行政院國家科學委員會補助專題研究計畫成果報告
探討抑制人類白血球誘發基質金屬蛋白酵素活化之藥物機轉及其對
活體再甦醒性傷害的保護作用之評估 (2/2)
The study of inhibitory mechanisms of anti-leukocyte agents on matrix metalloproteinase activation and evaluate the protective effects on resuscitation
injury in vivo (2/2)
計畫類別:個別型計畫
計畫編號:NSC 92-2320-B038 -016
執行期間: 92 年 08 月 01 日至 93 年 07 月 31 日
計畫主持人:蕭哲志 共同主持人:許準榕
計畫參與人員: 崔巧琪、黃麗曲
成果報告類型(依經費核定清單規定繳交): 完整報告
本成果報告包括以下應繳交之附件:
□赴國外出差或研習心得報告一份
□赴大陸地區出差或研習心得報告一份
□出席國際學術會議心得報告及發表之論文各一份
□國際合作研究計畫國外研究報告書一份
處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、
列管計畫及下列情形者外,得立即公開查詢
□涉及專利或其他智慧財產權,□一年□二年後可公開查 詢
執行單位:臺北醫學大學藥理學科 中 華 民 國 93 年 10 月 28 日
探討抑制人類白血球誘發基質金屬蛋白酵素活化之藥物機轉及其對活體再甦醒 性傷害的保護作用之評估 (2/2)
The study of inhibitory mechanisms of anti-leukocyte agents on matrix
metalloproteinase activation and evaluate the protective effects on resuscitation injury in vivo (2/2)
計畫編號:NSC 92-2320-B038 -016 執行期限:92 年 08 月 01 日至 93 年 07 月 31 日
主持人:蕭哲志 共同主持人:許準榕 一、中文摘要
Matrix metalloproteinases 簡 稱 MMPs,為一群結構類似且含鋅(zinc)金屬 離子之蛋白酵素。因MMPs的催化作用需 金屬離子加以活化,並且能夠催化分解維 持組織結構之細胞外基質蛋白,包括基質 與結締纖維組織,故稱之為基質金屬蛋白 酵素,而其對於組織之結構重組、修補與 破壞都扮演相當重要之角色。在大規模中 藥材萃取物及化學合成等藥物之成分篩選 實 驗 中 , 我 們 發 現 在histone deacetylase inhibitor II (HDI II) 及 cinnamophilin,皆 具有明顯抑制MMPs活化之作用。在電泳 酵素分析法中觀察到HDI II (0.01-5 µM)及 cinnamophilin (1-50 µM)確實有意義地依 濃度效應抑制對於LPS或MCP-1 誘發人類 單核球細胞之MMP-9 活性,此外以細胞存 活 率 測 定(MTT assay) 發 現 HDI II 及 cinnamophilin的抑制作用並非源自細胞之 損害。西方點墨法(Western blot)實驗發現 在 不 同 刺 激 下( 如 LPS 或 MCP-1) 細 胞 MMP-9 protein的表現量會隨著HDI II及 cinnamophilin濃度的增加而降低,故可證 實此兩種藥物作用在MMP-9 蛋白質表現 層面。並進一步以RT-PCR的實驗加以分 析 , 發 現HDI II 及 cinnamophilin 會 抑 制 MMP-9 mRNA的表現,更深入瞭解細胞轉 錄(transcription)之影響程度。同時我們也 更進一步探討HDI II及cinnamophilin在訊 息傳遞中作用機轉的方式,從實驗結果得 知cinnamophilin會明顯抑制由LPS刺激所 導致Inhibitor-κB-α (IκB-α)的降解作用,使
Nuclear factor-κB (NF-κB)無法進入細胞 核中與特定MMP-9 相關的DNA序列接 合 。 在Mitogen-activated protein kinases (MAPKs)方面,從目前實驗結果得知HDI II 及 cinnamophilin 對 於 LPS 誘 發 c-Jun-NH2-terminal kinase (JNK)活化具抑 制 作 用 , 但 對 於 extracellular signal-regulated kinases (ERKs)並無直接的 影響。HDI II及cinnamophilin對於MCP-1 誘發細胞趨化作用具抑制效果。另外,從 流 式 細 胞 儀 的 結 果 發 現 HDI II 及 cinnamophilin並不會抑制THP-1 活化後細 胞表面CD11b的表現。活體再甦醒性傷害 實驗中,的確發現實驗動物依時間關係其 肝臟與腎臟功能受損。在目前活體藥物處 理下HDI II類似作用藥valproic acid效果不 佳,而以抗白血球作用藥YC-1 具部分改善 效果。
綜合目前實驗的結果,發現HDI II 及 cinnamophilin 的確具有抑制 MMP-9 表現 之活性,而在 LPS 或 MCP-1 刺激方面其
作用機轉可能主要藉由影響 NF-κB 或
JNK 的訊號傳遞過程。未來也將會進行更 多相關之實驗及其他活體實驗以瞭解其是 否具有抗發炎療效之功能。
關鍵詞: 基質金屬酵素、細胞訊息、再甦 醒性傷害
Abstract
Matrix Metalloproteinases (MMPs) are a family of over 20 zinc-containing enzymes that cleave the various components of extracellular matrix. Because the catalytic
ability of MMPs needs to be activated by metal ions, and because they could catalyze and degrade tissue structure maintaining extracellular matrix protein (ECM), including ground substances and connecting fibers, they are named matrix metalloproteinase. Thus, it plays an important role in tissue structure remodeling, repairing and destroys. According to previous experiments, we found that histone deacetylase inhibitor II (HDI II) and cinnamophilin showed obviously inhibitory effect on MMPs activation. We observed that HDI II and cinnamophilin significantly and concentration-dependently inhibit MMP-9 activation induced by LPS and MCP-1 by zymographic method. Also, we found that the inhibitory effect of HDI II and cinnamophilin was not due to impairment of cellular viability by MTT tests.
According to Western blot method, we found that various stimulator-induced expression of MMP-9 protein is concentration-dependent inhibition by HDI II and cinnamophilin. This indicated that these two compounds have effect on the protein expression of MMP-9. By using RT-PCR method, we found that HDI II and cinnamophilin can inhibit the expression of MMP-9 mRNA, thus have deeper influence on the level of MMP-9 transcription. At the same time, we investigated the mechanism of action of HDI II and cinnamophilin in various signaling pathways. We found that cinnamophilin could significantly inhibit the degradation of inhibitor-κB-α (IκB-α) induced by LPS. Therefore, nuclear factor-κB (NF-κB) may not translocate for
transcription. Furthermore, in mitogen-activate protein kinases (MAPKs) aspect, HDI II and cinnamophilin showed direct influence on phosphorylated activation of c-Jun-NH2-terminal kinases (JNK) activation. However, both reagents did not show direct influence on phosphorylated activation of extracellualr signal-regulated kinases (ERKs) activation.
Besides, the results of flow cytometry showed that HDI II and cinnamophilin did not inhibit the expression of surface protein, CD11b, on THP-1 cells. In summary, we
found that HDI II and cinnamophilin have inhibitory effect on MMP-9 expression, and its main mechanism of action might through NF-κB or MAPK signal pathway on LPS and MCP-1 stimulation. According to the resuscitation injury in vivo studies, we found the functions of liver and kidney are graduately decay by the time. Valproic acid as function as HDI II exerted slightly effect, though, anti-leukocyte agent, YC-1 exerted some protective effects. It will be interesting to further investigate its anti-inflammatory therapeutic profile via other related experiments and animal model in vivo.
Keywords: Matrix Metalloproteinases, signal transduction, the resuscitation injury
二、緣由與目的
2-1 Matrix metalloproteinases 簡介 2-1-1. Structure and Function
