Association Analysis Between Tourette’s Syndrome and Two Dopamine Gene (DAT1, DBH) in Taiwanese Children
I-Ching Chou1,2, Wei-De Lin3,4, Chung-Hsing Wang1, Yu-Tzu Chang1, Zheng-Nan Chin1, Chang-Hai Tsai1,5,Fuu-Jen Tsai1,3
1 Departments of Pediatrics, Children’s Medical Center, China Medical University Hospital, Taichung, Taiwan
2 Graduate Institute of Integrated Medicine, College of Chinese Medicine, China Medical University, Taichung, Taiwan
3 Department of Medical Research, China Medical University and Hospital, Taichung, Taiwan
4 School of Post Baccalaureate Chinese Medicine, China Medical University, Taichung, Taiwan
5 Asia University, Taichung, Taiwan
Short title: DAT1 and DBH Polymorphisms in Tourette’s Syndrome Reprints and correspondence to: Fuu-Jen Tsai, MD, PhD.
Departments of Pediatrics and Medical Research, China Medical University Hospital No.2 Yuh-Der Road, Taichung, Taiwan
Telephone: 886-4-22052121 Ext 2066
Fax: 886-4-22032798, E-mail: d0704@ mail .cmuh.org.tw Abstract
defect in the dopamine system. Several candidate gene polymorphisms have been implicated in attention deficit hyperactivity disorder (ADHD), including dopamine transporter (DAT1) and dopamine beta hydroxylase gene (DBH). High rate of comorbidity between ADHD and TS indicates that they may share the same
pathophysiology. We aimed to test the hypothesis that the dopamine gene might play a role in TS.
Methods: By performing an association study, we collected an independent sample of patients from the midland region of Taiwan, and investigated whether DAT1 and DBH gene polymorphisms can be used as markers of susceptibility to TS. A total of 160 children with TS and 83 normal control subjects were included in the study. Polymerase chain reaction was used to identify the polymorphism of the DAT1 (40bp VNTR) and DBH (TaqI A2) gene. Genotypes and allelic frequencies for the DAT1 and DBH gene polymorphisms in both groups were compared.
Results: The results showed that genotypes and allelic frequencies in both groups were not significantly different. The most common genotype for DAT1 (40bp VNTR) in group 1 was 10/10 homozygote, and in group 2 was also 10/10 homozygote. The most common genotype for DBH (TaqI A2) in group 1 was T homozygote, and in group 2 was also T homozygote.
markers for prediction of the susceptibility of TS.
Key words: dopamine beta hydroxylase gene, dopamine transporter gene, polymorphism , Tourette’s syndrome
Introduction
Gilles de la Tourette syndrome (TS) is a neuropsychilatric disorder characterized by both motor and vocal tics. In addition, affected individuals frequently display symptoms such as attention deficit hyperactivity disorder and/or obsessive
compulsive disorder. In the 1970s, investigators first demonstrated that TS shows a familial concentration [1]. TS was then demonstrated to be transmitted vertically from generation to generation, and studies of twin pairs confirmed a genetic influence [2-3]. To date, the gene search in TS has been unsuccessful [4] and is illustrative of the many factors that can complicate genetic analysis of complex human traits [5].
The pathogenesis of TS remains obscure. Current evidence suggests that TS may result from a defect in the dopamine system [6-10]. Studies have focused mainly on the dopamine transporter gene (DAT1 40bp VNTR), and the dopamine beta hydroxylase gene (DBH TaqI A2) with attention deficit hyperactivity disorder (ADHD) [11-13]. ADHD is common in TS probands and is reported to affect about 50% to 70% of referred TS cases [14-16]. These observations lead us to test the polygenic hypothesis by examining the potential effect of DAT1 and DBH in TS. We previously used single nucleotide polymorphisms as a tool in genetic studies of polygenic disorders [17-21]. The single nucleotide polymorphisms are markers that may provide a new way to identify complex gene-associated diseases such as TS. In
this study, we tested the hypothesis that genetic variation in the DAT1 (40bp VNTR) and DBH (TaqI A2) gene confers susceptibility to TS. Two SNPs markers have been identified in, respectively, allowing researchers to detect disease-causing gene associations [22].
