國立臺灣大學醫學院口腔生物科學研究所 碩士論文
Graduate Institute of Oral Biology, College of Medicine
National Taiwan University Master Thesis
探討口腔鱗狀上皮細胞癌中表現 B7-H1和PD-1之浸潤T淋巴球
Identification of B7-H1 and PD-1 expressing T cells infiltrated in human oral squamous cell carcinoma
歐育志 Yu-Chih Ou
指導教授:賈景山 博士 Advisor: Jean-San Chia, Ph.D.
中華民國九十八年七月
July, 2009
國立臺灣大學碩士學位論文 口試委員會審定書
探討口腔鱗狀上皮細胞癌中表現 B7-H1和PD-1之浸潤T淋巴球
Identification of B7-H1 and PD-1 expressing T cells infiltrated in human oral squamous cell carcinoma
本論文係歐育志君(R96450009)在國立臺灣大學口腔生 物科學研究所完成之碩士學位論文,於民國九十八年六月二 十四日承下列考試委員審查通過及口試及格,特此證明
指導教授 賈景山博士
口腔生物科學研究所所長 郭彥彬博士
口試時間 中華民國九十八年六月二十四日
致謝
這兩年的台大研究生活非常的充實,首先感謝賈景山教授讓我接 觸臨床的研究工作,讓我不只是學到實驗的技術,還有很多的邏輯思 考和待人處世的方式,感謝許秉寧老師和李建國老師對我論文以及口 試所給的指導,感謝秋月學姐對我實驗技術、實驗方向和論文思考所 給予的指導,感謝珮茹學姐和淑惠學姐在我剛進實驗室時對我的耐心 指導,感謝惠婷學姐、筱菁學姐、佩旻學姐,感謝奇玄學長、達哥、
韋君學姐、雅婷學姐、宛伶、中帆、旻潔、鴻偉,感謝我母親以及姐 姐的支持,最後感謝律穎的陪伴。
摘要
在口腔中超過95%的惡性腫瘤為口腔鱗狀上皮細胞癌,並且為台灣六大常見 侵襲癌症死因之ㄧ。人類產生在不同器官或組織的癌症,其疾病起因除了基因毒 性和致癌基因突變外,免疫編輯(immune-editing)也被視為一個重要的因素,目前 已經有研究指出調節性T細胞(Tregs)和表現抑制性B7分子的細胞對於腫瘤微環境 的免疫抑制以及腫瘤生長都有貢獻。在本論文中發現口腔癌腫瘤組織抑制性 B7-H1分子和調節性T細胞的表現相關性,接著分析口腔癌病人表現抑制性B7-H1 分子的腫瘤浸潤調節性T細胞。研究發現腫瘤組織CD4+CD25+Foxp3+調節性T細 胞有88.08%表現B7-H1,比起病人PBMC(64.71%)(p= 0.001)和正常人PBMC (73.78%) (p=0.04)都高出許多,並且發現腫瘤組織B7-H1+Foxp3+和Foxp3+細胞之 百分比在TIL呈現中度相關(r=0.61),而在CD4+ T細胞內則是呈現高度相關 (r=0.75)。TIL內CD4+ CD25+Foxp3-活化T細胞之B7-H1接受體PD-1(programmed death-1)的平均表現螢光表現強度大約是PBMC的六倍高,CD4+CD25-Foxp3-未活 化T細胞在TIL大約是PBMC的五倍高,然而CD4+CD25+Foxp3+調節性T在TIL大 約只有PBMC的二倍高,並且發現B7-H1+Foxp3+表現百分比和PD-1表現強度在 腫瘤組織CD4+T 細胞(r=0.6)具有中度相關性。此外在組織免疫染色也發現,口 腔癌腫瘤組織中有Foxp3+ B7-H1+、Foxp3+ B7-H1-和PD-1+的浸潤T細胞。因此 本論文發現有表現共同抑制性B7-H1分子的調節性T細胞亞群,浸潤於口腔鱗狀 上皮細胞癌中,並且可能是藉由和PD-1的結合而具有免疫調節的功能。
Abstract
Oral squamous cell carcinoma(OSCC) accounts for over 90% of the oral
malignant neoplasms, ranks as the sixth major cause of cancer mortality rate in Taiwan and the incidence rate is raising. In addition to mutation of oncogene and gene toxicity, the cancer immune-editing is one of the important factors in human cancer currently.
The regulatory T cell and high levels of expression of coinhibitory B7 molecules are the mediators of immunosuppression in tumor microenviriment. Therefore, we
investigated the relationship of B7-H1 and regulatory T cells in human oral squamous cell carcinoma.The results indicate that, in TIL, proportion of distinct CD25+ Foxp3+
cells in the CD4+ subset (88.08%) were enriched relative to that found in PBMC (64.71 %) from OSCC patients(p= 0.001)and PBMC(73.78%) from healthy donor (p=0.04).The linear correction analysis indicated that Foxp3+ and B7-H1+Foxp3+
Tregs were positively correlated in TIL(R=0.61) and CD4+ T cell(R=0.75). The PD-1 (the receptor of B7-H1) average expression on CD4+CD25-Foxp3- and CD4+CD25+
Foxp3- in TIL is five- or six-fold to PBMC , but it is only two fold in CD4+CD25+
Foxp3+ regulatory T cell. The linear analysis indicated that PD-1 average expression and proportion of B7-H1+Foxp3+ Tregs on CD4+T cell were positively correlated (R=0.60).Furthermore, we also observe the B7-H1+ Foxp3+Tregs and PD-1+ T cell in the immunohistochemistry. Therefore, B7-H1+Tregs and PD-1+ T cell infiltrated in tumor may play a immunoregulatory role in oral squamous cell carcinoma.
目錄
口試委員會審定書……….………1
致謝……….………2
中文摘要………..………3
英文摘要……….……….………4
目錄………..………5
第一章、 緒論 第一節、研究背景 ㄧ、口腔鱗狀上皮細胞癌……….………..9
二、口腔癌免疫抑制作用……….……..11
三、調節性 T 細胞(regulatory T cells) ……….……….12
四、抑制性B7分子(inhibitory B7 molecules) ……….…………..15
第二節、研究動機與目的……….……...20
第二章、 實驗分法與材料 第一節、檢體的收集及來源……….…………21
第二節、人類口腔鱗狀上皮細胞癌組織內淋巴球的分離與純化………….…22
第三節、人類周邊血液單核球的分離與純化……….…………22
第四節、細胞表面染色……….………23
第五節、細胞內染色……….………23
第六節、腫瘤組織和正常組織之 RNA 萃取……….………24
第七節、即時定量聚合酶連鎖反應(real-time PCR)……….……..24
第八節、口腔癌組織及正常人組織的包埋和冷凍切片……….…………24
第九節、組織免疫染色……….………25
第十節、反應性 T 細胞及調節性 T 細胞篩選分離方法及細胞增生和抑制試驗 ….……….……….25
第十一節、周邊血液單核球與口腔鱗狀癌細胞株體外共同培養方法….….26
第三章、 結果
第一節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤CD4+CD25+淋巴球,其
B7-H1共同抑制分子的細胞表現百分比和表現強度…………...……27
第二節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤 CD4+T 細胞,B7-H1 和 Foxp3、CD25 之表現有相關性……….….………...…27 第三節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤 CD4+Foxp3+調節性 T 細胞
表現較高比例且較高量的 B7-H1,且B7-H1+Foxp3+細胞和 Foxp3+細 胞有相關性………...…….…….…28 第四節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤 CD4+CD25+Foxp3+調節性
T 細胞表現較高比例且較高量的 B7-H1……….……..……29 第五節、口腔鱗狀細胞癌腫瘤組織 CD4+CD25+Foxp3-和 CD4+CD25-Foxp3-T 細 胞 表 現 較 高 量 的 PD-1 , 且 CD25+Foxp3+B7-H1+ T 細 胞 和 Foxp3-PD-1+ T 細胞有相關性………...30 第六節、口腔鱗狀細胞癌細胞株 SAS 誘導 CD4+CD25+細胞之 B7-H1 表現
.………....31 第七節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤 B7-H1+CD4+CD25+
Foxp3+細胞表現較高比例且較高量的 ICOS……….……….……....31
第八節、腫瘤組織表現較高量B7-H1、Foxp3、IL-10、IFN-γ、CD4、PD-1、
B7-DC和B7-H4之RNA,且Foxp3和B7-H1有相關性…….………....32
第 九 節 、 口 腔 鱗 狀 細 胞 癌 病 患 分 離 出 調 節 性 T 細 胞 具 有 抑 制 細 胞 增 生 的 功 能….………...…………....……...…..32
第四章、 討論……….………..….…….34 第五章、 參考文獻………..………..…….……38 第六章、 圖表