Matrix metalloproteinases 簡 稱 MMPs,為一群結構類似且含鋅(zinc ion) 金屬離子之蛋白酵素(Kotra et al., 2001)。
MMP 的催化作用需金屬離子加以活化,
並 有 催 化 水 解 細 胞 外 基 質 蛋 白 (extracellular matrix proteins)的能力。且其 特 有 的 基 因 序 列 可 加 以 區 別 相 近 的 MMPs,因而稱之為基質金屬蛋白酵素 (Stöcker et al., 1995)。目前已有 28 種 MMPs 被識別出來,並依其只要催化之結締性受 質 成 分 被 分 成 五 類 : gelatinases 、 stromelysins 、 membrance-bound enzymes 、collagenases 與其它 MMPs,
且持續的在增加中(Sternlicht and Werb, 2001)。(A)~(H) 依其功能與結構區分成八 類,以茲瞭解MMPs 序列的相互關係:
(A) Minimal Domain MMPs: MMP-7, 26 (B) Simple Hemopexin Domain-Containing MMPs: MMP-1, 3, 8, 10, 12, 13, 18, 19, 20, 22, 27
(C) Gelatin-binding MMPs: MMP-2 and MMP-9 (which are focused studying in our lab)
(D) Furin-activated Secreted MMPs:
MMP-11, MMP28
(E) Transmembrane MMPs: MMP-14, 15, 16, 27
(F) GPI-linked MMPs: MMP-17, MMP-25 (G) Vitronectin-like Insert Linker-less MMPs: MMP-21
(H) Cystein/Proline-Rich IL-1 Receptor-like Domain MMPs: MMP-23
所有的MMPs 都有N-terminal signal sequence (或叫做 “pro” domain),會在它 合成進入ER後被移除掉。大部分的MMPs 也因此而被分泌出來。N-terminal sequence 可 說 是target 了 MMPs 的 分 泌 。 然 而 , MMPs 有 六 個 transmembrane domains 並 且以細胞表面的酵素被表現。在MMPs的 主要結構骨架至少可分為三部分, MMPs 的pro domain在pre domain之後,有維持酵 素latency的功能,通常被酵素抑制呈不活 化 狀 態 。 以MMP-7 為 例 (Gaire et al., 1994),第一段為signal sequence,第二段 為游離的thiol (SH) group peptide,及一段 含鋅離子之催化區。Pro domain含有保守 的 “systeine switch” sequence 和 中 央 的 cysteine residue在活化的區域與zinc 結合 (Nagase &Woessner, 1999)。在proteolytic cleavage之後,被抑制的區域就會被釋放,
而酵素因此而活化。Latent形式的MMPs 會 在 許 多 不 同 媒 介 的 作 用 下 被 活 化 (Springman & Harold, 1990; Min, 2002)。
例如protease的作用; 形式結構上的破壞 與擾亂物(conformational perturbants)如加 熱處理; sodium dodecyl sulfate與NaSCN;
重金屬如Au、Hg(II)和organomercurcials;
氧化物如NaOCl; disulfide化合物如氧化的 glutathione; sulfhydryl 烷 化 劑 如 N-ethylmaleimide (NEM)等等。 有許多的 報告表示MMPs的latent form也有自發性 的活化功能。 “Cysteine-switch” 的model 表現了在活化的區域,Cys-Zn2+ 之間的相 互 作 用 會 短 暫 的 分 離 (transient dissociation)。藉由此模式而讓Cys與SH reagents作用。Cys的變形模式接下來便會 防止Cys與Zn2+再相互作用,因此,SH基 便 永 久 與Zn2+分 開 , 而proMMPs 就 會 autolytically表現propeptide。ProMMPs會藉
由 上 述 的 物 理 或 化 學 作 用 而 產 生
“cystein-switch”形式。然而,經由APMA 或 其 它SH 的 作 用 , 化 學 性 的 改 變 在 propeptide中的Cys75 而破壞Cys與Zn2+之 間 的 協 調 , 卻 不 足 以 活 化proMMP-3 。 proMMPs 經由APMA與SH作用而被活化 的精確機轉,目前並不是十分清楚。僅管 如此,cysteine-switch的前提闡釋了Cys與 Zn2+之間的破壞,對於proMMPs的活化是 必須的條件(Woessner, 1999)。
除 了 MMP-7 (matrilysin) 、 MMP-26(endometase/matrilysin-2) 與 MMP-23 , 所 有 的 MMPs 都 有 hemopexin/vitronectin-like domain 並 以 hinge 或 link region 與催化 domain 連接。
Hemopexin domain 會影響 TIMP 與一些特 定基質的binding、也會影響 membrane 的 活化與一些proteolytic 的活性。(Sternlicht, 2001) 。舉例來說,hemopexin-like domain 對 MMP-9 在 細 胞 表 面 的 活 化 與 collagenase 的 活 性 表 現 都 是 必 要 的 (Mengshol, 2002)。
因為 Gelatinases A 與 B (MMP-2 與 MMP-9) 的 催 化 區 域 中 含 有 三 個 head-to-tail cystein-rich repeats,而特別被 區別出來。這些 inserts 類似 fibronection 的collagen-binding type II repeats,而需要 與collagen 和 elastin 結合與切除(cleavage) (Murphy.1994; Shipley 1996)。除此之外,
MMP-9 為 MMPs sequence 中最常且複雜 者 (92 kDa gelatinase B),在 hinge region 的底部含有獨特的 type V collagen-like inserts,而此 insert 的重要性與功能仍然未 知 。 Membrane-type 的 MT-MMPs 有 single-pass transmembrane domain 與短的 cytoplasmic C-terminal tail (如 MMPs 14、
15、16 與 24) 或是 C-terminal hydrophobic 的 區 域 (Itoh. 1999; Kojima, 2000),以 glycophosphatidyl inositol (GPI) membrane-anchoring signal 表 現 ( 如 MMP-17 和 MMP-25)。這些 MMPs 的 regions 在一些重要的 proteolytic events,
到專一性的細胞表面區域的 localizations 上都扮演了不可缺的角色。
MMPs 的 其 他 特 徵 包 括 : furin-susceptible site transmembrane
domain 、 cytoplasmic tail 、
glycophosphatidyl inositol-anchoring domain、 cysteine/proline 及 interleukin-1
receptor etc。
MMPs 最初被定義為一群酵素可被 金屬螯合劑與內生性抑制劑所抑制,而為 活化型可受有機汞所活化;其所能催化水 解的細胞外基質蛋白至少一種以上,例如 collagen、elastin、laminin 等。因此其對 於組織 modeling、repairing 與 destroying,
皆 扮 演 了 重 要 的 角 色(Woessner et al., 1991)。
MMPs 在生理情況下之內生性抑制劑 為 TIMPs (tissue inhibitors of metalloprotinases),TIMPs 屬於低分子量 (20~29kDa) 的蛋白質,主要分泌在細胞表 面而與 membrane-bound 的蛋白質相關。
它能夠專一性的抑制活化型態的 MMPs
(Baker, 2002)。目前已有四種 TIMPs 被發 現(如 TIMP-2、TIMP-3 與 TIMP-4 能與 membrane 形式的 MMP-14 結合),不同的 TIMPs 對於不同的 MMPs 抑制的能力也有 差別。每種 TIMP 對於 MMP 的親和力也 有 所 不 同(Brew, 2000) , 如 TIMP-1 為 MMP-9 的主要抑制劑。TIMPs 對於 MMPs 活性的抑制是為一種相當重要的嚴密調控 機制(Curran, 1999)。
在許多病理與生理情況下,MMPs 亦 扮 演 了 重 要 的 角 色 。 包 括 了 embryonic tissue morphogenesis 、 tissue repair 、 angiogenesis atherosclerosis 、 arthritis 、 gastric ulcer、tumor invasion 與 metastasis 等。MMPs 亦與細胞外基質 ECM 分解破 壞(extracellular matrix)的 components 息息 相關。
一般而言,不同類型之細胞有其特殊 分泌之MMPs。如 HUVEC (人類臍帶代靜 脈內皮細胞) ,主要是以 constitutive 分泌 的MMP-2 為主(De Lorenzo et al., 2000)。
而活化的人類單核球細胞或巨噬細胞,其 分 泌 主 要 是 以 inducible 的 MMP-9 為 主,藉由破壞base membrane 使單核球滲 出 至 組 織 (extravasation) (Lepidi et al., 2001)。
2-1-2. MMPs and Disease
結 締 組 織 的 完 整 性 依 賴 於
extracellular matrix 在分解(degradation)與 修復(repair)之間的平衡性。而蛋白質水解 酵素在 extracellular matrix 的 degradation 上扮演了rate-limiting 的一員。目前 MMPs 無疑已被認為在 degrade collagen 與其他 extracellar matrix 大分子上扮演了相當重 要的角色(Loftus, 2002)。通常 MMPs 會以 低濃度的表現量在正常成人體內,而當正 常或病理方面的 remodeling process 產生 時,MMPs 的量會則會提升。MMPs 與傷 口 癒 合 的 過 程 息 息 相 關 (Wysocki, 1993),但在與 extracellular matrix 的破壞 所造成的其它相關疾病上,則有更多的文 獻與研究討論。第一個與MMPs 相關的疾 病就是rheumatoid arthritis (RA)。環繞於關 節周圍,由 collagen 所組成的軟骨、骨頭 和 腱 被 MMPs- 特 別 是 collagenase 所 destructed (Elliott, 2001; Brinckerhoff, 2002)。因此,在 arthritic diseases 方面,
MMPs 的抑制劑被認為是一種重要的治療 目標 (Mengshol, 2002)。另外 MMPs 還參 與了cell growth、invasion、angiogenesis、
atheroma 與 cancer metastasis (Visse, 2003;