Methods
n=160 in DBH, respectively) and normal control subjects (group 2: n=83). This study was approved by the Ethics Committee of the China Medical University Hospital, Taichung, Taiwan. All parents signed informed consent before blood tests were performed. TS subjects and the controls were both recruited from the midland regions of Taiwan. Diagnosis of TS followed the criteria of the Diagnostic and Statistical Manual of Mental Disorders, 4th edition (DSM-IV) [23]. In the criteria for TS are as follows: the presence of multiple motor and at least one vocal tic (not necessarily concurrently); a waxing and waning course with tics evolving in a progressive manner; the presence of tic symptoms for at least 1 year; the onset of symptoms before age 21; the absence of a precipitation illness (e.g., encephalitis, stroke, or degenerative disease) or medication; the observation of tics by a knowledgeable neurologist; and marked distress or significant impairment in social, occupational, or other important areas of functioning. The pediatric neurologist, I-Ching Chou
examined the children and made sure that all cases were from unrelated kindred. The 83 controls were healthy volunteers with no history of psychiatric treatment.
All children underwent peripheral blood sampling for genotype analyses. Genomic DNA was isolated from peripheral blood by mean of a DNA extractor kit (Genomaker DNA extraction kit; Blossom, Taipei, Taiwan). A total of 50 ng of genomic DNA was mixed with 20 pmol of each polymerase chain reaction (PCR)
primer in a total volume of 25 ul containing 10mM Tris-hydrochloride, pH 8.3; 50mM potassium chloride; 2.0mM magenesium chloride; 0.2mM each
deoxyribonucleotide triphosphate; and 1 U of DNA polymerase (Amplitaq; Perkin Elmer, Foster City, Calif., USA). Four PCR primers were used to amplify the
correlated gene. The sequences of these primers were as following (from 5 to 3 end): DBH (444 g/a): upstream, CCTGGAGCCCAGTGCTTGTC; downstream,
ACGCCCTCCTGGGTACTCGC; and DAT1: upstream, TGTGGTGTAGGGAACGGCCTGAGA; downstream,
AAATTCCAGTGGGGTCCCTTCCTG. The PCR conditions were as follows: 35 cycles at 95ºC for 30 sec, 60º C for DBH(444 g/a), and 66.5º C for DAT1 for 30 sec, and 72ºC for 45 sec, then stood at 72C for 7 minutes and hold at 4C. The
polymorphisms were analyzed by PCR amplification followed by restriction analysis: EcoNI for DBH (444 g/a). The PCR products were directly analyzed on 3% agarose gel by electrophoresis, and each allele was recognized according to its size.
Allelic frequencies were expressed as a percentage of the total number of alleles. Genotypes and allelic frequencies for DAT1 and DBH polymorphisms in both groups were compared. The SAS system with 2 test was used for statistical analyses. A value of p<0.05 was considered statistically significant.
