國立臺灣大學生命科學院植物科學研究所 碩士論文
Institute of Plant Biology College of Life Science National Taiwan University
Master Thesis
ERF19 和 NINJA 抑制複合體參與阿拉伯芥抗灰霉病的 功能性分析
Functional analysis of ERF19 and NINJA repressor complex involved in plant defense against Botrytis cinerea
蔣貝爾 Beier Jiang
指導教授﹕金洛仁 博士 Advisor: Laurent Zimmerli, Ph.D
中華民國 105 年 7 月 July 2016
誌謝
窗外的蟬鳴似乎在回應結束最後一個實驗后我這顆焦躁而空虛的內心。兩年 前剛進實驗室時,艱難地適應了世界各地的英語口音,日漸發現大家都是把 sh*t, f*ck 放在嘴邊的人,令我倍感欣慰,在這樣自由的環境里過的很輕鬆。
對每一位實驗室的成員都充滿感激,以下是肉麻的點名大告白。感謝我們的 老闆滷蛋申請到雄厚的經費讓大家肆意燃燒,雖然大家經常罵他,但是我還是覺 得他是個很有趣又大方的人;我的小老闆品堯學長兢兢業業地教我很多很多實驗 思路和方法還要督促我不要懶惰要早去實驗室雖然我無法做到;鈺鴻學長每天都 在開黃腔以及言語性騷擾大家因此在這種強壓之下我練就了一身反黃腔及性騷擾 的能力想必日後定會派上用場;來自香港的運動選手陳晶和我分享旅行見聞;達 哥每次都會很熱情的回答任何問題雖然有時候我聽不懂;太一一直安慰鼓勵我碩 一那個題目做得出來讓我意識到不要一味的相信好朋友即使他很聰明;友慧姐姐 一直誇我可愛差點讓我自己都相信了其實妳最可愛了啦;富淳是我見過最最認真 細緻的女生多虧了妳讓我克服了對軟體小蟲兒的恐懼;好姐妹好麻吉老麥姐姐是 我最大的八卦來源讓我的實驗生活時刻有驚喜與驚嚇;高冷學弟義竣極大的提升 了我們家的平均顏值是我來實驗室的精神動力每次在我耍寶說笑話的時候很配合 的大笑是對我這個搞笑藝人的肯定;句點王冠宇想法獨特能懂我的笑點還會接大 家無法繼續的梗;中合雖然來得時間不長卻已然被我們污染是個可教之才。除了 實驗室的夥伴,浙滬美少女們也是我堅持下去的最大動力,燊琪聞喆華仔還有每 天和我告白一百遍的室友奚夢源,吃喝玩樂是你們溜。我的最最好朋友金今大哥 在电网上班非常不認真每天都秒回我的微信陪我嘮嗑。最感謝爸爸一直支持我的 所有選擇,看到你白頭髮越來越多真的很難過因為我也遺傳到這一點。
結束也是新的開始,不說了我要去吃飯了。
摘要
植物在受到病原菌攻擊時會啟動免疫反應從而引起一系列免疫基因的表達。
這個過程需要非常精細的轉錄水平上的重新編碼。舉例來說,乙烯調控因子在植 物調節免疫反應中有著重要的作用。在我們實驗室之前的研究中發現,阿拉伯芥 過表達 ERF19 基因時表現出比野生型更加容易被灰霉病菌感染的性狀。這種性狀 是通過抑制免疫基因 PDF1.2a 以及 PR3 的表達所導致的。在我的研究中,構建了 ERF19 的功能缺失植株,發現它們表現出比野生型更加抗灰霉病菌感染的性狀。
通過原生質體瞬時表達實驗發現 ERF19 是轉錄激活子而 ERF19 過表達植株中的 免疫基因被抑制。於是我通過酵母雙雜交,蛋白質免疫共沉澱以及雙分子螢光光 互補實驗發現 ERF19 蛋白可以和 NINJA 抑制複合體有交互作用。更有趣的是,
在 ninja 突變體中過表達 ERF19 蛋白會嚴重改變阿拉伯芥的生長表型以及抗病表 型。所有這些新的發現對於研究 ERF 轉錄因子參與植物免疫反應有嶄新的意義。
關鍵字:阿拉伯芥;灰黴菌;乙烯反應因子;NINJA 抑制子;自然免疫
Abstract
When attacked by pathogens, plants activate the immune responses to regulate the inducible expression of a large set of defense genes. This process requires effective transcriptional reprogramming. For example, ethylene responsive factor (ERF)
transcription factors (TFs) have been shown to play critical roles in regulating immune responses in plants. In the Zimmerli laboratory, overexpression of ERF19 was shown to increase Arabidopsis susceptibility to Botrytis cinerea (B. cinerea) via repression of the induction of the jasmonic acid (JA) defense marker genes PDF1.2a and PR3. In this study, I generated pERF19:ERF19-EAR transgenic lines and these loss-of-function plants exhibited increased resistance to B. cinerea, when compared to wild type (WT) plants. ERF19 acted as a transcriptional activator, however defense genes were
repressed in ERF19 overexpression lines. Using the yeast two hybrid, Co-IP and BiFC assays, ERF19 was shown to associate with the NINJA repressor complex. Moreover, overexpression of ERF19 in Arabidopsis ninja mutant strongly changes the
developmental pattern and defense phenotype of the plant. Notably, a strong increased susceptibility to B. cinerea was observed. All these findings provide new insight on how ERF TFs regulate plant immunity.
Key-words: Arabidopsis thaliana; Botrytis cinerea; ethylene responsive factor; NINJA repressor complex; innate immunity
Contents
誌謝 ... i
摘要 ... ii
Abstract ... iii
Contents ... iv
Introduction ... 1
Arabidopsis defenses against B. cinerea ... 1
Pathogen-associated molecular patterns-triggered immunity and effector- triggered immunity ... 1
Chitin-triggered immunity ... 2
ERF transcription factors ... 3
Roles of ERF proteins in biotic stress ... 4
The role of NINJA in JA signaling ... 4
EAR motif-mediated transcriptional repression in plants ... 5
Mechanism of action of the co-repressor TPL ... 6
HDACs participate in the defense response ... 6
Materials and Methods ... 8
Plant materials and growth conditions ... 8
Transgenic plants ... 8
Pathogen infection assay ... 9
β-estradiol induction before pathogen infection ... 9
PAMP treatment ... 9
Genomic DNA extraction ... 10
RNA extraction and gene expression analysis ... 10
Protoplast transactivation assay ... 11
Transient expression in Nicotiana Benthamiana ... 11
Subcellular localization in Nicotiana benthamiana leaves ... 12
Yeast two-hybrid assays ... 12
Bimolecular Fluorescence Complementation assays ... 12
Co-immunoprecipitation assay in Arabidopsis protoplast ... 13
SDS-PAGE and Immunoblotting ... 13
Accession numbers ... 14
Results ... 15
Loss of ERF19 function is linked with enhanced resistance to B. cinerea ... 15
pERF19:ERF19-SRDX transgenic lines display enhanced expression of defense gene ... 15
Generation of Arabidopsis ERF19 artificial microRNA transgenic lines ... 16
Subcellular localization of ERF19 in Nicotiana benthamiana ... 17
ERF19 directly interacts with NINJA in vitro ... 17
ERF19 associates with the NINJA repressor complex in vivo ... 18
ERF19 associates with HDA6 and HDA19 in vivo ... 18
Arabidopsis ninja mutant exhibit enhanced resistance while NINJA overexpression lines are more susceptible to B. cinerea ... 19 Overexpress ERF19 in Arabidopsis ninja mutant showed altered developmental
phenotype ... 20
Lines overexpressing ERF19 in the ninja mutant exhibits strong increased susceptibility to B. cinerea... 20
Discussion ... 21
Future Perspectives ... 24
Figures ... 25
Figure 1. Disease responses of pERF19:ERF19-SRDX transgenic plants to B. cinerea infection. ... 25
Figure 2. Response of ERF19 to amiRNA. ... 27
Figure 3. Subcellular localization of ERF19. ... 28
Figure 4. Direct interaction of ERF19 with NINJA but not with TPL, HDA6 or HDA19 by Y2H ... 29
Figure 5. ERF19 associates with TPL in vivo. ... 30
Figure 6. Association of ERF19 with NINJA, HDA6 and HDA19 but not with TPL by BiFC ... 31
Figure 7. Disease responses of ninja mutant and NINJA overexpression plants to B. cinerea infection. ... 32
Figure 8. Development and defense phenotype of ninja mutant overexpressing ERF19. ... 33
Figure 9. Model for ERF19-mediated defense repression in Arabidopsis. ... 34
Supplementary Figures ... 35 Figure S1. Constitutive and inducible overexpression lines of ERF19 were more
susceptible to B. cinerea infection than WT. ... 35
Figure S2. ERF19 associates with NINJA, HDA6 and HDA19 by Co-IP in Arabidopsis protoplast. ... 36
Figure S3. NINJA repress the activation of ERF19. ... 37
Tables ... 39
Table 1. Mutant alleles used in this study ... 39
Table 2. Primers used for ninja mutant check ... 39
Table 3. Primers used for ERF19 amiRNA plasmid construction ... 39
Table 4. Primers used for plasmid construction ... 40
Table 5. Primers used for qRT-PCR ... 40
Introduction Arabidopsis defenses against B. cinerea
B. cinerea is considered to be the second most important fungal plant pathogen (Dean et al., 2012). B. cinerea is a necrotrophic pathogen that can infect more than 200 plant species by killing the host cells and then extracting nutrients from the dead host cells. When attacked by B. cinerea, about one-third of the Arabidopsis genome is turned up or down during the first 48 h after infection, before the development of significant lesion (Windram et al., 2012). This differentially gene expression is regulated by a set of TFs. Two major groups of TFs involved in defense against B. cinerea are the WRKY (Eulgem and Somssich, 2007) and APETALA2 (AP2)/ERF families (Xu et al., 2011).