圖一、PBMC 和 TIL CD4+CD25+T 細胞之 B7-H1 共同抑制分子細胞群體表現 百分比和平均細胞螢光表現強度………...………..50
圖二、統計 PBMC 和 TIL 的 CD4+CD25+T 細胞之 B7-H1 共同抑制分子細胞 群體表現百分比和平均細胞螢光表現強度……… …....51 圖三、TIL CD4+CD25high、intermediate、low、negative T 細胞之 B7-H1 共同 抑制分子細胞群體表現百分比和平均細胞螢光表現強度….…………52 圖四、PBMC 和 TIL CD4+Foxp3+ T 細胞之 B7-H1 共同抑制分子細胞群體表 現百分比和平均細胞螢光表現強度……….……53 圖五、統計 PBMC 和 TIL CD4+Foxp3+ T 細胞的 B7-H1 共同抑制分子細胞群 體表現百分比和平均細胞螢光表現強度……….…………54 圖六、腫瘤組織Foxp3+B7-H1+T 細胞和 Foxp3+T 細胞的相關性………55 圖七、PBMC 和 TIL 的 CD4+CD25+Foxp3+、CD4+CD25+Foxp3-、
CD4+CD25-Foxp3- T 細胞之 B7-H1 共同抑制分子細胞群體表現百分比 和平均細胞螢光表現強度……….…..56 圖八、統計PBMC 和 TIL 的 CD4+CD25+Foxp3+、CD4+CD25+Foxp3-、
CD4+CD25-Foxp3- T 細胞之 B7-H1 共同抑制分子細胞群體表現百分比 和平均細胞螢光表現強度………..….57 圖九、PBMC 和 TIL 的 CD4+CD25+Foxp3+、CD4+CD25+Foxp3-、
CD4+CD25-Foxp3- T 細胞之 PD-1 共同抑制分子的細胞螢光表現強 度………...………...…….58
圖十、腫瘤組織Foxp3+B7-H1+T 細胞和 PD-1 T 細胞的相關性………...…59 圖十一、口腔鱗狀細胞癌腫瘤組織經組織切片和免疫染色所見到的
CD4+CD25+ Foxp3+B7-H1+調節性 T 細胞和 PD-1 浸潤細胞...…60
圖十二、口腔鱗狀細胞癌細胞株SAS 誘導 CD4+CD25+T 細胞之 B7-H1 表 現……….……….………61 圖十三、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤CD4+Foxp3+細胞的 B7-H1 和 ICOS 表現相關性……….……..62 圖十四、在腫瘤組織RNA level B7-H1 和相關分子的表現量以及其相關性 ……….…………..63 圖十五、口腔鱗狀細胞癌病患之PBMC CD4+CD25hi 細胞的細胞增生抑制試 驗……….………..…64
附圖一、調節性 T 細胞免疫抑制機制……….…….65 附圖二、B7 分子的表現和功能……….…….……...65
第一章 、緒論
第一節 、研究背景
一、 口腔鱗狀上皮細胞癌(oral squamous cell carcinoma)
癌症的發生是由於基因的突變或缺失,使得原本正常的細胞轉型變成持續不
斷生長的癌細胞,口腔癌就是屬於口腔及口咽部任何一部位所發生的癌症。口腔 包括的部位有唇、頰黏膜(唇和臉頰的內襯)、牙齒、齒槽、舌頭下方的口腔底 部、前三分之二的舌頭、口腔頂部的前面部分(硬顎)、牙齦及智齒後方的小區 域,口咽部則包含了後三分之一的舌頭、軟顎、扁桃腺及喉嚨後壁,另外唾液腺 遍布口腔並製造唾液,以保持口腔的濕潤及幫助消化食物,其中口腔癌發生的部 位以舌部和頰粘膜最多。口腔癌從組織學上可分為有鱗狀細胞癌(squamous cell carcinoma)、疣狀癌、腺樣囊狀癌、黏液表皮樣癌、黑色素瘤,其中又以鱗狀細 胞癌佔絕大多數(約佔90%)(Hunter et al., 2005)。
在臺灣地區,根據 2009 年行政院衛生局癌症資料統計,口腔癌是男女兩性 癌症發生率之第六名,而且高居男性癌症發生率的第四名(Department of Health, 2009)。2009 年的死亡原因統計資料則顯示,口腔癌在台灣癌症死亡率排名從 1981 年的第十一位,到1996 年上升至第七位,近來在 2009 年則再上升至第六位,其 中男性死亡率更是由原本的第八位上升至第四位,死亡成長率16.91%,為惡性腫 瘤之首。口腔癌的發生率及死亡率近年以來節節上升,並且超過了鼻咽癌,不僅 罹病平均年齡下降,而且每年發現的新病例及死亡人數都超過千人,已成為國人 頭頸部癌症的第一位,由此可見口腔癌對台灣人民健康威脅之嚴重性。
台灣地區口腔癌好發年齡集中於40 歲至 70 歲,平均年齡 54 歲,其中男性 52 歲、女性53 歲,兩者無顯著差別,而發生於男女之比率為 3:1,其中男性相對比較
容易罹患口腔癌,可能是由於90%以上口腔癌病患有嚼檳榔習慣,也因此顯示嚼
檳榔在台灣地區的口腔癌佔極關鍵地位。罹患口腔鱗狀上皮細胞癌的原因,一直都 是多因性,在歐美國家和日本族群中,抽煙和酗酒是口腔癌最主要的風險因子 (LaVecchiaet al., 1997);而在臺灣和東南亞國家口腔癌的高發生率與咀嚼檳榔的習 性有關。
臺灣地區檳榔的成分含有檳榔子(areca nut)、荖花(unriped fruit of Piper Linn)、荖葉(betel leaf of Piper Linn)、荖籐(Piper betle linn)、石灰(slaked lime)
及香料。檳榔子成分中的檳榔鹼(arecaidine)和檳榔素 (arecoline),對於體外培養 的人類面頰上皮細胞,有遺傳毒性致癌作用(Shirname et al., 1983; Shirname et al., 1984),並且會導致發生 DNA 受損機率增加(Dave et al., 1992),具有致突變性的 潛力。此外檳榔萃取物也會促使口腔黏膜細胞合成膠原蛋白質的量增加,使分解 及吞食膠原蛋白質的能力下降,形成過量之膠原纖維堆積,而產生口腔癌前病 變。荖花內含致癌物黃樟素(safrole),荖葉的致突變性潛力很低,荖籐則是在 長期嚼食情況下,可能產生強烈化學性刺激的亞硝基致癌物質。紅灰和石灰會使 口腔環境變成鹼性,在鹼性環境檳榔塊中一些成份會釋放出一些游離基,而游離 基易引起細胞變性。因此檳榔中複雜的成份,會使口腔產生癌前病變,像是口腔 黏膜纖維化、白斑、紅斑、疣狀增生等等,如果再加上酒精和香菸對於口腔黏膜 刺激產生的協同作用(Lee et al., 2003),很容易就會導致口腔癌的發生。
分析鱗狀細胞癌相關基因和分子的變異,在東南亞地區不同於西方地區人 口,有族群之間的差異性(Scully et al., 2000)。因此可以推測,台灣地區人口罹患 口腔鱗狀上皮細胞癌,也可能由於種族和地區的區別,存在不同的免疫調節或免 疫抑制作用。
二、口腔癌免疫抑制作用(immunosuppression)
免疫編輯(immunoediting)在致癌機轉中扮演重要角色,即是癌病變組織會引 導周圍環境,形成一個抑制型免疫反應,藉此讓癌細胞擴散(Kim et al., 2007)。免 疫編輯過程大致分為三個時期:排除癌細胞時期(elimination phase)、癌細胞和宿主 免疫系統平衡時期(equilibrium phase),以及癌細胞逃離宿主免疫系統時期(escape phase)。排除時期(elimination phase)代表免疫監測(immunosurveillance)的概念,免 疫系統去除變異的癌細胞,在平衡時期(equilibrium phase) ,則是免疫細胞和腫瘤 細胞之間互相作用,之後的逃離宿主免疫系統時期(escape phase),腫瘤細胞誘發 免疫抑制使免疫細胞死亡或是呈現不活化狀態,也可能腫瘤細胞變異或是減少免 疫性,使得只有少數的免疫細胞能辨識毒殺腫瘤細胞,造成腫瘤持續生長侵略的 環境。
近年來有研究指出免疫抑制和口腔鱗狀上皮細胞癌的發生是具有關連性,在 肝臟移植和骨髓移植的病人具有免疫抑制作用產生後,患者會有口腔白斑癌前病 變產生(Hernandez et al., 2003),甚至會導致口腔癌(Bhatia et al., 2001)。此外參與 對抗腫瘤的免疫細胞,主要有自然殺手細胞(NK cell)和毒殺型T細胞(cytotoxic T-cell),在頭頸癌轉移的病人中,發現多數的自然殺手細胞、及T細胞具有較低的 毒殺能力(Schantz et al., 1986) ,或是其功能根本被口腔癌細胞所抑制(Jewett et al., 2006; Laad et al., 1996)。而且研究顯示頭頸癌病患的周邊血液,其自然殺手細胞、
T細胞、樹突細胞以及腫瘤浸潤的淋巴細胞,皆發現有不正常訊息傳遞路徑,也 會誘發細胞自然凋亡和使細胞增生能力降低(Hoffmann et al., 2002b; Reichert et al., 2002),其可以作為免疫細胞受腫瘤抑制的指標(Hoffmann et al., 2002a)。儘管如 此,關於口腔鱗狀細胞癌免疫抑制的調節機制仍然不是非常清楚,因此癌症的免 疫抑制作用成為癌症治療的熱門研究主題。
目前的研究指出,跟癌症免疫抑制有相關的分子和細胞分為三類,(1)失去功 能的抗原呈現細胞(antigen-presenting cells),包括樹突狀細胞(dendritic cells)、巨 食細胞(macrophage)和骨髓性抑制細胞(myeloid-derived suppressor);(2)調節性T細 胞(regulatory T cells);(3)高度表現抑制性B7分子的免疫細胞(抗原呈現細胞、T和B 細胞)、腫瘤細胞(tumor cell)、基質細胞(stroma cell),這些分子和細胞被認為和腫 瘤逃避免疫系統攻擊有高度的相關性(Zou et al., 2008)。
三、調節性T細胞(regulatory T cells,Tregs)
免疫的耐受性調節可以分為兩種,分別是在初級淋巴組織(primary lymphoid organs)所進行細胞族群刪除(clonal deletion)的中樞耐受性(central tolerance),以及 在人體周邊由調節性T細胞所維持的周邊耐受性(peripheral tolerance)。調節性T細 胞(regulatory T cell,Tregs)是一群具有負調節機體免疫反應的淋巴細胞,可以維 持自體抗原的耐受性和避免免疫反應過度。這群具有負調節功能的細胞,1970年 Gershon等人曾命名為抑制T細胞(suppressor T cell),因為缺乏明確的表面標誌,
研究長期處於停滯不前的困境;直到1995年Sakaguchi等人,提出自體耐受性 (self-tolerance)主要是藉由一群CD4+ CD25+ T淋巴細胞執行,而這群細胞被稱之 為調節性T細胞(Sakaguchi et al., 1995)。2001年Bennett等人研究IPEX (immune dysregulation, polyendocrinopathy, enteropathy,X-linked)自體免疫疾病時,發現病人 有轉錄因子Foxp3的基因缺失(Bennett et al., 2001),並且在日後研究也發現Foxp3 和調節性T細胞的分化(Wan et al., 2007)、抑制功能、細胞增生以及細胞代謝(Gavin et al., 2007; Lin et al.,2007).有關聯性,而Foxp3在調節性T細胞的表現可以維持其 表現型和抑制功能 (Williams et al., 2007),因此Foxp3的表現普遍被認定是調節性 T細胞的特徵。
