Marine Biology 107, 75-81 (1990)
M a r i n e
... B i o l o g y
© Springer-Verlag 1990Relationship between growth rate and age at recruitment
of
Anguilla japonica
civets in a Taiwan estuary
as inferred from otolith growth increments
W.-N. Tzeng
Department of Zoology, National Taiwan University, Taipei, Taiwan 10764, Republic of China Date of final manuscript acceptance: August 3, 1990. Communicated by M. Anraku, Tokyo
Abstract. The growth history and recruitment dynamics
of eel
(Anguilla japonica)
elvers were studied. Observa- tions were based on growth increments in sagittal otoliths of elvers collected from Shuang-Chi River estuary off northeastern Taiwan, from November 1985 to February 1986. Total lengths of elvers upon arrival at the estuary were similar in most case; mean total lengths were from 55.99 to 59.06 ram. Daily ages of elvers at arrival ranged from 112.8___9.4 ( ± S D ) to 156.5+13.5 d, indicating that migration of eel larvae from their oceanic spawning ground to the estuary requires 4 to 5 too. Elver hatching dates, back-calculated from estimated daily ages, indicat- ed that the spawning season lasted 5 mo (from late June to early October). Furthermore, the earlier eels spawned, the earlier elvers reached the estuary. The transition in growth history during the larval stage was obvious, as indicated from the change in increment width in elver otoliths. The inverse correlation between daily age and mean daily growth rates of fish length and otolith indicat- ed that the age of elvers upon arrival at the estuary was susceptible to larval growth rate. In other words, the time taken on migration from oceanic spawning ground to the estuary was shorter for fast-growing larvae than for slow- growing ones.Introduction
Japanese eel,
Anguillajaponica,
is one of the most impor- tant cultured species in Taiwan and Japan. For cultiva- tion, large numbers of elvers are caught from the estuary during their upstream migration from November to March (Kuo 1971, Tzeng 1983, 1984, 1985). The spawn- ing ground of the eel is thought to be in the waters south of Okinawa and east of the Philippines (Matsui 1957). Eel larvae drift from oceanic spawning ground along with ocean current. Having attained their full term of growth, the larvae then undergo a metamorphosis from leptocephali into young transparent glass eels, or elvers. After this, they are endowed with increased activity, and so cross the coastal zones and make their ascent into freshwater. The eels live in rivers for 5 to 20 yr. During late Autumn when eels become premature, they migrate downstream to the ocean to spawn (Tesch 1977). In order to elucidate the early life history of the Japanese eel - including spawning ground, distribution, migration and growth history of leptocephali and elvers - a number of field investigations have been conducted since 1967. However, only 86 specimens of leptocephali have been obtained until now, and thus biological data has re- mained insufficient to clarify the early life history (Mat- sui et al. 1968, Tanaka 1975, Kajihara et al. 1987, Tabeta and Mochioka 1987, Kajihara 1988, Ozawa et al. 1989, Tsukamoto et al. 1989).
Since the daily formation of growth increments in fish otoliths was reported by Pannella (1971), aging of larval and juvenile fish by counting daily growth increments has become a commonly used technique for studying the early life history of fish. Examination of otolith micro- structure has proved to be a useful tool for estimating spawning and hatching dates (Ralston 1976, Struhsaker and Uchiyama 1976, Miller and Storck 1984), growth rate (Campana 1984, Volk et al. 1984, Penney and Evans 1985) and the transitions in growth history (Radtke and Dean 1982, Neilson et al. 1985, Victor
1986,
Chambers and Leggett 1987). The growth increments in otoliths of leptocephali and elvers were clearly revealed by Tabeta et al. (1987) and Tsukamoto et al. (1989), with the growth increments in otoliths of newly hatched larvae and elvers supposedly deposited on a daily basis (Lee and Lee1989,
Tsukamoto 1989, Umezawa et al. 1989). Thus, it is possi- ble to determine the age and to calculate the growth rate of elvers by counting otolith growth increments. In this paper, I intend to elucidate the early growth history of elvers through examination of daily age, hatching date and growth rate. A discussion on the recruitment dynam- ics of elvers in the estuary of Taiwan is also presented.
Materials and methods
A total of 208
Anguillajaponiea
elvers were collected using a hand- trawling net at night along the surf zone of the coastal waters of76 W.-N. Tzeng: Otolith growth increments in eel elvers Table 1.