WRKY and ERF TFs function in a diverse array of plant responses, including plant development, biotic and abiotic stresses, hormone signaling, primary and secondary metabolisms. WRKY3, 4, 8, 18, 33, 40, 60, and 70 have positive or negative roles in B.
cinerea immunity (Windram et al., 2012). For ERFs, ERF1, ERF5, ERF6, RAP2.2, ORA59 and ERF96 influence host susceptibility to B. cinerea (Berrocal‐Lobo et al., 2002; Pré et al., 2008; Moffat et al., 2012; Son et al., 2012; Zhao et al., 2012; Huang et al., 2016).
Pathogen-associated molecular patterns-triggered immunity and effector-triggered immunity
Pathogen-associated molecular patterns (PAMP)-triggered immunity (PTI) and effector-triggered immunity (ETI) are well-defined types of plant immunity acting against pathogens. PTI is the first line of defense the host activates to resist microbial infection.
The pattern-recognition receptors (PRRs) localized at the plasma membrane recognize PAMPs and trigger downstream defense signaling pathways. Classical examples of
PAMPs are structural molecules, such as bacterial flagellin, peptidoglycan, and lipopolysaccharides, oomycete glucans, and fungal chitin (Thomma et al., 2011). PTI is characterized by a wide range of physiological responses, including production of reactive oxygen species (ROS), mitogen activated protein kinase (MAPK) phosphorylation, calcium influx, ion channel activation, callose deposition, growth inhibition, and induction of defense-related genes (Macho and Zipfel, 2014). Successful pathogens are able to overcome PTI by secreting effectors into host cells, which can suppress PTI responses, resulting in effector-triggered susceptibility. Pathogenic bacteria typically inject such effectors directly into the host cytoplasm by means of their type III secretion machinery. Plants have also evolved a system to fight against effectors through the cytoplasmic resistance (R) proteins that recognize single effectors. The majority of these R proteins are intracellular receptor proteins of the nucleotide binding–leucine-rich repeat (NB-LRR) type that can activate ETI.
Chitin-triggered immunity
Multiple PAMPs are involved in the interaction between B. cinerea and Arabidopsis.
In this thesis, we focused on the essential fungal cell wall component, chitin, a β-(1,4)- linked homopolymer of N-acetylglucosamine, and its constituent oligosaccharides. Chitin is a fungal PAMPs that activate numerous defense responses (Felix et al., 1993). The chitin receptor was first reported in rice with the identification of the chitin-elicitor binding protein (CEBiP) (Kaku et al., 2006). OsCEBiP forms a complex with the rice chitin-elicitor receptor kinase 1 (OsCERK1) to mediate rice PTI in response to chitin.
Similarly, in Arabidopsis, AtLYK5 is the primary receptor for chitin, forming a chitin inducible complex with AtCERK1 to induce plant immunity (Cao et al., 2014). In the latest study, AtLYK5 binds to chitin with a significantly higher affinity than AtCERK1.
In the absence of chitin, AtLYK5 exists in the cell as a homodimer form. Upon chitin binding, AtLYK5 dimerizes with AtCERK1 and activates its kinase activity, resulting in downstream chitin signal transduction (Cao et al., 2014).
ERF transcription factors
TFs are proteins that control the expression of downstream target genes by binding to specific DNA sequences. Proteins in the AP2/ERF superfamily have one or two conserved 60-amino-acid domain called the AP2/ERF domain (Nakano et al., 2006).
Depending on how many AP2 domains or other domains they possess, members of the AP2/ERF superfamily are classified into four subfamilies, ERF family, AP2 family, RAV family and Soloist (Licausi et al., 2013). This AP2/ERF domain can bind to the GCC box, a conserved motif present in the promoter regions of genes that respond to JA, ethylene (ET), pathogens and wounding stress (Ohme-Takagi and Shinshi, 1995; Brown et al., 2003). As transcription factors, ERF proteins can act as activators or repressors. For example, transient expression analyses in Arabidopsis leaves revealed that AtERF1, ERF2 and ERF5 function as activators of GCC box-mediated transcription, whilst ERF3, ERF4 and ERF7 act as repressors. The EDLL motif, which has a unique arrangement of acidic amino acids and hydrophobic leucines, was shown to be a strong activation domain and a useful tool to confer transcriptional activation potential to heterologous DNA- binding proteins (Tiwari et al., 2012). In contrast, ERF repressors also have specific repressor domains, the ERF-associated amphiphilic repression (EAR: LxLxLx or DLNxxP) motif (Ohta et al., 2001b), the TLLLFR motif (Matsui et al., 2008) and the B3 repression domain (BRD: RLFGV) (Ikeda and Ohme-Takagi, 2009). ERFs that contain an EAR- or BRD-motif suppress the transcription of target genes by recruiting the co- repressors TOPLESS (TPL) or TOPLESS-RELATED (TPR) (Causier et al., 2012b).
Roles of ERF proteins in biotic stress
ERF proteins were originally discovered as TFs that bind to the promoter regions of stress-responsive genes. To date, ERF proteins are known to be involved in biotic and abiotic stress responses, such as pathogen infection, salt stress, osmotic stress, wounding, drought, hypoxia, temperature stress and the signaling of stress-related hormones ET, JA and ABA.
Several ERF transcriptional activators induce enhanced disease resistance when overexpressed and enhanced susceptibility when mutated. For example, overexpression of the activators ERF1 and ERF2 in Arabidopsis induces the up-regulation of the defense genes PDF1.2 and PR3, and increased resistance to the necrotrophic pathogen Fusarium oxysporum (McGrath et al., 2005). Similarly, T-DNA insertion mutant of the transcriptional activator ERF14 shows lower PDF1.2 and PR3 gene expression and enhanced susceptibility to F. oxysporum infection (Oñate-Sánchez et al., 2007). On the contrary, overexpression lines of the repressor ERF4 are more sensitive to F. oxysporum infection while the mutant of ERF4 is more resistant (McGrath et al., 2005). In addition, overexpression of the activator ERF96 in Arabidopsis enhances the expression of the defense genes PDF1.2a, PR-3 and PR-4 as well as the TF ORA59 by directly binding to GCC elements present in their promoters (Catinot et al., 2015).
The role of NINJA in JA signaling
To respond to biotic and abiotic stresses, plants accumulate JAs as a signal hormone.
JA also regulate diverse processes such as pollen maturation and wound responses in Arabidopsis. In the absence of JA–isoleucine (JA–Ile), JA ZIM domain (JAZ) proteins block TFs such as MYC2 by directly interacting with them to repress JA-responsive gene
expression and anthocyanin biosynthesis (Chini et al., 2007). JAZ proteins activate this process through interaction with co-repressor complexes containing the Novel Interactor of JAZ (NINJA) and TOPLESS. When JA-Ile accumulates, the bioactive hormone binds the CORONATINE INSENSITIVE 1 (COI1) receptor in the Skp–Cullin–F-box-type E3 ubiquitin ligase (SCFCOI1) complex, thereby recruiting the JAZ proteins, targeting them for their degradation by the 26S proteasome, and ultimately leading to the release of the TFs that can modulate the expression of JA-responsive and anthocyanin synthesis genes (Chini et al., 2009).
EAR motif-mediated transcriptional repression in plants
EAR motif-mediated transcriptional repression is one of the principal mechanisms of plant gene regulation. EAR motifs directly interact with co-repressors, such as TPL and SAP18, to function as negative regulators in a broad range of developmental and physiological processes. SAP18 directly interacts with HDA19 and TPL function in conjunction with HDA19 to suppress the expression of target genes through chromatin modification (Song and Galbraith, 2006b). Chromatin modification is one of the principal mechanism that regulate gene expression in both animals and plants (Thiel et al., 2004).
Among them, histone deacetylation plays an important role in transcriptional repression.
For instance, the EAR motif containing class II ERFs, such as ERF3 and ERF4, which are known to function as active repressors in vitro and in vivo, have been shown to physically interact with AtSAP18, which in turn interacts and forms a repression complex with AtHDA19 (Ohta et al., 2001a; Yang et al., 2005; Song and Galbraith, 2006a). NINJA, which contains an LxLxL type of EAR motif in its amino-terminal region and functions as a negative regulator of JA responses, physically interacts with TPL in an EAR motif- dependent manner.
Mechanism of action of the co-repressor TPL
Co-repressors are transcriptional regulators that are incapable of independent DNA binding. Instead, they are recruited directly or indirectly by DNA-binding TFs to repress target gene expression. It is a very common mechanism to down-regulate gene expression in both animals and plants (Kao et al., 1998; Liu and Karmarkar, 2008). In animals, yeast and fungi, Groucho (Gro)/Tup1 family proteins are known as the most important co- repressors by repressing genetic loci in chromatin (Liu and Karmarkar, 2008). In plants, three families of Gro/Tup1 co-repressors have been identified, including LEUNIG (LUG), TOPLESS (TPL) and WUSCHEL-INTERACTING PROTEINS (WSIPs). These co- repressors are involved in a broad range of developmental processes. TPL and TPL- RELATED (TPR) proteins participate in the regulation of embryo development, auxin and JA signaling, plant immunity and meristem fate (Causier et al., 2012a). Current evidence suggests that the TPL/TPR family mediates transcriptional repression by acting with histone deacetylases to induce a repressive chromatin state at the target locus. And it has been reported that TPL associates with HDA6 and HDA19 to inhibit the expression of target genes (Zhu et al., 2010; Wang et al., 2013).