老鼠體內調節性T細胞的研究,目前已有三種不同的CD4調節性T細胞並被明 確的歸類,根據其表面標記特徵、產生細胞的來源和作用機制的不同,可分為 CD4+CD25+Foxp3+調節性T細胞、Tr1細胞和Th3細胞等多種亞型。根據細胞的來 源,可分為從胸腺發展而成的自然調節性T細胞CD4+CD25+ Foxp3+Treg (natural occurring Treg)(Sakaguchi, 2004),和在體表經由誘導產生的Tr1細胞和Th3細胞 (Walker et al., 2003)。自然調節性CD4+CD25+ Foxp3+Treg是目前研究較為徹底的 亞型,在正常人和小鼠的外周邊血及脾臟組織的CD4+T細胞,佔有5-10%(Zou, 2006),並且在細胞表面持續表達CD25分子(即為IL-2受體α鏈),更具有專一性 的標記因子Forkhead or winged helix of transcription factor P3,簡稱Foxp3/FOXP3
,此為轉錄因子與調節性T細胞的生成及功能有關(Hori et al., 2003),此外有研究 指出老鼠的調節性T細胞會表現GITR (glucocorticoid-induced tumour-necrosis factor (TNF)-receptor related protein)和CTLA4(cytotoxic T-lymphocyte-associated antigen 4),CTLA4會和活化T細胞上的B7分子結合,藉此抑制T細胞的活化或細 胞激素之生成,有助於研究老鼠的調節性T細胞和發展免疫治療。自然調節性T 細胞CD4+CD25+ Foxp3+其抑制的對象可能是功能性T細胞或是樹突狀細胞,其抑 制的機制可以藉由分泌抑制型細胞激素(IL-10、IL35、TGF-β)、分泌穿孔素和 顆粒酵素分解細胞、CD25競爭功能性T細胞活化所需的細胞激素IL-2以及一些未 知分子使得功能性T細胞之細胞周期停滯,另一方面自然調節性T細胞也可能藉由 CTLA4、LAG-3以及一些代謝物的形成(IDO等等),抑制樹突狀細胞的成熟和 協同刺激(Shevach et al ,2009;Vignali et al ,2008)(附圖一)。在體表誘導產生的 CD4+CD25+Treg和自然調節性T細胞表現的特徵相似,可以從非調節性的CD4+T 細胞誘導產生(Bluestone et al, 2003),第ㄧ型調節性T細胞(type 1 regulatory T cell, Tr1),為另一類亞型的CD4+Treg,Tr1不會表現或是表現低量CD25表面分子,是 屬於Foxp3-的細胞,和CD4+CD25+Treg表現的特徵不同(Maynard et al., 2007;
Strauss et al., 2007a),但能產生高量的IL-10、適量的TGF-β、少量的IL-2、IFN-γ,
但不產生IL-4 (Roncarolo et al., 2006),藉此也能與Th1和Th2之間作區別,Tr1需在 IL-10存在情況下誘導產生,細胞抑制免疫反應的主要機制,是藉由產生具有免疫 調節功能的細胞激素IL-10和TGF-β。另一種為Th3細胞,Th3型調節性T細胞,是 在研究口服耐受機制的過程中發現的,主要分泌TGF-β來調節免疫功能,對Th1 和Th2都具有抑制作用(Weiner, 2001)。
最早人類調節性T細胞的研究中,人類非小細胞肺癌也發現到CD4+CD25+調 節性T細胞,在腫瘤組織內有增加的情形(Woo et al., 2001),類似的研究也發表在 卵巢癌(Curiel et al., 2004)、乳癌及胰臟癌(Liyanage et al., 2002)、胃癌與食道癌 (Ichihara et al., 2003),且這些浸潤於腫瘤組織內的調節性T細胞,有抑制同源性T 細胞的能力(Woo et al., 2002)。Curiel等人發現在人類卵巢癌,大量CD4+CD25+
FOXP3+Treg存在腫瘤內,會抑制腫瘤抗原特異性T細胞的活化及增生,直接證實 調節性T細胞會在腫瘤不斷發展期間累積於腫瘤組織內,且這樣的調節性T細胞有 助於腫瘤生長(Curiel et al., 2004)。最近研究顯示出調節性T細胞,造成的免疫抑 制反應是一個腫瘤躲避免疫的關鍵機制,且成為主要阻礙腫瘤免疫治療的細胞 (Curiel, 2007)。在早期人類調節性T細胞的研究中,高度表現CD25的細胞
(CD25high/bright)具有較強的抑制能力(Baecher-Allan et al., 2001),約佔CD4+T細 胞的1-2%,然而健康的個體體內,不表現CD25high Foxp3high T 細胞群,且發現 有CD25low Foxp3low的細胞亞群,因此一般研究都認定CD25high Foxp3 high T細 胞為具有抑制功能的調節性T細胞。
調節性T細胞在小鼠癌症模型中已廣泛研究,並且明確定義出調節性T細胞及 其參與作用,但是在人體的免疫中,不同調節性T細胞亞群表現的細胞激素和抑 制作用,仍然沒有很明確的研究結果。近年來在頭頸部鱗狀上皮細胞癌中指出,
在腫瘤組織浸潤淋巴球中,發現有高量的CD25highFoxp3+抑制型T細胞存在
(Strauss et al., 2007b),並且指出頭頸部鱗狀上皮細胞癌病人的CD4+CD25high細胞 亞群,具有抑制T細胞活化生長的能力,且其抑制T細胞生長之能力比健康人的 CD4+CD25high細胞亞群更佳。
T淋巴球的活化及免疫功能被輔助刺激分子(co-stimulatory molecules)所調 節,CD28作為B7基因產物的接收器,主要功能為起始T細胞的免疫反應;CTLA-4 與B7基因產物有更高度的親和力,在T細胞活化後被誘導而表現,主要功能為負 調節T細胞的免疫反應;誘導性輔助刺激分子(inducible costimulatory molecule;
ICOS)屬於CD28/CTLA-4家族第三成員,主要表現在已活化的T細胞,研究顯示 ICOS-/- 細胞在體外培養失去活化以及增生的能力,此外,ICOS對於T細胞priming 以及產生Th2細胞激素扮演關鍵的角色。近期研究發現黑色素瘤病患,其腫瘤浸 潤淋巴細胞(tumor infiltrating lymphocyte,TIL)中分離出的CD4+CD25+Foxp3+調 節性T細胞會高度表現inducible costimulatory molecule(ICOS),並且證實這群 ICOS+Foxp3+nTregs其功能會相似表現IL-10的Tr1細胞(Ito et al., 2008),之後,自 然調節性T細胞更直接被區分為ICOS+Foxp3+Treg與ICOS-Foxp3+ Treg,並且在研 究中指出ICOS+Foxp3+ Treg可藉由表現IL-10抑制樹突細胞的功能以及表現 TGF-β抑制T細胞的功能,然而,ICOS-Foxp3+ Treg只會表現TGF-β。不僅是功能 上有所差異,這兩群調節性細胞在調控自身的存活與增殖上也有不同的特性。
四、抑制性B7分子(inhibitory B7 molecules)
免疫系統在正常的情況下,抗原呈現細胞(APCs)會吞噬抗原(antigen),經過組 裝 後 抗 原 呈 現 細 胞 會 將 主 要 組 織 相 容 性 複 合 體- 抗 原 複 合 物 (major histo- compatibility complex,MHC-antigen complex),呈現給T cell receptor(TCR) ( Jenkins et al ,1987),然而要完全活化T細胞,還必須靠抗原呈現細胞上的共同刺激分子 (costimulatory molecules) B7-1(CD80)和B7-2(CD86),B7-1和B7-2與T細胞表面的
CD28相互作用(Azuma and Freeman,1993),這樣的結合可以給與T細胞第二活化的 訊息,使得T細胞可以被完全活化,引發下游的免疫反應。為了避免T細胞的過度 活 化 ,T 細 胞 自 身 會 表 現 另 一 個 可 以 和 CD28 競 爭 CD80 、 CD86 的 接 受 體 CTLA-4(cytotoxic T-lymphocyte-associated antigen 4),CTLA-4會負向的調控T細胞 增生(Linsley et al,1991),所以T細胞的活化,可以由正向和負向的訊息維持巧妙 的平衡。
活化T細胞所表現的CD28及CTLA-4具有相同的配體,即是B7分子(CD80、
CD86),最近新的研究發現B7分子有基因同源性的相似分子,這些分子被歸類為 B7家族的成員,目前所被研究的B7家族有七個成員,分別為B7-1(CD80)、
B7-2(CD86)、B7h(B7-H2)、PD-L1(PD-ligand1)(B7-H1)、PD-L2(PD-ligand2) (B7-DC),以及最新成員B7-H3、B7x(B7-H4)。綜合近年來的研究將這些B7分子 以親緣關係(phylogenetic analysis)分為三類(Zang and Allison , 2007) (附圖二):第 一類包含B7-1、B7-2、B7h(ICOS-L),在傳遞共同刺激訊息扮演關鍵性的角色;
第二類則包含PD-L1(B7-H1)(CD274)和PD-L2(B7-DC) (CD273),其PD-L1, PD-L2/PD-1 路徑為共同抑制的功能,雖然其抑制功能和B7-1、B7-2/CTLA-4 pathway之差異尚未完全釐清,但已有文獻指出T細胞PD-1(programmed death-1) (CD279)和B7-H1、B7-DC的結合與周邊耐受性、腫瘤免疫、感染免疫、移植免疫、
過敏以及自體免疫疾病都有相關,PD-1的細胞質的尾端含有immunoreceptor tyrosine-based inhibitory motif (ITIM)以及immunoreceptor tyrosine switch motif (ITSM),前者是用來調控抑制訊號,後者則是負責PD-1與PD-L1結合後的訊號。
PD-1可表現在CD4+ T細胞,CD8+ T細胞,自然殺手T細胞,B細胞等;第三類包 含B7-H3、B7x(B7-H4),其相對應之受體尚待研究發現,其功能較偏向為共同抑 制;第三類包含B7-H3、B7x(B7-H4),其相對應之受體尚待研究發現,其功能較 偏向為共同抑制。
