行政院國家科學委員會補助專題研究計畫成果報告
※※※※※※※※※※※※※※※※※※※※※※※※※
※ ※
※ 台灣海峽生態系統動態整合研究 子計畫一: ※
※
台灣海峽浮游生物之生產力及食物階層動態(II)
※
※※※※※※※※※※※※※※※※※※※※※※※※※
計畫類別:□個別型計畫
■整合型計畫
計畫編號:NSC90-2611-M-110-016-OP4
執行期間:90 年 8 月 1 日至 91 年 7 月 31 日
計畫主持人:陳宏遠
共同主持人:
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
執行單位:國立中山大學海洋生物研究所
中
華
民
國
92 年
3 月 25 日
行政院國家科學委員會專題研究計畫成果報告
台灣海峽生態系統動態整合研究 子計畫 1:
台灣海峽浮游生物之生產力及食物階層動態 II
Taiwan Strait Ecosystem Dynamics Interdisciplinary Project
Plankton pr oductivity and tr ophodynamics in the Str ait of Taiwan II
計畫編號:NSC 90-2611-M-110-016-OP4
執行期限:90 年 8 月 1 日至 91 年 7 月 31 日
主持人:陳宏遠 國立中山大學海洋生物研究所
計畫參與人員:康一凡、黃珠美、陳攸里
I. 中文摘要 本研究擬探討台灣海峽冬季 (November, 2002) 浮游動物之空間分布,所採用的生物資料包括總葉 綠素甲濃度、<20µm 葉綠素濃度、<3µm 葉綠素 濃度、浮游動物密度、浮游動物比例;非生物因子 包括物理學及化學分析結果。若只以生物資料來進 行主成分分析,結果發現海峽可以擬略分為三環 境;海峽中線的高生產力海域,中國沿岸海域及台 灣沿岸海域。其中在海峽中線的最高 Chl a 測站及 台灣北部沿海測站並不屬於上述三環境中。海峽 中,Chl a 濃度隨水溫升高而增加,隨 NO3濃度及 NO3 / SRP 增加而減少。浮游動物現存量正比於各 體型浮游植物之現有量,但不包括>20µm 體型 者。橈足類 (200-500µm) 現存量正比於總浮游動物 現存量,並與<3µm Chla 與<20µm Chla 是正相 關,而與>20µm Chl a 無顯著相關。 關鍵詞:台灣海峽、浮游植物、浮游動物、體型、 食物階層動力學 AbstractSpatial distribution of zooplankton assembly in the Taiwan Strait during wintertime (November, 2002) were analyzed by incorporating biological data including biomasses (measured by Chla concentration) of total, <20µm and <3µm phytoplankton, zooplankton abundance, percentage occurrences of major zooplankton groups and physical and chemical hydrographic factors. When non-biological measurements were excluded, the results of principal component analyses indicate that the Strait can be classified into 3 environments with slight overlapping: a productive area in the middle of the Strait where a thermal and pigment front was located; the water surrounding the western coast of Taiwan and the water along the coast of china. The station on the front with high Chla concentration and a coastal station could not be assigned to the above 3 environments. Chla concentration increased with increasing water temperature, decreased with increasing NO# concentration and NO3/SRP. Zooplankton abundance
were positively regressed to Chla concentration of all size fractions except those >20 µm. Zooplankton biomass in the Strait showed a significant and positive correlation with the occurrence of copepods (200-500µm). The standing stock of the copepods related positively to<3µm Chla and<20µm Chla, and not related to>20µm Chla. It becomes obvious that the distribution of phytoplankton and zooplankton varied seasonally, with productivity center occurring in the northern Strait in the summer and in the mid-strait in the winter. Top-down control by zooplankton grazing on primary producer was suspected to be an importance forcing in the summer; the results from the wintertime study did not support the assertion. Seasonal variation in control forcing is suspected.