HDACs participate in the defense response
Defences against necrotrophic pathogens are mainly connected with the JA/ET pathway (Bari and Jones, 2009). The stimulation of ETI and PTI involves extensive reprogramming of plant transcription. And this process is well regulated by histone modification including histone acetylation (Ding and Wang, 2015). Histone acetyltransferases (HATs) and histone deacetylases (HDACs) determine the acetylation level of lysine residues on histone tails. While HATs are associated with gene activation,
HDACs are usually mediating gene repression (Tian et al., 2005). For example, HDA19 is a nuclear RPD3/HDA1 family protein that has HDAC activity in vitro. Overexpression of HDA19 enhances fungal resistance through the apparent activation of the ERF1, whereas silencing of HDA19 increases fungal susceptibility (Zhou et al., 2005). HDA6 is another RPD3/HDA1 family protein which is involved in the activation of JA-dependent defences (Wu et al., 2008). Interestingly, HDA6 interacts with COI1, an F-box protein that mediates JA signalling (Devoto et al., 2002). This interaction seems to have functional significance, as HDA6 is required for JA-dependent responses, including PDF1.2, VSP2 and ERF1 expression (Wu et al., 2008).
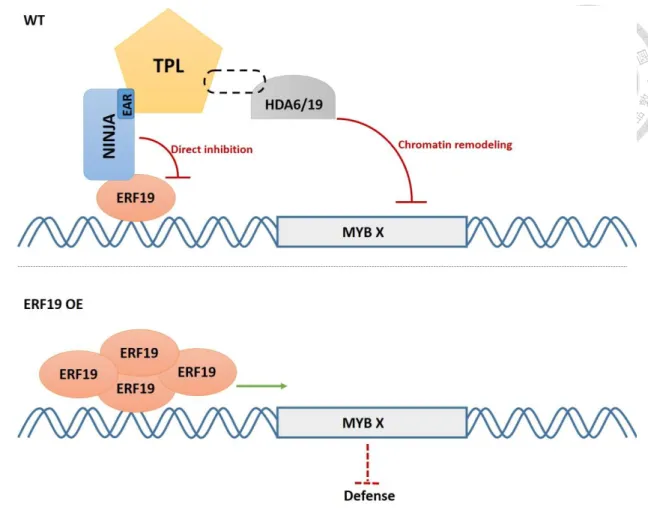
In order to discover new ERFs involved in the biotic stress response, previous members of the Zimmerli laboratory screened a pooled collection of Arabidopsis transgenic lines overexpressing HA-tagged ERF genes for their resistance to the necrotrophic fungal pathogen B. cinerea. They identified an ERF candidate, ERF19 (At1g22810) that when overexpressed in Arabidopsis triggers increased susceptibility to B. cinerea. However, how ERF19 regulate Arabidopsis defense responses is poorly understood. Here, I report on a molecular mechanism underlying the negative roles of ERF19 during the Arabidopsis defense response against B.cinerea. Specifically, I show that the NINJA repressor binds and inhibits the TF ERF19 through direct inhibition or recruitement of the co-repressor TPL. TPL in turn recruits HDACs to repress gene expression through chromatin remodeling. These findings provide new insight on how ERF TFs regulate plant immunity.
Materials and Methods Plant materials and growth conditions
All described mutants and transgenic lines are from Arabidopsis thaliana accession Columbia (Col) background. After seed stratification for 2 d at 4°C, plants were grown in commercial potting soil : perlite (3:2) at 22°C to 24°C day and 17°C to 19°C night temperatures under a 9-h-light/15-h-dark photoperiod. The lighting was supplied at an intensity of ~100 mE m–2 s–1 by fluorescent tubes. For seedling growth, seeds were surfaced-sterilized with 20% bleach solution for 5 min; then washed with sterile water four times to remove the bleach. Seedlings were grown on half-strength Murashige and Skoog solid medium (½ x MS, pH 5.7; Duchefa) supplemented with 0.5 g/L of MES hydrate (Bioman) and 0.7% (w/v) agar.
Transgenic plants
To generate inducible ERF19 microRNA lines, the XVE system was applied for β-estradiol-inducible expression. The gateway compatible binary pMDC7 vector (obtained from Ueli Grossniklaus, University of Zürich) was used for inducible expression. The ERF19 artificial microRNA (amiRNA) sequence was introduced into pMDC7 by LR reaction. ERF19 amiRNA sequence was identified using the online amiRNA design tool WMD3, a web application for the automated design of amiRNA (http://wmd3.weigelworld.org/). The construct was transferred to A. tumefaciens GV3101 in order to perform transformation of WT Arabidopsis by floral dipping.
Homozygote T3 Arabidopsis lines were selected on ½ MS solid medium plate using hygromycin B (Bioman Scientific Co., New Taipei, Taiwan) at the concentration of 15 μg/mL. T2 seeds of ERF19-SRDX driven by ERF19 native promoter were obtained from Nobutaka Mitsuda group in Japan (Saitama University). T3 lines were obtained from T2 seeds according to the 3:1 (Hgr-resistant/Hgr-sensitive) segregation ratio. For
ninja mutant overexpressing ERF19 transgenic plants, A. tumefaciens GV3101 containing pEG103-ERF19 was used for the transformation of ninja mutant plants. T3 lines were obtained from T2 seeds according to the 3:1 (Basta-resistant/Basta-sensitive) segregation ratio. Transgenic plants were also confirmed according to the GFP protein level extracted from leaves.
Pathogen infection assay
The fungus B. cinerea was obtained from C.Y. Chen (National Taiwan University) and grown at room temperature (18°C to 25°C) on potato dextrose broth (PDB) agar plates. For B. cinerea infection, spores were diluted to 105 spores/mL in half-strength PDB medium. Droplets (8 mL) of half-strength PDB with B. cinerea spores were deposited on leaf surfaces of 4-week-old plants (three leaves per plant). Leaves of the same age were used for droplet inoculation. Disease symptoms and lesion perimeters were determined at 2 or 3 DPI. At least 24 lesion diameters were evaluated for each biological replicate (eight plants).
β-estradiol induction before pathogen infection
A stock of 20 mM β-estradiol (Sigma-Aldrich) in 100% dimethyl sulfoxide (DMSO) was made and stored at −20°C in small aliquots (β-estradiol is light sensitive and its activity slowly declines in a light intensity-dependent manner). The DMSO alone has no effect on transgene expression. One day before pathogen infection, three leaves of the plant were induced by applying β-estradiol solution with an artist's paintbrush to sectors of the plant. For infection of β-estradiol induced transgenic plants, the B. cinerea PDB solution contained 20 μM (final concentration) β-estradiol.
PAMP treatment
For PDF1.2a expression, seedlings grown on ½ MS plates for 10 days were
transferred to liquid ½ MS medium one night before the addition of 20 mg/mL (final concentration) chitin from shrimp shells (Sigma-Aldrich), and samples for RNA extraction were collected at indicated time points.
Genomic DNA extraction
One rosette leaf from Arabidopsis plants was dipped into 50 μL DNA extraction buffer (10 mM Tris pH 8.2, 10 mM EDTA, 200 mM NaCl, 0.5% SDS). After grinded by micro-pestle, the solution was heated at 100℃ for 10 min. Then the solution was diluted in 1 mL sterile RO water. Two uL diluted solution was used for PCR analysis to confirm the three ninja mutant. The PCR products were then sequenced by DNA sequencing service providers.
RNA extraction and gene expression analysis
Total RNA was extracted and purified using the TriZol reagent according to the manufacturer's instructions (Invitrogen). Genomic DNA contaminations were removed using Qiagen RNase-Free DNase Set. For cDNA synthesis, 2 μg of total RNA were prepared in a volume of 22 μL DEPC-treated H2O and denatured at 65°C for 5 min.
Eighteen point 5 mL of master mix (16 μL M-MLV buffer, 1 mM dNTP, 5 mM OligoT, 100 U M-MLV reverse transcriptase, [Invitrogen]) was added into each tube and then incubated at 37°C for 1 h, 70°C for 10 min. The cDNA was then diluted 5 times before quantitative RT-PCR (qRT-PCR) analyses. qRT-PCR were conducted on a CFX Real- Time PCR Detection System (Bio-Rad). SYBR Green fast qPCR master mix (Bio-Rad;
1 μL of cDNA, 5 μL SYBR Green supermix, 5 μL filtered sterile H2O, 0.5 μL of 10 mM forward and reverse primers, in a total volume of 12 μL per well) was employed for the analysis. The cycling conditions were composed of an initial 3 min denaturation step at 95°C, followed by 40 cycles at 95°C for 3 s and 60°C for 30 s. Melting curve was run
from 65 to 95°C with 0.5-s time interval to ensure the specificity of product. Data were analyzed using Bio-Rad CFX manager software. UBQ10 was used as reference gene for normalization of gene expression levels in all samples. The WT without any treatment or mock treatment were considered as controls in each experiments (expression level = 1). Primer sequences are shown in Table 1.