1992年有研究在細胞凋亡細胞株發現PD-1是50–55-kDa第一型穿膜醣蛋 白,和CTLA-4、CD28有33%基因序列相等(Ishida et al,1992),之後在自體免疫疾 病的研究也證實PD-1和細胞凋亡的直接關連性(Nishimura et al., 1998) 。1999年發 現和CD28相似的分子ICOS(Swallow et al,1999),同年Dong也發現新的B7基因同 源性分子,命名為B7-H1,在這項研究中指出B7-H1會表現在人類活化的巨蝕細 胞、T細胞、B細胞,然而B7-H1的接受體並非已知的分子CTLA-4、CD28和ICOS,
在其功能方面發現B7-H1的確會影響T細胞的增生以及促進IL-10的產生,此外隔 年也發現了ICOS-L(B7-H2),也歸類為B7家族的新成員。2000年Freeman發現,
B7-H1的接受體是T細胞傳遞細胞凋亡訊息的PD-1(programmed death-1),所以 B7-H1也稱為PD ligand-1(PD-L1),此外在隔年也發現PD-1的第二個配體,命名為 PD ligand-2(B7-DC) (Latchman et al,2001 ),並且發現B7-DC只表現在樹突狀細胞 和巨蝕細胞 (Tseng et al,2001),PD-L1和PD-L2都是第一型穿膜醣蛋白,並且有 20%胺基酸和B7-1、B7-2相等。2001年同時也選殖出其他B7家族成員B7-H3 (Chapoval et al, 2001),而在2003年也發現了B7-H4(Sica et al, 2003),但是其接受 體仍然還在研究當中 。
近年來研究指出B7-H1除了會表現在活化的T細胞、B細胞、樹突狀細胞、
單核球/巨蝕細胞,也會表現在癌細胞(Dong et al,2002)、自然殺手細胞、活化的內 皮細胞、間質幹細胞以及細胞培養的骨髓性肥大細胞(Zou et al, 2008) ,並且有研 究指出IFN-γ會促進B7-H1在上皮細胞、內皮細胞、骨髓性細胞和癌細胞的表現 (Dong et al,2002),對於細胞本身可能有保護作用。多數的研究指出表現在癌細胞 和骨髓性細胞的B7-H1會促進T細胞細胞凋亡和抑制T細胞的活化和增生(Dong et al,2002 ; Curiel et al,2003),可能是由於B7-H1和PD-1的結合而抑制了T細胞接受體 的活化訊息傳遞(Freeman et al,2000),但也有少數的研究指出B7-H1的抑制功能是 不需要接受體PD-1的,其藉由CD80來傳遞抑制T細胞活化的訊息(Butte et al,2007)
。此外也有研究指出B7-H1的表現是細胞訊息傳遞的接受體,對於動物體內表現 B7-H1的癌細胞自身而言,有抗細胞凋亡的保護作用(Azuma et al,2008),還有動 物實驗研究指出天真T細胞表現的B7-H1和樹突狀細胞表現共同刺激分子有相關 (Talay et al,2009),因此B7-H1是一個在多種不同細胞都會表現並且是具有多種不 同功能的B7家族一員。然而對於B7-H1表現和功能的研究,一般是在癌細胞或是 抗原呈現細胞,對於B7-H1表現在T細胞的研究和其詳細的功能,仍然還是屬於比 較少被研究探討的。
對於B7-H1分子在人類癌症的研究探討,乳癌(Ghebeh et al,2006)、大腸癌、
胃癌(Wu, et al,2006)、肺癌(Konishi et al,2004)、卵巢癌(Hamanishi et al,.2007)、胰 臟癌(Nomi et al,2007)、腎細胞癌(Thompson et al,2006)、以及尿道癌(Nakanishi et al,2007)等等,在很多癌症組織都發現有高量B7-H1表現,並且經過統計發現B7-H1 表現量和腫瘤的大小、病程、轉移、預後、危險因子都有關聯性。其中在乳癌的 研究中發現,B7-H1在TIL(tumor infiltrate lymphocyte )中的表現和腫瘤大小、病程 以及腫瘤指標HER2表現有相關(Ghebeh et al,2006),並且在乳癌腫瘤組織切片中 可以發現Foxp3+調節性T細胞和表現B7-H1的T細胞共同浸潤到腫瘤組織(Ghebeh et al,2008)。另一方面在胰臟癌的研究指出,B7-H1的表現在胰臟癌病人腫瘤組織 相對於健康正常人胰臟高許多,而有B7-H1+的腫瘤組織也具有較高量的Foxp3+
調節性T細胞存在(Loos et al,2008),並且發現B7-H1和細胞激素IL-10在腫瘤組織 的表現量呈現正相關性(Geng et al,2008)。B7-H1表現在T細胞的研究還不是非常 多,2003年有類風濕性關節炎研究指出,T細胞上表現的B7-H1會接收自體抗體的 訊息,誘導T細胞產生細胞凋亡(Dong et al,2003),後來表現在T細胞上的B7-H1也 被認定具有抑制T細胞活化的功能(Seo et al,2006),並且在乳癌研究發現T細胞上 的B7-H1和一些危險因子都有相關性。此外有更進一步非霍奇金氏淋巴瘤(non- Hodgkin's lymphomas)和移植免疫的研究指出,表現在調節性T細胞上的B7-H1,
具有抑制T細胞的活化增生的功能(Yang et al,2006) (Kitazawa et al,2007)。
在許多不同腫瘤組織的研究發現,B7-H1的表現對於免疫抑制作用是具有相 關性的,並且可能和Foxp3+調節性T細胞的免疫調節作用也有關聯性。此外B7-DC 在肺癌(Konishi et al,2004)、卵巢癌(Hamanishi et al,2007)和胰臟癌(Nomi et al,2007) 被發現,屬於比較少在癌症中被發現的B7抑制性分子,另一個B7-H4則是會表現 在女性生殖道上皮細胞、腎臟、肺臟、胰臟等少數人體器官,卵巢癌的研究發現 有高量的B7-H4表現在巨蝕細胞(Kryczek et al.,2006a)以及在病人血清中也有低量 的B7-H4表現(Simon et al,2006),並且發現巨食細胞的B7-H4會降低抗原特異性T 細胞的活化增生,更進一步有研究指出卵巢癌調節性T細胞會促進巨食細胞分泌 IL-6和IL-10,而IL-6和IL-10會刺激巨食細胞自身B7-H4的表現,這些在腫瘤微 環境內大量表現B7-H4的巨食細胞,會直接抑制T細胞之細胞週期(Kryczek et al.,2006b) 。這些不同的B7抑制性分子PD-L1(B7-H1)、(B7-DC)和B7x(B7-H4),
可能在腫瘤微環境當中不同細胞上表現並且以不同的機制造促進免疫抑制,使腫 瘤可以逃避宿主免疫攻擊並且不斷生長,至於這些B7抑制性分子在腫瘤微環境中 所表現的是哪些細胞和其真正複雜免疫機制都還需要更進一步的探討和研究。
第二節、研究動機與目的
腫瘤細胞可能藉由不同方式來逃避宿主免疫反應,由本實驗室先前的研究結 果發現,口腔鱗狀細胞癌病患之腫瘤組織內浸潤CD3+CD4+之淋巴細胞中,
CD4+CD25+T 細 胞 佔 所 有 CD4+T 細 胞 比 例 有 22.81% , 而 病 患 的 周 邊 血 液 CD4+CD25+T細胞只有7.8%,因此存在於腫瘤組織內的CD4+CD25+T細胞,相對 於該病患的周邊血液CD4+CD25+T細胞比例高許多。存在於口腔鱗狀細胞癌組織 內的CD4+CD25+Foxp3+之調節性T細胞,大約有16.19%,而在病患的周邊血液 中 , 偵 測 到 比 例 大 約3.67% 的 CD4+CD25+Foxp3+ 之 調 節 性 T 細 胞 , 高 量 的 CD4+CD25+ Foxp3+之調節性T細胞存在口腔癌腫瘤組織內,且進一步研究發現 CD4+CD25high的調節性T細胞具有抑制T細胞活化增生的能力。
過去有研究指出,人類口腔癌細胞株和人類口腔鱗狀癌組織有B7-H1的大量 表現(Tsushima et al,2006),因此本論文中想要探討B7-H1分子在口腔鱗狀細胞癌 腫瘤組織之浸潤淋巴球的分布和表現,而浸潤在腫瘤組織中的調節性T細胞其免 疫抑制功能,是不是有可能和B7-H1的表現有相關性。若能釐清口腔鱗狀細胞癌 腫瘤組織中B7-H1和調節性T細胞免疫抑制的相關機制,未來可能將相關的調控分 子,作為治療時的標的,進而達到作為病患治療預後的目標,對於建立人類口腔 癌免疫療法,將有助於提供重要且必要性之訊息。
第二章、實驗方法及材料
第一節、檢體的收集及來源
本研究將以十七例口腔癌病人之腫瘤組織和周邊血液為研究材料,並蒐集健 康正常人之口腔組織和周邊血液作為對照比較,其中有十五名為男性,女性只有 兩名,病患的年齡從二十九歲至七十三歲。口腔鱗狀細胞癌的病患,在癌症分期 手術後,其口腔鱗狀上皮細胞癌組織收集成實驗組。每個病例的癌症分期(surgical stage),癌症病理分級(histological stage),淋巴或血管侵犯都會被紀錄,其中口腔 癌一般較容易轉移的部位除了頸部淋巴結外還包括肺臟、肝臟及骨骼。根據1997 年美國癌症醫學會(AJCC)的口腔癌分期的定義為標準:
零期:即原位癌。腫瘤細胞局限在口腔黏膜上皮內
第一期:腫瘤的最長徑小於或等於2公分且無頸部淋巴結及遠端轉移
第二期:腫瘤的最長徑大於2公分但不超過4公分,且無頸部淋巴結及遠端轉移 第三期:腫瘤的最長徑大於4公分或已轉移到同側頸部一個淋巴結,
此淋巴結之最長徑不超過3公分
第四期:有以下任何一種情形包括: 1.腫瘤侵犯鄰近的組織(如:穿過骨外層,
深入深層肌肉、上頷竇、皮膚等); 2.頸部淋巴結轉移的數目超過1個(不 論是在原發病灶的同側、對側或兩側都有),或是淋巴結的最大徑已超過 3公分; 3.無論腫瘤大小及淋巴結是否轉移,已發生遠處轉移者。
在開刀前,必須有完整且詳細的記錄記載病人過去病史,並且符合下列基本的要 求,才會納入本實驗之收集樣本中;第一為所有病患在接受手術前,皆有病理切 片的術前口腔臨狀上皮細胞癌之證明;第二為病患沒有明顯且嚴重局部發炎現 象;第三為術前沒有接受特殊非手術性的治療。納入本實驗的病患都必須接受檢 查,且沒有其他伴隨的併發症或疾病,且皆不患有人類後天免疫不全疾病的證 明,同時願意參與本實驗且簽署同意書。手術進行後,手術標本都會接受病理科 醫師的診斷,來排除其他有可能共存的惡性腫瘤。
第二節 、人類口腔鱗狀上皮細胞癌組織內淋巴球的分離與純化
人類口腔癌組織內淋巴球的分離方法,是根據1997年Bor-Ching Sheu等人,
所發表的方法加以改良(Sheu et al., 1997)。台大醫院接受手術治療的病人中,以無 菌方法收集口腔鱗狀細胞癌組織,並利用phosphate-buffered saline (PBS)將其上血 塊 沖 洗 乾 淨 , 之 後 用 刀 片 切 成 碎 片 , 加 入RPMI-1640 medium (Gibco, Life Technologies, Grand Island, NY, USA)研磨後,再以380 μm及45.7 μm篩孔之濾網分 別過濾,濾液以1200 rpm (RS-720, KUBOTA)離心10分鐘,再以1300 rpm (RS-720, KUBOTA)離心10分鐘,以不同濃度的Percoll solution分層(含20 %, 55 %, 100 %),
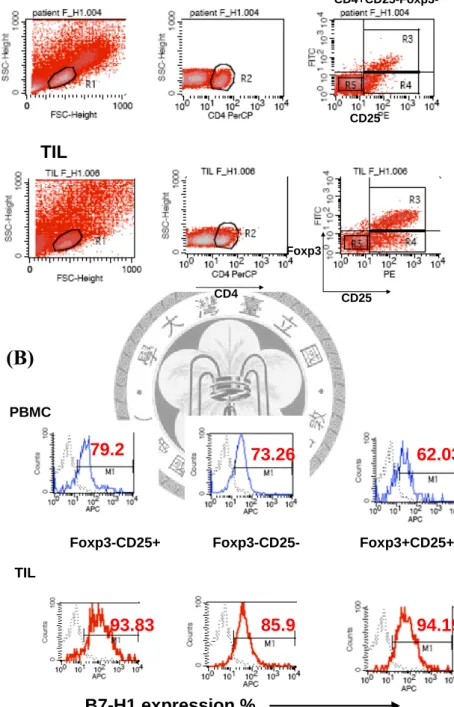