Chang, 2001; Egeblad, 2002)。MMPs 還可 以選擇性的 cleave matrix 的 protein,因 此,tumor-expressed-MMPs 便在 ECM 中 提供了空間讓tumor 細胞能通過 basement membrane 進行轉移。MMPs 在 metastasis 的 development 上因而扮演了重要的功能 (McCawley, 2000)。然而,一般而言,在早 期 tumor 形成期間,MMPs 的表現量會增 高。而在 invasive 與 metastatic 的 tumors
上,又有更高程度與更多的MMPs 表現。
由此可知,幾乎在所有 tumorgenesis 的階 段MMPs 都是需要的。在臨床方面,MMPs 的 表 現 程 度 可 作 為 許 多 tumor type 的 prognostic indicator (Stetler-Stevenson, 1996; Massova, , 1998)。因為 MMPs 參與 了 多 方 面 性 的 tumor 形 成 (tumor establishment 、 growth 、 angiogenesis 、 intravasation/extravasion 與 mastasis),抑制 MMPs 表現的研究在 anti-cancer 上也因而 被認為是重要的臨床implications (Bergers, 1999; Pupa, 2002)。
MMPs 在一些 lung diseases 方面 (Lim, 2003; Kelly, 2003),也有觀察到 MMP-9 的
量有增高的表現。研究發現,在asthma 的 病人中,inflammatory lung cells 因被刺激 而釋放出MMP-9,經由 cytokines 與 matrix bound growth factors 的調控,MMP-9 進而 modify 細胞的功能。 瞭解 MMP-9 在健康 與疾病上的角色因而變的相當重要,由於 broad spectrum 與專一性的抑制劑在不久 之後,將會從 bench knowledge 轉變成臨 床的試驗 (Atkinson, 2003)。
MMPs 被認為與早期粥狀動脈硬化板 塊(plaque)的發展有關。Plaque 的形成會 引起細胞的 migration 與 proliferation 並逐 漸在extracellular matrix 增加。與 MMP-9 相 關 的 basement membrane 因 此 而 degrade,而促使 vascular smooth muscle cell 的 migration 與 proliferation 。 由 macrophages 所釋放出的 MMPs 所造成的 matrix degradation 被認為是構成在粥狀動 脈 硬 化 板 塊 的 不 穩 定 或 剝 離 (atherosclerotic plaque instability or rupture) 或 是 動 派 瘤(aneurysm) 形 成 的 關 鍵 。 在 atherosclerotic plaque 的過程中,纖維帽的 剝離 (fibrous cap rupture)是觸發 coronary thrombosis 和 種 種 急 性 coronary syndromes 的重要因素 (Dzau, 2002; Libby, 2002; Loftus, 2002) 。 而 這 兩 種 源 自 於 plaque rupture 的 病 理 特 徵 會 造 成 著 macrophages 密度的增加、collagen 的 loss 並伴隨著 tensile strength 的減少(Mann, 1996; Chase, 2002) 。 有 學 者 在 人 類 atherosclerotic plaque 與一些動物模式的研 究中都發現有MMP-1、MMP-3 與 MMP-9 的大量表現(Loftus, 2002; Henney, 1991;
Gails, 1994)。由於 MMPs 活性的大量表現 會造成支持性的結締組織 matrix 形成 local destruction ,而與 plaque 的剝離息息 相關(Zhang, 1999)。因此,藉由抑制 MMPs 由 macrophages 中釋放,便能增進 plaque 的穩定性。
在各種發炎疾病中,MMPs 於疾病過 程 中 扮 演 了 相 當 重 要 的 角 色 。 而 chemokine MCP-1 與細胞激素如細菌性內 毒素 (LPS) 均會刺激或誘導 MMPs 的產 生,進而造成結締組織分解,並促使各種 不同生理與病理的傷害。
2-1-3 MCP-1 對單核球之作用
MCP-1 是 一 種 單 核 球 細 胞 的 chemotactic 與活化因子(activating factor,
簡稱 MACF)。MCP-1 有 chemoattract 單 核球細胞的能力。除此之外,在 in vitro 下 ,MCP-1 還 可 以 活 化 單 核 球 細 胞 (Mantovani, 1999),在癌症細胞株使其呈 cytostatic 狀態,使其細胞質的 free calcium 增加,進而造成並釋放出單核球細胞超氧 化 的 陰 離 子 並 且 釋 放 單 核 球 細 胞 lysosomal 酵素。目前的文獻植指出,在 人類單核球細胞中,MCP-1 有調控黏附因 子(adhesion molecules)和細胞激素產生表 現 的 能 力 。 並 且 , MCP-1 還 有 chemoattracting、活化並誘導 histamine 從 嗜鹼性白血球 (basophils) 中釋放出來。
MCP-1 在生物上所扮演的角色,則和發炎 與非發炎疾病的狀態有關。其特徵包括了 白 血 球 在 受 傷 區 域 的 遷 移 與 積 聚 (McQuibban, 2002)、在 artherogenesis 的傷 害形成上扮演著重要的角色(Terkeltaub, 1998)、delayed hypersensitivity 與活化單 核球細胞中細胞激素的分泌(Reape, 1999) 等等反應。 MCP-1 還可與 MIP1-α、
RANTES 與 MIP-1β結合。CCL2 (MCP-1) 與CCL3 (MIP-1α)、CCL5 (RANTES) 屬 CC Chemokines , 會 激 發 單 核 球 細 胞 MMP-9 的釋放 (Robinson, 2002)。
而 MMP 對於單核球的製造誘發在
cancer 、asthma 與 rheumatoid arthritis (RA) 的病理角色都極為重要。先前的研究指 出,chemokines 會調控在 peripheral blood lymopocytes 的 MMP-9,而與發炎反應有 關的細胞激素如 TNF-α 則會在其他的細 胞誘發出MMP-9 ; 而 chemokine 要激發出 MMP-9 則需有 autocrine TNF-α 的存在。
Chemokine 會造成單核球細胞和 THP-1 細胞中 TNF-α mRNA 和蛋白質釋放的量 增加。然而,對於 chemokines 在單核球
細胞的 MMP 調控機制的瞭解則比較少。
在本實驗中,我們對觀察了 MCP-1 對
THP-1 細胞的刺激與作用。
2-1-4 LPS
脂 多 醣 體 Lipopolysaccharide (LPS) 或稱為細菌性內毒素(endotoxin)會誘發人 類 單 核 球 細 胞 去 表 現 許 多 pro-inflammatory mediators (Guha, 2001;
Netea, 2002),其中包括了 pro-coagulant 分 子tissue factor (TF) 與細胞激素腫瘤壞死 因子 TNF-α。LPS 是最早被發現的內毒 素,位於格蘭氏陰性菌 (gram-negative) 的 細胞壁上,當細菌溶解時,LPS 會被裂解 釋出 (Raetz, 1986)。對於格蘭氏陰性菌 (gram-negative bacteria) 的感染,LPS 可當 作是一種發炎反應的主要源頭,而toll-like receptor 4 (簡稱 TLR4) 則在此現象中有其 不可缺少地位(Kiechl, 2002)。人類的單核 球細胞對於LPS 的刺激尤其特別敏感,並 反應產生許多發炎性細胞激素。LPS 會與 LPS-binding protein (簡稱 LBP)在 plasma 中 結 合 , 並 傳 送 到 細 胞 表 面 的 CD14 receptor 。 接 下 來 LPS 便 會 被 轉 換 至 transmembrane 的 signaling toll-like receptor 4 與 LTR4 的附加蛋白 MD2 上 (Guha & Mackman, 2001)。LPS 對於單核 球 與 巨 噬 細 胞 是 一 種 強 有 力 的 激 活 劑 (O’Connell, 1998),對於 host 細胞有保護 性與傷害性的反應。藉由研究在單核球細
胞中 LPS 的分子機制與發炎性基因的調
控 ,也許能夠在許多系統性的發炎反應症 狀 的 治 療 上 , 找 到 新 的 方 法(Guha &
Mackman, 2001)。因 LPS 可刺激單核球並 釋放 TNF-α,間接誘發 MMP-9 之表現,
在本實驗中,我們對觀察了LPS 對 THP-1 細胞的刺激與作用。
在東方的國家中,藥用性的植物已被 廣泛的使用在傳統醫療方面。在大規模藥 材萃物及化學合成等成分藥物成分篩選實 驗 後 , 我 們 發 現 , Cinnamomum philippinense 所 萃 取 的 天 然 物 cinnamophilin 和 HDI II (Histone Deacetylase Inhibitor II) 對 MMP-9 有明顯 的抑制作用。
2-1-5 Cinnamophilin 的藥理活性
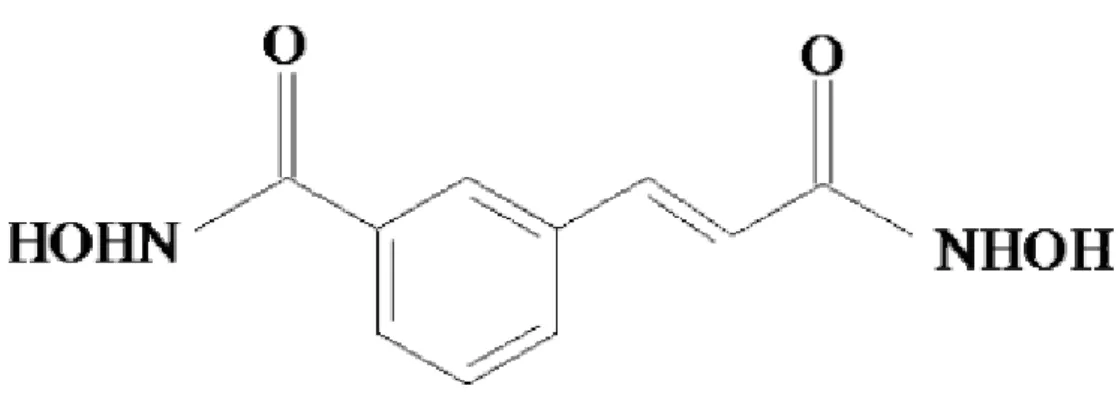
Cinnamophilin--((8R,8’)-4,4’-dihydeox y-3,3’-dimethoxy-7-oxo-8,8’-neo-lignan) , 簡 稱 MA-1 (Figure 1) , 是 一 種 由 Cinnamomum philippinense 中所萃取出來 的 天 然 物 , 已 被 確 定 有 free radical scavenging capacity (Hsiao, 2001),並且表 現出其特有的抗氧化與保護細胞免於氧化 的傷害,而在其他的in vitro model,其另 有抗脂質過氧化反應的能力。
過去的研究發現 (Cheng, 1995; Yu, 1994),在研究in vitro 人類platelet 、隔離 的大鼠主動脈、隔離的天竺鼠氣管與in vivo 小 鼠 與 天 竺 鼠 的 實 驗 中 , cinnamophilin 的藥理性作用除了有抑制 Thromboxane (TX) 合 成 、 阻 斷 voltage-dependent鈣離子通道的能力,還有 TXA2 antagonizing 的 作 用 ; cinnamophilin 是 為 一 種 選 擇 性 的 TXA2
的拮抗劑,並且能清楚的表現出抑制TX合 成的特徵。因此cinnamophilin被認為是一 種novel 的 TX 與TXA2雙重抑制劑 (dual inhibitor)。TXA2是一種由血液platelet 的 arachidonic acid (AA) 所 產 生 的 主 要 cyclo-oxygenase產物。TXA2 是一種非常 有效的platelet aggregator,vasoconstrictor 與bronchoconstrictor。 因此cinnamophilin 被認為在研究與治療與TXA2 相關的疾病 上可視為一項有用的工具。在骨骼肌肉 ischemic 的 model 中 (Cheng, 1995) , cinnamophilin 更 能 預 防 進 一 步 的 reperfusion 所造成的傷害。而在另外的一 項研究中,(Su, 1999) cinnamophilin 在大 鼠 心 臟 中 的 ischemia-reperfusion arrhythmia model表現出antiarrhythmic 的 能力經由抑制向外流的鉀current和向內流 的鈉與鈣current。
2-1-6 Histone Deacetylase Inhibitor HDI II 的藥理活性
Histone deacetylase inhibitor II