Pigmentati•nstages•fAngui••ajap•nicae•versc•••e•tedfr•mShuang-ChiRiverestuary.Pigmentati•nstagesafterStrubberg(19•3)
Group Collection date Sample size
No. fish in pigmentation stage
VA VB VIA1 VIA2 VIA3 VIA4 VIB
1 4November 1985 15 3 10 2 2 14December 1985 18 13 5 3 11, 16 January 1986 30 6 6 5 1 4 7 1 4 5 to 26 February 1986 145 112 24 7 2 Total 208 134 45 14 3 4 7 1
3°1
2o Gr0upl 1 0 0 i I I t t i i i r N Gr0up2 (]~ i i I I I I J I T M . 2o Gr0up3 1 I i i [ - " ~ l ~ i i i r~° t
~ [~
Gr0up4 - - , i i i t i T I I i i i I I I I I i i 5 0 5 2 5 4 5 6 5 8 6 0 6 2 6 4 6 6 6 BTolal Lenglh (ram)
Fig. 1.
Anguillajaponica.Frequency distribution of total length of
elvers collected from Shuang-Chi River estuary, Taiwan on four different dates. Group 1: November 4, 1985, n (sample size)= 15; Group 2: December 14, 1985, n = 18; Group 3: January 11 and 16, 1986, n=30; and Group 4: February 5 to 16, 1986, n=145
SEM (scanning electron microscope, Hitachi S-520) at 20 kV. The growth increment is a bipartite structure which consists of a cal- cium-rich incremental zone and an organic-rich discontinuous zone. After EDTA etching, these two zones became the "crest and trough" on the polished otolith surface, which could then be scanned by SEM.
Otolith radius, along the longest axis from the core to the peripheral margin, was measured and growth increment was count- ed from a SEM photograph at x 2000. Growth increments in each otolith were counted at least three times and the mean was adopted if the difference in the number of these three counts was less than
10. Otherwise, the otolith was disregarded.
In most marine fish, the initial otolith growth increment is deposited during their first feeding when the larvae complete yolk- sac absorption (Lough et al. 1982, McGurk 1984, Tzeng and Yu 1988). The yolk-sac of eel larvae was completely absorbed 4 to 6 d post-hatching (Umezawa et al. 1989). Therefore, the initial growth increment in otoliths of elvers was proposed to be deposited during the transitional stage when larvae switch from endogenous yolk-sac nutrition to exogenous feeding, and thus the daily age of elvers after hatching was estimated by the number of otolith growth increments with an additional 5-d yolk-sac period.
Mean daily growth rates of body and otolith of each elver were, respectively, calculated by the ratios of total length and maximum radius of otolith to estimated daily age. Furthermore, the relation- ships between body and otolith growth rates and daily age were fitted with regression lines using logarithmically transformed and arithmetic data, respectively (Snedecor and Cochran 1969). Tests for homogeneity and significant difference of total length and otolith radius with daily age were conducted using a one-way analysis of variance (ANOVA) and Scheff's multiple comparison, respectively (Sokal and Rohlf 1981).
Shuang-Chi River estuary, northeastern Taiwan during the fishing season (from November 1985 to February 1986) (Table 1). Elvers were immediately fixed with 95% alcohol, and total length was measured to the nearest 0.1 mm 1 wk post-fixation. The elver pig- mentation stage was determined according to the distribution of pigments on the body surface (Strubberg 1913, Bertin 1956). A total of 61 elvers at the early pigmentation stage, randomly selected from those collected on November 4 and December 14, 1985, January 11, 16 and February 16, 1986 (Groups 1 to 4), respectively, were used to examine otolith microstructure.
