轉錄因子Alpha-Pal/NRF-1在小腦發育及神經分化之角色
20
0
0
全文
(2) 附件一. 行政院國家科學委員會補助專題研究計畫. ■成 果 報 告 □期中進度報告. 轉錄因子 alpha-Pal/NRF-1 在小腦發育及神經分化之角色 Role of transcription factor alpha-Pal/NRF-1 in the cerebellar development and neuronal differentiation. 計畫類別:■ 個別型計畫 □ 整合型計畫 計畫編號:NSC 94-2320-B-006-053- 執行期間: 94 年 8 月 1 日 至 95 年. 7 月. 31 日. 計畫主持人:黃阿敏 共同主持人: 計畫參與人員:謝崇斌. 成果報告類型(依經費核定清單規定繳交):□精簡報告 ■完整報告 本成果報告包括以下應繳交之附件: □赴國外出差或研習心得報告一份 □赴大陸地區出差或研習心得報告一份 □出席國際學術會議心得報告及發表之論文各一份 □國際合作研究計畫國外研究報告書一份 處理方式:除產學合作研究計畫、提升產業技術及人才培育研究計畫、 列管計畫及下列情形者外,得立即公開查詢 □涉及專利或其他智慧財產權,□一年□二年後可公開查詢 執行單位: 中 華 民. 國. 95. 年. 10. 月. 28. 日.
(3) 中文摘要 Nuclear respiratory factor-1(NRF-1)是細胞核內的轉錄因子,在先前的研究,我們證實 NRF-1 可以調控人類 integrin-associated protein (IAP)基因啟動子的活性。我們的研究也發 現,在中樞神經系統中, IAP 與記憶形成和神經突生長有關。在斑馬魚中,與 NRF-1 有高 度相似性的基因 not really finished (nrf)在中樞神經發育扮演了重要的角色,然而在哺乳類中 NRF-1 對腦部發育的角色尚未清楚。因此,本研究主要探討 NRF-1 及 IAP 兩基因在哺乳類 腦發育過程中的表現情形及可能功能。原位雜交法(in situ hybridization)結果顯示,在胚胎 第 15 天,IAP mRNA 廣泛分布在腦中,隨後在發育過程分佈至特定腦區,包括端腦 (telencephalon)、間腦 (diencephalon),和菱腦唇 (rhombic lip)。在出生後時期,明顯的變化 發生在發育中的小腦。從出生後第 6 至第 14 天,IAP mRNA 表現隨著顆粒前驅細胞 (granular cell precursors, GCPs)由 external granular layer (EGL) 遷移至 internal granular layer (IGL)其 表現量逐漸增加。免疫組織染色法顯示 IAP 蛋白表現隨著小腦成熟,在分子層 (molecular layer)與 IGL 表現量亦逐漸上升。另外在 EGL 中,顆粒前驅細胞同時表現 IAP 與 TAG-1, 顯示 IAP 參與在顆粒前驅細胞遷移過程中。Gel mobility shifted assay (EMSA)顯示 NRF-1 在 IAP 啟動子的結合活性隨著小腦發育有所變化,與 IAP 基因表現一致。在神經突部份,調 控 NRF-1 與 IAP 對小腦神經細胞神經突生長有顯著的影響。過度表現 NRF-1 減少了神經突 的長度,而顯著抑制型 NRF-1 則降低其長度及數目。IAP form 4 cDNA 抑制了神經突伸長, 然而 antisense IAP cDNA 減少了神經突數目。這些結果顯示 NRF-1 及其下游基因 IAP 對小 腦的發育及其神經細胞的分化扮演了重要的角色。 關鍵字:轉錄因子 NRF-1、小腦發育、神經突生長 Abstract Nuclear respiratory factor 1 (NRF-1) is a nuclear transcription factor that was found to regulate the human integrin-associated protein (IAP or CD47). In the central nervous system (CNS), IAP is associated with memory formation and neurite outgrowth. Recently, we have found a novel function of NRF-1 in neurite outgrowth which is mediated by the expression of its downstream IAP gene. Not really finished (nrf) gene, a homologue of NRF-1 in the zebra fish, plays an important role in the development of CNS. However, the role of NRF-1 in the development of mammalian brain is unclear. In situ hybridization showed that IAP mRNA is widely expressed in the brain at embryonic day 15 (E15), and located distinctively in regions including telencephalon, diencephalon, and rhombic lip during embryonic development. Striking changes in the expression of IAP mRNA were found in the cerebellum at postnatal stages. Increased expression of IAP mRNA was found from postnatal 6 to 14 days (P6 to P14), which is correlated with the migration of granular cell precursors (GCPs). Immunohistochemistry also demonstrated that the expression of IAP in the molecular layer and internal granular layer of the cerebellum increases from P6 to P14. Co-localization of IAP and TAG-1 in the external granular layer (EGL) from P6 to P8 implied that IAP is involved in the migration of GCPs from EGL to IGL. Gel mobility shifted assay (EMSA) showed that the binding activity of NRF-1 to the IAP promoter in the cerebellum was correlated with IAP mRNA expression from P1 to adult. In the primary cerebellar cells, overexpression of full-length NRF-1 significantly decreased the length of neurites, and dominant negative NRF-1 decreased the length and number of neurites. IAP form.