Results
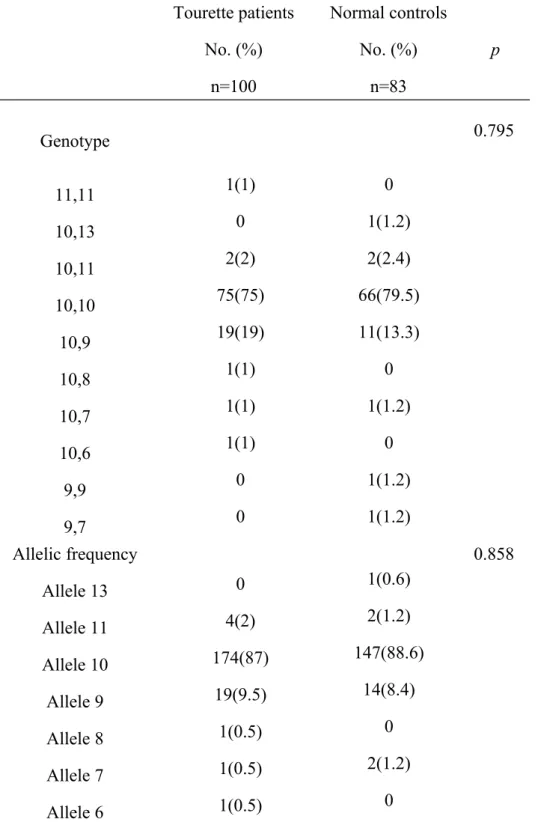
Genotype proportions and allele frequencies for the DAT1 and DBH in both groups were not significantly different (Tables 1, 2). The most common genotype for DAT1 in group 1 was 10/10 homozygote, and in group 2 was also 10/10 homozygote. The allele 10 frequencies for DAT1 in group 1 was 87% and in group 2 was 88.6% (Table 1).
Furthermore, the most common genotype for DBH in group 1 was T
homozygote, and in group 2 was also T homozygote. Proportions of T homozygote, T/C heterozygote and C homozygote for DBH were as follows: in group 1, 75.6%, 22.5% and 1.9%, respectively; and in group 2, 78.3%, 20.5% and 1.2%, respectively. The allele T and C frequencies for DBH in group 1 was 86.9% and 13.1%,
Discussion
Dopamine transport was first described 30 years ago [24]. DAT was itself identified and its molecular structure described a considerable number of years later [25]. The human DAT gene is localized on chromosome 5p15.3 [26]. A genetic polymorphism of a 40 bp variable tandem nucleotide repeat (VNTR) polymorphic sequence in the 3′ untranslated region of exon 15 of the gene is described [27]. This VNTR of exon 15 is repeated 3–11 times, most typically 10 times. The 10-repeat shows an ethnic heterogeneity with a frequency of 0.7 among Caucasians and Hispanics in USA, 0.54 in African Americans and 0.9 in Asians [28-30]. DATs are expressed in a small number of neurons in the brain, mainly in striatum and nucleus accumbens, but also in the globus pallidus, cingulate cortex, olfactory tubercle, amygdala and the midbrain [31]. The DAT, like the transporters for norepinephrine and serotonin, is a Na+/Cl− dependent transmembrane transport protein [32], which regulates the concentration of dopamine in the synaptic cleft.
DBH appears to be a strong candidate to investigate in TS, because it catalyzes the conversion of dopamine to norepinephrine and therefore influences both the dopaminergic and adrenergic systems. Serum DBH levels are under strong genetic control and show large interindividual variation [30]. Alleles of several
polymorphisms at the DBH locus have been found to be associated with serum DBH levels.
In the present study we did not find significant evidence for association in our TS samples. The role of dopaminergic system in the pathogenesis of TS is still unknown. Preliminary studies have suggested that the pathogenesis of tics involves neuronal activity within subcortical neuronal circuits [34]. Therefore, the classic
neurotransmitters, dopamine and serotonin, raised the possibility that they may be involved in the pathobiology of TS. However, other investigators have emphasized that abnormalities of dopamine fail to explain many clinical and laboratory
observations, including the description of unchanged tics in four adults who developed parkinsonism and received treatment with L-dopa [35].
Our review of the literature found that recent linkage studies have yield no positive results to dompaine D1-5 receptors [36-38], glycine alpha 1 subunit, GABAA receptor alpha-1, alpha-6, and gamma-2 subunits (GABRA1, GABRA6, GABRG2). GABAA receptor beta-1 and alpha-2 subunits (GABRB1, GABARA2), glutamate receptor GLUR1, the alpha adrenergic receptor ADRA1, the beta adrenergic receptor ADRB1 and the glucocorticoid receptor GRL [39]; norepinephrine transporter gene [40]; and catechol-o-methyltransferase [41]. Other investigators have sought to identify associations between TS and other movement disorders [42]. Further studies
will be required to confirm these assertions.