Sagittal otoliths of elvers were dissected from the otic vesicle under a binocular dissecting microscope using a pair of minute insect needles mounted on wooden rods. The dissected otoliths were then separated from extraneous organic materials, washed with distilled water, dried in air, and finally embedded in spur and hard- ened in an oven for 24 h at 70 °C. Embedded otoliths were ground and polished with 1000 to 2000 mesh polishing paper and alumina paste along the anterior-posterior direction in the frontal plane of the fish until otolith core was revealed. Polished otoliths were etched with 5% EDTA (ethylene diamine tetra-acetate; pH adjusted with NaOH to 7.4) for ca. 3 min, coated with gold and viewed under
Results
Total l e n g t h a n d o t o l i t h r a d i u s
T h e m o n t h l y l e n g t h - f r e q u e n c y d i s t r i b u t i o n o f
Anguilla
japonica
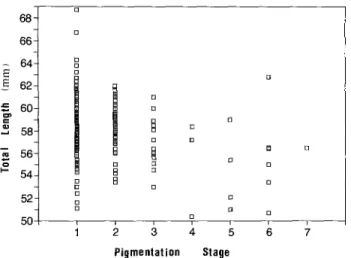
elvers are s h o w n i n Fig. 1. T o t a l l e n g t h s r a n g e d f r o m 49.6 to 68.7 m m , a n d m e a n t o t a l l e n g t h s for each s a m p l i n g p e r i o d ( G r o u p s 1 to 4 ) w e r e 56.59 + 2 . 2 9 m m , 56.88 + 2.07 ram, 55.99 + 3.00 m m a n d 59.06 + 2.77 m m , respectively. T h e m e a n t o t a l l e n g t h o f elvers a m o n g each s a m p l i n g p e r i o d was n o t s i g n i f i c a n t l y d i f f e r e n t (p > 0.05); however, the m e a n t o t a l l e n g t h o f elvers i n F e b r u a r y was significantly greater t h a n those in other m o n t h s (p < 0.01). T h e p i g m e n t a t i o n stages o f elvers collected i n the e s t u a r i n e r e g i o n were d o m i n a t e d b y Stages VA a n d VB, w i t h the o t h e r stages b e i n g relatively few (Table 1). Stage VA a n d VB elvers were believed to be the n e w recruits i n the estuary. Elver t o t a l l e n g t h seemed to r e d u c e slightly as the stages a d v a n c e d (Fig. 2) since the c o r r e l a t i o n coef-W.-N. Tzeng: Otolith growth increments in eel elvers 68~ 66 ] 64- 62~
= 6o~
s8 Z 56 ] ~" 54~ 52 ~ 50 ] i B t~i
[]
B 13 [] B [] [] 12 B [] J Pigmentation StageFig. 2. Anguilla japonica. Relationship between total length and pigmentation stages of elvers. Specimens as in Fig. 1. Scales 1 to 7 on X-axis represent pigmentation Stages VA, VB, VIA1, VIA2, VIA3, VIA4 and VIB, respectively (Strubberg 1913)
60
40
1213
0
2~r 40 1
2 00
4° 1
20"
0
I I
Group1
G roup
2I
I I ~ ~-" ]
I ' I
Group 3
I I rGroup4
100 120 140 160 180 200
Max Otolith Radius (prn)
Fig. 3. Anguilla japonica. Frequency distribution of maximum otolith radius of elvers collected from Shuang-Chi River estuary, Taiwan, on four different dates. Groups as in Fig. 1
77 of 138.3_9.0#m, 136.3±9.2#m, 143.5_16.1 #m and 142.9_+14.6/~m (Groups 1 to 4) for elvers collected on November 4 and December 14, 1985 and January 11, 16 and February 16, 1986, respectively (Fig. 3). These mean values showed no significant difference among groups (ANOVA, F-test, p>0.05). On the other hand, otolith growth was not obviously in step with the advancement of the pigmentation stage since the correlation coefficient between otolith radius and pigmentation stage was not significant (r = 0.149, p > 0.05). This indicated that the development of pigmentation of elvers in this estuary progresses over a short period during upstream migra- tion.