單核細胞球(mononuclear cell)會位於55 %及100 %的Percoll solution之間,而癌細 胞會位於20 %及55 %的Percoll solution之間,以此方法將口腔癌細胞和腫瘤內浸 潤淋巴球分離。將分離出來的腫瘤內浸潤單核細胞球,以anti-CD3、anti-CD4單 株抗體做螢光染色,並進行流式細胞儀(BD FACS Calibur)分析,藉由細胞的大小
(與FSC成正比),顆粒性(與SSC成正比)與CD3、CD4的標定來確定不同亞群 T淋巴球的分布。
第三節、人類周邊血液單核球的分離與純化
從醫院接受手術治療的病人中收集血液標本,於手術進行前抽取20cc 靜脈 血,置於含有肝素(heparin)的採血管中,先以 3000 rpm 離心 20 分鐘(RS-720, KUBOTA),吸取上層的血漿後,加入等量的 Hank's Buffered Salt Solution
(HBSS),再用 Ficoll hypaque (1.077 density,Pharmacia Biotech Co.)分離出周邊血 液中的單核細胞球(peripheral blood mononuclear cells, PBMCs),分離出的單核細 胞球懸浮於RPMI-1640 medium (Gibco, Life Technologies, Grand Island, NY, USA),使其濃度為 1×106 cells/ml。將分離出來的腫瘤內浸潤單核細胞球,以 anti-CD3、anti-CD4 單株抗體做螢光染色,並進行流式細胞儀(BD FACS Calibur)
分析,藉由細胞的大小(與FSC 成正比),顆粒性(與SSC 成正比)與 CD3、CD4 的標定來確定不同亞群T 淋巴球的分布。
第四節 細胞表面染色
將分離出的周邊血液和腫瘤組織內浸潤淋巴球,分別以螢光標定如下:
anti-CD4-FITC、anti-CD25-PE、anti- CD3-PerCP,以及標定 biotin-B7-H1(MIH1)、
biotin-B7-PD-1(MIH4)並以老鼠之 strepavidin-APC 當做第二級抗體(eBioscience, San Diego, Calif.), IgG1κ isotype control-APC(eBioscience, San Diego, Calif.)做為 對照,並以CellQuest software (Beckton-Dickinson Inc.)進行資料分析。
第五節 細胞內染色
為了偵測調節性T 細胞的轉錄因子 Foxp3,將分離出來的周邊血液單核細 胞,先分別加入anti-CD4-PerCP、anti-CD25-PE,以及標定 biotin-B7-H1(MIH1)、
biotin-B7-PD-1(MIH4)並以老鼠之 strepavidin-APC 當做第二級抗體(eBioscience, San Diego, Calif.), IgG1κ isotype control-APC(eBioscience, San Diego, Calif.)做為 對照,混合均勻後,於4°C 避光作用30 分鐘,利用 Cytofix/Cytoperm buffer (eBioscience, San Diego, Calif.)在冰上作用一小時,再利用一倍的 permeabilization buffer (eBioscience, San Diego, Calif.)連續三次沖洗細胞,使細胞固定及破壞細胞 膜讓通透性增加,在染胞內分子前,先使用2%胎牛血清(fetal calf serum)結合非特 異性抗原,再藉由anti-Foxp3單株抗體(eBioscience, San Diego, Calif.)偵測調節性 T 細胞,以流式细胞儀進行分析。以FSC、SSC、CD4定義出 CD4+淋巴球细胞,觀 察不同T 細胞亞群表現 B7-H1和 PD-1的百分比和表現強度。
第六節 腫瘤組織和正常組織之 RNA 萃取
將約 0.5mm3大小之腫瘤組織或口腔內非腫瘤組織磨碎後,加入Solution D (4 M Guanidinium iso-thiocyanate、25 mM Sodium citrate pH 7.0、0.5% Sarcosyl、0.1 M b-Mercaptoethanol),搖晃均勻後加入 2 M Sodium Acetate pH 4.0、Acid phenol (Water saturated)、Chloroform:isoamyl (49:1),搖晃均勻後冰上作用 20 分鐘,9000 rpm 4 度 C 20 分鐘,取上清液加入 1 倍體積 Ispropanol-20 度 C 沉澱一小時,9000 rpm 4 度 C 20 分鐘,Pellet 加 Solution D 0.5 ml,並加入 0.5 ml Ispropanol-20 度 C。
經由光譜分析儀(Ultrospec 3300 pro, Amersham Bioscience )定量 total RNA 後取 2 μg RNA 以 RQ1 DNase (Promega)於 65℃中作用 30 分鐘,進行 DNA 清除,進一 步利用M-MLV Reverse Transcriptase (Promega)以及 oligo(dT)於 37℃中作用 1 小 時以進行反轉錄的動作。
第七節 即時定量聚合酶連鎖反應(real-time PCR)
本實驗中所使用之 primers 是由 Primer express v3.0所設計,包括 B7-H1、
PD-1、IL-10、IFN-γ、Foxp3、CD4、B7-H4、B7-DC。cDNA 利用 POWER SYBR GREEN PCR Master Mix(Applied Biosystem)於 Applied Biosystem 7500 system 中 進行同步定量聚合酶連鎖反應。結果之不同基因Ct 值將以 GAPDH Ct 值作為基 準,使用公式如下:Relative expression = (2-dCt)。
第八節 口腔癌組織及正常組織的組織包埋和冷凍切片
口腔鱗狀上皮細胞癌腫瘤組織和正常組織經收集後,以Optimal Cutting
Temperature(O.C.T.) Tissue-Tek (Sakura, Tokyo, Japan)包埋組織,並存放於-80 度 C 之環境中,待組織冷凍後將組織切成厚度5μm 之組織切片,並將組織切片存放於 -80 度 C。
第九節 組織免疫染色(Immunohistochemistry)
組織冷凍切片和丙酮(acetone)反應 10 分鐘固定組織,用二次水(ddH2O)、
1XPBS wash 後,以 3% 過氧化氫(hydrogen peroxide)和組織切片反應 20 分鐘去除 內生性過氧化酵素活性(peroxidase activity),1X PBS wash 完後用疏水性(hydrphbic pen)筆劃出組織範圍,再以 10% goat serum blocking(Zymed Laboratories,histostain bulk kit) 20 分鐘,避免非特異性抗體和組織的結合,接著加入 1:50 的 mouse anti-human 抗體 anti-MIH1(B7-H1)、anti-MIH4(PD-1)、1:100 anti-Foxp3(eBioscience, San Diego, Calif.),在 4 度 C 反應 overnight,1X PBS wash 後加入 anti-mouse biotinylated secondary Ab(Zymed Laboratories,histostain bulk kit)室溫作用 20 分 鐘,1X PBS wash 後再加入 strepavidin-peroxidase conjugate(Zymed Laboratories ,histostain bulk kit)室溫作用 20 分鐘,1X PBS wash 後加入 DAB(Dako)呈色,並以 Mayer’s hematoxylin 做複染劑 5 分鐘,running water 之後經 50-75-85-95%酒精 10 次、100%酒精將組織脫水,最後將組織切片浸入 xylene 並以膠封片,最後在顯 微鏡下觀察。雙分子染色則是以1:100 mouse anti-human-Foxp-3 染色 24 小時之 後,以DAB(Dako)呈色,再以 1:50 的 mouse anti-human 抗體 anti-MIH1
(B7-H1),最後用 AEC(BiogeneX)呈色,再進行複染。
第十節、反應性T細胞及調節性T細胞篩選方法及細胞增生和抑制試驗
20 c.c.靜脈血,置於含有肝素(heparin)的採血管中,首先分離周邊血液的單核 球,進行anti-CD3-PerCP、anti-CD4-FITC、anti-CD25-PE 染色,利用流式細胞分 選儀(BD FACSAria Cell Sorter)分選出 CD3+CD4+CD25-細胞,即為反應細胞 (responder cells),CD3-細胞即為 B 細胞和單核球細胞,作為抗原呈現細胞(APC),
CD4+CD25high 細胞作為抑制細胞,將分選出的反應細胞(CD3+CD4+CD25-)標定 carboxyfluorescein diacetate succinimidyl ester (CFSE),放置 1x105細胞於96-well plate,再加入 RPMI-1640 medium (Gibco, Life Technologies, Grand Island, NY,
USA),以 plate-bound anti-CD3 (加入 100 μl 的 2.5 μg/ml anti-CD3 於 96 well 盤中,
於37℃作用 2 個小時後,以 PBS 清洗兩次後,即可加入細胞作用)和抗原呈現細 胞(APC)等刺激細胞增生,CD25high 細胞和 B7-H1 Ab preincubate 以及不和 B7-H1 Ab preincubate 兩組細胞,分別和有 CFSE 標定的反應 T 細胞以 1:1 或是 1:2 歷經 84-120 小時後,再利用流式細胞儀分析增生結果,藉此評估調節性 T 細胞,抑制 同源的CD4+CD25-T 細胞增生的能力和 B7-H1 分子是否相關。
第十一節、周邊血液單核球與口腔鱗狀癌細胞株在體外共同培養方法
20 c.c.靜脈血,置於含有肝素(heparin)的採血管中,放置1×106細胞於12-well plate,再放入照放線或不照放射線兩組2×105的口腔鱗狀癌細胞株SAS,一起培養 五天,培養後的細胞加入biotin-B7-H1(MIH1),且以老鼠之strepavidin-APC當做第 二級抗體(eBioscience, San Diego, Calif.), IgG1κ isotype control-APC(eBioscience, San Diego, Calif.) 做 為 對 照 , 並 分 別 加 入 anti-CD4 、 anti-CD25 及 單 株 抗 體 (Becton-Dickinson Immunocytometry Systems, Becton-Dickinson, San Jose, CA)和染 細胞內Foxp3染色(eBioscience,San Diego, Calif.),以流式细胞儀進行分析,利用 CellQueast software (BD Biosciences)整理分析實驗結果,觀察表現CD4+CD25+ T 細胞以及CD4+CD25+Foxp3+調節性T細胞表現B7-H1所佔的百分比和平均表現強 度。
