione S-transferase. It belongs to the
Tau class (Fig. S1) GST family.
According to the expression pattern under various conditions by Arabidopsis eFP Browser, AtGSTU17 can be induced by salt stress, osmotic stress, cold stress and drought stress (Fig. S3). AtGSTU17 was expressed in stems, leaves, and flowers, but has lower expression levels in root and matured siliques (Fig. S4). AtGSTU17 can be induced by ABA and expressed in mesophyll and guard cell (Fig. S5 and S6). The expression of AtGSTU17 also shows a circadian rhythm (Fig. S7).
AtGSTU17 Affects Arabidopsis Developments
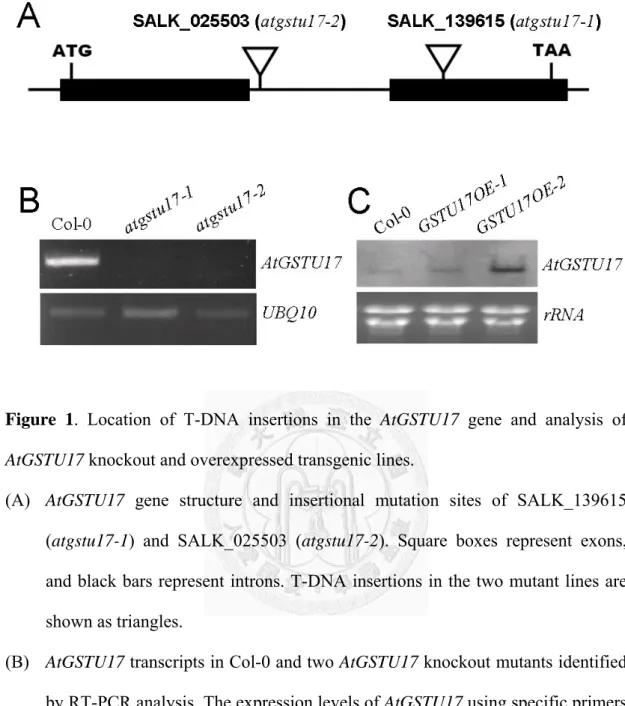
Two independent T-DNA insertions of AtGSTU17, atgstu17-1 (SALK_139615) and atgstu17-2 (SALK_025503) are located in the second exon and first intron of
AtGSTU17 (Fig. 1A). RT-PCR analyses indicated that atgstu17-1 and atgstu17-2 were
null mutants (Fig. 1B). First, We observed a delay flowering phenotype in atgstu17 mutants (atgstu17-1 and atgstu17-2) under long-day condition(16 h light/8 h dark)(Fig. 2A) and this phenomenon is more significant under short-day condition(12 h light/12 dark)(Fig. 3A). The atgstu17 mutants exhibited a smooth elliptical leaf shape, and a profusely growing root system (Fig. 3B) and delayed flowering time which produced a leaf number of 27 in contrast to 17 for wild-type (WT, Col-0) plants (Fig. 3C). In contrast, ectopic expression of AtGSTU17 under the control of the CaMV35S promoter in WT, GSTU17OE-1 and GSTU17OE-2, confirmed by RNA-gelblotting (Fig. 1C), resulted in an uneven leaf surface and root system similar to WT plants but had an earlier flowering time and only produced a leaf number of 12 (Fig.
3).
Tolerance to Drought and Salt Stresses of the atgstu17 Mutants
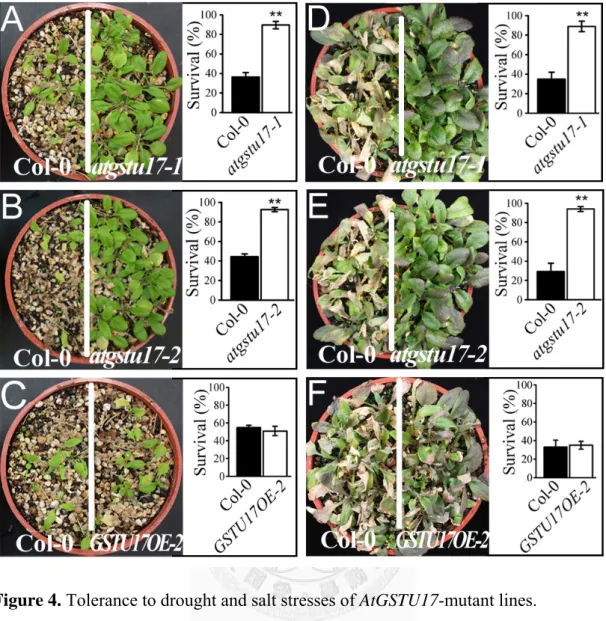
Since AtGSTU17 was induced by exogenous ABA (Jiang et al., 2010), we investigated the response to prolonged periods of drought and found that both
atgstu17-1 and atgstu17-2, had improved resistance to water deficits (Fig. 4A and 4B).
Nearly all of the atgstu17 plants had recovered and begun to grow again, while only 40% of the WT could resume growth. GSTU17OE plants did not exhibit a difference in water deficit compared to WT plants (Fig. 4C). Enhanced salt stress tolerance of the atgstu17-1 and atgstu17-2 was also observed (Fig. 4D and 4E). However,
GSTU17OE did not exhibit a difference in salt tolerance compared to WT plants (Fig.
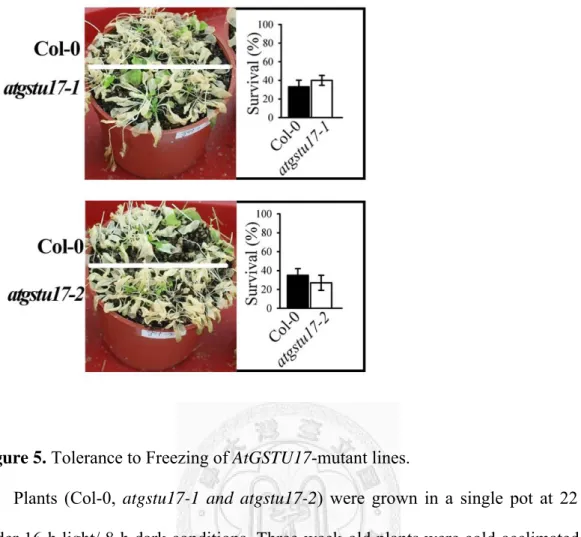
4F). But in freezing treatment, atgstu17-1 and atgstu17-2 did not show more tolerance compared to WT plants (Fig. 5)
To confirm that the mutated AtGSTU17 was the cause of these phenotypes, the
atgstu17-1 and atgstu17-2 were transformed with a cDNA of AtGSTU17 driven by the
35S promoter. These 35S:AtGSTU17/atgstu17 transgenic plants showed the leaf morphology, bolting time, and sensitivity to drought stress were similar to WT plants (Fig. 6), verifying the role of AtGSTU17 in plant growth and development.Effect of Abiotic Stresses on AtGSTU17 Gene Expression.