Otolith microstructure and growth increment
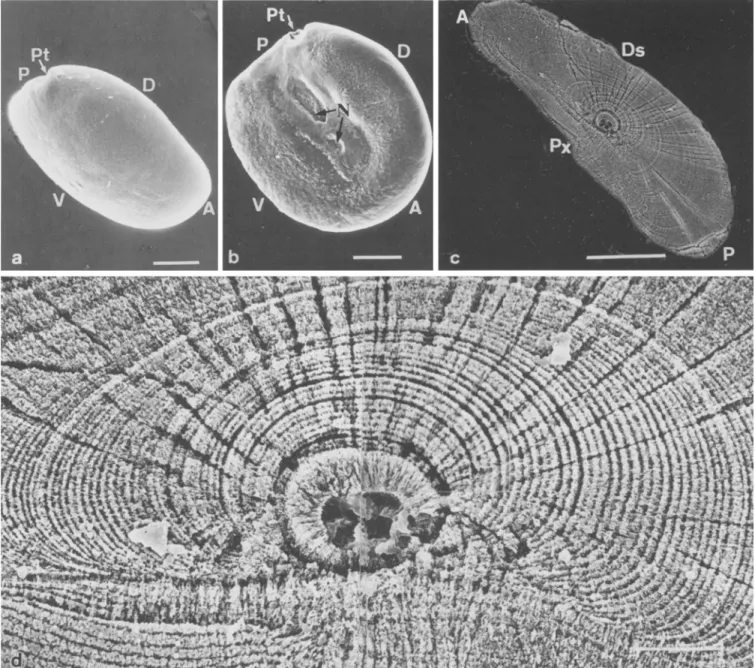
The sagittal otolith of elvers, as shown in Fig. 4 a, b, was subelliptical in external feature. The longest axis of the otolith was along the anterior-posterior direction, while the shortest axis was along the proximal-distal direction. The proximal side of the otolith resembled a horseshoe- shape with a postcaudal trough posteriorly and a U- shape cristae peripherally, two neuron insertion holes in the colliculum area and a sulcus acusticus located be- tween cristae and colliculum (Fig. 4 b).
The central part of a ground and etched otolith, as shown in Fig. 4 c, d, revealed a deeply etched core and amorphous nucleus, both ca. 17 #m in diameter. The peripheral part of the nucleus was alternately surrounded by a calcium-rich incremental zone (light band) and an organic-rich discontinuous zone (dark band). A single growth increment comprising an incremental zone plus a discontinuous zone was generally formed in a daily schedule. The number of growth increments in the otolith of this specimen was ca. 122 rings. The daily age of this elver was thus estimated at ca. 127 d post-hatching, in- cluding 5-d yolk-sac period adjustment.
The otolith growth rate was different in each direc- tion, being faster in the anterior-posterior direction due to the larger increment width there. Increment width was narrow in the distal side and the growth increment was interrupted in the proximal side (Fig. 4 c, d). The anteri- or-posterior direction of the otolith thus seemed the most appropriate for examining daily growth increment.
The change in otolith increment width (Fig. 4c, d) revealed the growth history for elvers for the period when the fish migrated from the oceanic spawning ground across Kuroshio to the estuary. Elvers grew fast during the early larval stage and slowed at 65 to 100 d old, which was followed by another fast-growing period.
ficient between total length and pigmentation stage was significant ( r = - 0 . 5 3 5 , p<0.01). Thus, elvers of the advanced pigmentation stage were not used in otolith growth increment studies - to minimize the effect of pigmentation stage on the evaluation of body growth rate.
The maximum otolith radius of elvers ranged from 120 to 190/~m (mode 130 to 140/~m) with a mean
Daily age and hatching date
The mean daily ages of elvers upon arrival at the estuary were estimated as 112.8 _ 9.4 d, 120.0_+ 10.4 d, 129.9_+ 9.7d and 156.5_+13.5d in Groups 1 to 4, respectively (Table 2). Except for Group 4, the mean daily age of elvers from different sampling dates was similar. Thus, the time taken for eel larvae to migrate from the spawn-
78 W.-N. Tzeng: Otolith growth increments in eel elvers
Fig. 4. Anguilla japonica. Scanning electron micrographs illustrat-
ing external feature and internal microstructure of sagittal otolith of an elver collected in the Shuang-Chi River estuary on December 14, 1985. (a) Dorsal-distal face of right otolith from a 58.7 mm total length (TL) elver. (b) Proximal face of left otolith from a 63.8 mm TL elver. (c) Growth increments in the otolith sectioned along anterior-posterior direction and frontal plane of a 56.0 mm TL
elver. (d) Enlargement of (c). Light (thick) and dark (thin) bands indicate the incremental and the discontinuous zones of the growth increments, respectively. Abbreviations are (A) anterior; (D) dor- sal; (Ds) distal side; (N) neuron insertion areas; (P) posterior; (Pt) postcaudal trough; (Px) proximal side; (V) ventral. Scale bar= 50/zm (a, b, c), 10/zm (d)
ing site to the estuary was ca. 4 mo. It was, however, ca. 5 m o for G r o u p 4.