第三章、結果
第一節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤CD4+CD25+淋巴 球,其B7-H1共同抑制分子的細胞表現百分比和表現強度
為了探討浸潤於口腔鱗狀上皮細胞癌中之淋巴細胞的特性,採用機械式磨碎
分離方法,將口腔癌組織中的腫瘤浸潤淋巴球(tumor-infiltrating lymphocyte,TIL) 分離出來,以PBMC當做對照組再藉由螢光免疫染色及流式細胞儀分析,圖一A 以其中一個病例顯示分析的方式,先以細胞的大小(與FSC成正比),顆粒性(與 SSC成正比)圈選出淋巴細胞(為R1),再與CD3+和CD4+的標定來確定T淋巴球的 分布,分析CD4+CD25+T細胞表現共同抑制分子B7-H1之比例。圖一B顯示兩個不 同病患,其CD4+CD25+細胞表現較高比例和較低比例的B7-H1分子,其中腫瘤浸 潤的B7-H1+CD4+CD25+T細胞,佔所有CD4+CD25+T細胞的比例(百分比%)和表 現量(平均螢光值MFI)都比周邊血液稍高。但是經過圖二的統計,腫瘤浸潤 B7-H1+CD4+CD25+T 細 胞 佔 所 有 CD4+CD25+T 細 胞 比 例 78.25%(MFI=64.08) (n=17),而病患的周邊血液只有64.76%( MFI=47.14) (n=17) (百分比例% p=0.09 , 平均螢光值MFI p=0.05),雖然在平均比例和平均表現量上腫瘤和周邊血液仍有 些微的不同,但是並沒有達到統計上的顯著差異(p<0.05)。
第二節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤
CD4+T 細胞,
B7-H1 和 Foxp3、CD25 之表現有相關性
利用流式細胞儀分析,將腫瘤浸潤並同時表現 CD4 及 CD25 的 T 細胞群,
根據細胞 CD25 的螢光強度,將細胞分為三群,CD25 表現螢光強度較高的細胞
群定義為 CD4+CD25high 細胞,將 CD25 表現螢光強度較低的細胞群定義為 CD4+CD25low 細胞,而另一群表現 CD25 螢光強度介於上述兩者中間,我們定 義為 CD4+CD25intermediate 細胞,上述細胞亞群分群以圖三 A 左圖表示。再將 分出的三群細胞,統計各亞群細胞表現B7-H1 之比例,結果以圖三 A 右圖顯示,
在 口 腔 癌 腫 瘤 組 織 內 ,CD4+CD25high 細 胞 亞 群 有 85.62% 表 現 B7-H1 , CD4+CD25intermediate 有 77.91%而 CD4+CD25low 有 73.05%(n=17),顯示 B7-H1 在三細胞亞群的平均表現比例和 CD25 的表現量(螢光值)有些微的相關性,但是 並沒有達到統計上的顯著差異(CD4+CD25high 和 CD4+CD25low p=0.07)。
過去有研究指出 CD25highFoxp3+調節性 T 細胞(Strauss et al., 2007b)具有抑 制T 細胞活化增生的功能,並且 CD25 和 Foxp3 具有正相關,為了分析 CD25、
Foxp3 和 B7-H1 的相關性,利用細胞內 Foxp3 以及細胞外 CD25、B7-H1 的免疫 螢光染色和流式細胞儀,圖三 B 將腫瘤浸潤 CD4+細胞分為 CD4+CD25high 和 CD4+CD25-兩群,發現 CD4+CD25high 表現較高量的 Foxp3 和 B7-H1,而 CD4+CD25-則是不表現 Foxp3 以及表現較低量的和 B7-H1。
第三節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤
CD4+Foxp3+調節 性 T 細胞表現較高比例且較高量的 B7-H1,且 B7-H1+
Foxp3+細胞和 Foxp3+細胞有相關性
由於研究觀察到 B7-H1 和 Foxp3 表現的相關性,更進一步想研究腫瘤組織和 周邊血液Foxp3+調節性 T 細胞表現 B7-H1 的比例和表現量,以細胞內外染色和 流式細胞儀偵測Foxp3 細胞表現 B7-H1 百分比和表現強度,並以健康成人周邊血 液當做對照,圖四A 可以看到選取 CD4+Foxp3+細胞的方式,圖四 B 為其中一個
病人的B7-H1 表現百分比和表現強度。圖五為經過統計結果顯示,在腫瘤組織浸 潤CD4+Foxp3+調節性 T 細胞所表現 B7-H1 的細胞比例 85.47% ,比病人 PBMC 63.20%(*p=0.005)和正常人 PBMC68.15%(**p=0.004)都來的高(n=12),而腫瘤組織 浸潤 CD4+Foxp3+調節性 T 細胞之 B7-H1 表現量(MFI=58.21)比正常人 PBMC 27.306(**p=0.001)和病人 PBMC 34.29 (*p=0.006)都高(n=13) 。
研究指出 B7-H1+CD4+Foxp3+調節性 T 細胞在腫瘤組織所佔的百分相對於周 邊比較高,因此認為B7-H1+CD4+Foxp3+調節性 T 細胞也許對於腫瘤生長或逃避 宿主免疫攻擊是非常重要的,利用流式細胞儀分析B7-H1+Foxp3+細胞和 Foxp3+
細胞在TIL 和 CD4+ T 細胞內所佔的百分比,圖六 A 我們可以統計出結果並發現 B7-H1+Foxp3+和 Foxp3+細胞之百分比在 TIL 呈現中度正相關(R=0.61),而圖六 B 在CD4+T 細胞內則是呈現高度正相關(R=0.75)。
第四節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤
CD4+CD25+
Foxp3+調節性 T 細胞表現較高比例且較高量的 B7-H1
為了釐清 CD4+CD25+Foxp3+調節性 T 細胞在腫瘤組織所扮演的角色,利用 細胞內和外螢光免疫螢光染色,分析 CD4+CD25+Foxp3+調節性 T 細胞、
CD4+CD25+Foxp3-活化 T 細胞、CD4+CD25-Foxp3-未活化 T 細胞,三細胞群 B7-H1 的表現百分比和表現量,並以病人和正常周邊血液作為對照,圖七 A 為細 胞選取的方式,圖七 B 為其中一個病人之三群細胞 B7-H1 的表現百分比和表現
量,圖八為經過統計後三群細胞的表現百分比和表現量,圖八 A 腫瘤組織內
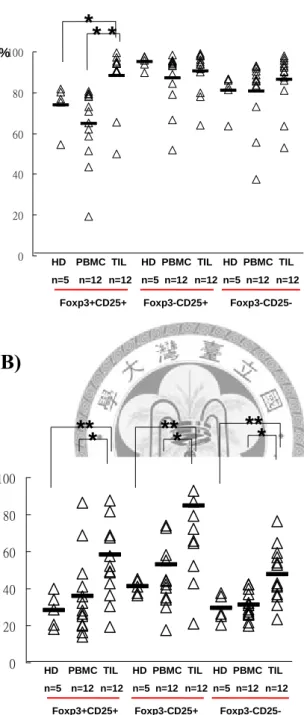
CD4+CD25+Foxp3+ 調 節 性 T 細 胞 有 88.08% 表 現 B7-H1 , 比 起 病 人 PBMC(64.71%)(**p= 0.001) 和 正 常 人 PBMC(73.78%)(*p=0.04) 都 高 許 多 , 在 CD4+CD25+Foxp3-活化 T 細胞、CD4+CD25-Foxp3-未活化 T 細胞則是沒有看到
顯著差異性。圖八B 腫瘤組織內 CD4+CD25+Foxp3+調節性 T 細胞有表現 B7-H1 之MFI=58.24,比起病人 PBMC(35.7)(*p= 0.02)和正常人 PBMC(28.28)(**p=0.002) 都高許多,而CD4+CD25+Foxp3-活化 T 細胞、CD4+CD25-Foxp3-未活化 T 細胞 的B7-H1 表現量也都有類似的情形。
第五節、口腔鱗狀細胞癌腫瘤組織
CD4+CD25+Foxp3-和
CD4+CD25-Foxp3- T 細胞表現較高量的 PD-1,且 CD25+
Foxp3+B7-H1+ T 細胞和 Foxp3-PD-1+ T 細胞有相關性
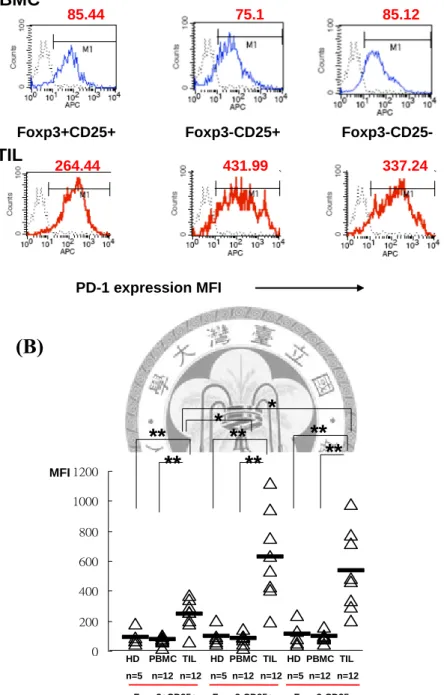
為了可以更進一步證實 B7-H1+CD4+Foxp3+調節性 T 細胞在腫瘤組織內的重 要性,利用細胞內和外螢光免疫螢光染色,分析CD4+CD25+Foxp3+調節性 T 細 胞、未活化T 細胞,三細胞群 PD-1 的表現量,並以病人和正常周邊血液作為對 照,圖九A 為其中一個病人之三群細胞 PD-1 的表現百分比和表現量,圖九 B 為 經過統計後三群細胞的表現百分比和表現量,腫瘤組織內 CD4+CD25+Foxp3-的 PD-1 平均表現量 MFI=629.25 ,而 CD4+CD25-Foxp3-平均表現量 MFI= 534.37,
兩群細胞比 CD4+CD25+Foxp3+調節性 T 細胞 MFI=250.19 高出許多(*p=0.02),
其中 CD4+CD25+Foxp3+調節性 T 細胞、CD4+CD25+Foxp3-活化 T 細胞、
CD4+CD25-Foxp3-未活化 T 細胞在腫瘤組織的 PD-1 表現量比病人和正常人 PBMC 都高許多,腫瘤組織內 CD4+CD25+Foxp3- 活化 T 細胞在 TIL 之 B7-H1 接受體PD-1 平均表現量大約是 PBMC 的六倍,CD4+CD25-Foxp3- T 細胞在 TIL 之PD-1 平均表現量大約是 PBMC 的五倍,然而 CD4+CD25+Foxp3+調節性 T 在 TIL 之 PD-1 平均表現量大約只有 PBMC 的二倍。
由於觀察到腫瘤組織調節性 T 細胞表現 B7-H1 和非調節性 T 細胞表現 PD-1 的相對關係,藉由螢光免疫染色和流式細胞儀分析後發現,圖十A B7-H1+Foxp3+
和PD-1 在 TIL(r=0.74)和 CD4+T 細胞(r=0.70)都具有高度正相關,圖十 B 並且發 現B7-H1+Foxp3+表現百分比和 Foxp3-細胞 PD-1 表現強度在腫瘤組織 CD4+T 細 胞(r=0.6)具有中度相關性。
此外我們利用口腔鱗狀細胞癌病人取下的腫瘤組織,進行組織免疫染色,圖 十一可以看到在口腔鱗狀細胞癌的腫瘤組織有B7-H1+CD4+Foxp3+調節性 T 細胞 的存在,並且也有表現B7-H1 的接受體 PD-1 的浸潤細胞。
第六節、口腔鱗狀細胞癌細胞株
SAS 誘導 CD4+CD25+表現 B7-H1
為了證實在口腔鱗狀細胞癌的腫瘤微環境會誘導CD4+CD25+ Foxp3+調節性 T 細胞表現 B7-H1,利用體外細胞共同培養的系統,放置 PBMC 1×106 細胞於 12-well plate,再放入不同細胞比例的口腔鱗狀細胞癌細胞株 SAS,或是照射過放 射線的SAS 細胞共同培養於 RPMI medium 中,一起培養五天,觀察 CD4+CD25+
T 細胞以及 CD4+CD25+Foxp3+調節性 T 細胞表現 B7-H1 之比例有無增加。 圖 十二顯示流式細胞儀資料分析後的結果以及細胞選取的方式,圖十二可以看到當 PBMC 單獨培養時,CD4+CD25+表現 B7-H1 的比例大約是 20.7%,而加入 1/5 細 胞數的口腔鱗狀細胞癌細胞株SAS 共同培養時,CD4+CD25+表現 B7-H1 的比例 上升到 56.25%,若是加入 1/5 細胞數的放射線照射口腔鱗狀細胞癌細胞株 SAS 共同培養時,CD4+CD25+表現 B7-H1 的比例上升到 50.27%。