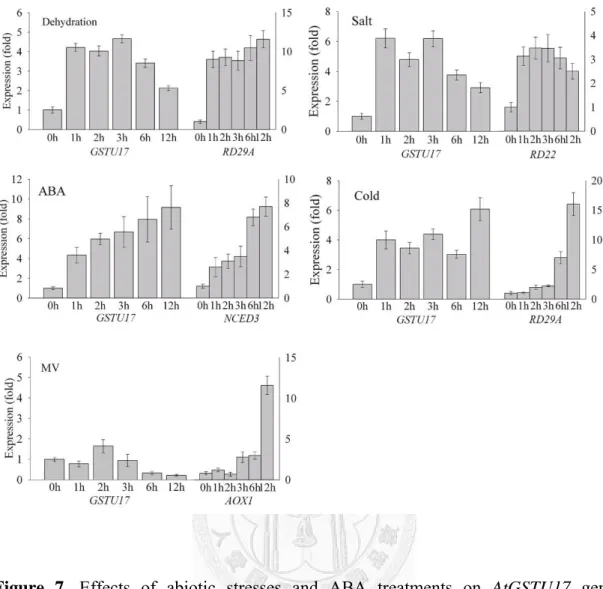
To assess if AtGSTU17 can play a role in plant stress responses, a wide spectrum of abiotic stressors was assessed. We observed up-regulation of AtGSTU17 mRNA levels after practically all employed stimuli (Fig. 7). The most pronounced and persistent induction took place after treatment with the stress hormone ABA, with the
transcript level steadily increased without saturation after 12 h of stimulation. Drought, cold and NaCl treatment resulted in similar transient mRNA elevation that declined within 12 h. The increase in AtGSTU17 transcript levels occurred as soon as the increase in RD29A and RD22 (marker genes for abiotic stresses) transcripts, or earlier than the marker gene of cold. AtGSTU17 could also be induced slightly by oxidative stress (paraquat).
Tissue-specific expression of the AtGSTU17 protein
It has been reported that most of the GFP fusions of AtGSTUs, including AtGSTU17 is localized in the cytosol (Dixon et al., 2009). For subcellular localization of the protein, AtGSTU17 cDNA was fused in frame to the N-terminal side of the GFP marker gene and expressed in transgenic Arabidopsis under the control of the cauliflower mosaic virus (CaMV) 35S promoter (Fig. 8A). Confocal imaging of GFP revealed that the AtGSTU17-GFP fusion protein accumulated in the cytosol and nucleus in the onion epidermal cells (Fig. 8B) and in the protoplast of Arabidopsis (Fig. 8C).
In transgenic Arabidopsis expressing the GUS reporter gene driven by the
AtGSTU17 promoter (Fig. 9A), GUS activity was mainly observed in guard cells
surrounding hydathodes, trichomes, the shoot and root apical meristems, lateral root primordial, and vascular tissues (Fig. 9, B-D). Strong GUS expression was observed in guard cells and lateral root primordia as well as vascular tissues under ABA treatment (Fig. 9E) and NaCl (Fig. 9F) and mannitol treatments (Fig. 9G).The atgstu17 Mutants Exhibit a Reduced Water Loss and Smaller Stomatal Aperture
As a reduction in stomatal pore aperture size mediated by ABA is a critical aspect
of the response of plants to drought stress, we speculated that the enhanced drought resistance of the atgstu17plants might be correlated with an altered response to water scarcity. Indeed, the rate of water loss from atgstu17s (atgstu17-1 and atgstu17-2) was lower than that from WT plants, as measured bythe fresh-weight loss of detached leaves (Fig. 10).
We found that stomata of atgstu17 had a constitutively reduced aperture (Fig. 11, A and B). ABA treatment reduced the stomatal aperture to a similar extent in mutant and WT plants when the smaller initial aperture of atgstu17 was taken into account.
This constitutively smaller aperture size of stomata may explain the observed lower water loss rate of detached leaves by the atgstu17 plants (Fig. 10). The stomatal aperture response to ABA treatment in GSTU17OE was similar to that of WT plants (data not shown).
The atgstu17 Mutants Show Altered Physiological Responses Regulated by ABA Since expression of AtGSTU17 was induced by ABA , we speculated that
germination of mutant seeds in responding to ABA might be altered. At a concentration of 2 μM ABA the germination rate of atgstu17 was 50% compared to no germination in WT seeds (Fig. 12). In contrast, germination of the GSTU17OE seeds was similar to that of WT plants. This indicates that germination of atgstu17 seeds was less sensitive to ABA.Root development is also sensitive to ABA (Sharp & LeNoble 2002, De Smet et al. 2006) and this was further studied in AtGSTU17 mutants. Under white light and unstressed condition for 2 weeks, root lengths among WT, atgstu17 and GSTU17OE plants were similar (Fig. 13A). However, the atgstu17 mutant was significantly less sensitive to ABA suppression of primary root and lateral root elongation (Fig. 13B), while GSTU17OE appeared to be more sensitive compared to WT plants (Fig. 13C).
To more-systematically evaluate the effects of ABA, mutant plants were grown on vertical half-strength MS agar plates supplemented with various concentrations of ABA (0~5 μM). Whereas primary root lengths were reduced by 50% in the presence of 5 μM ABA in 2-week-old atgstu17 plants, they were reduced by 70% in WT plants (Fig. 13D). The number of lateral roots longer than 0.5 cm per cm of primary root was significantly suppressed in atgstu17 compared to WT ones under control and ABA treatment conditions (Fig. 13E). The average lateral root length was much greater in the atgstu17 plant than WT plants in the control and in the presence of ABA (Fig.