The hatching dates o f elvers (Fig. 5), back-calculated f r o m their daily age, indicated that the spawning season o f the eel lasted ca. 5 m o , f r o m late June to early October. Fig. 5 also reveals that the earlier eels spawned the earlier elvers reached the estuary. However, the hatching dates o f elvers o f G r o u p s 3 and 4 overlapped, which m a y indi- cate that they originated f r o m the same spawning group, with G r o u p 4 elvers obviously delayed in their recruit- m e n t to the estuary.
Relationship between growth rates and daily age at recruitment
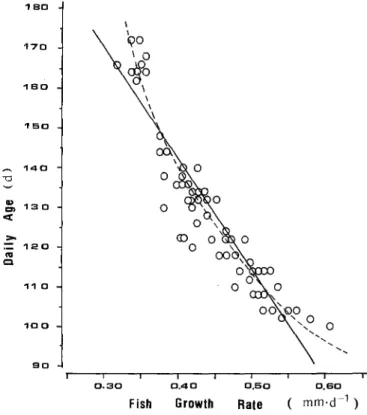
A highly negative correlation was observed between daily age at estuary arrival and m e a n daily growth rates of b o t h total length and otolith radius (Figs. 6 and 7). The relationship between daily age ( T D, d) and m e a n daily fish growth rate (GL, m m d - 1) was calculated by either a linear model:
W.-N. Tzeng: Otolith growth increments in eel elvers
Table 2. Frequency distribution of daily age of Anguilla japonica
elvers sampled from Shuang-Chi River estuary northeastern Taiwan on four different dates. Group 1: November 4, 1985; Group 2: December 14, 1985; Group 3: January 16, 1986; and G r o u p 4 : February 16, 1986. Test of significant difference of mean daily age between groups was made using Scheffs method. The lines connected between groups indicated non-significant difference (p > 0.05), those without lines were significantly different (p < 0.01) Age (d) No. fish
Group 1 G r o u p 2 G r o u p 3 G r o u p 4 9 0 - 95 95-100 100-105 105-110 110-115 115-120 120-125 125-130 130-135 135-140 140-145 145-150 150-155 155-160 160-165 165-170 170-175 175-180 Total
Mean daily age (-+SD) Significance test i 4 2 2 0 1 2 6 0 2 4 0 1 2 4 1 0 1 1 4 4 3 0 1 14 18 14 15 112.8 120.0 129.9 156.5 _+9.4 __10.4 -+9.7 _+13.5
79
1 8 01.o
\
1so
1 5 0~ 0
0
.~-- 1 2 0 1 1 0 1 O O O " x 9 0 i I I ! i l i i 0 , 3 0 0 , 4 0 0 . 5 0 0 . 6 0Fish Growth Rate
( m m . d 1 )Fig. 6. Anguilla japonica. Regressions of daily age (TD) on mean daily growth rate of total length (GL) for elvers collected from Shuang-Chi River estuary, Taiwan. Continuous line: T D = 250.62- 270.50 GL; dashed line: Ln TD=4.09849--0.92582 Ln G L 1 4 0 -CD 1 5 0 = . m 14. o Z 4 0 E3 1 n
Group4
I
Group3
,
,
I ] I
,
0 IJUL
Group2
Group1
I I IAUG SEP OCT 1 9 8 5
Hatching Date
Fig. 5. Anguilla japonica. Frequency distribution of hatching dates, classified by 10 d, of elvers collected from Shuang-Chi River estu- ary, Taiwan on four different dates. Groups as in Fig. 1
1 8 0 1 7 0 1 6 0 1 5 0 1 4 0 1 3 0 . ~ 1 2 0 g~ 1 1 0 1 0 0 9 0
\
oo
0
\ \ ~8
oo °
o Oo \
o
o o o \ \ o
0 00 0 ~
i n I I 0 . 5 0 0 - 8 0 1 . Q O 1 . 2 0 1 - 4 0Otolith Growth Rate
( lurn'd -1 )Fig. 7. Anguilla japonica, Regressions of daily age (To) on mean daily otolith growth rate (G R) for elvers collected from Shuang-Chi River estuary, Taiwan. Continuous line: T o = 241.67-- 101.96 G R ; dashed line: Ln T D =4.92320--0.82796 Ln GR
80
or a curvilinear model:
Ln T~=4.09849-0.92582 Ln G L ( r = - 0 . 9 5 6 , n=61).
(2)
Similarly, the relationship between daily age (T D, d) and mean daily otolith growth rate (GR, pm d-1) was calculated by either a linear model:
T~=241.67-101.96 GR (r=--0.816, n = 6 1 ) , (3) or a curvilinear model:
Ln T D = 4.92320--0.82796 Ln G R (4) (r= -0.828, n=61).