The etiology of TS is therefore unknown. In fact, TS of children may involve a complex interaction between the environmental influences, especially infection, autoimmune contributions, epigenetic factors and genetic factors. Our study suggests that the DBH and DAT1 gene may not contribute to the etiology of TS. Further studies could be focused on the analysis of other dopaminergic genes in TS patients. Our results could provide the database for further survey of the DBH and DAT1 gene polymorphism.
Acknowledgements
This study was supported in part by the China Medical University Hospital (grant number DMR-102-038).
Conflict of Interest:
All authors fulfilled the condition for authorship. There was no commercial support in performing this study and submitting this manuscript.
syndrome: clinical, genetic, psychologic, and biochemical aspects in 21 selected families. Neurology 1977;27:115-24.
2. Price RA, Kidd KK, Cohen DJ, Pauls DL, Leckman JF. A twin study of Tourette syndrome. Arch Gen Psychiatry 1985;42:815-20.
3. Pauls DL, Leckman JF. The inheritance of Gilles de la Tourette's syndrome and associated behaviors. Evidence for autosomal dominant transmission. N Engl J
Med 1986;315:993-7.
4. Heutink P, Breedveld GJ, Niermeijer MF, et al. Progress in gene localization. In Handbook of Tourette’s syndrome and related tic and behavioral disorders.
Kurlan R (editor). New York: Marcel Dekker 1993;317-35.
5. Comings DE. Tourette's syndrome and psychiatric disorders. Br J Psychiatry 1995; 166:399.
6. Leckman JF, Riddle MA, Cohen DJ. Pathobiology of Tourette’s syndrome. In Cohen DJ, Bruun RD, Leckman JF (eds): “ Tourette’s syndrome and tic disorder: clinical understanding and treatment.” New York: Wiley & Sons, 1992;103-16. 7. Shapiro E, Shapiro AK, Fulop G, Hubbard M, Mandeli J, Nordlie J, et al.
Controlled study of haloperidol, pimozide and placebo for the treatment of Gilles de la Tourette's syndrome. Arch Gen Psychiatry 1989; 46:722-30.
8. Jankovic J, Glaze DG, Frost JD. Effect of tetrabenazine on tics and sleep of Gilles de la Tourette's syndrome. Neurology 1984; 34:688-92.
9. Sweet RD, Bruun R, Shapiro E, Shapiro AK. Presynaptic catecholamine antagonists as treatment for Tourette syndrome: Effects of alpha methyl para tyrosine and tetrabenazine. Arch Gen Psychiatry 1974; 31:857-861.
10. Golden GS. The use of stimulants in the treatment of Tourette’s syndrome. In Cohen DJ, Bruun RD, Leckman JF (eds):”Tourette’s syndrome and tic disorder: Clinical understanding and treatment.” New York: Wiley & Son 1992;317-25. 11. Cook EH Jr, Stein MA, Krasowski MD, Cox NJ, Olkon DM, Kieffer JE, et al.
Association of attention-deficit disorder and the dopamine transporter gene. Am
J Hum Genet 1995;56:993-8.
12. Daly G, Hawi Z, Fitzgerald M, Gill M. Mapping susceptibility loci in attention deficit hyperactivity disorder: preferential transmission of parental alleles at DAT1, DBH and DRD5 to affected children. Mol Psychiatry 1999;4:192-6. 13. Smith KM, Daly M, Fischer M, Yiannoutsos CT, Bauer L, Barkley R, et al.
Association of the dopamine beta hydroxylase gene with attention deficit
hyperactivity disorder: genetic analysis of the Milwaukee longitudinal study. Am
J Med Genet B Neuropsychiatr Genet 2003;119:77-85.