13F).
In contrast, primary root lengths of GSTU17OE were similar to the WT plants in control condition but were reduced in the presence of 5 μM ABA (Fig. 13, C and G).
The lateral root number per cm of primary root of the GSTU17OE is significantly greater than the WT plants only in the 5 μM ABA treatments, but the average lateral root length of the GSTU17OE did not differ from the WT plants (Fig. 13, H and I).
The atgstu17 Has Higher GSH and ABA Contents Compared to WT Plant
Using the model xenobiotic substrate CDNB as well as BITC (benzylisothiocyanate) to measure GSH-conjugating activities, AtGSTU17 exhibited high specific activity when compare to other GSTs (Dixon et al., 2009). To investigate whether AtGSTU17 mutations affected GSH levels during vegetative growth, we measured GSH in leaves and roots of WT and all mutant plants under normal growth conditions. GSH levels in the atgstu17s were significantly higher than that in WT plants (Fig. 14A). GSH contents in roots of atgstu17s were also greater than that of WT and GSTU17OE plants (Fig. 14A). In previous report, the GSH/GSSG ratio was significantly higher in the atgstu17 than in the WT plants under normal growth conditions (Jiang et al., 2010). Taken together, loss-of-function of AtGSTU17
contributed to an increased level of GSH and redox potential in plants growing in normal growth conditions.
Because the stress tolerant phenotype of the atgstu17, we suspected that ABA content might be altered in the AtGSTU17-mutant plants. Surprisingly, the ABA content in leaves was respectively 2- and 2.3-fold significantly higher in the
atgstu17-1 and atgstu17-2 than in WT plants, whereas content was lower in the
overexpressors than in WT plants (Fig. 14B). ABA-deficient mutant aba2 (GLUCOSE INSENSITIVE 1 or GIN1), which contains low level of ABA, was used as a reference, and had approximately 1/3 of the WT level of ABA.Exogenous GSH Induced ABA Accumulation in Planta
We hypothesized that the accumulation of ABA in the atgstu17 lines was resulted from higher GSH content. To test this hypothesis, WT plants were grown in the solution with or without GSH for two weeks, and the leaf ABA content was measured.
The plants grown in solution containing 200 μM GSH accumulated an ABA level that was 1.8-fold greater than the level detected in plants without exogenous GSH treatment (Fig. 15). Plants treated with 400 μM GSH showed 1.4-fold higher ABA content. Our results support that ABA accumulation can be promoted by exogenous GSH treatment.
Effects of Exogenous GSH and ABA on Seed Germination, Stomata Aperture Size, Root architecture and Stress Tolerances
Knowing that GSH and ABA accumulated to higher extents in the atgstu17, we investigated the effect of these two chemicals separately and in combination on Arabidopsis. We found that GSH was able to suppress the germination inhibition caused by ABA. When treated with both GSH and ABA, seeds have a higher
germination rate than seeds treated with ABA alone (Fig. 16A). The effect of GSH on the intrinsic stomata aperture size was not known. Figure 16, B and C showed that stomatal apertures were significantly smaller in plants grown in the GSH-containing solutions.
To evaluate the effect of GSH and ABA on the root architecture, WT seedlings were grown on vertical 1/2 MS agar plates supplemented with various concentrations of GSH and/or ABA for 2 weeks. Whereas primary root lengths were reduced by 33%
in the presence of 3 μM ABA in 2-week-old WT plants, they were increased by 62%
in plants growing in the 25 and 50 μM GSH (Fig. 17, A and B). However, higher GSH concentrations repressed primary root growth. The number of lateral root longer than 0.5 cm per cm of primary root was significantly increased in various concentrations of GSH (Fig. 17, A and C). The combinations of 3 μM ABA and GSHs slightly suppressed the lateral root number (Fig. 17C). The average lateral root length was much greater in WT seedlings receiving 3 μM ABA, and combinations of 3 μM ABA and GSHs (Fig. 17, A and D). GSH treatment alone had no effect on the lateral root length. We concluded that GSH and ABA have different effect on the root growth and development.
atgstu17 Phenotypes were Abolished by GSH Synthesis Inhibitor
To further elucidate the effect of reduced level of GSH in the atgstu17 on the phenotypes, we grew atgstu17 mutants on vertical 1/2 MS agar plate containing 3μM BSO. BSO is a highly specific inhibitor of the first enzyme of GSH biosynthesis, and its application results in the depletion of cellular GSH (Vernoux et al., 2000). The GSH level in BSO treated leaves was reduced compared to the mutants without BSO treatment (Fig. 18A). The atgstu17 seedlings exhibited root development similar to WT plants by 2 weeks after germination on medium containing 3 μM BSO (Fig. 18B
and 18C), specially in lateral root development and (Fig. 18D and 18E). These observations support that root architecture of atgstu17 to some extent is attributed to the GSH content. Those results support that GSH accumulation is important factor in root development in atgstu17s.
Effects of GSH and ABA on Stress Tolerances
We know the concentrations of GSH and ABA in atgstu17s contribute to stomatal closed and root development. Therefore, we want to confirm the effects of GSH and ABA on stress tolerances in atgstu17s. To assess if exogenous ABA and/or GSH could confer drought tolerances, we grew WT plants in water containing GSH or ABA or combinations of GSH and ABA. We found that plants growing in water containing 400 μM GSH in the presence or absence of ABA recovered and resumed growth from the drought stress test, while no plants could resume growth in water only (Fig. 19B). Exogenous GSH gave better protection than ABA in drought conditions. For salt-tolerance test, all of the WT plants receiving ABA and/or GSH exhibited enhanced salt stress tolerance and much-delayed leaf chlorosis (Fig. 19B).
All of the plants growing in water containing 3 μM ABA or a combination of ABA and GSH resumed growth. Three μM ABA gave better protection than GSH in saline conditions.
From the experiment in Figure 19, we learned that increased GSH level confers drought and salt tolerance of WT plants. To test this observation using different approach, we grew the atgstu17 in water containing 20 and 50 μM BSO for two weeks and found that it exhibited reduced drought tolerance compared to control mutants (Fig. 20A). BSO treatment also reduces the bolting time of the atgstu17s (Fig.
20B). This experiment confirms the direct link between the phenotype of the atgstu17 and accumulation of GSH.