(m-Carboxycinnamic acid bis-hydroxamide),簡稱 HDI II (Figure 2),
則被認為是一種有效力的誘導劑,形成 transformed 細胞生長遏止並且造成末端 的 分 化 。 在 人 類 的 neuroblastoma 細胞 中,則會誘發CD95/CD95 ligand 的表現與 apoptosis。Histone 的 acetylation 在調控 transcription 上扮演著重要的角色。Histone acetylation 的 程 度 是 依 照 histoneacetyl transferases (HATs) 和 histone deacetylases (HDACs) 這兩種酵素 families 的活性而作 調控 (Archer & Kurdistani, 2003)。它是經 由控制 chromatin 的結構改變而進而調控 transcription 與 gene 表現。 Histone 是組 成nucleosome 的主要蛋白質成分,histone 與 DNA 相互作用而形成 nucleosomes。
Nucleosomes 包 含 了 緊 緊 依 附 在 DNA phosphate backbone 的 高 charged hypoacetylated histones,transcription 也因 而被抑制。 當 histone acetylation 時,這 種 高 charged 的 狀 態 會 被 中 和 , transcription factors 、 調 控 因 子 與 RNA polymerases 才 依 此 途 徑 去 產 生 開 放 的 DNA 結構,使 transcription 進行,相關的 gene 也才能產生 (Marks, 2001)。
一般而言, histone 的 actylation 是與 transcription 的 activation 有關; 而 histone deactylation 則 與 transcription 的 repression 相 關 。 是 否 histone 會 hyperacetylation 或 deacertylation 則被認 為是與其專一性的 target gene 有關。 如 hydeoxamic acid-based histone deacetylase inhibitos 會限制 tumor 細胞的生長,並會 選擇性的作用在gene 上,去調控大約 2 % tumor 細胞的 gene 表現 (Kramer, 2001)。
最近有文獻指出,有許多malignancies , 特 別 是 leukemias,與不正常的 histone deacetylases (HDACs) 或是 histone aceyl transferases 如 EP300 的突變有關。最近 有新的活體研究發現指出 HDAC 的抑制 劑在的治療cancer 上,與一般抗癌藥物相 較,是為更有效的治療目標去對抗難治療 的癌症(Batova, 2002)。Histone deacetylase inhibitors 在不同種類的 transformed 細胞 中,或是在tumor-bearing 的動物中,皆能 有效的引起生長停滯、分化或是 apoptotic 細 胞 死 亡 。 目 前 已 有 一 些 histone deacetylase inhibitors 如 FR901228 、 oxamflatin,hydroxamic acid-based histone deacetylase inhibitors MS-275 、 SAHA pyroxamide 、與 m-carboxy-cinnamic acid bishydroxamate (CBHA-HDI II 的 synonym) 等與 tumor 相關的動物實驗在 進行當中(Marks, 2001)。研究指出,CBHA 在 有 人 類 neuroblastoma 異 種 皮 移 植 的 nude 老 鼠 中 , 是 為 一 種 非 常 有 效 的 抗 tumor 生 長 的 抑 制 劑 (Coffey, 2001) 。 CBHA、SAHA 與 pyroxamide 皆能有效抑 制tumor 生長,當它們的有效劑量隨著動 物體重上升、或是各方面動物屍體的組織 研 究 而 增 加 時 , Histone deactylase inhibitors 只會造成微量,或甚至沒有藥毒
性的產生。Ailenberg 的研究發現,histone deacetylase inhibitor trichostatin A (TSA) 可以選則性的抑制 3T3 細胞中 gelatinase A 的表現與活化。Gelatinase A mRNA 和 gelatinolytic 的活性都呈現了 dose-related 減少變化。而其他 gelatinase A activating 的複合物如 MT1-MMP 和 TIMP-2 則沒 有變化 (Ailenberg, 2001) 。
2-1-7 基因表現之訊息傳遞路徑 Nuclear factor-κB (NF-κB)
Nuclear factor-κB (簡稱 NF-κB),屬於 transcription factors 家族中的一員。文獻 指出,NF-κB 與 Activator protein-1 (AP-1) 的binding 在人類和兔子的 smooth muscle cells 與 dermal fibroblasts 中 能 構 成 MMP-1、MMP-3 與 MMP-9 的 upregulation (Bond, 2001; Bond, 1999)。而在人類的 macrophages 和兔子的 foam cells 中(Chase, 2002),同樣觀察到了 NF-κB 與 MMP-1 和 MMP-3 的分泌有關。NF-κB 與許多和發 炎反應相關的genes 表現有關 (Feldmann, 2002),舉例來說,在 arteriosclerosis lesion 形成的初期(initial phase),許多在內皮細 胞中表現的proinflammatory genes (如 LPS (O’Connell, 1998)、TNF-α、IL-1 等)與它 們proinflammatory receptors 之間的作用都 是 NF-κB dependent。 同樣在接下來的 lesion progressing phase 中 , 有 白 血 球 monocyte 的 infiltrating 作用,將導致這些 與發炎反應相關的 genes 表現。而這些 genes 皆是 NF-κB dependent 的(Martin, 2000)。NF-κB 是由 Rel A (p65)、RelB、
c-Rel、NF-κ B(p50) 與 NF-κB2 (p52) 所組 成 。 後 兩 者 分 別 是 由 inactive precursor molecule p105 與 p100 所合成。NF-κB 的 家 族 成 員 還 包 括 了 inhibitory subunits:
IκBα、IκBβ與IκBε。NF-κB subunits 形成 了 homo- 與 heterodimers , 而 p65/p50 heterodimer 是最 predominant 的。NF-κB 經由外界的一些 stimuli 與 inflammaroty factors (如 TNF- α、IL-1、LPS、advanced glycation end products (AGEs) 、 hyperglycemia、plate-activating factor、sheer stress 等) 刺激之後(Pahl, 1999),IKKs 會 phosphrylate IκB α,並在 polyubiquitinated
與 proteosme 的 complex system 作用下被 degrade 掉。IκB α的 degradation 進而使 NF-κ B translocate 到 nucleus 中使一些特 定的基因進行active transcription。
在進來幾年,許多的研究對 NF-κB pathway 在治療上的調節作用(modulation) 有有逐漸增加的興趣。由實驗得知,NF-κ B 在治療方面的調控機制包括了基因上的 操 作 ( 如 Decoy oligodeoxynucleotides (ODN)、IκB mutation、Iκ B gene transfer) 到藥理上NF- κB pathway 中特定步驟的抑 制劑如 NSAIDs、proteosine inhibitors、
glucocorticoids 等(Zingarelli,2003)。
2-1-8 Mitogen-activated protein kinase (MAPKs)
Mitogen-activated protein kinase (MAPK) 構成了由細胞表面到核內的主 要訊息傳遞路徑。這些訊息的傳遞控制了 相當複雜的訊息作用程序,並在許多方面 調控了mammalian 細胞的生理作用。其中 包括了胚胎發育、細胞的分化、增生與死 亡 (Ip, 1998; Chuang, 2000)。另外在短期 的變化方面,MAPKs 則與體內平衡與急 性賀爾蒙的反應有關 (English, 2002)。
MAPKs 屬於 serine-threonine kinase 酵素,
主要的mammalian MAPKs 為 extracelullar signal-regulated kinase (ERK) 、 c-JUN N-terminal kinase (JNK) 與 p38 kinase。
MAPKs 會經由 Thr 與 Tyr 的區域磷酸化而 產生活性。一種含MAPK kinase kinase 的 protein kinase 能夠磷酸化和活化一種或多 種的MAPK kinase (MKK),也因此磷酸化
與活化了下游的每一個 MAPK ,進而
translocate 進入細胞核,促使轉錄因子 AP-1 的活化與基因的表現。
一般而言,一些growth factors 會活化 ERK 的訊息傳遞路徑,在細胞的增生則扮 演了決定性的角色 (Hills, 1995)。相反 的,基因毒性物與細胞激素會刺激JNK 與 p38 的訊息傳遞路徑,並傳遞 stress 的反 應、生長停頓與apoptosis。MAPK 在生理 上的所表現的複雜性在於對於不同種類的 細胞,它能部分性的依其活性的持續期間 作用去調控細胞的基因表現。如ERK 短暫 性的活化會導致細胞增生,而持續性的作 用 則 傳 達 了 生 長 停 止 或 分 化 的 訊 息
(Alblas &Yen, 1998)。相對而言,JNK 與 p38 短暫性的誘發則提供了細胞生存的訊 息。然而,持續性的活化卻與apoptosis 的 誘發有關 (Guo & Roulston, 1998)。有研究 指出,LPS 誘發單核球細胞 MMP-1 的產 生是經由 ERK1/2 與 p38 的調控,而對於 單 核 球 細 胞 與 初 級 星 細 胞 (primary astrocytes) MMP-9 的 產 生 則 主 要 經 由 ERK1/2 的 訊 息 傳 遞 路 徑 (Lai & Lee, 2003)。
2-2 實驗目的
在 我 們 的 實 驗 中 , 以 人 類 單 核 球 細 胞 THP-1 為 實 驗 細 胞 , 藉 以 了 解 cinnamophilin 與 HDI-II 等 藥 物 對 MCP-1 與 LPS 所引出或誘發之 MMP-9 蛋 白 質 活 性 的 表 現 與 影 響 。 我 們 以 Zymography 、Western Blot 與 RT-PCR 等 實驗試著了解與觀察 MMP-9 蛋白質與 mRNA 的表現,並以 MTT Assay 觀察細胞 存活率的情形。而之後我們探討了之中的 藥理作用和機轉等問題,試著進一步的瞭 解cinnamophilin 與 HDI II 在細胞訊息傳 遞路徑如MAPK 或 NF-κB 等機制的影響 程度。另外藉由細胞的 migration assay,
我 們 觀 察 了 MCP-1 所 induce 的 細 胞 migration 情形。期許能夠藉由 YC-1、
cinnamophilin 與 HDI II 類 似 作 用 藥 valproic acid 等藥物之研究,對與 MMP 有關的病理反應如出血後再甦醒性傷害,