Protoplast transactivation assay
Protoplasts were prepared from 4-week-old Arabidopsis leaves as described (Yoo et al., 2007), and co-transformed with a reporter plasmid carrying UAS-Luciferase (LUC) gene, effector plasmids carrying GUS, ERF96, ERF19 or ERF19-SRDX genes, and a reference plasmid carrying the RENILLA LUCIFERASE (REN) gene. The effector plasmid vector was pGWHA with a CaMV35S promoter. This plasmid was used to create the GUS, ERF96, ERF19 or ERF19-SRDX effector plasmids by LR reaction. One hundred μL of a suspension of 2.5 × 104 protoplast/mL was transfected with 10 μg of each of the 3 plasmids. After 18 h, the transfected protoplasts were harvested and the LUC assay was performed with the Dual-Luciferase Reporter Assay System (Promega, http://worldwide.promega.com/). Normalized LUC activity measured after transfection of the reporter UAS-LUC with GUS effector plasmid was set arbitrarily at 1.
Transient expression in Nicotiana Benthamiana
A. tumefaciens GV3101 strains carrying plasmids were grown overnight at 28°C in 5 mL LB medium with 2.5 μg/mL of rifampicin, 10 μg/mL of gentamycin and 50 μg/mL of kanamycin (the negative control A. tumefaciens GV3101 without binary vector was grown only with rifampicin). A. tumefaciens cells were harvested by centrifugation and re-suspended to OD600 0.4 in MMA induction buffer (5 g MS salts,
20 g sucrose, 1.95 g MES per liter, 200 μM acetosyringone, pH to 5.6 with 1M NaOH).
Bacterial suspensions were then incubated at room temperature for at least 3 h prior to infiltration through the abaxial surface into the leaf, using a syringe with no needle.
Subcellular localization in Nicotiana benthamiana leaves
For transient expression of GFP fusion proteins, the A. tumefaciens GV3101 containing the plasmid 35S-ERF19-GFP was infiltrated into 4-week-old N.
benthamiana leaves. GFP fluorescence was investigated 3 days after infiltration in excised leaf sections that included the infiltrated zone and 1-3 cm of surrounding non- infiltrated tissue using a confocal laser scanning microscope (LSM 780; Carl Zeiss).
Yeast two-hybrid assays
AD and BD constructs based on the GAL4 system were co-transformed into yeast strain AH109 using an LiAc-mediated yeast transformation protocol according to the manufacturer’s instructions (Clontech). At least 10 co-transformed yeast colonies were then plated on SD-His/Leu/Trp and SD-His/Ade/Leu/Trp plates supplemented with 3- amino-1,2,4-triazole (3-AT) (10mM) or X-α-galactosidase (X-α-Gal) (40 mg/L). The plates were incubated for 3 d at 30°C to test nutritional marker (His and Ade) gene expression and α-galactosidase activity of the MEL1 reporter protein.
Bimolecular Fluorescence Complementation assays
Full-length coding sequences of ERF19, NINJA, TPL, HDA6, HDA19 and GUS with and without stop codon amplified from cDNA of Arabidopsis Col-0 were inserted into entry vector pCR8/GW/TOPO and subcloned into YN (pDEST-VYNE and pDEST-VYNER) and YC (pDEST-VYCE and pDEST-VYCER) vector through the LR reaction. The constructs were transformed into N. Benthamiana by Agro infiltration for transient expression. YFP fluorescence was investigated 3 days after infiltration in
excised leaf sections that included the infiltrated zone and 1-3 cm of surrounding non- infiltrated tissue using a confocal laser scanning microscope (LSM 780; Carl Zeiss).
Co-immunoprecipitation assay in Arabidopsis protoplast
For Co-IP assays, the plasmids of 35S: ERF19-3HA (PCR amplified from pGBW14-ERF19 by using pGBW14-FP and pGBW14-RP, then cloned into pCR8/TOPO/TA cloning vector [Invitrogen]) and 35S: NINJA-GFP, 35S: TPL-GFP and 35S: GFP (modified from pEarlyGate103 with removing gateway cassette by using XhoI restriction enzyme) were transformed into 4-week-old Arabidopsis protoplasts.
After 18 h, total proteins were extracted with 0.5 mL protein extraction buffer (50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 10% glycerol, 10 mM DTT, 10 mM EDTA, 1 mM NaF, 1 mM Na2MoO4·2H2O, 1% [w/v] polyvinylpyrrolidone, 1% [v/v] IGEPAL CA- 630 [Sigma-Aldrich] and 1% [v/v] Roche protease inhibitor cocktail) and incubated with gentle shaking at 4°C for 1 h. Samples were then centrifuged at 14,000 rpm for 15 min at 4°C. Proteins were separated by 10% SDS–PAGE. Supernatants were incubated with 20 μL GFP-Trap (Chromotek) for 4 h at 4°C. Following incubation, beads were washed four times with TBS containing 0.5% (v/v) IGEPALCA-630. Proteins were separated by 10% SDS–PAGE.
SDS-PAGE and Immunoblotting
Ten percent SDS-PAGE gels were run at 80 to 140 V for 2 h before electroblotting on a polyvinylidene difluoride membrane (Millipore) at 100 V for 1 h at 4°C.
Membranes were rinsed in Tris-buffered saline (TBS) and blocked in 5% (w/v) nonfat milk powder in 0.1% (v/v) TBS-Tween for 1 h. Primary antibodies were diluted in TBS- Tween solution to the following concentrations and incubated overnight: anti-GFP (Santa Cruz), 1:3000; anti-HA (Santa Cruz), 1:3000. Membranes were washed 3 times
in TBS-Tween before 1 h of incubation with the secondary antibodies anti-mouse-HA (Santa Cruz) diluted 1:3000 or anti-rabbit-GFP (Santa Cruz) diluted 1:3000. Signals were visualized using an enhanced chemiluminescence system (Immobilon Western;
Millipore) and a LAS-3000 (Fujifilm) scanner following the manufacturer’s instructions.
Accession numbers
Sequence data from this thesis can be found in the EMBL/Genbank data libraries under accession numbers: ERF19 (AT1G22810), NINJA (AT4G28910), TPL (AT1G15750), PDF1.2a (AT5G44420), UBQ10 (AT4G05320).
Results
Loss of ERF19 function is linked with enhanced resistance to B. cinerea
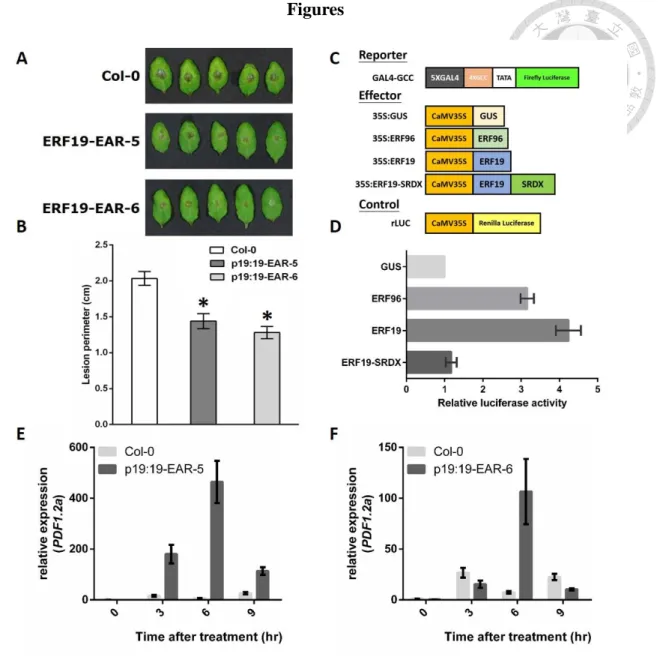
Previous studies in the Zimmerli laboratory show that both constitutive and inducible overexpression of ERF19 enhance Arabidopsis susceptibility to B. cinerea (Fig. S1). To confirm that ERF19 plays a negative role in Arabidopsis defense, we examined the resistance response of pERF19:ERF19-SRDX transgenic plants that demonstrate a lost in ERF19 activity after B. cinerea infection. For that purpose, 4- week-old WT and transgenic plants were droplet-inoculated with a suspension containing 105 B. cinerea spores per mL. Three days after infection, pERF19:ERF19- SRDX transgenic lines exhibited smaller lesions on leaves compared with WT plants (Fig. 1A, B). Protoplast transactivation assay (PTA) was used to confirm that the SRDX motif block the activation activity of ERF19. The PTA resulted in a 4.3 fold increase of luciferase activity when using GAL4DB-ERF19 as effector plasmid instead of GAL4DB-GUS effector plasmids. However, SRDX (LDLDLELRLGFA) fused at the C-terminus of ERF19 repressed the luciferase activity to the basal level (Fig. 1C, D).
This result indicated that SRDX fragment efficiently repress the activation activity of ERF19. The opposite infection phenotype between ERF19 overexpression plants and pERF19:ERF19-SRDX transgenic plants confirmed the negative role of ERF19 in Arabidopsis defense against B. cinerea.
pERF19:ERF19-SRDX transgenic lines display enhanced expression of defense gene
To examine whether the altered disease resistance phenotype of the pERF19:ERF19-SRDX transgenic lines were associated with altered defense gene expression, we analyzed transcript levels of defense-related genes using the qPCR
method. Seven-day-old WT and pERF19:ERF19-SRDX transgenic plants were treated with 20 μM chitin. We conducted quantitative real-time PCR on RNAs extracted from leaves of the transgenic plants (T3 lines 5-5 and 6-9) and WT. The expression level of PDF1.2a were comparable between the Col-0 and ERF19-SRDX plants in mock controls, suggesting that suppression of ERF19 target genes did not affect the expression of this defense gene. Consistent with infection phenotype on plants, all these transgenic lines revealed higher PDF1.2a gene expression after chitin treatment (Fig 1E, F). These results suggest that loss of ERF19 function results in enhanced sensitivity to the fungal PAMP, chitin.