14. Comings DE, Comings BG. Tourette syndrome: clinical and psychological aspects of 250 cases. Am J Hum Genet 1985;37:435-50.
15. Comings DE, Gade R, Wu S, Chiu C, Dietz G, Muhleman D, et al. Studies of the potential role of the dopamine D1 receptor gene in addictive behaviors. Mol
Psychiatry 1997;2:44-56.
16. Comings DE, Comings BG. Tourette's syndrome and attention deficit disorder with hyperactivity. Arch Gen Psychiatry. 1987;44:1023-6
17. Sun SS、Chou IC、Lai YH、Kao CH. 99mTc-ECD SPECT image in children with Gilles de la Tourette’s syndrome: a preliminary report. Ann Nucl Med Sci 2001:149-53.
18. Chen WY, Lin CY, Chou IC, Tsai FJ, Sun SS. The Role of Tc-ECD Brain SPECT in differentiating Tourette's Syndrome from Chronic Tic Disorder. Ann Nucl Med Sci. 2003;16:59-63.
19. Chou IC, Tsai CH, Lee CC, Kuo HT, Hsu YA, Li CI, et al. Association analysis between Tourette’s syndrome and dopamine D1 receptor gene in Taiwanese children. Psychiatr Genet.2004;14:219-21.
20. Chou IC, Tsai CH, Wan L, Tsai FJ. Association study between Tourette's syndrome and polymorphisms of noradrenergic genes (ADRA2A,ADRA2C).
Psychiatr Genet.2007:359.
21. Chang YT、Li YF、Muo CH、Chen SC、Chen CN、Kuo HT、Lin HC、Sung FC、Tsai CH、Chou IC. Correlation of Tourette’s syndrome and allergic disease: nationwide population based case-control study. J Dev Behav Pediatr.2011;
2:98-102.
22. Internet at http:// www.ncbi.nlm.nih.gov/SNP
23. Comings DE. DSM-IV criteria for Tourette's. J Am Acad Child Adolesc
Psychiatry1995;34:401-2.
24. Iversen LL. Role of transmitter uptake mechanisms in synaptic neurotransmission. Br J Pharmacol 1971;41:571-91.
25. Giros B, el Mestikawy S, Godinot N, Zheng K, Han H, Yang-Feng T,et al. Cloning, pharmacological characterization, and chromosome assignment of the human dopamine transporter. Mol Pharmacol 1992;42:383-90.
26. Donovan DM, Vandenbergh DJ, Perry MP, Bird GS, Ingersoll R, Nanthakumar E, et al. Human and mouse dopamine transporter genes: conservation of 5′-flanking sequence elements and gene structures. Brain Res Mol Brain Res 1995;30:327–5.
27. Vandenbergh DJ, Persico AM, Hawkins AL, Griffin CA, Li X, Jabs EW, et al. Human dopamine transporter gene (DAT1) maps to chromosome 5p15.3 and displays a VNTR. Genomics 1992;14:1104–6.
28. Byerley W, Hoff M, Holik J, Caron MG, Giros B. VNTR polymorphism for the human dopamine transporter gene (DAT1). Hum Mol Genet 1993;2:335.
29. Sano A, Kondoh K, Kakimoto Y, Kondo I. A 40-nucleotide repeat polymorphism in the human dopamine transporter gene. Hum Genet 1993;91:405–6.
30. Doucette-Stamm LA, Blakely DJ, Tian J, Mockus S, Mao JI. Population genetic study of the human dopamine transporter gene (DAT1). Genet Epidemiol 1995;12:303–8.
31. Leighton PW, Le Couteur DG, Pang CC, McCann SJ, Chan D, Law LK, et al. The dopamine transporter gene and Parkinson's disease in a Chinese population.
Neurology 1997;49:1577–9.