能進一步的評估與瞭解其治療的可能性。
三、結果與討論 3-1 結果
3-1-1. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II)對人類單 核球細胞 (THP-1)以 LPS 或 MCP-1 所誘 發的MMP-9 酵素活性
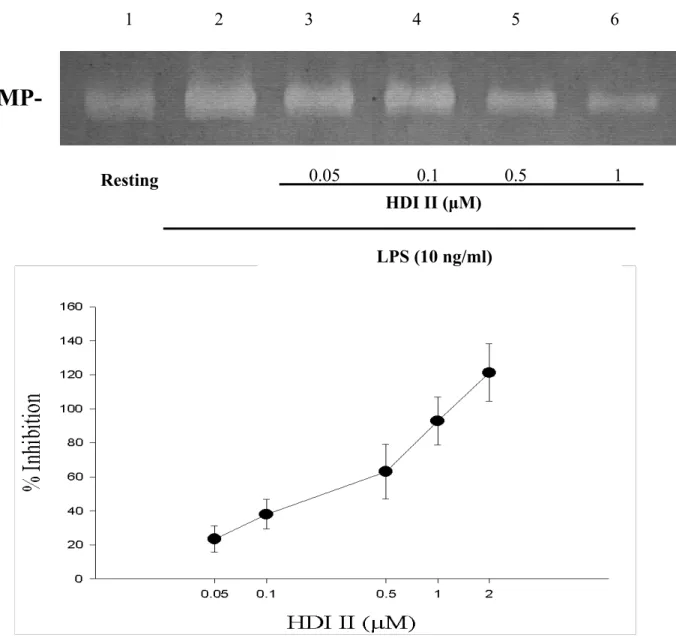
由 之 前 已 建 立 之 刺 激 物 質 誘 發 MMP-9 活 性 的 表 現 條 件 的 gelatine zymography之方法,投與不同濃度之藥物 以 觀 察 藥 物 對LPS 與 MCP-1 所 誘 發 的 MMP-9 在THP-1 細胞中的影響。我們之 前在THP-1 細胞中分別給予不同濃度的 MCP-1 與LPS處理 24 小時後,以gelatine zymography的方法觀察發現以細胞濃度為 1×106/ml時,240 ng/ml 的MCP-1 與 10
ng/ml的LPS能誘發MMP-9 (非MMP-2) 在 THP-1 細胞中明顯的表現,之後,依此條 件再加以cinnamophilin 或HDI II等藥物。
由gelatine zymography的方法發現,隨著 cinnamophilin濃度(1 µM、5 µM、10 µM、
20 µM、50µM),或是HDI II (0.05 µM、0.1 µM、0.5 µM、1 µM、2 µM) 濃度的增高,
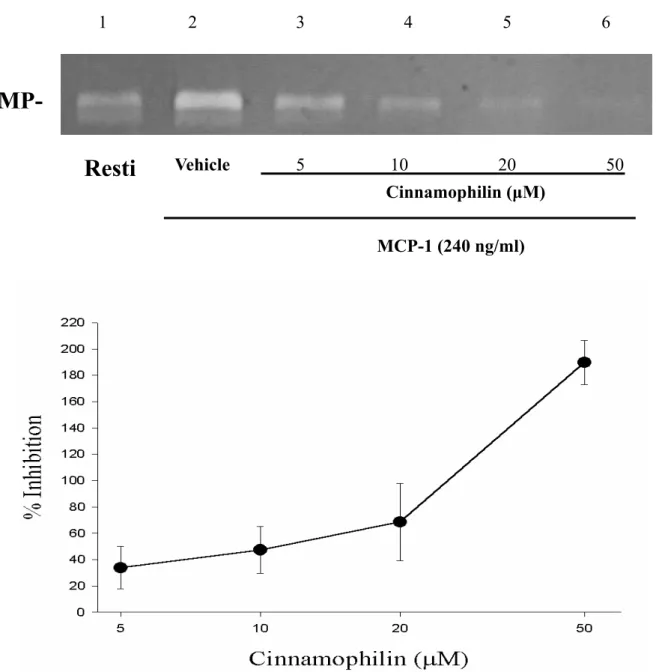
MMP-9 活性的表現呈現逐漸減少而被抑 制 的 現 象 (concentration-dependent inhibition)。Cinnamophilin對MCP-1 刺激作 用的抑制百分比(In %)分別為 33.8 ±16.1
% (5 µM),47.3 ±17.7 % (10 µM),68.5 ± 29.9 % (20 µM),189.9±16.8 % (50 µM),
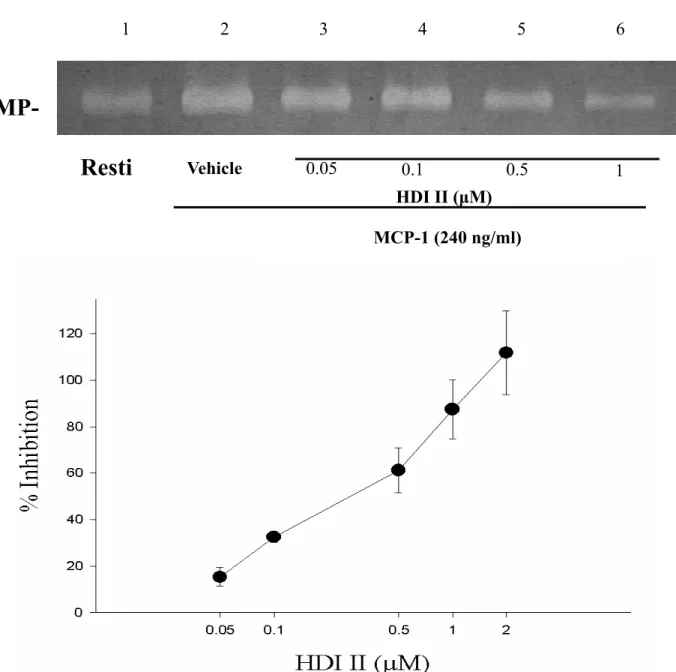
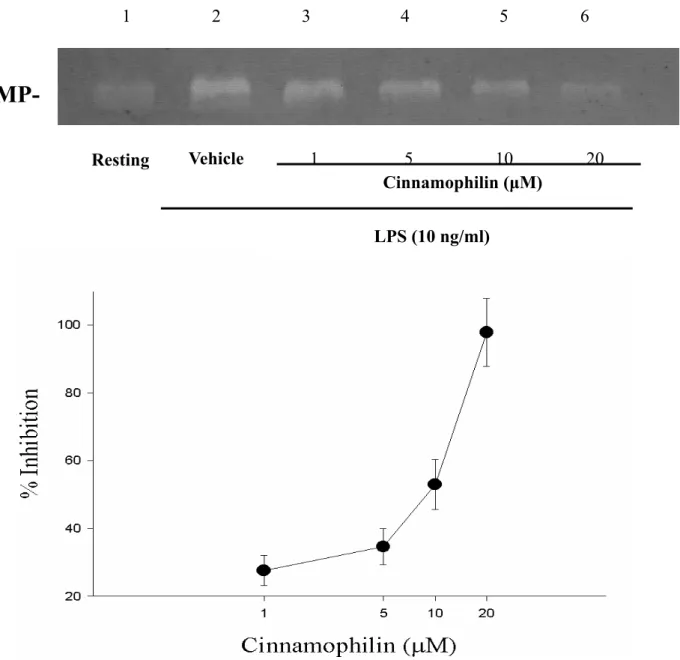
其抑制 50 % 反應濃度(IC50)為 11.6 ±3.0 µM (n = 3,Figure 3)。另外,cinnamophilin 對LPS刺激作用的抑制百分比(In %) 分別 為27.6 ±4.45 % (1 µM),42.2 ±11.5 % (5 µM),60.5 ±14.7 % (10 µM),92.9 ±9.7 % (20 µM),其抑制 50 % 反應濃度(IC50)為 8.1 ±1.5 µM (n=3-4,Figure 5)。而HDI II 對MCP-1 刺激作用的抑制百分比(In %)分 別為15.4 ±3.9 % (0.05 µM),32.5 ±2.4 % (0.1 µM),61.2 ±9.4 % (0.5 µM),87.5 ± 12.7 % (1 µM),其抑制 50 % 反應濃度 (IC50)為 0.37 ±0.04 µM (n = 3-5,Figure 4)。另一方面,HDI II對LPS刺激作用的抑 制百分比(In %) 分別為 23.4 ±7.9 % (0.05 M),38.0±8.7 % (0.1 M),63.1 ±15.9 % (0.5 µM),92.8 ±14.1 % (1 µM),其抑制 50 % 反應濃度(IC50)為 0.43 ±0.16 µM (n
= 3-4,Figure 6)。無論是LPS或是MCP-1 在THP-1 細胞中所誘發之MMP-9 皆能被 cinnamophilin 或HDI II等藥物隨濃度上升 而 呈 現 活 性 減 少 之 反 應 , 此 為 一 種 concentration-dependent inhibition。
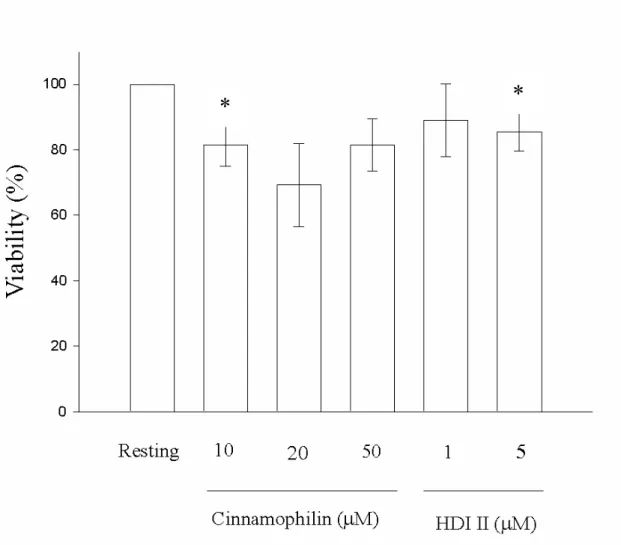
3-1-2.探討藥物對細胞存活率的影響 為了證明cinnamophilin與HDI II等藥物 對於THP-1 細胞中MMP-9 活性的抑制並 非引起細胞減少或死亡,而導致MMP-9 的活性減少,我們使用了MTT assay的方 式。將1×106/ml的細胞種在 24 well中,以 cinnamophilin (10 µM、20 µM、50 µM)或 HDI II (1 µM、5 µM)處理 22 小時候,再 加入MTT試劑處理細胞 2 小時。存活細胞 的粒腺體內的酵素會將MTT試劑還原成
formazan紫色結晶。之後再加入DMSO溶 解結晶以550 nm測其吸光值,細胞之存活 率 與resting 組 比 較 之 後 以 百 分 比 表 示 (Figure 7)。我們發現cinnamophilin 在 20 µM (69.3 ±12.7 %) 及 50 µM (81.5 ±8.0 %) 時對細胞的存活率有些許的影響,但在統 計上並不具又意義。HDI II在 1 µM (89 ± 11.2 %) 時對細胞的存活率沒有影響,但 在濃度增至5 M (85 ±5.9 %) 時,約影 響了15 % 的細胞存活。
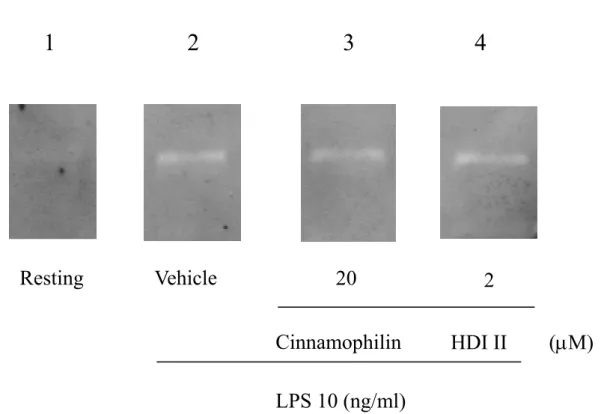
3-1-3.探討 Cinnamophilin 與 HDI II 對 MMP-9 酵素本生活性之影響
在直接將cinnamophilin 與 HDI II 加入 reacting buffer 中 培 養 , 再 依 照 gelatin zymography 步驟染色所得知結果 (Figure 8),我們觀察到藥物的投與並不會對 LPS 刺激下的lytic bands 產生抑制的作用,表 示出藥物對於 MMP-9 本生的酵素活性不 是直接的抑制作用,而是經由抑制MMP-9 蛋白質的表現而與細胞內的轉錄作用相 關,或藉由影響訊息傳遞路徑如NF-κB 或 MAPK 等訊息傳遞而對 MMP-9 產生影響 與抑制作用。
3-1-4. 探 討 Cinnamophilin 或 Histone deactylase inhibitor II (HDI II)對 THP-1
細胞以LPS 所誘發的 MMP-9 蛋白質表現
之作用
由之前的實驗結果得知,cinnamophilin 或 Histone deactylase inhibitor II (HDI II) 能抑制在人類單核球細胞 (THP-1)中,
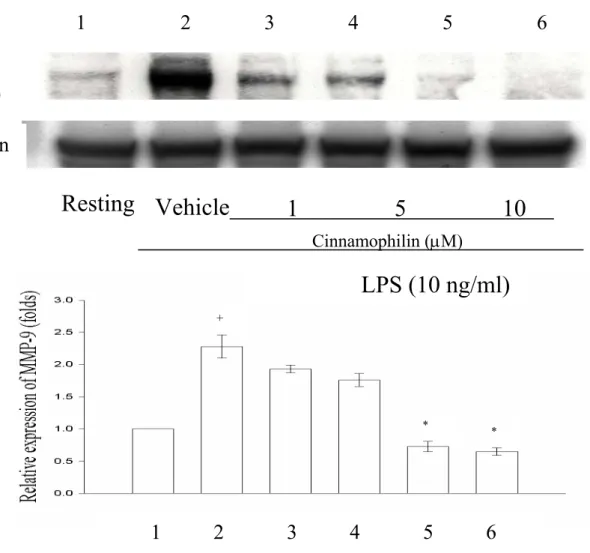
LPS 或是 MCP-1 所誘發的 MMP-9 酵素活 性,並由MTT assay 證明了 MMP-9 的活 性抑制並非源由藥物所影起之細胞毒性作 用。 欲瞭解 Cinnamophilin 與 HDI II 對 MMP-9 的抑制是否藉由減少其蛋白質的 表現,我們使用了 Western Blot 來分析 cinnamophilin 與 HDI II 對 MMP-9 蛋白質 的表現的影響。我們將THP-1 細胞處理並 培養24 後,取得其萃取物(cell lysate)。由 Figure 9 與 Figure 10 可觀察到,在有外在 LPS 的刺激之下,細胞萃取物有大量被 LPS 誘發出來的 MMP-9 蛋白質表現(lane 1,resting)。反之,在沒有外在刺激劑時 (lane 2),細胞萃取物只有非常微量 MMP-9 蛋 白 質 的 表 現 。 在 投 與 不 同 濃 度 的
cinnamophilin (1 µM、5 µM、10 µM、20 µM),或是 HDI II (0.1 µM、0.5 µM、1 µM、
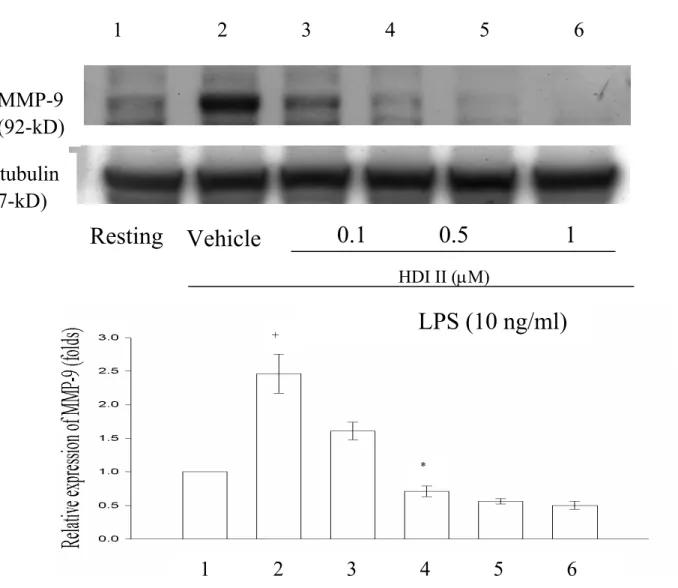
2 µM) 時,由 lane 3 至 lane 6 可以觀察到 MMP-9 蛋白質的表現隨藥物濃度逐漸增 高,而呈現逐漸減少的情況。Figure 9- cinnamophilin 在 LPS 刺激作用下,relative MMP-9 的表現為: 1 (Resting),2.28 ±0.18 (Vehicle),1.93 ±0.06 (1 µM),1.76 ±0.1 (5 µM),0.73 ±0.08 (10 µM),0.65 ±0.06 (20 µM)。Cinnamophilin 對 LPS 刺激作用的抑 制百分比(In %) 分別為 26.3 ±6.1 % (1 µM),40.3 ±5.6 % (5 µM),96.6 ±25.1 % (10 µM),104.9 ±23.1 % (20 µM)。Figure 10 顯示 HDI II 在 LPS 刺激作用下,relative MMP-9 的表現為: 1 (Resting),2.46 ±0.32 (Vehicle),1.61 ±0.13 (0.1 µM),0.71 ±0.08 (0.5 µM),0.56 ±0.04 (1 µM),0.5 ±0.06 (2 µM)。對 LPS 刺激作用的抑制百分比(In %) 分別為39 ±10.2 % (0.1 µM),70.2 ±5.4 % (0.5 µM),75.6 ±7.2 % (1 µM),92 ±21.3 % (2 µM)。Cinnamophilin 與 HDI II 這兩種 藥物接皆能對在THP-1 細胞中,LPS 所誘 發 之 MMP-9 的 蛋 白 質 產 生 concentration-dependent 的抑制作用。
3-1-5. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II) 對 THP-1 細胞以 LPS 所誘發釋放 TIMP-1 蛋白質 之作用
在組織中,TIMP-1 對於MMPs是最主 要的內生性調控者與活性抑制劑 (Nagase, 1998),目前已知的TIMPs (TIMP-1 至 4) 有四種。每種TIMPs對於MMPs的親和力與 抑制力都不盡相同,TIMP-1 便為MMP-9 的主要內生性抑制劑 (Brew, 2000)。為了 觀察與瞭解MMP-9 表現量的減少是否與 其內生性的抑制因子TIMP-1 有關,我們取 實驗所得之上清液 (supernatant) 並使用 了Human TIMP-1 ELISA system 來測其 TIMP-1 的變化。由Figure 11 與 12 中可以 觀 察 到 , 在 沒 有 外 界 的 刺 激 之 下 (Resting),以每 106的細胞為單位,便可偵 測到些許程度TIMP-1 的蛋白質表現(分別 為84.4 ±14.8 與 77.8 ±12.9 pg/106 cells)。
在經LPS刺激後,TIMP-1 的量由Figure 11 可觀察到有一些上升的情形(109.5 ±21.7
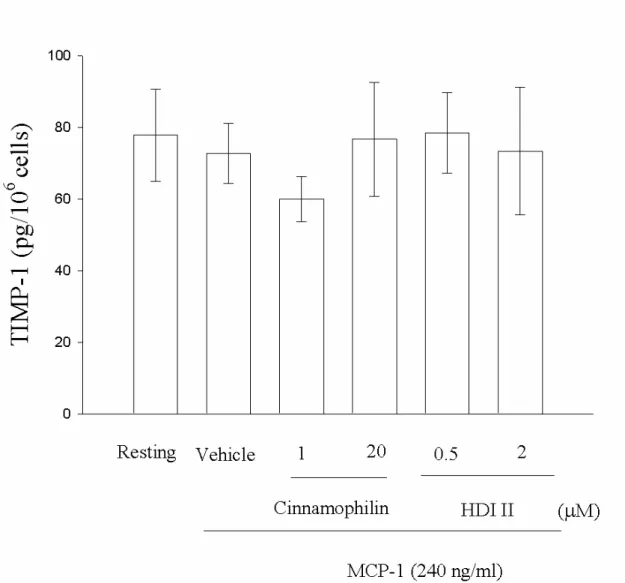
pg/106 cells),但在統計上並無明顯的差 異。而在經MCP-1 刺激的組別中(Figure 12),TIMP-1 雖有些微的下降(72.2±8.3 pg/106 cells),但與Resting幾乎沒有明顯的 差 異 。 在 分 別 加 以 藥 物 處 理 後 (cinnamophilin 1 µM、20 µM; HDI II 0.5 µM、2 µM),藥物對於LPS刺激下TIMP-1 蛋 白 質 的 表 現 皆 無 明 顯 的 影 響 : cinnamophilin 1 µM (105.1 ± 9.2 pg/106 cells)、20 µM (102.5 ±12.6 pg/106 cells) ; HDI II 0.5 µM (105.7 ±2.4 pg/106 cells)、2 µM (97.5 ±7.6 pg/106 cells)。而在MCP-1 的刺激之下,同樣投與相同條件的藥物濃 度(cinnamophilin 1 µM、20 µM; HDI II 0.5 µM、2 µM),我們可以觀察到較低濃度(1 µM) 的 cinnamophilin 似 乎 會 減 少 些 許 TIMP-1 蛋 白 的 表 現 (59.9 ± 6.4 pg/106 cells),但在統計上並無意義。而在有 20 µM cinnamophilin (76.7 ±15.9 pg/106 cells) 與 HDI II 0.5 µM (78.5 ±11.3 pg/106 cells)、2 M (73.4 ±17.9 pg/106 cells) 存在時,對 於MCP-1 刺激下TIMP-1 蛋白質的表現在 統計上仍無明顯的影響。本實驗所有的數 值皆為三次獨立實驗後的平均值。