Generation of Arabidopsis ERF19 artificial microRNA transgenic lines
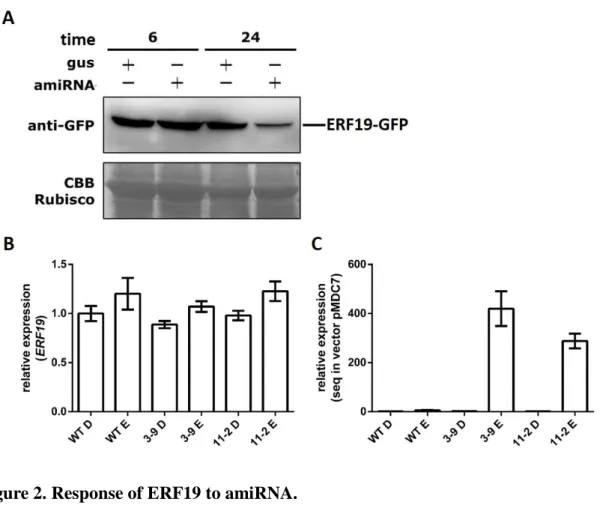
In addition to pERF19:ERF19-SRDX transgenic plants, we also generated β- estradiol induced microRNA-mediated suppression lines to do loss of ERF19 function analysis. Protein degradation in protoplast was used to confirm the efficiency of amiRNA construct. We observed a substantial reduction of ERF19-GFP protein in protoplast by its optimal amiRNA, at 24 h after amiRNA transfection and β-estradiol treatment (Fig. 2A). Two amiRERF19 lines (3-9 and 11-2) were selected for further functional studies. In our experiment, no visible morphological or vegetable growth or reproductive development defect was observed in the amiRERF19 plants. qRT-PCR analysis revealed that the expression levels of ERF19 in these two lines were similar with that in WT plants (Fig. 2B), however the transfected fragment were highly induced in both amiRNA lines (Fig. 2C). The results indicated that the amiRNA was expressed in both of the T3 amiRNA lines however ERF19 was not repressed efficiently in this condition. To solve this problem, other concentration of β-estradiol will be tried out in future experiments. Further B. cinerea infection analyses will be performed in these
ERF19 amiRNA transgenic plants.
Subcellular localization of ERF19 in Nicotiana benthamiana
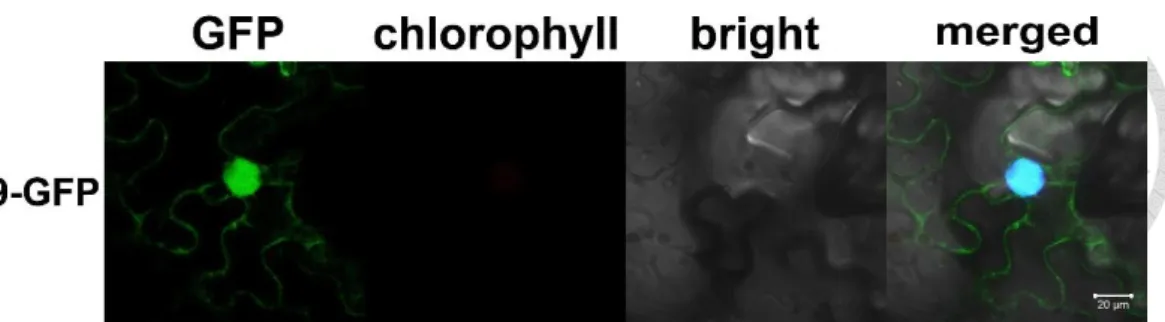
To examine the subcellular localization of ERF19, Agrobacterium tumefaciens carrying pEG103-ERF19 and the nuclear marker protein NLS-mCherry were injected into leaves of 4-week-old N. benthamiana with a 1:1 ratio. The ERF19-GFP fusion was mainly localized to the nucleus mimicking the NLS-mCherry signal localization, and also other cytoplasmic organelles (Fig. 3). However, in this study we focused on nucleus-localized transcriptional activity of ERF19, the cytoplasmic localization may have other not yet discovered functions.
ERF19 directly interacts with NINJA in vitro
Previous studies in the Zimmerli laboratory showed that ERF19 act as a transcriptional activator. However, the expression of defense genes in ERF19 overexpression lines is lower than in WT after B. cinerea inoculation or chitin treatment.
We thus hypothesized that some transcriptional co-repressors may work together with ERF19 to down-regulate the expression of defense genes. EAR motif-mediated transcriptional repression is one of the principal mechanisms in plant gene regulation.
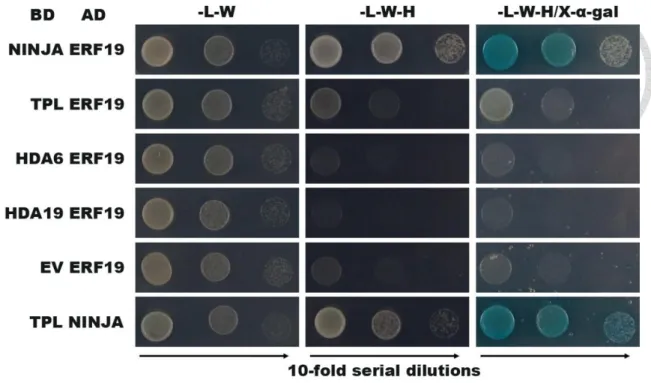
Many ERF repressors associate with the NINJA repressor complex to repress their target genes. So we hypothesized that ERF19 repress downstream genes by association with the NINJA repressor complex. Interactions were first evaluated with the yeast two hybrid (Y2H) assay. The documented interaction between NINJA and TPL was used as a positive control (Pauwels et al., 2010). Positive interactions were observed for ERF19 and NINJA, while no interaction was detected between ERF19 and TPL or in the negative control reactions (Fig. 4). The observed direct interaction of ERF19 with
NINJA suggests that NINJA may be the co-repressor involved in the process of the ERF19-mediated defense response against B. cinerea.
ERF19 associates with the NINJA repressor complex in vivo
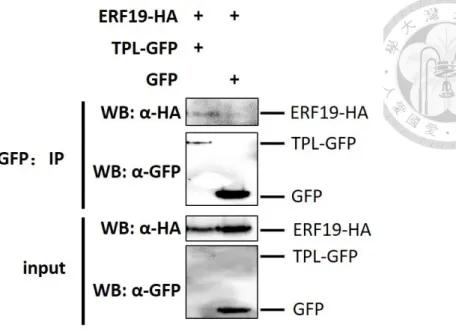
Next, associations were also evaluated by Co-Immunoprecipitation (Co-IP) in Arabidopsis protoplasts and by bimolecular fluorescence complementation (BiFC) assays in N. benthamiana leaves. For the Co-IP experiment, NINJA-GFP or TPL-GFP or GFP alone were transiently coexpressed with ERF19 epitope-tagged with 3HA in Arabidopsis protoplast. Equal amounts of GFP were immunoprecipitated with GFP- Trap beads and the presence of ERF19-3HA was analyzed using anti-HA immunoblotting. ERF19-3HA could be detected with NINJA (Fig. S2) and TPL (Fig.
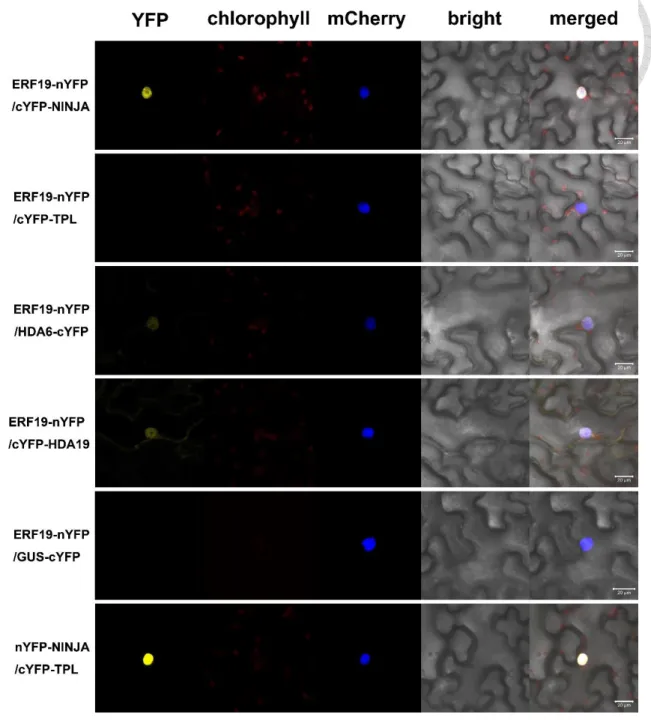
5), although the signal with TPL was very weak. In BiFC assay, YFP fluorescence was detected in the combination of ERF19-YN/YC-NINJA, but generally not of ERF19- YN/YC-TPL (Fig. 6). YN-NINJA/YC-TPL was used as positive control while ERF19- YN/GUS-YC was used as negative control. Taken together, these experiments confirmed the ERF19/NINJA protein association identified in the Y2H assay. The association of ERF19 and TPL detected in Co-IP, but not by Y2H or BiFC, suggested that these two proteins physically work in the same complex, but ERF19 does not directly interact with TPL.