Keywords: Taiwan Strait, phytoplankton, zooplankton, size-fractionation, trophodynamics II. Intr oduction
The objective of the present study was to describe the trophic relationships among phytoplankton and zooplankton in the Taiwan Strait. The Taiwan Strait is a meridional channel that allows water exchange among the East China Sea, the South China Sea and the Kuroshio Current. The combinations of seasonal alternation of the northeastern monsoon in winter and the southwestern monsoon in summer and the bottom topography of the Strait determine the circulation (Jan et al., 2002). Although there is often a northward current on the Taiwan side and a southward current in the Mainland side, the controlling forcing leads to winter blocking of the northward current from the Kuroshio, spring renewal of the northward intrusion, minimal blocking of northward intrusion in summer by the water from the South China Sea and fall emergence of China Coastal Current. Oligotrophic water from the Kuroshio and the South China Sea meets relatively eutrophic water of China Coastal Current in the Strait with clear seasonal distinction. In warm seasons, warm, saline and oligotrophic water flood the Strait; While cold,
diluted and eutrophic water occupies much of the Mainland side and the northern part of the Strait during winter. Previously, based on the results of a cruise conducted in June of 2002, we found that the Strait can be classified into 3 distinctive environments: a productive Chinese coastal water in the northeastern corner of the Strait near the river mouth of the Meng-Chiang; a medium productive range in the Taiwan side of the northern Strait, opposite to the productive Chinese coastal water; and an oligotrophic water covering the rest of the strait, including the mid-strait and the Peng-Hu Channel where the South China Sea water enters the Strait. In the productive Chinese coastal water, phytoplankton abundance expressed by chlorophyll a concentration was high and most of the phytoplankton were larger than 20µm. The large sized (>20µm) phytoplankton supported a zooplankton population relative high in carnivores such as sagittae and lucifers. The least productive waters covering much of the Strait, on the other hand, were characterized by <20µm phytoplankton and high occurrence of copepods. The unproductive waters in the Peng-Hu Channel and along the Taiwan coast were characterized by the abundance (>30%) of dinoflagellate Noctiluca sp.
The present paper reports the summary results of the cruise observations in the Taiwan Strait during the second year of the ‘Taiwan Strait Ecosystem Dynamics’ interdisciplinary project. Although effective sampling in wintertime in the Taiwan Strait has been difficult, we were able to complete a relatively thorough survey in the Strait in November of 2002 when the northeastern monsoon prevailed in the Strait. The data set represents an observation of the hydrographic and biological variations during the cold season.
III. Mater ials and Methods
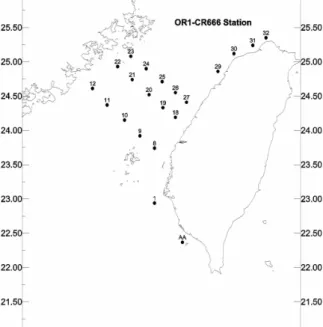
We collected plankton samples and measured physiochemical properties of seawater in the Taiwan Strait during a cruise (OR666) aboard R/V Ocean Researcher I in November 24-30, 2002. Fig. 1 depicts the location of the 50 preset sampling stations for the integrated project. In the present study, 21 stations were sampled, including Stations 1, 8, 9, 10, 11,12, 18, 19, 20, 21, 22, 23, 24, 25, 26 27, 28, 29, 30, 31 and 32. CTD data and seawater samples were collected using 20-l Go-Flo bottles attached to a rosette multi-sampler mounted on a CTD (SBE 9/11plus, SeaBird Inc., WA). Plankton samples were collected in oblique tows using a three-layered plankton net, each with a mesh size of 20, 200 and 500 µm, respectively. The setup allowed simultaneous collection of 3 size classes: 20-200 µm, 200-500 µm and >500 µm. Two equal portions of each size-fractionated plankton sample were taken and one portion was preserved with neutral formalin for taxonomic examination; and the other portion was picked over to collect larval fish and then rinsed with
distilled water, blotted dry and kept at -20oC. Larval fish were also washed and frozen.
Figure 1. Location of the sampling stations during the cruise ORI cr666 in the Taiwan Strait.