3-1-6. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II)在單核球 細胞 (THP-1)以 LPS 所誘發的 MMP-9 mRNA 之影響
細胞內mRNA 的表現是經由 RNA 萃取 之 後 , 以 reverse transcriptase reaction (RT-PCR)的方法完成。使用 MMP-9 與 GAPDH 的專一性 primers 進行 MMP-9 與 GAPDH 之 cDNA 產物,再以 1 % 的 agarose gel 的電泳分析。House-keeping gene,GAPDH 在此為 internal control。由 Figure 13 與 14 中,我們可觀察到 MMP-9 mRNA 程度經由 LPS 的誘發而上升(lane 2)。而在給予藥物之後(lane 3 與 lane 4),
mRNA 量的表現就會被藥物所抑制,並表 現出 concentration-dependent inhibition 的 現象。 Figure 13 顯示 cinnamophilin 在 LPS 刺激作用下,relative MMP-9 mRNA 的表 現為: 1 (Resting),1.55 ±0.11 (Vehicle),
1.36 ±0.17 (5 µM),1.07 ±0.06 (20 µM),
而cinnamophilin 對 LPS 刺激作用的抑制百
分比(In %) 分別為 31.8 ±6.1 % (5 µM),
63.8 ±4.2 % (20 µM)。Figure 14: HDI II 在 LPS 刺激作用下,relative MMP-9 mRNA 的 表 現 為 : 1 (Resting) , 1.50 ± 0.15 (Vehicle),1.21 ±0.07 (0.5 µM),0.89 ±0.1 (2 µM); HDI II 對 LPS 刺激作用的抑制百 分比(In %)為 34.7 ±10.8 % (0.5 µM),當藥 物濃度在與2 µM (lane 3)時,到達了近乎 100 % (97.6 ±24.1 %) 的 mRNA 抑制作 用。
3-1-7. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II)對 THP-1 細胞以LPS 誘發 IκB-α之降解作用
單 核 球 細 胞(monocyte) 的 吞 噬 細 胞 (phagocytes)在免疫與發炎相關的反應作 用上扮演了不可缺的重要角色。基於細菌 的脂多醣體 lipopolysaccharide (LPS)會誘 導 單 核 球 細 胞 經 由 活 化 轉 錄 因 子 NF-κB/Rel,而進而表現許多與發炎反應有 關 的 基 因 (O’Connell, 1998; Martin, 2000),我們利用了觀察 NF-κB 的抑制因 子(inhibitory subunit),IκB-α,在 Western blot 中蛋白質量的表現情形,來瞭解藥物 對於 IκB-α磷酸化的分解作用與在 NF-κB 的訊息路徑上所扮演的任務。我們首先觀 察LPS 在不同之時間點對 IκB-α蛋白質表 現所造成之引響。由 Figure 15 中,我們 觀察到當由 LPS 刺激 90 分鐘時所取得之 細胞lysate (lane 4),IκB-α的表現呈現明顯 的降解作用,隨後又回到了basal level 基 準值(lane 5,120 min ; lane 6,180 min)。
而在其他時間點的 IκB-α表現(lane 2,30 min ; lane 3,60 min),則與 Resting 組無 異。我們接下來便以 90 分鐘為 LPS 刺激 的 基 準 , 分 別 投 與 不 同 濃 度 之 藥 物 (Cinnamophilin 10 µM、20 µM 與 HDI II 2 µM、5 µM)。若將 Resting 設為 100 % 的 basal level (Figure 16),在 LPS 刺激之後,
relative IκB-α的蛋白質含量降低為 80.3 ± 7.8 %,我們觀察到隨著 cinnamophilin 濃 度的上升,IκB-α也逐漸回升到與 Resting 相當的含量: 98.3 ±4 .1 % (10 µM),118.2
±9.2 % (20 µM)。而此現象在 Figure 17 中,在LPS 刺激之後,relative IκB-α的蛋 白質含量降低為 67.4 ±1.2 %,但在給予
HDI II 之後,relative IκB-α 的含量反因 藥物濃度的增加而呈現減少的狀況: lane 3, 68.4 ±4.3 % (2 µM); lane 4, 53.1 ±2.3 % (20 µM)。根據由 Figure 16 所關察到的情 形,我們可以推斷cinnamophilin 可能會經 由抑制IκB-α蛋白質的 degradation 而進一 步的減少NF-κB translocate 至細胞核內的 作用。也因此,一些與發炎反應相關的基
因也無法經由 NF-κB 的活化而進行運
作。LPS 誘導出來的 MMP-9 也因此被抑 制而無法表現。而HDI II 對抑制 IκB-α蛋 白質的degradation 作用並不明顯,則需進 一步的加以探討。
3-1-8. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II)對 THP-1 細胞以LPS 誘發 p65 之影響
在使用TransAMTM NF-κB p65 NF- κB p50 Transcription Factor Assay kits之前,我 們先使用Western blot的方式來觀察採用 NE-PERTM nuclear and cytoplasmic extraction reagents kit抽出之THP-1 核內物 質中,p65 的蛋白質是否會受到藥物的抑 制 。 由 Figure 18 中 可 以 觀 察 到 cinnamophilin (lane 3: 20 µM) 能抑制在人 類單核球細胞 (THP-1) 核內,LPS所誘發 的p65 蛋白質表現 (lane 2, Vehicle),但 Histone deactylase inhibitor II (HDI II) 則 無此抑制作用(lane 4: 2 µM),反而呈現出 加 強 p65 蛋 白 質 的 表 現 。 2,2,5,7,8-Pentamethyl-6-hydroxychromane (PMC) 在 10 µΜ 時,對於NF-κB 是一種 非常強效的抑制劑 (Suzuki, 1993),因此我 們 使 用PMC (lane 5: 10 µΜ) 來 當 作 control。可發現PMC對 LPS所誘發的p65 蛋白質表現有明顯的抑制作用。而HDI II 會加強p65 蛋白質的表現可能是由於正向 回饋的的作用。
3-1-9. 探 討 Cinnamophilin 與 Histone deactylase inhibitor II (HDI II)對 THP-1 以 LPS 誘發釋放 ERK、JNK 等 MAPKs 之活化作用
Mitogen-activated protein kinase (MAPK) 構成了由細胞表面到核內的主 要訊息傳遞路徑。由於主要的 mammalian MAPKs 為 extracelullar signal-regulated
kinase (ERK) 、 c-JUN N-terminal kinase (JNK),而 ERK 與 JNK 易受細胞激素的影 響而被活化。我們利用了觀察JNK 與 ERK 在 Western blot 中蛋白質量的表現情形,
來瞭解LPS 是否經由 JNK 或 ERK 等訊息 路徑而誘發 MMP-9 的表現,並觀察藥物
在其中所扮演的角色。我們首先觀察 LPS
在不同之時間點對 JNK 或 ERK 蛋白質表 現所造成之引響,發現在15 分鐘時,JNK 蛋白質的表現最為明顯,而ERK 則無顯著 的變化 (data not shown)。之後我們便以 15 分鐘為LPS 刺激之基準,由 Figure 19 中,
我們觀察到在分別投與不同濃度之藥物 cinnamophilin 20 M (lane 3) 與 HDI II 2 M (lane 4) 之後,relative JNK 的表現為:
1 (Resting),1.63 ±0.12 (Vehicle),1.22 ± 0.02 (cinnamophilin 20 µΜ),0.89 ±0.02 (HDI II 2 µΜ)。Cinnamophilin 對於 LPS 刺 激作用的抑制百分比(In %)為 58.7 ±13.6
%,而 HDI II 對 LPS 刺激作用的抑制百分 比(In %)則為 100.6 ±9.4 % 。由於在投與 藥物之後,ERK 蛋白質的表現作用沒有明 顯的改變(Figure 20),因此,LPS 誘導出來 的 MMP-9 可能經由 JNK 的訊息傳遞路 徑。我們可以推斷HDI II 可能會經由抑制 JNK蛋白質的表現再進一步的抑制轉錄因 子 AP-1 的活化與 MMP-9 的表現,而 cinnamophilin 對於 JNK 蛋白質的抑制作 用則不如HDI II 顯著。
3-1-10.探討藥物對 MCP-1 誘導人類單核 球細胞 (THP-1) 進行 migration 之影響
當有病理性發炎反應產生時,單核球細 胞 (monocyte) 會 遷 移 到 vascular subendothelium 中,而這種 migration 作用 在atherosclerosis 與 rheumatoid arthritis 中 皆扮演了非常重要的角色(Falcone, 2001;
Kintscher, 2000; Zhu, 1999)。穿越內皮細胞 的的單核球 migration 作用是一種多階段 性的機轉,最先會經由內皮細胞上細胞激 素所調控的黏附因子(adhesion molecules) 與循環中的單核球(circulating monocyte) 的黏附作用。而後,黏附的單核球便會朝 向特定的chemokines (chemotaxis) 移動穿 越 毗 連 的 內 皮 細 胞 , 再 經 過 basement membrane 到受傷發炎的組織中,此時便需
要 MMPs 的作用去分解細胞外的基質(如 basal laminae 與 interstitial stroma)而使單 核球移入組織中(Ebnet & Jordan, 1999)。而 C-C chemokine MCP-1,對於單核球細胞的 migration,便是一種強效的趨化作用劑 (chemoattractant)。由於 MCP-1 能夠調控單 核球細胞的活化與遷移作用(Zhu, 1999),
我們的實驗便在觀察藥物對 MCP-1 所誘 發之 THP-1 細胞 migration 的情形,是否 有所影響。經過六個小時之後,由Figure 21 與 Figure 22 (100X)中可觀察到,在沒有 MCP-1 作用下,幾乎沒有 THP-1 細胞 migrate 到 lower chamber 中(lane 1: 2.4 ± 0.4 cells,n = 4),MCP-1(100 ng/ml) 激發 了約 34.84 倍的誘發反應(lane 2: 82.8 ± 24.4 cells,n = 4,p ﹤0.05)。若給予 50 ng/ml 的 MCP-1,則觀察到對 THP-1 細胞 的migration 約有 21.89 倍的誘發反應(52 ± 17.8 cells,n =2,data not shown)。MCP-1 所誘發的 THP-1 細胞的 migration 反應,
會隨著 cinnamophilin 濃度的增加而被減 弱。可觀察到Figure 21 中,lane 3: 35 ±12.9 cells,n = 4 (5 µΜ) 與 lane 4: 9.8 ±6.3 cells,n = 4 (20 µΜ)。而隨著 HDI II 濃度 (Figure 22: lane 3)的增加雖呈現上升的現 象,但仍對 MCP-1 所誘發的 THP-1 細胞 的migration 反應有抑制的作用:18.7 ±4.5 cells,n = 4 (0.5 µΜ) 與 lane 4: 52.2 ±17.7 cells,n = 4 (2 µΜ)。若延長作用時間為九 小時,在 100X 下觀察到為 24.5 ±11.5 cells (Resting),166.8±21.8 cells (MCP-1:
100 ng/ml),158 ±3 cells (cinnamophilin 5 µΜ),61 ±6 cells (cinnamophilin 20 µΜ),
129.5 ±14.5 cells (HDI II 0.5 µΜ),97.3 ± 8.3 cells (cinnamophilin 20 µΜ) 。MCP-1 所誘發的 THP-1 細胞的 migration 反應,
皆會隨著藥物濃度的增加有被減弱的表現 (n=2,data not shown)。
3-1-11. 探 討 藥 物 對 THP-1 細 胞 表 面 CD11b 表現之影響
為了測定LPS 調控單核球 THP-1 細胞 表面integrin CD11b 的功能, 與觀察藥物 在此所扮演的角色,我們先將THP-1 細胞 處理過LPS 與藥物後,以流式細胞儀計算 細胞表面 integrin CD11b 的變化情形。由
Figure 23 中可以觀察到在 LPS 存在下,細 胞表面 CD11b 的表現呈現有意義的增加 (lane 2: 1.31 ±0.1 folds,p<0.05,n = 4)。
Cinnamophilin (20 µΜ)與 HDI II (2 µΜ)會 些許提升 CD11b 的表現 (lane 3: 1.48 ± 0.06 folds,n = 4; lane 4: 1.34 ±0.07 folds,
n = 4),而 5 µΜ的 HDI II 則能抑制 LPS 誘 發的CD11b 表現(lane 5: 1.1 ±0.04 folds,n
= 2)。但與 lane 2 相比,藥物的給予對 CD11b 蛋白質的表現在統計上並無意義。
3-1-12.探討在 THP-1 細胞中 MCP-1 與 LPS 對 於 MMP-9 的 產 生 和 內 生 性 的 TNF-α之關係
過去的文獻曾指出,在單核球細胞中,
tumor necrosis factor-α (TNF-α) 與tumor 所促進產生MMP-9 有關。給予TNF-α的抗 體會抑制MMP-9 的生成、TNF-α mRNA 的合成與proMMP-9 蛋白質的釋放(Leber, 1998)。過去的研究曾指出,在單核球與巨 噬細胞中,LPS對於TNF- α的生成是一種 強效的誘發劑。在THP-1 與PBMCs中,LPS 皆 會 誘 導 非 常 顯 著 的 TNF- α 生 成
Mangalam & Haversen, 2001)。而Stephen 的研究指出, chemokine如MCP-1 要激發 出 MMP-9 的 表 現 是 需 有 autocrine 或 endogenous 的 TNF-α 存 在 (Stephen, 2002)。由於上述的種種研究顯示了TNF- α 對於MMP-9 在表現上重要性,我們取實驗 所 得 之 上 清 液 (supernatant) 並 使 用 了 Human TNF- ELISA s ystem測其TNF- α 的變化。由Figure 24 與 25 中可以觀察 到,在沒有LPS的刺激之下(Resting),以每 106的細胞為單位,便可偵測到些許程度 TNF- α的蛋白質表現 (17.3 ±10.3 pg/106 cells)。在經LPS (10 ng/ml) 刺激後,可觀 察TNF- α的量有一些上升的情形(26.2 ± 3.1 pg/106 cells,n = 3),但在統計上並無 明顯的差異。相對的,在高濃度100 ng/ml 的LPS刺激之下,TNF- α的量則有明顯上 升的情形(57.1±10 pg/106 cells,p<0.05,n = 2) 。而在經MCP-1 刺激的組別中(Figure 26、27),TNF- α的量與Resting (29.6 ±5.3 pg/106 cells,n = 3) 相比雖有些許上升 (42.8 ±8.3 pg/106 cells,n = 3),但在統計 上沒有明顯的差異。在分別加以藥物處理
後(cinnamophilin 1、5、20 µΜ),藥物對於 LPS刺激下細胞之TNF- α蛋白質皆呈現顯 著的上升:Cinnamophilin 1 µΜ (62.7±5.5 pg/106 cells,n =3,p<0.05)、5 µΜ (83.2±
9.7 pg/106 cells, n =3,p <0.005) 、20 µΜ (71.2 ±2.5 pg/106 cells,n =3,p<0.001) 。 在Figure 25 中,HDI II對於在LPS刺激下 TNF- α蛋白質的表現皆無明顯的影響:
HDI II 0.5 µΜ (42 ±6.1 pg/106 cells,n
=3)、1 µΜ (44.4 ±6.9 pg/106 cells,n =3)、
2 µΜ (32.3 ±12.8 pg/106 cells,n =3)。而在 MCP-1 的刺激之下(Figure 26、27),同樣 投與相同條件的物濃度(cinnamophilin 1、
20 µΜ; HDI II 0.5、2 µM),我們可以觀察 到在MCP-1 刺激下TNF-α蛋白質的表現在 cinnamophilin為 20 µΜ時97.9 ±29.7 pg/106 cells,n =3),比cinnamophilin為 1 µΜ時49.6
±2.6 pg/106 cells,n =3) 多,但與Vehicle 相比在統計上皆無明顯的差異。較高濃度 2 µM的HDI II (52.4 ±15.5 pg/106 cells, n
=3) 與 0.5 µM的HDI II (69.3 ±12.5 pg/106 cells,n =3) 相比似乎能減少些許TNF-α 蛋白的表現,在統計上卻無明顯的差異。
藥物與MCP-1 同時存在時,當與Resting相 較,可以觀察到對TNF-α蛋白質的表現似 乎有加強的效果。我們的結果表示了LPS 與MCP-1 雖能些略使TNF-α蛋白質表現量 上升,但與Resting相比在統計上卻無有意 義的差別。而藥物的作用並不是藉由影響 內生性TNF-α而對MMP-9 產生抑制作用。
3-1-13. 探 討 在 出 血 後 再 甦 醒 性 傷 害 中 MMP 抑制藥物之研究
由血壓之變化(Figure 28)、腎臟功能 (Figure 29-30)與肝臟功能(Figur 31-32)評 估顯示大鼠出血後再甦醒性傷害為多重器 官損傷。Anti-leukocyte agents 如 YC-1 (10 mg/kg)與 HDI II 類似作用藥 valproic acid (150 mg/kg)等藥物,雖對 MMP 有所部分 影響,但對出血後再甦醒性傷害作用僅部 分之作用(劑量或 PK 問題),仍須進一步探 討。
3-2 討論
白血球(leukocyte)滲入(extravasation) 與浸潤(infiltration)作用到受傷或是有發炎 反應的區域是一種複雜且多步驟的連鎖過
程。白血球對於內皮細胞產生的附著作用 (adhesion) 導 致 了 一 連 串 滾 動 (rolling) 、 loose 與 firm adhesion 再經由vessel wall 與basement membrane 遷移(migration) 滲 入 到extracellular matrix 中 。 白 血 球 的 extravasation 與 migration 等作用必須要 有 分 解 基 質 的 酵 素 , 特 別 是 matrix metalloptoinases (MMP)存在,才能讓白血 球穿越 basement membrane,到達組織的 基 質 中 (Lindhout, 1999; Ebnet, 1999;
Baggiolini, 1998)。這種白血球的 migration 作用在 athersclerotic lesion、thrombosis 的 產生與 rheumatoid arthritis 中皆扮演了非 常重要的角色(Chase, 2002; Falcone, 2001;
kintscher, 2000; Zhu, 1999) 。 而 因 為 Gelatinases MMP-2 與MMP-9 屬於第四 型的collagenases (Type IV collagenases),
能degrade basement membrane 的 蛋 白 質,在這個作用機制中便有其專一性的重 要 性 (Overall, 2001) 。 C-C chemokine MCP-1 對於單核球細胞的migration是一種 強效化學趨化作用劑(chemoattractant),
chemokines可以調控MMP-9 在peripheral blood lymphocytes 中 的 表 現 (Johnatty, 1997) 與發炎性細胞激素如TNF-α的生成 (Leber, 1998) , 然 而 在 monocyte 中 chemokines 如何調控MMP的表現,目前 目前所知的卻不多(Robinson, 2002); 而 LPS則和許多發炎等疾病息息相關,我們 的實驗中,使用了白血球中的單核球細胞 THP-1 來觀察在有外在LPS、MCP-1 與 TNF-α (data snot shown)的刺激之下,
THP-1 所產生MMP-9 反應與在有給予 cinnamophilin 與 HDI II 等藥物之下,藥 物 對 MMP-9 所 產 生 的 抑 制 作 用 。 Cinnamophilin 是 一 種 由 Cinnamomum philippinense 中所粹取出來的天然物,具 有free radical scavenging capacity (Hsiao, 2001),並且表現出其特有的抗氧化與保護 細胞免於氧化的傷害,在研究與治療與 TXA2 相關的疾病上被視為有用的工具。
在骨骼肌肉ischaemic 的model中(Cheng, 1995),cinnamophilin 更能預防進一步的 reperfusion 所 造 成 的 傷 害 。 而 histone deacetylase inhibitor trichostatin A (TSA) 可擇選則性的抑制 3T3 細胞中gelatinase
A 的表現與活化。另外,histone deacetylase (HDAC) inhibitors 在 不 同 種 類 的 transformed 細胞中,或是在tumor-bearing 的動物中,皆能有效的引起生長停滯、分 化或是apoptotic 細胞死亡。HDAC的抑制 劑 在 治 療cancer上,與一般抗癌藥物相 較,是為更有效的治療目標去對抗難治療 的癌症(Batova, 2002)。在參考許多學者對 於 cinnamophilin 與 histone deacetylase inhibitor 文獻之後,使我們對於這兩種藥 物在發炎性反應和MMP-9 的作用上產生 了興趣。由zymography的分析可以發現,
cinnamophilin或HDI II 能抑制在人類單核 球細胞 (THP-1)中,LPS或是MCP-1 所誘 發的MMP-9 酵素活性。在LPS (10 ng/ml) 的刺激之下,cinnamophilin與HDI II對於 MMP-9 的抑制 50 % 反應濃度(IC50)為 8.11 ±1.52 µM (n=3-4) 與 0.43 ±0.16 µM (n = 3-4)。而在MCP-1 (240 ng/ml)的刺激之 下,cinnamophilin 與HDI II對於MMP-9 的抑制 50 % 反應濃度(IC50)為 11.63 ± 3.04 µM (n = 3) 與 0.37 ±0.04 µM (n = 3-5)。無論是LPS或是MCP-1 在THP-1 細 胞 中 所 誘 發 之 MMP-9 皆 能 被 cinnamophilin 或HDI II等藥物隨濃度上升 而 呈 現 活 性 減 少 之 反 應 , 而 呈 現 出 concentration-dependent 的抑制作用。
為了證明cinnamophilin 與 HDI II 等藥 物對於THP-1 細胞中 MMP-9 活性的抑制 並非引起細胞減少或死亡,我們使用了 MTT assay 來 觀 察 。 高 濃 度 的 cinnamophilin (50 µM)並不會影響細胞的 存活率,但HDI II 到達 2 µM 時,則對細 胞的存活率有些許影響。我們可觀察到藥 物對於 MMP-9 本生的酵素活性不是直接 的抑制作用。