32. Nakatome M, Honda K, Tun Z, Kato Y, Harihara S, Omoto K, et al. Genetic polymorphism of the 3′ VNTR region of the human dopaminergic function gene DAT1 (human dopamine transporter gene) in the Mongolian population. Hum
Biol 1996;68:509–15.
33. Craig SP, Buckle VJ, Lamouroux A, Mallet J, Craig IW. Localization of the human dopamine beta hydroxylase (DBH) gene to chromosome 9q34. Cytogenet
34. Peterson BS, Skudlarski P, Anderson AW, Zhang H, Gatenby JC, Lacadie CM, et al. A functional magnetic resonance imaging study of tic suppression in Tourette syndrome. Arch Gen Psychiatry 1998;55:326-33.
35. Kumar R, Lang AE. Coexistence of tics and parkinsonism: evidence for non-dopaminergic mechanisms in tic pathogenesis. Neurology 1997; 49:1699-701. 36. Barr CL, Wigg KG, Zovko E, Sandor P, Tsui LC. No evidence for a major gene
effect of the dopamine D4 receptor gene in the susceptibility to Gilles de la Tourette syndrome in five Canadian families. Am J Med Genet 1996; 67:301-5. 37. Barr CL, Wigg KG, Zovko E, Sandor P, Tsui LC. Linkage study of the dopamine
D5 receptor gene and Gilles de la Tourette syndrome. Am J Med Genet 1997;74:58-61.
38. Devor EJ, Dill-Devor RM, Magee HJ. The Bal I and Msp I polymorphisms in the dopamine D3 receptor gene display, linkage disequilibrium with each other but no association with Tourette syndrome. Psychiatr Genet 1998; 8:49-52.
39. Brett PM, Curtis D, Robertson MM, Gurling HM. Neuroreceptor subunit genes and the genetic susceptibility to Gilles de la Tourette syndrome. Biol Psychiatry 1997; 42:941-7.
40. Stober G, Hebebrand J, Cichon S, Brüss M, Bönisch H, Lehmkuhl G, et al. Tourette syndrome and the norepinephrine transporter gene: results of a
41. Barr CL, Wigg KG, Sandor P. Catechol-O-methyltransferase and Gilles de la Tourette syndrome. Mol Psychiatry 1999; 4:492-5.
42. Nemeth AH, Mills KR, Elston JS, Williams A, Dunne E, Hyman NM. Do the same genes predispose to Gilles de la Tourette syndrome and dystonia? Report of a new family and review of the literature. Mov Disord 1999;14:826-31.
Table 1. Genotypes for DAT1(40bp VNTR) polymorphisms in children with Tourette’s syndrome and normal controls
Tourette patients No. (%) n=100 Normal controls No. (%) n=83 p Genotype 1(1) 0 2(2) 75(75) 19(19) 1(1) 1(1) 1(1) 0 0 0 1(1.2) 2(2.4) 66(79.5) 11(13.3) 0 1(1.2) 0 1(1.2) 1(1.2) 0.795 11,11 10,13 10,11 10,10 10,9 10,8 10,7 10,6 9,9 9,7 Allelic frequency Allele 13 Allele 11 Allele 10 Allele 9 Allele 8 Allele 7 Allele 6 0 4(2) 174(87) 19(9.5) 1(0.5) 1(0.5) 1(0.5) 1(0.6) 2(1.2) 147(88.6) 14(8.4) 0 2(1.2) 0 0.858
* p-value were calculated by 2 test.
Table 2. Genotypes and allele frequencies for DBH (TaqI A2) polymorphisms in children with Tourette’s syndrome and normal
controls Tourette patients No. (%) n=160 Normal controls No. (%) n=83 p Genotype T/T 0.900 121(75.6) 65(78.3) C/T 36(22.5) 17(20.5) C/C Allelic frequency 3(1.9) 1(1.2) 0.700 Allele T 278(86.9) 147(88.6) Allele C 42(13.1) 19(11.4) * p-value were calculated by 2 test.