Size fractionated (<20 µm and > 20 µm) estimations of chlorophyll a concentration were carried out by on board filtration procedures. Chlorophyll a concentration was determined fluorometrically on extracted samples. Concentrations of nitrate and phosphate were measured following the pink azo dye method and the molybdenum blue method, respectively. Particulate organic matter was obtained by filtering seawater with pre-combusted GF/F filters.
All frozen samples (particulate organic matter, plankton and larval fish) were freeze-dried, pulverized and frozen until isotope analysis. Quantitative determinations of δ15N were carried out using an ANCA GC-mass spectrometer (Europa Scientific, Crewe, UK). Measurements were made relative to the isotopic compositions of and nitrogen in air (δ15N), where
δ15N = [(Rsample-Rstandard)-1] x 1000 and
R = 15N/14N
Analysis of replicates usually showed agreement at the 0.2‰ levels for δ15N or better. The isotope data was not reported here because the maintenance problems with the ANCA.
Regression analyses between size-fractionated zooplankton abundance and chla concentrations of coupling phytoplankton biomass were performed. Principal component analysis was performed to describe the distribution of the zooplankton in the Strait and its coupling with the environment.
Non-biological data including temperature, salinity and concentrations of nitrate, phosphate and particulate organic nitrogen as well as biological data including chlorophyll concentration, densities of major zooplankton groups were tested exclusively or concurrently.
IV. Results and Discussion
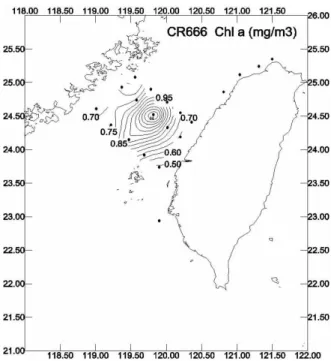
The cruise 666 was conducted in November when the Northeastern Monsoon prevailed in the Taiwan Strait. The China Coastal Current flooded the northern part of the Strait and the northward Kuroshio occupied the southern half. The two water masses met in the center of the Strait. Satellite image of the sea surface water temperature taken during the cruise time (Fig.2) clearly indicated the existence of a thermal front at where the two water masses engaged each other. The front occurred next to the Chiang-Yun Rise. A significant branch of the strong China Coastal Current with the assistance of the prevailing NE monsoon crossed the Strait from the Mainland side to the Taiwan side. A thermal front center was clearly observed at water near Station 20. Similarly, distribution of surface Chla concentration shows a highest value at Station 20. It then decreased towards both coasts of the Strait. The patterns of <3
µm and 3-20 µm size fractions of phytoplankton are similar to that of the total chla concentration, i.e., no difference was found between the two size fractions. In our observation in the summer of 2001, small phytoplankton (<3µm) became predominated towards the southern half of the Strait. Surface nitrate concentration shows a decreasing trend from the Chinese coast towards the Taiwan coast, with 10µM near the China coast and1 µM near Taiwan coast. There was n o high value of nitrate concentration as
Figure 2. Satellite image of sea surface water
temperature distribution in waters surrounding Taiwan
during the cr666 cruise.
Fig.3 Surface chlorophyll a concentration distribution in Taiwan Strait, measured in mid-November, 2002. seen in the surface temperature and Chla concentration profiles. Distribution of (NO3+NO2)/SRP also shows similar trend as does the nitrate concentration pattern.
The results of regression analyses indicated some significant relationship between phytoplankton biomass (Chla concentration) and zooplankton abundance (individual per m3): Both of the abundance of >200µm and >500µm zooplankton were positively regressed to total Chla concentration (P=0.044 and 0.048, respectively). The abundance of 200-500µm zooplankton was positively regressed to Chla concentration of <20µm phytoplankton (P=0.047). The abundance of200-500µm zooplankton was positively regressed to Chla concentration of <3µm phytoplankton (P=0.016). Multiple regression analyses show that the abundance of Noctiluca, a >200µm dinoflagellates, was positively regressed to surface water temperature and negatively regressed to nitrate concentration. The abundance of 200-500µm copepods was positively regressed to total Chla concentration, <3µm Chla and <20µm phytoplankton, but not >20µm phytoplankton.