ERF19 associates with HDA6 and HDA19 in vivo
Co-repressors commonly recruit chromatin-remodeling factors to actively repress gene expression, and prior genetic analyses implicated the RPD3-like HDA19 in TPL- dependent transcriptional regulation (Long et al., 2006). HDA6 is another RPD3/HDA1 family protein involved in the activation of JA-dependent defenses (Wu et al., 2008).
Therefore, we assessed the ability of ERF19 to associate with HDA19 and HDA6 in planta using Co-IP (Fig. S2) and BiFC. By BiFC, weak YFP fluorescence was detected in the combination of ERF19-YN/HDA6-YC and ERF19-YN/YC-HDA19 (Fig. 6).
Combined with the result of Y2H that the combination of BD-HDA6/AD-ERF19 and BD-HDA19/AD-ERF19 could not grow on –L-W-H media (Fig. 4), we concluded that the interaction between ERF19 and HDA19/HDA6 is not direct, as adapter proteins may facilitate association between the two factors. Regardless, these findings support our view that the activation activity of ERF19 is repressed by the NINJA repressor complex.
Arabidopsis ninja mutant exhibit enhanced resistance while NINJA overexpression lines are more susceptible to B. cinerea
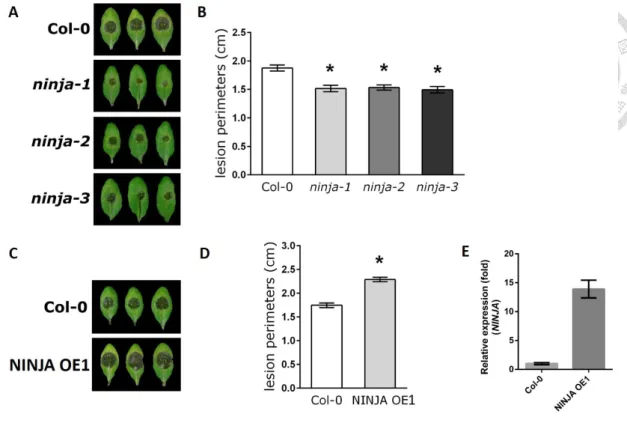
We next explored the possible role of NINJA in immunity against B. cinerea. Three ninja mutants (ninja-1, ninja-2 and ninja-3) (Acosta et al., 2013) and one NINJA overexpression line (NINJA OE1) (Pauwels et al., 2010) were droplet-inoculated with 105 spores per mL of B. cinerea. Three days after infection, the lesions were significantly smaller on leaves of ninja mutants when compared with WT plants (Fig.
7A, B). By contrast, the NINJA overexpression line (NINJA OE1) was much more susceptible than WT plants (Fig. 7C, D). Importantly, the NINJA RNA level of OE1 was 14 times higher than WT (Fig. 7E). The 3 ninja mutant were EMS point mutation lines, which were confirm by sequencing of genomic DNA (data not shown). These results suggested that loss of NINJA function increases defense responses, consistent with the published view that NINJA is a negative regulator in the JA signaling pathway that acts against B. cinerea infection (Acosta et al., 2013).
Overexpress ERF19 in Arabidopsis ninja mutant showed altered developmental phenotype
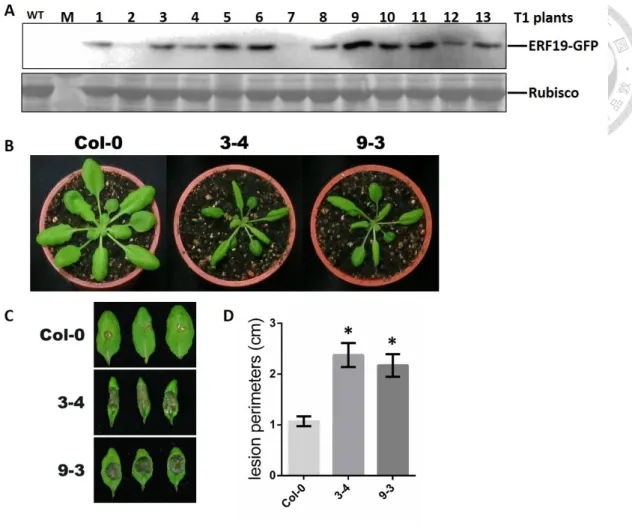
To analyze the genetic relationship of ERF19 and NINJA in the Arabidopsis defense response to B. cinerea, ninja-1 mutant overexpressing ERF19 transgenic lines were generated. Since ninja-1 mutant has the shortest functional sequence of NINJA and NINJA gene expression was well repressed, we chose this line to overexpress ERF19. Interestingly, the transgenic plants show altered development phenotype from T1 plants, with down-curled leaves and shorter petioles (Fig. 8A). Thirteen T1 plants were screened by growing them on basta plate. Each plants exhibited different ERF19 protein level (Fig. 8B) and only seven of them produced mature seeds. Interestingly, three of the seven T2 line seeds demonstrated no germination. Finally, we could obtain 4 viable T2 lines (2, 3, 7 and 9). The more variant developmental phenotype of line 3 and 9 corresponds to the higher ERF19 protein level.
Lines overexpressing ERF19 in the ninja mutant exhibits strong increased susceptibility to B. cinerea
Four-week-old ninja mutant overexpressing ERF19 T3 transgenic lines 2-5 and 3- 4 and WT plants were droplet-inoculated with a suspension containing 105 B. cinerea spores per mL. Two days after infection, ninja mutant overexpressing ERF19 developed significantly larger lesions than WT controls (Fig. 8C). The infection phenotype was much stronger than ERF19 overexpression in Col-0 background (Fig. S1), suggesting that the co-repressor NINJA possesses a repressor function in ERF19-mediated defense response. These results also suggest a genetic interaction between ERF19 and NINJA, in both developmental and immunity responses.
Discussion
Previous studies in the Zimmerli laboratory showed that overexpression of ERF19 increased Arabidopsis susceptibility to the necrotrophic pathogen B. cinerea, suggesting that ERF19 plays a negative role in the Arabidopsis defense response against B. cinerea. However, there is no appropriate T-DNA insertion mutant line for ERF19 gene. In this thesis, we generated inducible ERF19 artificial microRNA transgenic plants and pERF19:ERF19-SRDX transgenic plants to perform loss of ERF19 function analyses. The pERF19:ERF19-SRDX transgenic plants were more resistant to B.
cinerea infection than WT plants and had higher PTI marker gene expression when treated with the PAMP chitin. Since ERF19 overexpression lines were more susceptible to B. cinerea while loss of ERF19 function lines were more resistant to B. cinerea than WT plants, I postulate for a negative role of ERF19 in the plant defense response against necrotrophic fungal pathogens.
ERF19 plays negative roles in defense however it acts as a transcriptional activator when analyzed with the PTA technique. We hypothesized that the transcription factor ERF19 repress its target genes by association with other transcriptional regulators that act as repressors, or ERF19 activates transcriptional repressors which directly repress defense-related genes. In general, HDACs have been found to mediate gene repression (Tanaka et al., 2008) and to participate in the process of plant immunity (Zhou et al., 2005; Ding and Wang, 2015). For example, overexpression of HDA19 enhances fungal resistance through the apparent activation of ERF1, whereas silencing of the gene increases fungal susceptibility (Zhou et al., 2005). Another histone deacetylase, HDA6 is also involved in the activation of JA-dependent defenses (Wu et al., 2008).
Interestingly, ERF19 associated with HDA6 and HDA19 when analyzed with the Co- IP (Fig. S2B) and BiFC (Fig. 6) techniques. This observation suggests that ERF19 may
regulate the defense process against B. cinerea through epigenetic controls. In addition, there are mainly four active repression motifs in plants, and the EAR motif (LxLxL or DLNxxP) is linked with the chromatin modifier, HDA19 (Kagale and Rozwadowski, 2010). Among 147 AP2/ERF proteins in Arabidopsis, 23 contain an EAR (or EAR-like) motif or a B3 repression domain (BRD: RLFGV) (Nakano et al., 2006). However, neither motifs are present in the ERF19 sequence. The JA signaling pathway is an important reference model for the understanding of ERF19 role in defense. The transcriptional activator MYC2 is blocked by JAZ1, NINJA, TPL, and the HDAC repressor complex in the absence of JA–isoleucine (Zhang et al., 2015). In this model, NINJA that possesses an EAR motif, plays the role of co-repressor of MYC2. We thus postulated that the activation activity of ERF19 is also repressed by association with the NINJA repressor complex. ERF19 indeed directly interacted with NINJA in vivo and in vitro. In addition, ERF19 also associated with TPL when analyzed by CoIP but not with the BiFC technique, suggesting these 2 proteins are in the same complex but do not interact directly. TPL is known to function in conjunction with HDA19. In our BiFC result, TPL did not associate with HDA6 or HDA19 (data not shown), which means unknown adapter proteins may facilitate the association between TPL and HDACs. The direct interaction of ERF19 and NINJA suggests that the NINJA repressor binds and inhibits the TF ERF19 through direct inhibition or through the recruitment of TPL co-repressors. TPL in turn could recruits HDACs to repress gene expression through chromatin remodeling.