經由Western blot 分析,cinnamophilin 或 Histone deacetylase inhibitor II (HDI II) 能抑制在人類單核球細胞 (THP-1)中,
LPS 所 誘 發 的 MMP-9 蛋 白 質 表 現 。 Cinnamophilin 與 HDI II 對 MMP-9 的抑制 是藉由減少其蛋白質的表現,而這兩種藥 物皆能對在THP-1 細胞中,LPS 所誘發之
MMP-9 的 蛋 白 質 產 生
concentration-dependent 的抑制作用。經由
TIMP-1 ELISA 的分析,我們可以發現藥 物對於 MMP-9 表現量的減少與其內生性 的抑制因子TIMP-1 並不相關。而 RT-PCR 的實驗結果表示出cinnamophilin 與 HDI II 能夠concentration-dependent 的抑制由 LPS 所誘發的MMP-9 mRNA 的產生。由以上 的實驗結果,我們可以推論cinnamophilin 與 HDI II 的抑制作用在發生在 MMP-9 transcription 的 upstream。
許多常見的 inducible MMPs (MMPs 1、3、9 與 13) 含有 activator protein 1 (AP-1) 轉錄因子結合區域(binding site) 在它們近 側的 promoters 上,並需要經由細胞激素 的作用(如 TNF-α、IL-1)而被誘發。AP-1 因 子 是 由 Fos 與 Jun 蛋 白 所 組 成 的 heterodimers , 這 些 蛋 白 質 會 經 由 mitogen-actived protinse kinase (MAPKs)的 訊息傳遞路徑而被合成與活化(Firestein, 1999)。在許多組織細胞實驗,藉由阻隔 MAPKs 的路徑便能有效的抑制 MMPs 基 因的表現,而在動物模式中抑制 arthritis 的形成(Mengshol, 2002)。Nuclear factor κB (NF-κB) 的訊息傳遞路徑在對於 MMPs 的 抑制上提供了另一個有效的目標。這條訊 息傳遞路徑控制了許多誘發性的發炎基因 與MMPs 的表現。在接下來的實驗中,我 們便對NF-κB 與 MAPKs kinase 等訊息傳 遞路徑加以討論。藉由針對這些訊息傳遞 路徑的研究,進接便能鑑別出藥物對於調 控MMPs 合成的可能目標。我們利用了觀 察NF-κB 的抑制因子(inhibitory subunit),
IκB-α,在 Western blot 中蛋白質量的表現 情形,來瞭解藥物對於 IκB-α磷酸化的分 解作用與在 NF-κB 的訊息路徑上所扮演 的角色。發現 LPS 刺激 90 分鐘時所取得 之細胞lysate,IκB-α 的表現呈現明顯的降 解作用。以 90 分鐘為 LPS 刺激的基準,
分別投與不同濃度之藥物後,可觀查到 cinnamophilin 能抑制在 THP-1 中,LPS 所 引發的IκB-α degradation,而 HDI II 對此 作用則較不明顯。目前在癌症的治療上,
histone acetylation 調控機制的研究被認 為是可行的治療對策。過去的研究曾指 出,在 in vitro 模式中,給予細胞 histone deacetylse inhibitors 如 butyric acid 或
trichostatin (TSA),細胞會呈現 apoptosis、
生長停頓、分化與基因表現改變的情形,
指示出 hyperacetylation 與 carcinogenesis 之間的關連性(Archer, 1999)。而 HDAC inhibitors 是如何調控這整個機轉目前並不 是 非 常 瞭 解 (Kramer, 2001; Mayo, 2003) 。 若 排 除 藥 物 毒 性 的 影 響 , hydroxamic acid-based HDAC inhibitors 如 TSA 與 suberoylanilide hydroxamic acid (SAHA),會選則性的作用在基因上,在細
胞中進而去改變只有約 2 % 的基因表現
(Ailenberg, 2001; Marks & Kramer, 2001;
Archer, 1999)。SAHA 會增加調控 tumor 的基因而進一步的表現其 antitumor 的作 用。並且在 in vitro 與 in vivo model 中,
SHAH 能減少 proinflammatory cytokines 的產生(Leoni, 2002),給予小鼠單一口服計 量的 SAHA (10-50 mg/kg, per os) 能夠 dose-independently 的減少由 LPS 所誘發的 TNF- α、IL-1β、L-6 與 IFN-γ 等細胞激 素。而 TSA 能夠 dose-related 的減少與抑 制 fibroblast 3T3 細 胞 中 , gelatinase A mRNA 與 gelatinolytic 的活性 (Ailenberg, 2001)。而在 STO 細胞(mice embryonic fibroblasts)中,亦可觀察到相似的結果。
因 此 , 有 學 者 認 為 可 藉 由 監 控 與 研 究 gelatinase A 的表現程度,而當作早期階段
的生物參數來監測 TSA 在臨床試驗上的
功效。若從結晶學上的研究發現去解釋,
HDAC 屬於一種包含了 zinc 在其催化區域 的金屬蛋白(Finnin, 1999)。一旦 hydroxate 如 TSA 與 zinc 結合,便能抑制住催化區 域的活性而抑制MMPs 的活性。然而 TSA 與 HDI II 的作用都是經由抑制 MMPs (gelatinse A 與 gelatinase B)的 mRNA。因 此,對於疾病或癌症的早期治療可能有其 功效。Histone deacetylase inhibitor II 與 TSA 同屬於 hydroxamic acid-based HDAC inhibitors (Marks, 2001) ,是否可能藉由基 因 的 改 變 , 而 對 gelatinase B , MMP-9 mRNA 與 gelatinolytic 的活性產生抑制,
則 需 進 一 步 的 研 究 。 我 們 結 果 發 現 , cinnamophilin 可能會經由抑制 IκB-α蛋白 質的 degradation 而進一步的減少 NF- B translocate 至細胞核內的作用。雖曾有文 獻指出 LPS 所誘發單核球細胞中 MMP-9
的生成,主要是經由ERK 1/2 這條訊息傳 遞路徑(Lai, 2003),但在 THP-1 細胞中,
以 我 們 的 實 驗 結 果 觀 察 ,LPS 所 誘 發 MMP-9 蛋白質的表現作用,可能經由 JNK 的 訊 息 傳 遞 路 徑 。 我 們 可 以 推 斷 cinnamophilin 與 HDI II 可能會經由抑制 JNK蛋白質的表現而進一步的抑制轉錄因 子AP-1 的活化與 MMP-9 基因的表現,而 並非經由訊息傳遞路徑 ERK 與 p38 (data not shown)等 MAPKs pathways。
當組織受傷或是有發炎反應產生時,
白 血 球(leukocyte) 會 滲 入 (extravasation) 與浸潤(infiltration)到受傷或是有發炎反應 的區域,其中需要MMPs 的作用去分解細 胞外的基質(如 basal laminae 與 interstitial stroma)而使單核球移入組織中(Ebnet &
Jordan, 1999)。由於 MCP-1 對於單核球細 胞的migration,便是一種強效的趨向作用 劑(chemoattractant),能夠調控單核球細胞 的活化與遷移作用(Zhu, 1999),而與這種 複雜且多步驟的連鎖過程息息相關。 我們 觀察藥物對 MCP-1 所誘發之 THP-1 細胞 migration 的情形。在 migration 實驗中可 觀 察 到 ,MCP-1 (100 ng/ml) 加強了約 34.84 倍 的 趨 化 反 應 , 比 50 ng/ml 的 MCP-1 21.89 倍高。而在藥物存在下,
MCP-1 所誘發的 THP-1 細胞的 migration 反應,會隨著cinnamophilin 濃度的增加而 被減弱,而隨著HDI II 濃度的增加呈現上 升的現象,但仍對MCP-1 所誘發的 THP-1 細胞的migration 反應有抑制的作用。由以 上的實驗我們可以推斷,cinnamophilin 與 HDI II 皆能對在 THP-1 細胞中,MCP-1 所 誘發的細胞migration 作用,產生一些程度 上的抑制作用,但與Vehicle 相比並無統計 上的意義。
白血球(leukocyte)遷移到受傷或是有 發炎反應的區域,是一連串複雜的過程。
除了有之前提到的 MMPs 與 chemokines 的參與之外,還包括了 seletin 和 integrin adhesion molecules 如 ICAM-1、VCAM-1 的作用。單核球細胞能表現 β1 integrin CD49d/CD29 與 2 integrin CD11a/CD18、
CD11b/CD18 、 CD11c/CD18 等 (Han, 2003)。Integrins 不但能調控朝發炎區域移
動的白血球與專一性的ligand 結合,還能 將訊息傳入細胞並且控制一些effectors 的 功能。基於在白血球上,CD11b/CD18 是 最多的 β 2 integrin,作為許多種類 ligands 如iC3b、ICAM-1、coagulation factor X 與 fibrinogen 的接受體,並在 locomotion、
chemotaxis 與 phagocytosis 上有其重要性 (Monneret & Rubel, 2003; Schwarz, 2002),我們測定了 LPS 調控單核球 THP-1 細胞表面integrin CD11b 的功能與觀察藥 物在此所扮演的角色,並以流式細胞儀計 算細胞表面 integrin CD11b 的變化情 形。由Figure 23 中可以觀察到在 LPS 能 誘發細胞表面CD11b 的表現 (lane 2: 1.31
±0.1 folds,p<0.05,n = 4),而 5 M 的 HDI II 則能抑制 LPS 誘發的 CD11b 表現 (lane 5: 1.1 ±0.04 folds,n = 2)。MA-1(20 µM)與 HDI II (2 µM)會些許提升 CD11b 的 表現 (lane 3: 1.48 ±0.06 folds,n = 4; lane 4: 1.34 ±0.07 folds,n = 4),但與 lane 2 相 比,藥物的給予對CD11b 蛋白質的表現在 統計上並無意義。由初步實驗的結果我們 可推斷,藥物可選擇性的抑制 MMP-9 蛋 白質的表現。
過去的研究曾指出,在單核球與巨噬細 胞中,LPS對於TNF-α的生成是一種強效 的誘發劑。在THP-1 與PBMCs中,LPS皆 會誘導非常顯著的TNF-α生成Mangalam
& Haversen, 2001)。而Robinson的研究指 出, chemokine如MCP-1 要激發出MMP-9 的 表 現 是 需 有autocrine 或 endogenous 的 TNF-α 存在(Robinson, 2002)。基於上述的 種種研究顯示了TNF-α對於MMP-9 在表 現上重要性,我們觀察了內生性的TNF-α 的表現情形。細胞在經LPS (10 ng/ml) 與 MCP-1(240 ng/ml)刺激後,可觀察TNF-α 的量有一些上升的情形(26.2 ±3.1 pg/106 cells,n = 3 與 29.6 ±5.3 pg/106 cells,n = 3),但在與Resting相比在統計上並無明顯 的差異(Vehicles: Figure 24-27)。相對的,
在 高 濃 度 100 ng/ml 的 LPS 的 刺 激 之 下 (Figure 31 & 32),TNF-α的量則有明顯上 升的情形(57.1 ±10 pg/106 cells,p < 0.05,
n = 2) 。 在 分 別 加 以 藥 物 處 理 後 (cinnamophilin 1、5、20 µM),藥物對於