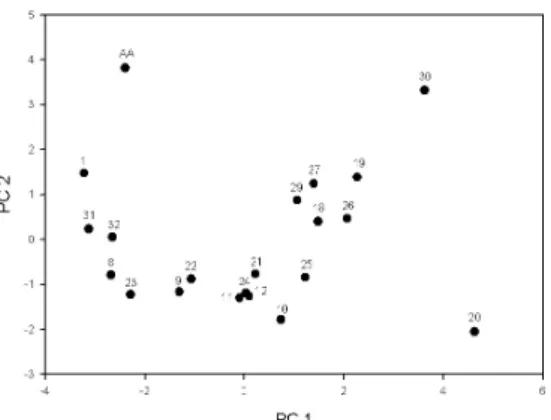
When all size fractions of Chla and >200µm zooplankton were compared, principle component analyses reveal that the first axis is an expression of biomass of all phytoplankton, <3µm phytoplankton and <20µm phytoplankton. The second axis is a description of changes mainly in abundances of ostracoda and Noctiluca. The first 2 axes alone are accounted for 68.0% of the total variation in the data set of plankton community and environmental factors. Scatter diagram of PCA 2 against PCA 1 (Figure 4)
shows that the 21 stations can be divided into several clusters. The first cluster has 5 stations (AA, 1, 31,32 and 8) located along the coast of Taiwan. The second cluster was constituted of Stations 18, 19, 21, 25, 26, 27 and 29. These 6 stations were located at the front where the two water masses met. Seven stations (9, 10, 11, 12, 21, 22, 23 and 24) could be grouped into the third cluster. These stations were more affected by the China Coastal Current. Two stations, 20 and 30, could not be grouped into the above three groups. Station 20 was the front station and station 30 was located at northern coast of Taiwan where high pigment concentration was observed.
Figure 4. Scatter diagram of plots of the first two PCA axes of plankton community structure data. Numbers correspond to the sampling stations.
Unlike in summer when the Strait of Taiwan is widely flooded with warm and oligotrophic water from the South China Sea and phytoplankton cell size is mostly small than 20 µm, winter phytoplankton assembly did not show geographical size spectrum pattern. In the northern Strait, water was entrophic, especially along the coast of China.
It becomes obvious that the distribution of phytoplankton and zooplankton varied seasonally, with productivity center occurring in the northern Strait in the summer and in the mid-strait in the winter. Top-down control by zooplankton grazing on primary producer was suspected to be an importance forcing in the summer (Acuna et al. 2002; Strom et al. 2001); the results from the wintertime study did not support the assertion. Seasonal variation in control forcing is suspected.
V. Refer ence
Acuna, J. L., Deibel, D., Saunders, P. A., Booth, P., Hatfielf, E., Klein, B., Mei, Z. P., Rivkin, R. (2002) Phytoplankton ingestion by
appendicularians in the North Water. Deep_sea Research II 49: 5101-5115.
Jan, S., Wang J., Chern, C. S., and Chao, S. Y.(2002) Seasonal variation of the circulation in the Taiwan Strait. J. Mar. Syst. 35: 249-268.
Jan, S., Chern, C. S., and Wang, J.(2002)Transition of
Tidal waves from the East to South China Seas over the Taiwan Strait: influence of the abrupt step in the topography. J. Oceanogr. 58: 837-850. Kidwai, S. and Amjad, S. (2000) Zooplankton:
pre-southwest and northeast monsoons of 1993 to 1994, from the North Arabian Sea. Mar. Biol. 136: 561-571.
Hulot, F. D., Lacroix, G., Lescher-Moutoue, F. and Loreau, M. (2000) Functional diversity governs ecosystem response to nutrient enrichment. Nature 405: 340-344.
Lalli, C. M. and Parsons, T. R (1993) Biological oceanography: an introduction. Pergamon Press, Oxford.
Strom, S. L., Brainard, M. A., Holmes, J. L. and Olson, M. B. (2001) Phytoplankton blooms are strongly impacted by microzooplankton graing in coastal North Pacific waters. Mar. Biol. 138: 355-368.