To test whether ERF19 needs NINJA to negatively regulate plant defenses, we first performed infection on ninja mutant and NINJA overexpression lines. Consistent with the published view that NINJA is a negative regulator in JA signaling, ninja mutants were more resistant while a NINJA overexpression line was more susceptible to B.
cinerea infection. Next we also performed infection on lines overexpression ERF19 in the ninja mutant background. The developmental phenotype of these transgenic lines were completely changed with short petioles and down-curled leaves. In addition, these transgenic plants were much more susceptible to B. cinerea infection than lines overexpressing ERF19 only. In addition, PTA performed by Jason showed that NINJA repress the activation activity of ERF19 (Fig. S3). Together, these results suggested that NINJA has an active role in blocking the function of ERF19 during the Arabidopsis resistance to B. cinerea.
To find out the direct target of the TF ERF19, the PhD student Yao in the Zimmerli laboratory performed microarray experiments at the Affymetrix Gene Expression Service Lab at Academia Sinica (Taipei, Taiwan). Microarray analysis coupled to chromatin immunoprecipitation (Chip)-qPCR of Arabidopsis overexpressing ERF19 revealed that ERF19 directly binds to the promoter region of the TF MYB X but not the defense gene PDF1.2a (Fig. S4). To further elucidate the mechanism of action of ERF19, Chip-qPCR will be used to test whether MYB X directly binds to the promoter region of defense-related genes.
In conclusion, we built a model to illustrate how ERF19 regulate its target genes.
In normal condition, NINJA physically interacts with and represses ERF19, then the EAR motif of NINJA interacts with TPL and TPL recruit HDA6/19 to block the target genes of ERF19. Upon overexpression of ERF19, the ERF19 target gene MYB X is efficiently activated to repress defense-related genes and compromise disease resistance.
Future Perspectives
This work suggests a model explaining how ERF19 regulates Arabidopsis defense against B. cinerea. Except for the direct interaction of ERF19 and NINJA, we will further analyze the interaction domain of these 2 proteins. This study may give us new ideas of how ERF/AP2 family proteins regulate transcription. And the PhD student Yao in the Zimmerli laboratory performed Microarray coupled to Chip-qPCR analysis to find that ERF19 directly binds to the promoter region of the TF MYB X. So in the future, we will use PTA to test whether MYB X represses the transcription of PDF1.2 or other defense-related genes. We will also generate Arabidopsis MYB X-GFP overexpression lines and then use Chip-qPCR analysis to check whether MYB X directly binds to defense-related genes.
Figures
Figure 1. Disease responses of pERF19:ERF19-SRDX transgenic plants to B.
cinerea infection.
(A and B) B. cinerea-mediated infection. Four-week-old Arabidopsis was droplet- inoculated with B. cinerea (droplets of 8 μL with 105 spores/mL) and symptoms (A) or lesion perimeters (B) were evaluated at 3 dpi. For (B), values are means ± SD of 3 biological replicates each with 8 technical repeats (n = 24). Asterisks indicate a significant difference to the WT control as determined by t-test analysis (P < 0.05). (C) Schematic diagram of the effector and reporter constructs used in PTA. (D) PTA.
Relative luciferase activities were evaluated in mesophyll protoplasts co-transfected
with the reporter plasmid (35S-UAS-LUCIFERASE) and the effector plasmids (35S- GUS, 35S- ERF96, 35S-ERF19 and 35S:ERF19-SRDX) and a calibrator plasmid encoding the Renilla luciferase. The effector plasmids encoding exogenous protein GUS (35S-GUS) was used as negative control. All values were normalized to the Renilla luciferase activity and a value of 1 was attributed to the luciferase activity measured in protoplast transformed with the reporter 35S-GUS. (E and F) Relative PDF1.2a expression levels in 7-day-old WT and pERF19:ERF19-EAR-5-5 (E) and pERF19:ERF19-EAR-6-9 (F) plants 3, 6 and 9 h after 20 μM chitin treatment. Relative expression levels were compared to WT at time 0 (defined value of 1) by qRT-PCR analyses. Transcript levels were normalized to UBQ10. Values are means ± SD of 3 technical repeats (n = 3).
Figure 2. Response of ERF19 to amiRNA.
(A) Degradation of ERF19 when treated with β-estradiol induced microRNA. Levels of accumulated ERF19-GFP protein in Arabidopsis protoplast after 20 μM β-estradiol treatment for 6 and 24 h. (B) Relative ERF19 gene expression levels in ERF19 amiRNA T3 transgenic plants. (C) Relative sequence in the pMDC7 vector gene expression levels in ERF19 amiRNA T3 transgenic plants. D, DMSO treatment; E, β-estradiol treatment.
Figure 3. Subcellular localization of ERF19.
GFP-fused ERF19 was transiently expressed in N. benthamiana leaves by agrobacterium-mediated infiltration and visualized at 3 d post-infiltration using a confocal microscope. Bars, 20 µm. The same results were observed in three independent experients.
Figure 4. Direct interaction of ERF19 with NINJA but not with TPL, HDA6 or HDA19 by Y2H
Yeast strains expressing the indicated GAL4AD and GAL4BD fusions were grown on dropout media lacking leucine (−L), tryptophan (−W), and/or histidine (−H). Activation of the GAL4-regulated HIS reporter genes is seen as growth on –LWH media and of the MEL1 gene as a blue staining after overlaying the cells with X-α-Gal.
Figure 5. ERF19 associates with TPL in vivo.
Analysis of association between ERF19 and TPL by Co-IP assay in 4-week-old Arabidopsis protoplast. Total protein extracts (input) and IP-proteins were detected using immunoblotting with α-GFP or α-HA antibody. Experiments were repeated three times with similar results.
Figure 6. Association of ERF19 with NINJA, HDA6 and HDA19 but not with TPL by BiFC
BiFC assay of the ERF19 and NINJA, TPL, HDA6, HDA19 association in N.
benthamiana leaves infiltrated with A. tumefaciens (OD600 = 0.4). The combination ERF19-nYFP/GUS-cYFP was used as negative control. The fluorescent signals were detected using a confocal microscope 3 d after infiltration. Bars = 20 μm. This experiment was repeated three times with the same results.
Figure 7. Disease responses of ninja mutant and NINJA overexpression plants to B. cinerea infection.
(A, B) B. cinerea-mediated infection. Four-week-old Arabidopsis ninja-1, ninja-2, ninja-3 and WT were droplet-inoculated with B. cinerea. Symptoms (A) or lesion perimeters (B) were evaluated at 3 dpi. (C, D) Four-week-old Arabidopsis NINJA overexpression (OE1) line and WT were droplet-inoculated with B. cinerea. Symptoms (C) or lesion perimeters (D) were evaluated at 3 dpi. For (B, D), values are means ± SD of 3 biological replicates each with 24 technical repeats (n = 72). Asterisks indicate a significant difference to the WT control as determined by t-test analysis (P < 0.05). (E) Relative NINJA expression levels in 4-week-old WT and NINJA OE1 line. Relative expression levels were compared with WT (defined value of 1) by qRT-PCR analyses.
Transcript levels were normalized to UBQ10. Results were average ± SD of three biological replicates each consisting of three technical repeats (n=9).
Figure 8. Development and defense phenotype of ninja mutant overexpressing ERF19.
(A) ERF19 proteins level in 13 ninja mutant overexpressing ERF19 T1 plants. (B) Morphological phenotype observed in 4-week-old ninja mutant expressing ERF19 T3 transgenic plants 3-4 and 9-3 growing side by side with a WT plant. (C and D) B.
cinerea-mediated infection. Four-week-old Arabidopsis WT and transgenic plants were droplet-inoculated with B. cinerea (droplets of 8 mL with 105 spores/mL) and symptoms (C) or lesion perimeters (D) were evaluated at 2 dpi. For (D), values are means ± SD of 3 biological replicates each with 8 technical repeats (n = 24). Asterisks indicate a significant difference to the WT control as determined by t-test analysis (P <
0.05).
Figure 9. Model for ERF19-mediated defense repression in Arabidopsis.
In normal condition, NINJA physically interacts with and represses ERF19, then the EAR motif of NINJA interacts with TPL and TPL recruit HDA6/19 to block MYB X, the target gene of ERF19. Overexpression of ERF19 in Arabidopsis, the ERF19 target gene MYB X is efficiently induced to repress defense-related genes and compromise disease resistance.
Supplementary Figures
Figure S1. Constitutive and inducible overexpression lines of ERF19 were more susceptible to B. cinerea infection than WT.
(A and B) B. cinerea-mediated infection. Four-week-old Arabidopsis WT and ERF19 constitutive overexpression lines were droplet-inoculated with B. cinerea. Symptoms (A) and lesion perimeters (B) were evaluated at 3 dpi. (C and D) B. cinerea-mediated infection. Four-week-old Arabidopsis WT and ERF19 β-estradiol inducible overexpression line were droplet-inoculated with B. cinerea after DMSO or β-estradiol treatment. Symptoms (C) and lesion perimeters (D) were evaluated at 3 dpi. For (B, D), values are means ± SD of 3 biological replicates each with 8 technical repeats (n = 24).
Asterisks indicate a significant difference to the WT control as determined by t-test analysis (P < 0.05).
Figure S2. ERF19 associates with NINJA, HDA6 and HDA19 by Co-IP in Arabidopsis protoplast.
Total proteins (input) of Arabidopsis protoplast expressing NINJA-GFP, HDA6-GFP, HDA19-GFP with ERF19-HA were immunoprecipitated with anti-GFP antibodies and then analyzed by immunoblot with an anti-HA antibody.
Figure S3. NINJA repress the activation of ERF19.
(A) Schematic diagram of the effector and reporter constructs used in PTA. (B) Relative luciferase activity measured 16 h after mesophyll protoplast transformation. The ERF3RD and ERF19 effector plasmids were under the control of the CaMV 35S promoter with GAL4 binding domain, whereas NINJA effector plasmid was under the control of the CaMV 35S promoter only. Presence and absence of the effector plasmids were indicated by + and -, respectively.