Global Gene Expression in AtGSTU17-Knockout Plants Identified by a GeneChip Analysis
Microarray is a powerful tool to analyze the global gene expression. AsGATU17 can regulate the root development, stomatal closure and abiotic stresses by accumulating GSH and ABA. But the molecular mechanism is unclear. We were interested in determining if the altered gene expression conferred stress tolerance and in assigning a function to AtGSTU17 as a negative regulatory component of the stress response. A microarray analysis was employed for atgstu17-2 plants using an Agilent Arabidopsis 2 Oligo Microarray (Agilent Technologies, http://www.agilent.com) which covers >21,000 Arabidopsis genes. Total RNA extracted from 3-week-old seedlings of Col-0 and AtGSTU17 mutants growing under normal growth conditions was used. Three experiments were performed for each line using different labels, Cy3 or Cy5. Each experiment shows consistent expression profiling using Hierarchical Clustering (Fig. 21). As observed, 1320 genes were found to be upregulated > 2-fold, and 888 genes were found to be expressed by < 0.5-fold compared to WT plants. A large number of transcriptome changes being observed, indicated that the AtGSTU17 gene has an immense influence to the entire genome. Since the potential functions of limited genes in the dataset were studied, we offer selected comments and observations to illustrate important themes.
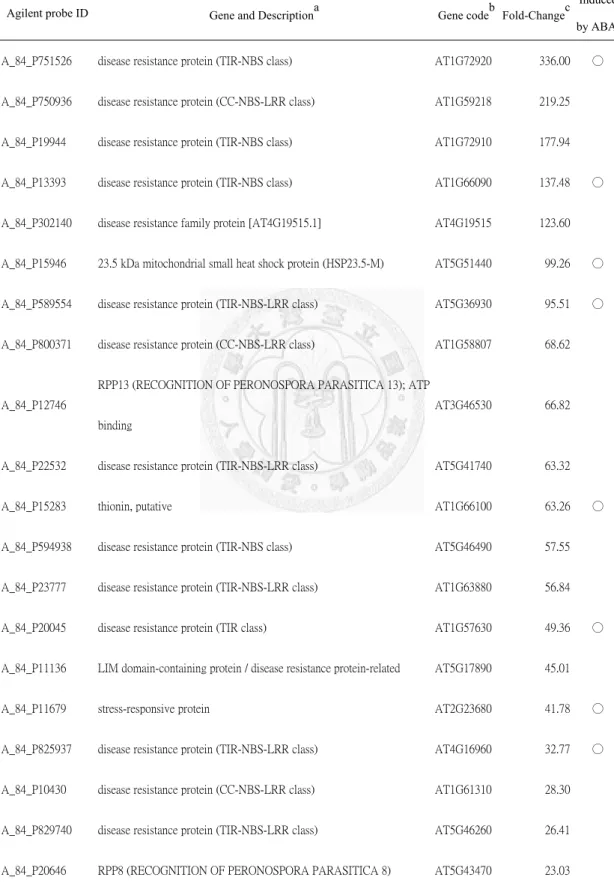
Expression profiles of genes with known function are presented in Table 1 where only genes the expressions of which had increased by a ratio of > 1.5-fold or reduced by < 0.5-fold in atgstu17-2 plants compared to WT plants are listed. Among downregulated genes in atgstu17, PIP2;8 (plasma membrane intrinsic protein 2;8, At2g16850, 0.127-fold) and ATP binding/kinase/protein Ser/Thr kinase (At1g51830, 0.035-fold) showed great repression in atgstu17-mutants. Other genes repressed in
atgstu17 were protein serine/threonine phosphatase (At5g26010, 0.46-fold), and
PIP2;7 (At4g35100, 0.46-fold). Most downregulated genes help improve stress
tolerance. It was interesting to note that many protein kinases were repressed, as well as many transcription factor genes in various families including MYB, bZIP, WRKY, and especially a large number of Zinc finger genes; these results imply that the AtGSTU17 serves as an upstream signaling component under abiotic stress conditions.The increased expression of FLC (At5g10140, 4.34-fold) and repression of the FT (At1g65480, 0.30-fold) agree with the a delayed-flowering phenotype.
Among upregulated genes in atgstu17 mutants, AtPLC8 (Phosphoinositide- specific phospholipase C family protein, At3g47290, 173-fold), AtPLC9 (At3g47220, 20-fold) which catalyzes the hydrolysis of phosphatidylinositol 4,5-bisphosphate into the two second messengers, inositol 1,4,5-trisphosphate and diacylglycerol, and
AREB1 (At1g45249, 24-fold), an ABA-induced ABRE-binding bZIP gene, were
highly induced. In addition, several other drought stress-tolerant genes were also induced including XERICO (At2g04240, 2.31-fold), one of the RING zinc-finger genes, RAP2.4 (At1g78080, 2.23-fold), an AP2/DREB-type transcription factor gene,ENH1 (ENHANCER OF SOS3-1, At5g17170, 2.42-fold), a chloroplast-localized
protein gene, and AnnAt1 (ANNEXIN ARABIDOPSIS 1, At1g35720, 4.16-fold), a Ca2+-dependent membrane-binding protein annexin gene. AtMYB88 (At2g02820;2.54-fold) has the function of generating normal stomatal patterning (Lai, et al., 2005).
The expressions of XERICO, RAP2.4, ENH1, and AtMYB88 were ABA-independent.