The inverse relationships between daily age and mean daily growth rates of fish length and otolith radius indicated that fast-growing elvers arrived at the estuary earlier than slow-growing ones. The time required for migration from spawning ground to the estuary could be reduced from ca. 160 to 95 d by an accompanied increase in growth rate from 0.35 to 0.60 mm d - t, or alternatively in otolith growth rate from 0.80 to 1.4 #m d-1.
Discussion
In order to accurately estimate the daily age of Anguilla
japonica elvers, the time of the formation of the initial otolith growth increment must be known, a time which is still controversial among investigators (Tabeta et al. 1987, Tsukamoto et al. 1989, Umezawa et al. 1989). The most desirable method of validating ring-deposition rate is to rear larvae through the entire larval stage, from eggs to metamorphosis, sampling reasonable numbers of individ- uals at frequent intervals throughout (Geffen 1987). Until recently, however, rearing techniques had not yet been established; the artificially hatched eel larvae used for validating initial growth increment in the experiment of Umezawa et al. (1989) all died before the first feeding stage when the larvae had completed yolk-sac absorp- tion. Like most marine fish, the initial growth increment in otoliths of eel larvae was probably deposited at first feeding (upon complete absorption of the yolk-sac) (Lough et al. 1982, Radtke and Dean 1982, Campana 1984, McGurk 1984, Neilson et al. 1985, Tzeng and Yu 1988). The yolk-sac of eel larvae was completely absorbed 4 to 6 d post-hatching (Umezawa et al. 1989). Thus, an adjustment for the 5-d yolk-sac period was considered when the number of growth increments in otoliths was applied to estimated daily age after hatching.
The particular elver shown in Fig. 4c, d revealed moderate growth between 1 and 65 d old, grew slowly between 65 and 100 d old, and sped up again after 100 d. This growth pattern was very similar to that reported by Tabeta et al. (1987), who investigated elvers of a different year and location. The slow-growing incre- ments (Fig. 4c, d) were deposited during the period mid-September to mid-October, the timing of which seemed to be independent of the low temperature in winter months. Eel larvae are subjected to an extremely different environment during their early life history - they
W.-N. Tzeng: Otolith growth increments in eel elvers drift with the ocean current and migrate across the Kuroshio current to the coastal waters, then enter the estuary (Tesch 1977). Larvae metamorphose from leptocephali to elvers before entering coastal waters. The changes in otolith increment width might reflect the tran- sition of developmental stage and change of their habitats (Neilson et al. 1985, Victor 1986, Chambers and Leggett 1987).
The daily age of elvers at the time of arrival at the estuary was estimated to be 112.8 to 156.5 d post-hatch- ing (Table 2). Thus, the duration of Japanese eel lepto- cephalus and elver stages in the ocean was estimated to be ca. 4 to 5 too. This is rather short in comparison with larvae of European eel, which take ca. 2.5 yr to migrate from spawning site to coastal waters (Bertin 1956, Tesch 1977). Larval life stage duration is associated with the distance between spawning site and estuary. The spawn- ing ground of Japanese eel is not far from the estuary of Taiwan, and it is thought to be in the waters south of Okinawa and east of Taiwan and Luzon (Matsui 1957, Kajihara 1988, Ozawa et al. 1989).
The growth rate variation in the early life of fishes has a pronounced effect on recruitment (Larkin 1978, Houde 1987). The inverse relationship between daily age of elvers upon arrival at the estuary and the mean daily growth rates of fish and otolith (Figs. 6 and 7) indicated that the age of elvers upon arrival at the estuary was susceptible to larval growth rate. Fast-growing eel larvae arrived at the estuarine area quicker than slow-growing ones. In gener- al, fast-growing larvae metamorphose early and swim quickly to the preferred habitat (Hunter 1972, Miller et al. 1988). In contrast, slow-growing larvae prolong larval stage duration and delay metamorphosis; they are unable to swim as fast as the fast-growing ones, but delay recruit- ment to the estuary (Victor 1986). This phenomenon has been observed in Group 4 elvers, which delayed recruit- ment to the estuary and were of older daily age (Table 2, Fig. 5). Furthermore, such larvae are more vulnerable to predation, which might cause poor recruitment. Thus, larval growth rate influences not only the recruitment age but the year-class strength of elvers recruited into the estuary. The latter assumption will be clarified by future investigation.