Figure S4. ERF19 binds to the promoter region of MYB X but not PDF1.2a. Chip- qPCR analysis of ERF19 binding to the promoter region of MYB X. Four-week-old Arabidopsis WT, ERF19 β-estradiol-inducible overexpression plants and ERF19 overexpression plants were used for Chip analysis. Fold enrichment of each fragment was calculated by normalizing the amount of target DNA fragments against the input DNA.
Tables Table 1. Mutant alleles used in this study
Allele Mutation Location in gene (nt)
ninja-1 C→T transition; causes P224STOP substitution 673 ninja-2 G→A transition in first base of last intron; destroys
donor splice site; transcript including this intron codes for 17 altered amino acids after position S297 and prior to a premature STOP codon
893
ninja-3 2-bp deletion; causes frameshift that codes for 46 altered amino acids after position T363 and prior to a premature STOP codon
1164-1165
Table 2. Primers used for ninja mutant check Primer 5'-3' sequence
ninja-F1 TGGTGGTTCTTCTTCCAACC
ninja-R1 GCAACAGGTTGTTTGCCTTC ninja-R2 TCAGGTGTGAGCTGACGCT
Table 3. Primers used for ERF19 amiRNA plasmid construction Primer name 5'-3' sequence
micro-I GATGTAATACGTCGTGGTATCTCTCTCTCTTTTGTATTCC
micro-II GAGAGATACCACGACGTATTACATCAAAGAGAATCAATGA
micro-III GAGAAATACCACGACCTATTACTTCACAGGTCGTGATATG
micro-IV GAAGTAATAGGTCGTGGTATTTCTCTACATATATATTCCT
Primer A CTGCAAGGCGATTAAGTTGGGTAAC
Primer B GCGGATAACAATTTCACACAGGAAACAG
Table 4. Primers used for plasmid construction Gene or motif Primer 5'-3' sequence
ERF19 FP ATGCGTCGACTCAAACGTGATCGTGGC
w stop-RP TCAAACGTGATCGTGGCCGCCCA
w/o stop-RP2 AACGTGATCGTGGCCGCCCA
NINJA FP ATGGACGATGATAATGGGCTCGA
w stop-RP TCAGGTGTGAGCTGACGCTGCA
w/o stop-RP2 GGTGTGAGCTGACGCTGCA
TPL FP ATGTCTTCTCTTAGTAGAGAGCTCGTTTTCTTG
w stop-RP TCATCTCTGAGGCTGATCAGATGCAGAG
w/o stop-RP2 TCTCTGAGGCTGATCAGATGCAGAG
HDA6 FP ATGGAGGCAGACGAAAGCG
w stop-RP TTAAGACGATGGAGGATTCACGT
w/o stop-RP2 AGACGATGGAGGATTCACGT
HDA19 FP ATGGATACTGGCGGCAATTC
w stop-RP TTATGTTTTAGGAGGAAACGCCT
w/o stop-RP2 TGTTTTAGGAGGAAACGCCT
SRDX RP TTAAGCGAAACCCAAACGGAGTTC
Table 5. Primers used for qRT-PCR
Gene or motif Primer 5'-3' sequence
UBQ10
FP GGCCTTGTATAATCCCTGATGAAT
RP TGTTTCCGTTCCTGTTATCTCTTT
ERF19
FP GAAATGGGGCAAATGGGTATC
RP CTGCTGTTGAGAATGAACCTAA
NINJA
FP CGGATACTCACCAGTCCAGC
RP GGTTGTTTGCCTTCGCCTTT
PDF1.2a
FP AATCTTTGGTGCTAAATCGTGTG
RP CAACGGGAAAATAAACATTAAAA
pMDC7
FP ATTGTTATCCGCAAGGGCGA
RP TGATACGGACGAAAGCTGGG
References
Acosta, I.F., Gasperini, D., Chetelat, A., Stolz, S., Santuari, L., and Farmer, E.E.
(2013). Role of NINJA in root jasmonate signaling. Proceedings of the National Academy of Sciences of the United States of America 110, 15473-15478.
Bari, R., and Jones, J.D. (2009). Role of plant hormones in plant defence responses.
Plant Mol Biol 69, 473-488.
Berrocal‐Lobo, M., Molina, A., and Solano, R. (2002). Constitutive expression of ETHYLENE‐RESPONSE‐FACTOR1 in Arabidopsis confers resistance to several necrotrophic fungi. The Plant Journal 29, 23-32.
Brown, R.L., Kazan, K., McGrath, K.C., Maclean, D.J., and Manners, J.M. (2003).
A role for the GCC-box in jasmonate-mediated activation of the PDF1. 2 gene of Arabidopsis. Plant physiology 132, 1020-1032.
Cao, Y., Liang, Y., Tanaka, K., Nguyen, C.T., Jedrzejczak, R.P., Joachimiak, A., and Stacey, G. (2014). The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1.
eLife 3.
Catinot, J., Huang, J.B., Huang, P.Y., Tseng, M.Y., Chen, Y.L., Gu, S.Y., Lo, W.S., Wang, L.C., Chen, Y.R., and Zimmerli, L. (2015). ETHYLENE RESPONSE FACTOR 96 positively regulates Arabidopsis resistance to necrotrophic pathogens by direct binding to GCC elements of jasmonate - and ethylene- responsive defence genes. Plant Cell Environ 38, 2721-2734.
Causier, B., Lloyd, J., Stevens, L., and Davies, B. (2012a). TOPLESS co-repressor interactions and their evolutionary conservation in plants. Plant signaling &
behavior 7, 325-328.
Causier, B., Ashworth, M., Guo, W., and Davies, B. (2012b). The TOPLESS
interactome: a framework for gene repression in Arabidopsis. Plant physiology 158, 423-438.
Chini, A., Boter, M., and Solano, R. (2009). Plant oxylipins: COI1/JAZs/MYC2 as the core jasmonic acid‐signalling module. Febs Journal 276, 4682-4692.
Chini, A., Fonseca, S., Fernandez, G., Adie, B., Chico, J., Lorenzo, O., Garcia- Casado, G., Lopez-Vidriero, I., Lozano, F., and Ponce, M. (2007). The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448, 666-671.
Dean, R., Van Kan, J.A., Pretorius, Z.A., Hammond-Kosack, K.E., Di Pietro, A., Spanu, P.D., Rudd, J.J., Dickman, M., Kahmann, R., Ellis, J., and Foster, G.D. (2012). The Top 10 fungal pathogens in molecular plant pathology.
Molecular plant pathology 13, 414-430.
Ding, B., and Wang, G.-L. (2015). Chromatin versus pathogens: the function of epigenetics in plant immunity. Frontiers in plant science 6.
Eulgem, T., and Somssich, I.E. (2007). Networks of WRKY transcription factors in defense signaling. Current opinion in plant biology 10, 366-371.
Felix, G., Regenass, M., and Boller, T. (1993). Specific perception of subnanomolar concentrations of chitin fragments by tomato cells: induction of extracellular alkalinization, changes in protein phosphorylation, and establishment of a refractory state. The Plant Journal 4, 307-316.
Huang, P.Y., Catinot, J., and Zimmerli, L. (2016). Ethylene response factors in Arabidopsis immunity. J Exp Bot 67, 1231-1241.
Ikeda, M., and Ohme-Takagi, M. (2009). A novel group of transcriptional repressors in Arabidopsis. Plant Cell Physiol 50, 970-975.
Kagale, S., and Rozwadowski, K. (2010). Small yet effective: the ethylene responsive
element binding factor-associated amphiphilic repression (EAR) motif. Plant signaling & behavior 5, 691-694.
Kaku, H., Nishizawa, Y., Ishii-Minami, N., Akimoto-Tomiyama, C., Dohmae, N., Takio, K., Minami, E., and Shibuya, N. (2006). Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor.
Proceedings of the National Academy of Sciences 103, 11086-11091.
Kao, H.-Y., Ordentlich, P., Koyano-Nakagawa, N., Tang, Z., Downes, M., Kintner, C.R., Evans, R.M., and Kadesch, T. (1998). A histone deacetylase corepressor complex regulates the Notch signal transduction pathway. Genes &
development 12, 2269-2277.
Licausi, F., Ohme-Takagi, M., and Perata, P. (2013). APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors: mediators of stress responses and developmental programs. New Phytol 199, 639-649.
Liu, Z., and Karmarkar, V. (2008). Groucho/Tup1 family co-repressors in plant development. Trends in plant science 13, 137-144.
Long, J.A., Ohno, C., Smith, Z.R., and Meyerowitz, E.M. (2006). TOPLESS regulates apical embryonic fate in Arabidopsis. Science 312, 1520-1523.
Macho, A.P., and Zipfel, C. (2014). Plant PRRs and the activation of innate immune signaling. Mol Cell 54, 263-272.
Matsui, K., Umemura, Y., and Ohme-Takagi, M. (2008). AtMYBL2, a protein with a single MYB domain, acts as a negative regulator of anthocyanin biosynthesis in Arabidopsis. The Plant journal : for cell and molecular biology 55, 954-967.
McGrath, K.C., Dombrecht, B., Manners, J.M., Schenk, P.M., Edgar, C.I., Maclean, D.J., Scheible, W.-R., Udvardi, M.K., and Kazan, K. (2005).
Repressor-and activator-type ethylene response factors functioning in