Even though we did not observe a freezing-tolerant phenotype, atgstu17 did exhibit elevated cold-responsive genes; for example, KIN1 (At5g15960, 4.59-fold),
KIN2/COR6.6 (At5g15970, 5.46-fold) and COR15b (At2g42530, 3.93-fold), three late
embryogenesis-abundant proteins also induced by ABA.Expression of Selected Genes from the Microarray Dataset in the AtGSTU17 Mutant Lines and WT Plants
For further validation of genes that are upregulated or downregulated in the
atgstu17 mutants, we performed a real-time PCR analysis. The total RNAs isolated
from the WT and atgstu17-2 plants under normal growth conditions were used for the real-time PCR analysis (Fig. 22). Also GSTU17OE and atgstu17-1 were included for comparison.XERICO, AREB1, Bax inhibitor-1, AnnAt1, COR15b, and AtMYB88 of the atgstu17-1- and atgstu17-2-mutant lines were all upregulated to expression levels
similar to those of the array dataset (Fig. 22). These genes maintained similar expression levels as WT in GSTU17OE-1 and GSTU17OE-2 plants except forXERICO, which had a slightly lower level. The PI-PLC gene was difficult to quantify
and exhibited great variations in multiples of expression because the gene probably has no expression in WT and GSTU17OE lines, and the residual activity influenced the measurement of the knockout mutant lines. The expression levels of four downregulated genes also agreed with the array dataset (Fig. 22). Again these genes in GSTU17OE plants maintained similar expression levels as in WT plants except forERD5 and hydrolase which had higher expressions than WT plants. In conclusion, the
global expression of atgstu17-2 based on the microarray dataset was validated and can be used to interpret the function of AtGSTU17 in Arabidopsis.AtGSTU17 Regulation of ABA-Downstream and ABA-independent Gene Expressions under Dehydration Conditions
Using an Agilent GeneSpring GX Analysis to screen our microarray dataset, we found that a total of 112 genes were categorized as stress-responsive genes (Table 2).
Among them, only 42 genes were activated by ABA treatment to 1.3-fold compared to those without treatment by screening them from the two websites, the Arabidopsis
eFP Browser and AtGenExpress Visualization Tool (Table 2). Thus AtGSTU17 has an immense influence on ABA-independent gene expressions as well as classical ABA signaling.
To clarify this point, we assayed one ABA-independent gene, XERICO, altered in
atgstu17 and six ABA-downstream genes, ABI1, AREB1, AREB2, AtMYC2, COR15B,
and RD29B, for comparison (Fig. 23). Under control conditions, gene expressions were the same as those seen in the array dataset. After withholding water for 5 days,XERICO of atgstu17-1 and atgstu17-2 showed greater induction over the WT and
overexpressing mutant lines.ABI1 expressions were the same for the WT and AtGSTU17-mutant lines
indicating no effect on the master regulator of the ABA response. AREB1 under normal conditions exhibited high expression in the atgstu17-mutant lines, which had no effect on the induction of downstream genes, because AREB1 activity is regulated by ABA-dependent multi-site phosphorylation of conserved domains (Furihata et al., 2006; Fujii et al., 2007). After withholding water for 5 days, atgstu17-mutant lines unexpectedly had lower expression of AREB1 than the WT, and RD29B was also strongly suppressed. AREB2 of atgstu17 plants also was repressed compared to WT plants under drought treatment, while AtMYC2 was similar to the WT, but was lower than the GSTU17OE mutant lines. In conclusion, these experiments provide conclusive evidence that AtGSTU17 plays a role as a regulatory component of the ABA-dependent and -independent signaling pathways upstream of many well-studied transcription factors.AtGSTU17 Induced by ABA-Dependent and -Independent Pathways under Drought Stress Treatment
We hypothesize that AtGSTU17 gene expression is only ABA-dependent during dehydration treatment. To test this, two ABA-deficient mutants, aba2 and nced3,
which cannot synthesize ABA because of a lack of the short-chain dehydrogenase/
reductase (SDR) (Cheng et al., 2002), and NCED (9-cisepoxy- carotenoid dioxygenase) were respectively assayed under the normal growth conditions and under dehydration treatment. Surprisingly, the ABA-deficient mutants had 3-fold higher expression of AtGSTU17 than the WT under normal growth conditions, but the aba2- and nced3-mutant lines respectively exhibited 80% and 60% expressions of
AtGSTU17 after 5 days of withholding water (Fig. 23). Taken together, this
experiment indicates that the expression of AtGSTU17 is partially induced by ABA-independent drought stress.DISCUSSION
atgstu17 plants accumulated higher level of GSH in shoot and root
Recent studies have shown that AtGSTU17 transcripts were induced by far-red light irradiation and regulated by different photoreceptors, especially phyA. Its loss-of-function mutants resulted in a long-hypocotyl phenotype under FR light and delayed flowering under long-day conditions (Jiang et al., 2010). In this study, we extend the function of AtGSTU17, the first member of the large GST family in
Arabidopsis thaliana, by playing a negative role in drought and salt stress tolerance.
The basic observation of accumulation of GSH in shoot and root in the atgstu17 (Fig.
14A) agrees with AtGSTU17 having high activity among GSTs in Arabidopsis when tested with different substrates (Dixon et al., 2009). Without GST mediation GSH can not be utilized as the substrate for the xenobiotic detoxification (Rouhier et al., 2008).
As there are no xenobiotics in the system, the turnover of GSH by the GST may be a contributing function but the co-substrate is unknown. The higher GSH content also could be contributed from the increased expression of GSH2 in the atgstu17 mutants (Table 1).
AtGSTU17 Plays a Negative Role in Drought and Salt Stress Tolerance
Stress tolerant phenotype of atgstu17 can be well-explained by the greater GSH and ABA accumulation, and gene expression patterns. According to microarray dataset of atgstu17 plants (Table 1), among upregulated genes, AREB1 and several other drought stress-tolerant genes including XERICO, RAP2.4, ENH1, and AnnAt1 were induced. The expressions of XERICO, RAP2.4, ENH1, and AtMYB88 are ABA-independent according to the Arabidopsis eFP Browser. XERICO overexpression exhibited a marked increase in drought tolerance (Ko et al., 2006).
RAP2.4 was upregulated by drought and salt treatment, and enhanced drought
tolerance when overexpressed (Lin et al., 2008). AnnAt1 induced by ABA and NaCltreatment when overexpressed were more drought tolerant than WT plants (Konopka-Postupolska et al., 2009). ENH1 functions in detoxification resulting from salt stress by participating in a salt-tolerance pathway (Zhu et al., 2007). In conclusion, ABA-dependent and ABA-independent stress tolerant transcription factors and other genes are activated in atgstu17 plants leading to drought and salt tolerant phenotype.