第七節、口腔鱗狀細胞癌病患分離出之腫瘤內浸潤
B7-H1+CD4+
CD25+ Foxp3+細胞表現較高比例且較高量的 ICOS
為 了 進 一 步 了 解 B7-H1+CD4+CD25+ Foxp3+ 的 細 胞 特 性 , 我 們 想 B7-H1+CD4+CD25+ Foxp3+的表現型,有研究指出 ICOS+FOXP3+ Treg 可藉由表 現 IL-10 抑制樹突細胞的功能 (Ito et al., 2008),圖十三是 B7-H1+和 ICOS 在 CD4+Foxp3+細胞群的表現關聯性和細胞選取的方式,可以發現 ICOS+FOXP3+
Treg 表現較高比例且較高量的 B7-H1,在 n=3 統計雖然百分比只有 10%的差異 性,而表現量有兩倍的差異性,由於個體數目和範圍的限制,並沒有達到統計上 的意義。
第八節、腫瘤組織表現較高量B7-H1、Foxp3、IL-10、IFN-γ、CD4、
PD-1、B7-DC和B7-H4之RNA,且Foxp3和B7-H1有相關性
為了更證實B7-H1和Foxp3表現的相關性,我們萃取出腫瘤組織中的RNA,以 同一個病人的非腫瘤口腔組織作為對照,並且進行B7-H1、Foxp3和其他分子的即 時聚合酵素連鎖反應,發現腫瘤組織所表現的B7-H1(0.0081) 和Foxp3 (0.0195),
都比非腫瘤組織高(0.0009)(0.0002),B7-H1大約是九倍而Foxp3則是兩百倍,並且 統計十個不同口腔癌病人腫瘤組織,圖十四B結果顯示B7-H1和Foxp3的表現量具 有中度相關(r=0.54) 。圖十四A腫瘤組織的RNA可以看到IL-10、IFN-γ、CD4、
PD-1、B7-DC、B7-H4、Foxp3和B7-H1和的表現量都比正常組織高出許多。
第九節、口腔鱗狀細胞癌病患分離出調節性T細胞具有抑制細胞增生 的功能
為了證實B7-H1+CD4+CD25+ Foxp3+具有抑制T細胞增生的功能,並且可能 有一部分是藉由B7-H1而抑制T細胞增生,圖十五A將細胞刺激五天後我們比較中
和B7-H1的CD4+CD25hi抑制功能是否和CD4+CD25hi有所差異性,研究發現單獨 只有responder cell(CD4+CD25-)加入CD3抗體和APC(CD3-)刺激,可以看到64.43%
的細胞增生,當加入1:1的CD4+CD25hi細胞時,可以看到28.94%的細胞增生,
PBMC的調節性T細胞達到50%以上的抑制效果,如果先和B7-H1抗體結合的 CD4+CD25hi則是看到21.61%的細胞增生,和CD4+CD25hi細胞抑制效果沒有太大 差異性。圖十五B當刺激三天時可以看到19.72%的細胞增生,當加入CD4+CD25hi 細胞時可以看到10.47%的細胞增生,而先和B7-H1抗體結合的CD4+CD25hi細胞 則是有10.29%的細胞增生,和CD4+CD25hi細胞的抑制效果仍然沒有太大的差異 性。
第四章、討論
B7-H1在許多惡性腫瘤都被研究過,包括卵巢癌,結腸癌,頭頸部的鱗狀細 胞癌(Strome et al. 2003),非小細胞肺癌(Konishi et al. 2004),腎細胞癌(Thompson et al. 2004),食道癌(Ohigashi et al. 2005),乳癌(Ghebeh et al. 2006),胃癌(Wu et al.
2006),口腔鱗狀細胞癌(Tsushima et al. 2006)等。許多腫瘤浸潤性T淋巴球之 Foxp3+調節性T細胞確實有上升的情形,並且與術後的復發情形,存活率,淋巴 轉移等臨床病理危險因子有關(Bate et al. 2006, Liyanage et al. 2002)。在腫瘤浸潤 性淋巴球中若有Foxp3、B7-H1 及其接受體PD-1分子一起表現於相同的腫瘤組織 中,可能會有協同作用使病人的免疫反應減弱(Ghebeh et al. 2008)。但也有報告指 出B7-H1及PD-1兩者皆會表現在CD4+CD25+調節性T細胞,但是他們真正的功能 至今尚未明瞭。
本論文由口腔鱗狀細胞癌病患分離出之腫瘤內浸潤CD4+CD25+ T細胞,研究 發現其B7-H1共同抑制分子的細胞表現比例,在每一病患個體比起PBMC的 CD4+CD25+ T細胞都高,平均值比例雖然有一些不同,但是可能由於每一個病患 體內T細胞活化程度有很大的差別,百分比的分布非常廣,因此沒有達到統計上 顯著的差異。此外我們將腫瘤浸潤CD4+ T細胞分成CD4+CD25high細胞、
CD4+CD25intermediate細胞、CD4+CD25low細胞和CD4+CD25-細胞,發現三細胞 亞群隨著CD25的表現量下降其B7-H1表現百分比也會下降,雖然沒有統計上的顯 著差異,但是由於也發現Foxp3的和B7-H1的螢光表現量可能有關聯性,因此 Foxp3+細胞和B7-H1+細胞在腫瘤組織免疫調控可能有相似或協同的作用。
過去研究在乳癌腫瘤組織切片中可以發現Foxp3+調節性T細胞和表現B7-H1/
PD-1的T細胞共同浸潤到腫瘤組織(Ghebeh et al,2008),而有B7-H1+的腫瘤組織 也具有較高量的Foxp3+調節性T細胞存在(Loos et al,2008),並且Foxp3+調節性T
細胞和B7-H1有正相關性,此外非霍奇金氏淋巴瘤(non-Hodgkin's lymphomas) 和移植免疫的研究指出,表現在調節性T細胞上的B7-H1,具有抑制T細胞的活化 增生的功能(Yang et al,2006) (Kitazawa et al,2007)。由於我們也在口腔癌腫瘤組織 看到Foxp3和B7-H1的相關性,因此我們想進一步研究,口腔鱗狀細胞癌腫瘤組織 浸潤Foxp3+調節性T細胞是否會表現B7-H1,B7-H1在很多腫瘤組織和抗原呈現細 胞都會表現,並且在活化的CD4+都會看到B7-H1,因此我們必須詳細分析不同亞 群細胞表現B7-H1的百分比和表現量,才能説明B7-H1的表現在其中的相對重要 性。
在本論文研究發現CD4+Foxp3+調節性T細胞會表現B7-H1,並且在腫瘤組織 比病人和正常PBMC的表現比例(p=0.005) (p=0.004)和表現量(p=0.0007)都高,因 此我們想進一步研究B7-H1+CD4+Foxp3+調節性T細胞在腫瘤組織的重要性,有 可能B7-H1+CD4+Foxp3+在腫瘤微環境扮演重要的免疫抑制角色。此外我們也發 現了B7-H1+Foxp3+和Foxp3+細胞在腫瘤組織有正相關性,B7-H1+Foxp3+和 Foxp3+細胞之百分比在TIL呈現中度正相關(r=0.61),而在CD4+T細胞內則是呈現 高度正相關(r=0.75),因此更加支持了本研究的假設。
接著我們進一步了解B7-H1在腫瘤組織淋巴球表現的分布,將PBMC和TIL 分為CD4+CD25+Foxp3+調節性T細胞、CD4+CD25+Foxp3-活化T細胞、
CD4+CD25-Foxp3-未活化T細胞,分析三細胞群B7-H1的表現百分比和表現量,研 究發現不論是在正常人或病人的PBMC,其中CD4+CD25+Foxp3+調節性T細胞 B7-H1的表現比例在週邊血液都是三群T細胞中最低的,然而在腫瘤組織的 CD4+CD25+Foxp3+調節性T細胞則是比病人PBMC(**p= 0.001)和正常人
PBMC(*p=0.04)表現較高比例的B7-H1,雖然三群T細胞在腫瘤組織都可觀察到平 均螢光表現強度都有上升,但是卻只有CD4+CD25+Foxp3+調節性T細胞在腫瘤局
部可以看到B7-H1的表現比例有明顯變化,有可能是口腔癌在腫瘤組織誘導出一 群表現B7-H1的調節性T細胞。CD4+CD25+Foxp3-活化T細胞、CD4+CD25-Foxp3- 未活化T細胞在周邊表現的B7-H1可能有其他我們未知的功能,因此我們認為只有 CD4+CD25+Foxp3+調節性T細胞所表現的B7-H1,對於腫瘤微環境的免疫抑制較 有相關性,此外三群T細胞在腫瘤微環境都可以看到B7-H1平均表現量的上升,可 能是由於大量活化細胞激素所影響,導致我們看到的差異性。
為了證實 CD4+CD25+B7-H1+Foxp3+T 細胞,對於口腔鱗狀細胞癌的免疫抑 制是有影響的,因此我們以同樣方式,將PBMC 和 TIL 分為 CD4+CD25+Foxp3+
調節性T 細胞、CD4+CD25+Foxp3-活化 T 細胞、CD4+CD25-Foxp3-未活化 T 細 胞,分析三細胞群的PD-1 表現百分比和表現量,研究發現 PD-1 在三群細胞的表 現 比 例 都 會 有 上 升 的 現 象 , 但 是 在 平 均 螢 光 表 現 量 則 是 可 以 發 現 , CD4+CD25+Foxp3-活化 T 細胞在 TIL 之 PD-1 平均表現量大約是 PBMC 的六倍 高,而CD4+CD25-Foxp3-未活化 T 細胞在 TIL 之 PD-1 平均表現量大約是 PBMC 的五倍高,然而CD4+CD25+Foxp3+調節性 T 在 TIL 之 PD-1 平均表現量大約只 有 PBMC 的 二 倍 高 , 因 此 本 論 文 發 現 可 能 是 口 腔 癌 腫 瘤 組 織 的 高 比 例 CD4+CD25+B7-H1+Foxp3+T 細 胞 , 結 合 CD4+CD25+Foxp3- 活 化 T 細 胞 、 CD4+CD25-Foxp3-未活化 T 細胞上高表現量的 PD-1,使得 T 細胞的功能受到抑 制,進而讓腫瘤環境產生免疫抑制,使腫瘤不斷生長。
此外我們有利用腫瘤組織萃取的 RNA 進行即時聚合酵素鏈反應後,也發現 PD-1 和 B7-H1 在腫瘤組織的高表現量,並且經過統計也看到 PD-1 和 B7-H1 具 有中度相關(r=0.54),在流式細胞儀經過分析後也發現 B7-H1+Foxp3+和 PD-1 在 TIL(r=0.74)和 CD4+T 細胞(r=0.70)都具有高度正相關,B7-H1+Foxp3+表現百分比 和Foxp3-細胞 PD-1 表現強度在腫瘤組織 CD4+T 細胞(r=0.6)具有中度相關性。
這樣的假設也利用口腔鱗狀細胞癌腫瘤組織進行組織免疫染色來證實,發現 有B7-H1+CD4+Foxp3+調節性 T 細胞的存在,也有表現 B7-H1 的接受體 PD-1 的 浸潤細胞,因此 CD4+CD25+B7-H1+Foxp3+T 細胞的確有可能藉由和 PD-1 的結 合影響T 細胞功能。藉由口腔鱗狀癌細胞細胞株 SAS 和 PBMC 的體外共同培養 系統,可以觀察到SAS 會誘導 CD4+CD25+表現較高比例的 B7-H1,因此我們初 步認為口腔癌細胞會影響CD4+CD25+調節性 T 細胞的 B7-H1 表現。
進一步我們發現腫瘤組織 CD4+CD25+Foxp3+ICOS+細胞表現比例稍高的 B7-H1,並且表現兩倍量的 B7-H1,因此可能 B7-H1 和 ICOS 的表現可能有相關 性。此外在腫瘤組織的RNA 可以看到較高量 IL-10、IFN-γ、CD4、PD-1、B7-DC、
B7-H4、Foxp3 和 B7-H1,因此可以了解其對於腫瘤組織的免疫調節重要性。