Acknowledgements. This study was conducted with the financial support of the National Science Council, Republic of China (Project No. NSC 77-0211-B002-18 and 79-0211-B002-09). The au- thor is grateful to Professor O. Tabeta for providing the otolith handling technique; Miss S. Y. Yu for handling specimen; Mr W. C. Chen for preparing the manuscript; and two anonymous reviewers for helpful comments.
Literature cited
Bertin, L. (1956). Eels - a biological study. Cleaver-Hume Press, London
Campana, S. E. (1984). Microstructural growth patterns in the otoliths of larval and juvenile starry flounder, Platichthys stella-
tus. Can. J. Zool. 62:1507-1512
Chambers, R. C., Leggett, W. C. (1987). Size and age at meta- morphosis in marine fishes: an analysis of laboratory-reared winter flounder (Pseudopleuroneetes americanus) with a review
W.-N. Tzeng: Otolith growth increments in eel elvers
of variation in other species. Can. J. Fish. aquat. Science 44: 1936-1947
Geffen, A. J. (1987). Methods of validating daily increment deposi- tion in otoliths of larval fish. In: Summerfelt, R. C., Hall, G. E. (eds.) Age and growth of fish. Iowa State University Press, Ames, Iowa, p. 223-240
Houde, E. D. (1987). Fish early life dynamics and recruitment variability. Am. Fish. Soc. Syrup. 2 : 1 7 - 2 9
Hunter, J. R. (1972). Swimming and feeding behavior of larval
anchovy, Engraulis mordax. Fish. Bull. U.S. 70:821-838
Kajihara, T. (1988). Distribution of Anguilla japonica leptocephali
in western Pacific during September 1986. Bull. Jap. Soc. Scient. Fish. 54:929-933
Kajihara, T., Tsukamoto, K., Otake, T., M0chioka, N., Hasumoto, H., Oya, M., Tabeta, O. (1987). Sampling leptocephali with reference to the diel vertical migration and the gears. Bull. Jap. Soc. Scient. Fish. 54:941 946
Kuo, H. (1971). Approach of eel elvers to the land in Taiwan. Aquaculture (Yoshoku) 1 : 5 2 - 5 6 (in Japanese)
Larkin, P. A. (1978). Fisheries management - an essay for ecolo- gists. A. Rev. Ecol. Syst. 9 : 5 7 - 7 3
Lee, T. W., Lee, K. S. (1989). Daily growth increments and lunar
pattern in otolith of the eel, Anguilla japonica, in the fresh-
water. Bull. Korean Fish. Soc. 2 2 : 3 6 - 4 0
Lough, R. G., Pennington, M., Bolz, G. R., Rosenberg, A. A.
(1982). Age and growth of larval Atlantic herring, Clupea
harengus L., in the Gulf of Maine-Georges Bank region based on otolith growth increments. Fish. Bull. U.S. 80:187-199 Matsui, I. (1957). On the records of a leptocephalus and catadro-
mous eels of Anguillajaponica in the waters around Japan with
a presumption of their spawning places. J. Shimonoseki Coll. Fish. 7:151-167
Matsui, I., Takai, T., Kataoka, S. (1968). Anguillid leptocephalus found in the Japan current and its adjacent waters. J. Shimono- seki Coll. Fish. 17:17-23
McGurk, M. IZ (1984). Ring deposition in the otoliths of larval
Pacific herring Clupea harengus pallasis. Fish. Bull. U.S. 82:
113 120
Miller, S. J., Storck, T. (1984). Temporal spawning distribution of large mouth bass and young-of-year growth, determined from daily otolith rings. Trans. Am. Fish. Soc. 113:571-578 Miller, T. J., Crowder, L. B., Rice, J. A., Marshall, E. A. (1988).