When a gene is repressed and can confer stress tolerance, this gene is usually considered to be a negative regulator of a stress response. Just a few example, knockout mutants of ABI1, ABI2, Nuclear protein X1 (NPX1), and Altered expression
of APX2 8 (ALX8) were more stress tolerant than WT plants, and are considered to be
negative regulators (Merlot et al., 2001; Kim et al., 2009; Wilson et al., 2009). In this paper, we show that atgstu17 exhibited altered transcriptome, metabolites, and morphology of the rosette, and enhanced abiotic stress tolerance. Thus AtGSTU17 plays a negative regulator of drought and salt stress response.The Role of GSH and ABA in Enhancing Drought and Salt Tolerance
An unexpected observation in this study is exogenous GSH treatment associated with the accumulation of ABA in Arabidopsis (Fig. 15). It is surprising that, despite the widely assumed involvement of GSH in abiotic stress signaling in plants, no GSH-mediated drought and salt stress tolerance in plant have been reported in literatures. These facts underline the link between the loss-of-function of AtGSTU17 gene and all the phenotypes we have found in the atgstu17 mutants.
GSH is a determinant of the cellular redox balance, and a major cellular antioxidant. In addition, GSH is an important cellular signaling compound influence many fundamental cellular processes (Foyer and Noctor, 2005). Genetic and other evidence shows that GSH concentration is important in many physiological responses (Foyer and Noctor 2009, and the reference therein). Exposure to drought and salt
stress caused increased formation of ROS and thus oxidative stress. The plants GSH and glutaredoxins are implicated in the response to oxidative and are involved in both detoxification of ROS and transmission of the redox signal (Meyer 2008). The GSH pools in atgstu17 were 35% higher compared to WT plants (Fig. 14A), and the GSH/GSSG ratio in the atgstu17 was also significantly higher than WT plants (Jiang et al., 2010). The redox potential difference might be the cause of tolerant phenotype of the atgstu17 under conditions of stress.
When GSH is depleted, plants frequently exhibit decreased sensitivity to oxidative stress (Kushnir et al., 1995; Grant et al., 1996). Several transgenic plants with elevated levels of GSH have been shown to be resistant to oxidative stress (Foyer et al., 1995; Wellburn et al., 1998). In addition, elevated GSH synthesis by increasing γ-glutamylcysteine synthetase (γ-ECS) activity was shown to correlate with Cd resistance in cultured tomato cells (Chen and Goldsborough, 1994) and chilling tolerance (Kocsy et al., 2000, 2001). During the growing stage, an exogenous supply of GSH alone at 400 μM had an obvious effect of enhancing drought tolerance of WT plants. The effect of enhancing salt tolerance was also evident even at 200 μM GSH because chlorosis of leaf tissues was much delayed compared to WT plants without GSH treatment. Furthermore, if WT plants were exposed to a combination of ABA and GSH, the fitness increased dramatically (Fig. 19), much better than the performance produced by each chemical independently against drought and salt stresses.
An activation of ABA accumulation in Arabidopsis by exogenous GSH provides a link between GSH and drought and salt stress tolerance. ABA accumulation was correlated with increased drought tolerance (Thompson et al., 2007), and conferred drought tolerance in mutants like enhanced drought tolerance1 (a homeodomain-START transcription factor), poly(ADP-ribose) polymerase (PARP),
npx1, isopentenyltransferase (IPT), XERICO (a RING-H2 gene), and alx8 (Ko et al.,
2006; Vanderauwera et al., 2007; Rivero et al., 2007; Yu et al., 2008; Kim et al., 2009;Wilson et al., 2009). In atgstu17 plants the higher ABA levels compared to WT plants under non-stressed conditions are consistent with levels of stress tolerance in these mutant plants. Exogenous ABA increased the drought tolerance compared with plant without ABA treatment was demonstrated previously (Huang et al., 2008).
GSH’s Effects on the Stomata Aperture Size and Root Patterning
It was interesting to find that the constitutive stomatal aperture was smaller when the WT plants were grown in a solution containing GSH a situation similar to smaller stomatal aperture in atgstu17 mutants (Fig. B and C). This probably is an effect of ABA because plants growing in the GSH solution exhibited greater ABA content (Fig.
15).
The astonishing root architecture might have been due to higher levels of GSH accumulation in the loss-of-function atgstu17 plants, which is consistent with high levels of endogenous GSH enhancing cell division in the root meristematic region leading to root elongation (Vernoux et al., 2000), and exogenous GSH on auxin-induced in vitro root formation (Imin et al., 2007). Low concentrations of GSH (< 50 μM) enhanced primary root elongation, but high concentrations suppressed root growth (Fig. 17). Our observation indicates the effect of an exogenous GSH in modulating the root growth pattern of WT plants, and can mimic the phenotype in the
atgstu17 plants. The effect of ABA on the root architecture differed from that of GSH.
When applied exogenously in a well-watered condition, ABA acts as a growth inhibitor to suppress primary root growth (Sharp et al., 2004; Bai et al., 2009) while drought stress inhibits lateral root development of soil-grown plants (Xiong et al., 2006). ABA treatment reduced primary root length but encouraged lateral root growth
(Fig. 17). It was apparent that the root systems in the atgstu17 plants reflected the effects of different combinations of GSH and ABA.
Our findings support a model as summarized in Figure 24 whereby repressed
AtGSTU17 expression is linked to the accumulation of GSH which associates with the
accumulation of ABA. The phenotypes of atgstu17 can be mostly attributed to the combined effects of GSH and ABA, which explain the molecular mechanism of the repression of AtGSTU17 gene expression in modulating ABA sensitivity of seed germination, stomatal aperture, root architecture, and drought- and salt-stress tolerance. There must be some mechanism that regulates or represses the expression of AtGSTU17 in stressful conditions. If this did not occur in the Col-0 background,AtGSTU17 would be a target of natural selection in order for plant to adapt to adverse
environments. The link between genotypes and the expressions of specific GST genes was found in wheat which differed in drought tolerance (Galle et al., 2009). Thus in the whole plant, the repression of AtGSTU17 may play a role of find-tuning GSH homeostasis, redox status and stress-responsive genes in adaptation to changes in environmental signals.REFERENCE
Bai L, Zhang G, Zhou Y, Zhang Z, Wang W, Du Y, Wu Z, Song C-P (2009)
Plasma membrane-associated proline-rich extensin-like receptor kinase 4, a novel regulator of Ca2+ signalling, is required for abscisic acid responses in Arabidopsisthaliana. Plant J 60: 314–327.
Chen IC, Huang IC, Liu MJ, Wang ZG, Chung SS, Hsieh HL (2007) Glutathione
S-transferase interacting with far-red insensitive 219 is involved in phytochrome A-mediated signaling in Arabidopsis. Plant Physiol 143: 1189-1202Chen J, Goldsborough PB (1994) Increased activity of γ-glutamylcysteine
synthetase in tomato cells selected for cadmium tolerance. Plant Physiol 106:233–239.