雖然在細胞增生實驗當中,無論在強烈(五天)或微弱(三天)的刺激下,都沒有 看到B7-H1 對 CD4+CD25+ T 細胞抑制功能的貢獻,但是有可能在腫瘤浸潤中表 現比例高且量多B7-H1 的 CD4+CD25+ T 細胞才會有明顯的功能,因此在週邊血 液的細胞比較不容易看到 B7-H1 的免疫抑制功能。此外過去有研究認為 B7-H1 對 於 細 胞 本 身 具 有 促 進 或 抑 制 細 胞 凋 亡 的 功 能 , 因 此 在 腫 瘤 組 織 的 CD4+CD25+B7-H1+Foxp3+T 細胞,也有可能是調節性 T 細胞本身的訊息傳遞調 控接受體。此外由於可以明顯觀察到腫瘤組織 PD-1 表現量的上升,因此可能還 有其他的例如腫瘤細胞或是抗原呈現細胞表現B7-H1,可以和 PD-1 作用。
第五章、參考文獻
1. Azuma, M., D. Ito, H. Yagita, K. Okumura, J. H. Phillips, L. L. Lanier, and C.
Somoza. 1993. B70 antigen is a second ligand for CTLA-4 and CD28. Nature 366:76-79.
2. Azuma, T., S. Yao, G. Zhu, A. S. Flies, S. J. Flies, and L. Chen. 2008. B7-H1 is a ubiquitous antiapoptotic receptor on cancer cells. Blood 111:3635-3643.
3. Baecher-Allan, C., J. A. Brown, G. J. Freeman, and D. A. Hafler. 2001.
CD4+CD25high regulatory cells in human peripheral blood. J Immunol 167:1245-1253.
4. Bennett, C. L., J. Christie, F. Ramsdell, M. E. Brunkow, P. J. Ferguson, L.
Whitesell, T. E. Kelly, F. T. Saulsbury, P. F. Chance, and H. D. Ochs. 2001. The immune dysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) is caused by mutations of FOXP3. Nat Genet 27:20-21.
5. Bhatia, S., A. D. Louie, R. Bhatia, M. R. O'Donnell, H. Fung, A. Kashyap, A.
Krishnan, A. Molina, A. Nademanee, J. C. Niland, P. A. Parker, D. S. Snyder, R.
Spielberger, A. Stein, and S. J. Forman. 2001. Solid cancers after bone marrow transplantation. J Clin Oncol 19:464-471.
6. Bluestone, J. A., and A. K. Abbas. 2003. Natural versus adaptive regulatory T cells. Nat Rev Immunol 3:253-257.
7. Butte, M. J., M. E. Keir, T. B. Phamduy, A. H. Sharpe, and G. J. Freeman. 2007.
Programmed death-1 ligand 1 interacts specifically with the B7-1 costimulatory molecule to inhibit T cell responses. Immunity 27:111-122.
8. Chapoval, A. I., J. Ni, J. S. Lau, R. A. Wilcox, D. B. Flies, D. Liu, H. Dong, G.
L. Sica, G. Zhu, K. Tamada, and L. Chen. 2001. B7-H3: a costimulatory
2:269-274.
9. Curiel, T. J. 2007. Tregs and rethinking cancer immunotherapy. J Clin Invest 117:1167-1174.
10. Curiel, T. J., G. Coukos, L. Zou, X. Alvarez, P. Cheng, P. Mottram, M.
Evdemon-Hogan, J. R. Conejo-Garcia, L. Zhang, M. Burow, Y. Zhu, S. Wei, I.
Kryczek, B. Daniel, A. Gordon, L. Myers, A. Lackner, M. L. Disis, K. L.
Knutson, L. Chen, and W. Zou. 2004. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival.
Nat Med 10:942-949.
11. Curiel, T. J., S. Wei, H. Dong, X. Alvarez, P. Cheng, P. Mottram, R. Krzysiek, K. L. Knutson, B. Daniel, M. C. Zimmermann, O. David, M. Burow, A.
Gordon, N. Dhurandhar, L. Myers, R. Berggren, A. Hemminki, R. D. Alvarez, D. Emilie, D. T. Curiel, L. Chen, and W. Zou. 2003. Blockade of B7-H1 improves myeloid dendritic cell-mediated antitumor immunity. Nat Med 9:562-567.
12. Dave, B. J., A. H. Trivedi, and S. G. Adhvaryu. 1992. Role of areca nut consumption in the cause of oral cancers. A cytogenetic assessment. Cancer 70:1017-1023.
13. Dong, H., S. E. Strome, E. L. Matteson, K. G. Moder, D. B. Flies, G. Zhu, H.
Tamura, C. L. Driscoll, and L. Chen. 2003. Costimulating aberrant T cell responses by B7-H1 autoantibodies in rheumatoid arthritis. J Clin Invest 111:363-370.
14. Dong, H., S. E. Strome, D. R. Salomao, H. Tamura, F. Hirano, D. B. Flies, P. C.
Roche, J. Lu, G. Zhu, K. Tamada, V. A. Lennon, E. Celis, and L. Chen. 2002.
Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of
immune evasion. Nat Med 8:793-800.
15. Dong, H., G. Zhu, K. Tamada, and L. Chen. 1999. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion.
Nat Med 5:1365-1369.
16. Freeman, G. J., J. G. Gribben, V. A. Boussiotis, J. W. Ng, V. A. Restivo, Jr., L. A.
Lombard, G. S. Gray, and L. M. Nadler. 1993. Cloning of B7-2: a CTLA-4 counter-receptor that costimulates human T cell proliferation. Science 262:909-911.
17. Freeman, G. J., A. J. Long, Y. Iwai, K. Bourque, T. Chernova, H. Nishimura, L.
J. Fitz, N. Malenkovich, T. Okazaki, M. C. Byrne, H. F. Horton, L. Fouser, L.
Carter, V. Ling, M. R. Bowman, B. M. Carreno, M. Collins, C. R. Wood, and T.
Honjo. 2000. Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation. J Exp Med 192:1027-1034.
18. Gavin, M. A., J. P. Rasmussen, J. D. Fontenot, V. Vasta, V. C. Manganiello, J. A.
Beavo, and A. Y. Rudensky. 2007. Foxp3-dependent programme of regulatory T-cell differentiation. Nature 445:771-775.
19. Geng, L., D. Huang, J. Liu, Y. Qian, J. Deng, D. Li, Z. Hu, J. Zhang, G. Jiang, and S. Zheng. 2008. B7-H1 up-regulated expression in human pancreatic carcinoma tissue associates with tumor progression. J Cancer Res Clin Oncol 134:1021-1027.
20. Ghebeh, H., E. Barhoush, A. Tulbah, N. Elkum, T. Al-Tweigeri, and S.
Dermime. 2008. FOXP3+ Tregs and B7-H1+/PD-1+ T lymphocytes
co-infiltrate the tumor tissues of high-risk breast cancer patients: Implication for immunotherapy. BMC Cancer 8:57.