Larval size and recruitment mechanisms in fishes: toward a conceptual framework. Can. J. Fish. aquat. Science 45:1657 1670
Neilson, J. D., Green, G. H., Bottom, D. (1985). Estuarine growth
of juvenile chinook salmon Oncorhynchus tshawytscha as in-
ferred from otolith mieros~ructure. Can. J. Fish. aquat. Science 42:899-908
Ozawa, T., Tabeta, O., Mochioka, N. (1989). Anguillid leptocephali from the western north Pacific east of Luzon in 1988. Bull. Jap. Soc. Scient. Fish. 55:627-632
Pannella, G. (1971). Fish otoliths: daily growth layers and periodi-
cal patterns. Science, N.Y. 173 : 1 1 2 4 - 1 1 2 7
Penney, R. W., Evans, G. T. (1985). Growth histories of larval
redfish (Sebastes spp.) on an offshore Atlantic fishing bank
determined by otolith increment analysis. Can. J. Fish. aquat. Science 42:1452-1464
Radtke, R. L., Dean, J. M. (1982). Increment formation in the otoliths of embryos, larvae and juveniles of the mummichog,
Fundulus heteroclitus. Fish. Bull. U.S. 80:201-215
81 Ralston, S. (1976). Age determination of a tropical reef butterflyfish
utilizing daily growth rings of otoliths. Fish. Bull. U.S. 74: 990-994
Snedecor, G. W., Cochran, W. G. (1969). Statistical methods. Iowa State University Press, Ames, Iowa
Sokal, R. R., Rohlf, F. J. (1981). Biometry (2nd ed.). W. H. Freeman and Co., San Francisco
Strubberg, A. C. (1913). The metamorphosis of elvers as influenced by outward conditions. Medd. Komm. Havunders. Ser. Fisk. 4: 1-11
Struhsaker, P., Uchiyama, J. H. (1976). Age and growth of the nehu,
Stolephorus purpureus (Pisces: Engraulidae) from the Hawaiian Islands as indicated by daily growth increments of sagittae. Fish. Bull. U.S. 7 4 : 9 - 1 7
Tabeta, O., Mochioka, N. (1987). Anguillid leptocephali from the waters south of Okinawa and east of Taiwan and Luzon, western north Pacific in 1986. Bull. Jap. Soc. Scient. Fish. 54: 935-940
Tabeta, O., Tanaka, K., Yamada, J., Tzeng, W. N. (1987). Aspects of
the early life history of the Japanese eel Anguillajaponica deter-
mined from otolith microstructure. Bull. Jap. Soc. Scient. Fish. 53:1727-1734
Tanaka, S. (1975). Collection of leptocephali of the Japanese eel in waters south of the Okinawa Islands. Bull. Jap. Soc. Scient. Fish. 41:129-136
Tesch, F. W. (1977). The eel - biology and management of anguillid eels. Chapman and Hall Press, London
Tsukamoto, K. (1989). Otolith daily increments in the Japanese eel. Bull. Jap. Soc. Scient. Fish. 55:1017-1021
Tsukamoto, K., Umezawa, A., Tabeta, O., Mochioka, N., Kaji-
hara, % (1989). Age and birth date of Anguillajaponica lepto-
cephali collected in western north Pacific in September 1986. Bull. Jap. Soc. Scient. Fish. 55:1023-1028
Tzeng, W. N. (1983). Seasonal and long-term changes of the catch of anguillid elvers in Taiwan. J. Fish. Soc. Taiwan 1 0 : 1 - 7 (in Chinese with English summary)
Tzeng, W. N. (1984). Dispersion and upstream migration of
marked anguillid eel, Anguilla japonica elvers in the estuary of
the Shuang River, Taiwan. Bull. Jap. Soc. Fish. Oceanogr. 45: 10-20
Tzeng, W. N. (1985). Immigration timing and activity rhythms of the
eel, Anguilla japonica, elvers in the estuary of northern Taiwan
with emphasis on environmental influences. Bull. Jap. Soc. Fish. Oceanogr. 47/48 : 11-28
Tzeng, W. N., Yu, S. Y. (1988). Daily growth increments in otoliths
of milkfish, Chanos ehanos (Forsskfil), larvae. J. Fish Biol. 32:
495-504
Umezawa, A., Tsukamoto, K., Tabeta, O., Yamakawa, H. (1989). Daily growth increments in the larval otolith of the Japanese eel,
Anguillajaponica. Jap. J. Iehthyol. 35:440-444
Victor, B. C. (1986). Delayed metamorphosis with reduced larval
growth in a coral reef fish (Thalassoma bifasciatum). Can. J.
Fish. aquat. Science 43: 1208-1213
Volk, E. C., Wissmar, R. C., Simenstad, C. A., Eggers, D. M. (1984). Relationship between otolith microstructure and the
growth of juvenile chum salmon (Oncorhynehus keta) under