Cheng WH, Endo A, Zhou L, Penney J, Chen HC, et al. (2002) A unique
short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 14: 2723–2743.Cummins L, Cole DJ, Edwards R (1999) A role for glutathione transferases
functioning as glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant J 18: 285-292.De Smet I, Zhang H, Inze D, Beeckman T (2006) A novel role for abscisic acid
emerges from underground. Trends Plant Sci 11: 434-439.Deng Z, Zhang X, Tang W, Oses-Prieto JA, Suzuki N, Gendron JM, Chen H, Guan S, Chalkley RJ, Peterman TK, Burlingame AL, Wang Z (2007) A
proteomic study of brassinosteroid response in Arabidopsis.Mol Cell Proteomics. 6:2058–2071.
Dixon DP, Hawkins T, Hussey PJ, Edwards R (2009) Enzyme activities and
subcellular localization of members of the Arabidopsis glutathione transferase superfamily. J Exp Bot 60: 1207-1218Edwards R, Dixon DP (2005) Plant glutathione transferases. Methods Enzymol 401:
169–18
Edwards R, Dixon DP, Walbot V (2000) Plant glutathione S-transferases: enzymes
with multiple functions in sickness and in health. Trends Plant Sci 5: 193-196Foyer CH, Noctor G (2005) Redox homeostasis and antioxidant signaling: a
metabolic interface between stress perception and physiological responses. PlantCell, 17, 1866–1875.
Foyer CH, Noctor G (2009) Redox regulation in photosynthetic organisms: signaling,
acclimation, and practical implications. Antiox Red Signa 11: 861-905.Foyer CH, Souriau N, Perret S, Lelandais M, Kunert K-J, Pruvost C, Jouanin L
(1995) Overexpression of glutathione reductase but not glutathione synthetase leads to increases in antioxidant capacity and resistance to photoinhibition in poplar trees.Plant Physiol 109: 1047–1057.
Furihata T, Maruyama K, Fujita Y, Umezawa T, Yoshida R, et al. (2006)
Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc Natl Acad Sci USA 103: 1988–1993.Fujii H, Verslues PE, Zhu JK (2007) Identification of two protein kinases required
for abscisic acid regulation of seed germination, root growth, and gene expression in Arabidopsis. Plant Cell 19: 485–494.Gallé A, Csiszár J, Secenji M, Guóth A, Cseuz L, Tari I, Györgyey J, Erdei L
(2009) Glutathione transferase activity and expression patterns during grain filling in flag leaves of wheat genotypes differing in drought tolerance: response to water deficit. J. Plant Physiol 166: 1878-1891Grant CM, MacIver FH, Dawes IW (1996) Glutathione is an essential metabolite
required for resistance to oxidative stress in the yeast Saccharomyces cerevisiae.Current Genetics 29: 511–15.
Griffith OW (1980) Determination of glutathione and glutathione disulfide using
glutathione reductase and 2-vinylpyridine. Anal Biochem 106: 207–212.Hsu YT, Kao CH (2003) Role of abscisic acid in cadmium tolerance of rice
(Oryza sativa L.) seedlings. Plant Cell Environ 26: 867–874Huang D, Wu W, Abrams SR, Cutler AJ (2008) The relationship of drought-related
gene expression in Arabidopsis thaliana to hormonal and environmental factors. J Exp Bot 59: 2991–3007.Imin N, Nizamidin M, Wu T, Rolfe BG (2007) Factors involved in root formation in Medicago truncatula. J Exp Bot 58: 439–451.
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-glucuronidase
as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901-3907
Ji W, Zhu Y, Li Y, Yang L, Zhao X, Cai H, Bai X (2010) Over-expression of a
glutathione S-transferase gene, GsGST, from wild soybean (Glycine soja) enhances drought and salt tolerance in transgenic tobacco. Biotechnol Lett 32: 1173-1179.Jiang HW, Liu MJ, Chen IC, Huang CH, Chao LY, Hsieh HL (2010) A
glutathione S-transferase regulated by light and hormones participates in the modulation of Arabidopsis seedling development. Plant Physiol 154: 1646-1658.Kim MJ, Shih R, Schachtman DP (2009) A nuclear fctor regulates abscisic acid
responses in Arabidopsis. Plant Physiol151: 1433–1445.
Kitamura S, Shikazono N, Tanaka A (2004) TRANSPARENT TESTA 19 is involved
in the accumulation of both anthocyanins and proanthocyaniins in Arabiopsis. Plant J 37: 104-114Ko JH, Yang SH, Han KH (2006) Upregulation of an Arabidopsis RING-H2 gene, XERICO, confers drought tolerance through increased abscisic acid biosynthesis.
Plant J 47: 343–355
Kocsy G, von Ballmoos P, Suter M, Ruegsegger A, Galli U, Szalai G, Galiba G, Brunold C (2000) Inhibition of glutathione synthesis reduces chilling tolerance in
maize. Planta 211: 528–536.Kocsy G, von Ballmoos P, Ruegsegger A, Szalai G, Galiba G, Brunold C (2001)
Increasing the glutathione content in a chilling-sensitive maize genotype using safeners increased protection against chilling-induced injury. Plant Physiol 127:1147–1156.
Konopka-Postupolska D, Clark G, Goch G, Debski J, Floras K, Cantero A, Fijolek B, Roux S, Hennig J (2009) The role of Annexin 1 in drought stress in Arabidopsis. Plant Physiol 150: 1394–1410
Kushnir S, Babiychuk E, Kampfaenkel K, Belles-Boix E, Van Montagu M, Inze D (1995) Characterisation of Arabidopsis thaliana cDNAs that render yeasts
tolerant toward the thiol-oxidising drug diamide. Proc Natl Acad Sci USA 92:10580–10584.
Lai B, Nadeau JA, Lucas J, Lee EK, Nakagawa T, et al. (2005) The Arabidopsis
R2R3 MYB proteins FOUR LIPS and MYB88 restrict divisions late in the stomatal cell lineage. Plant Cell 17: 2754–2767.Lappartient AD, Touraine B (1997) Glutathiowe-mediated regulation of ATP
sulfurylase activity, SO42- uptake, and oxidative stress response in intact canola
roots. Plant Physiol 114: 177-183.Lin RC, Park HJ, Wang HY (2008) Role of Arabidopsis RAP2.4 in regulating light
and ethylene-mediated developmental processes and drought stress tolerance. Mol Plant 1: 42 -57Loggini B, Scartazza A, Brugnoli E, Navari-Izzo F (1999) Antioxidative defense
system, pigment composition, and photosynthetic efficiency in two wheat cultivars subjected to drought. Plant Physiol 119: 1091–1100.Loyall L, Uchida K, Braun S, Furuya M, Frohnmeyer H (2000) Glutathione and a
UV light-induced glutathione S-transferase are involved in signalling to chalcone synthase in cell cultures. Plant J 25: 237–245.Marrs KA (1996) The functions and regulation of glutathione S-transferases in plants.
Annu Rev Plant Physiol Plant Mol Biol 47: 127-158.
Marrs KA, Walbot V (1997) Expression and RNA splicing of the maize glutathione
S-transferase Bronze2 gene is regulated by cadmium and other stresses. Plant Physiol 113: 93-102.Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J (2001) The ABI1and ABI2
protein phosphatases 2C act in a negative feedback regulatory loop of the abscisic acid signalling pathway. Plant J 25: 295–303.Meyer JM (2008) The integration of glutathione homeostasis and redox signaling. J.
Plant Physiol 165: 1390-1403.
Moons A (2003) Osgstu3 and osgtu4, encoding tau class glutathione S-transferases,
are heavy metal- and hypoxic stress-induced and differentially salt stress-responsive in rice roots. FEBS Letters 553: 427–432Rivero RM, Kojima M, Gepstein A, Sakakibara H, Mittler R, Gepstein S, Blumwald E (2007) Delayed leaf senescence induces extreme drought tolerance in
a flowering plant. Proc Natl Acad Sci USA 104: 19631–19636.Rouhier N, Lemaire SD, Jacquot J (2008) The role of glutathione in photosynthetic
organisms: emerging functions for glutaredoxins and glutathionylation. Annu Rev Plant Biol 59: 143–66.Roxas VP, Lodhi SA, Garrett DK, Mahan JR, Allen RD (2000) Stress tolerance in
transgenic tobacco seedlings that overexpress glutathione S-transferase/ glutathione peroxidase. Plant Cell Physiol 41: 1229–1234Sappl PG, Carroll AJ, Clifton R, Lister R, Whelan J, Millar AH, Singh KB (2009)
The Arabidopsis glutathione transferase gene family displays complex stress regulation and co-silencing multiple genes results in altered metabolic sensitivity to oxidative stress. Plant J 58: 53–68.
Sharp RE, LeNoble ME (2002) ABA, ethylene and the control of shoot and root
growth under water stress. J Exp Bot 53: 33-37.Sharp RE, Poroyko V, Hejlek LG, Spollen WG, Springer GK, Bohnert HJ, Nguyen HT (2004) Root growth maintenance during water deficits: physiology to
functional genomics. J Exp Bot 55: 2343–2351.Sheehan D, Meade G, Foley VM, Dowd CA (2001) Structure, function and
evolution of glutathione S-transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily. Biochem J 360: 1-16.Tausz M, Pilch B, Rennenberg H, Grill D, Herschbach C (2004) Root uptake,
transport, and metabolism of externally applied glutathione in Phaseolus vulgaris seedlings. J. Plant Physiol 161: 347–349.Tepperman JM, Zhu T, Chang HS, Wang X, Quail PH (2001) Multiple
transcription-factor genes are early targets of phytochrome A signaling. Proc Natl Acad Sci USA 98: 9437-9442Thompson AJ, Andrews J, Mulholland BJ, McKee JMT, Hilton HW, Horridge JS, Farquhar GD, Smeeton RC, Smillie IRA, Black CR, Taylor IB (2007)
Overproduction of abscisic acid in tomato increases transpiration efficiency and root hydraulic conductivity and influences leaf expansion. Plant Physiol 143:1905–1917.
Vanderauwera S, Block MD, Van de Steene N, van de Cotte B, Metzlaff M, Van Breusegem F (2007) Silencing of poly(ADP-ribose) polymerase in plants alters
abiotic stress signal transduction. Proc Natl Acad Sci USA 104: 15150–15155.Vernoux T, Wilson RC, Seeley KA, Reichheld JP, Muroy S, Brown S, Maughan
SC, Cobbett CS, Van Montagu M, Inzé D, May MJ, Sung ZR (2000) The ROOT MERISTEMLESS1/CADMIUM SENSITIVE2 gene defines a
glutathione-dependent pathway involved in initiation and maintenance of cell division during postembryonic root development. Plant Cell 12: 97-110.Wellburn FAM, Creissen GP, Lake JA, Mullineaux PM, Wellburn AR (1998)
Tolerance to atmospheric ozone in transgenic tobacco over-expressing glutathione synthetase in plastids. Physiol Plant 104: 623–629.Wilson PB, Estavillo GM, Field KJ, Pornsiriwong W, Carroll AJ, Howell KA, Woo NS, Lake JA, Smith SM, Millar AH, von Caemmerer S, Pogson BJ (2009)
The nucleotidase/phosphatase SAL1 is a negative regulator of drought tolerance in Arabidopsis. Plant J 58: 299-317.Xiong L, Wang R-G, Mao G, Koczan GM (2006) Identification of drought
tolerance determinants by genetic analysis of root response to drought stress and abscisic acid. Plant Physiol 142: 1065–1074.Yu H, Chen X, Hong Y-Y, Wang Y, Xu P, Ke S-D, Liu H-Y, Zhu J-K, Oliver DJ, Xiang C-B (2008) Activated expression of an Arabidopsis HD-START protein
confers drought tolerance with improved root system and reduced stomatal density.Plant Cell 20: 1134–1151.
Zhu J, Fu X, Koo YD, Zhu JK, Jenney FE Jr, Adams MW, Zhu Y, Shi H, Yun DJ, Hasegawa PM, Bressan RA (2007) An enhancer mutant of Arabidopsis salt overly sensitive 3 mediates both ion homeostasis and the oxidative stress response.
Mol Cell Biol 27: 5214–5224.