(4) 4 cDNA inhibited neurite elongation; however antisense IAP cDNA reduced the number of neurites. These results suggest that NRF-1 and its downstream gene IAP play a role in the cerebellar development and neuronal differentiation. Key words: Nuclear respiratory factor-1, cerebellar development, neurite outgrowth. 研究目的 1. 2. 3. 4.. To examine the expression of IAP mRNA during brain development. To investigate the expression of IAP mRNA and protein during cerebellar development. To characterize the relationship between NRF-1 and IAP during cerebellar development. To examine the effects of NRF-1 and IAP on the neurite outgrowth in primary cerebellar neurons.. 前言 Nuclear respiratory factor-1 (NRF-1) is a nuclear transcription factor which was originally found to regulate the eukaryotic initiation factor 2gene and genes related to mitochondrial functions and cell cycle regulation (Chau et al., 1992; Efiok and Safer, 2002; Scarpulla, 2002). NRF-1 belongs to a novel transcription factor family with leucine zipper DNA-binding domain in the N-terminal and trans-activation domain in the C-terminal half (Virbasius et al., 1993; Gomez-Cuadrado et al., 1995). The DNA-binding domain of NRF-1 shows high homology with two genes on the invertebrate including of sea urchin P3A2 and Drosophila erect wing gene (ewg) (Virbasius et al., 1993; Efiok, et al., 1994), which have been implicated in the embryonic or larval development. Furthermore, not really finished (nrf) gene of the zebrafish showed 91% identity with human NRF-1 and the high expression in the developing central nervous system (CNS) implicated it plays an important role during brain development in zebrafish (Becker et al., 1997). However, the role of NRF-1 during brain development in mammalian is still unclear. In the previous study, we have found that NRF-1 was a critical transcription factor of the human intergrin-associated protein (IAP) gene. IAP belongs to the member of the immunoglobulin (Ig) superfamily, possessing an Ig-V–like extracellular region, five putative transmembrane domains, and a short cytoplasmic tail (Brown and Frazier, 2001). IAP is implicated in the regulation of multiple cellular processes including neutrophil migration (Cooper et al., 1995; Parkos et al., 1996), T-cell activation (Reinhold et al., 1997), and platelet activation (Chung et al., 1997). In the CNS, the expression level of IAP in the hippocampus was associated with memory performance (Huang et al., 1998). Studies showed that IAP promotes neurite outgrowth in the neuroblastoma cell (Miyashita et al., 2004) or primary cortical and hippocampal culture neurons (Numakawa et al., 2004; Ohnishi et al., 2005). Furthermore, we have found that N56R5, a stable cell line overexpressing NRF-1, bears more and longer neurites, and this promotion of neurite outgrowth is mediated by the downstream gene IAP (Chang et al., 2005). The effects of NRF-1 in neurite outgrowth were also found in the primary cortical neurons (Chang et al., 2005). These line of evidences indicated that NRF-1 and IAP in involved in neuronal differentiation. However, little is known about the effects of NRF-1 and IAP in brain development. In this study, we will.
(5) investigate the role of NRF-1 and its downstream gene IAP in the mammalian brain development.. 研究方法 In situ hybridization Brain sections. Postnatal ICR mice (P1, P6, P7, P8, P10, P12, P14, P21, P28 and 3-month old adult) were sacrificed and brains were removed immediately and placed in ice-cold phosphate-buffered saline for 5 min. For embryos, the pregnant ICR mice were sacrificed immediately and embryos were removed and placed in ice-cold MEM medium. Brains and embryos were embedded with O.T.C. and frozen at -55 °C for 10 min, and then stored at -80 °C. Serial sections at 10-μmthickness were cut on cryostat, thaw-mounted onto 100 g/ml poly-l-lysine-coated slides and stored at -80 °C until use. Frozen sections were fixed by 4% paraformaldehyde for 2 hr and then digested with 0.05 g/ml proteinase K at 37°C for 7.5 min in the buffer containing 0.05 M Tris-HCl, pH 8.0 and 0.005M EDTA. Acetylation was performed by immersion of brain sections twice for 5 min in the solution containing 0.25% acetic acid in 1M Tris-HCl, pH7.5. The sections were then dehydrated through a series of ethanol solution from 50, 70, 80, 95, to 100% (15 sec each) and stored at -80 °C. cRNA probes. The mouse IAP cDNA were amplified by RT-PCR (Forward primer: rIAPF1, 5’ -TCC AGA GAA GGC AAA ACA GT-3’ ; reverse primer: rIAPR1, 5’ - GGC AAT GGT GAA AGA GGT C-3’ ). The PCR products were inserted into the pCRII vector (Invitrogen). The result plasmid was linearized by HindⅢ and used as a template for in vitro transcription. A 20 μl-reaction mixture containing 8 mM Tris-HCl, pH 7.9, 1.2 mM magnesium chloride, 0.4 mM spermidine, 2 mM sodium chloride, 10 mM DTT, 20 U RNase inhibitor, 3.3 mM rATP, rCTP and rGTP mixture, 0.1 mM rUTP, 60 μCi α-35S rUTP and 10 U T7 RNA polymerase (Promega) was incubated at 37 °C for 1 h and then the template DNA was removed by the DNaseⅠtreatment at 37 °C. Labeled cRNA probes were then purified by Sephadex G-50 columns. Hybridization, RNase A treatment, post-hybridization washing, and x-ray film exposure. Labeled IAP cRNA(10× 103 cpm)probes were pre-heated at 65 °C for 5 min in the hybridization solution (50% formamide ,4xSSC,1xDe nha r t ’ ssolution, 0.25 mg/ml tRNA, 10% dentran sulfate, 0.5 mg/ml salmon sperm DNA, and 1 mM DTT). After pre-heating, 200 μl of probes were applied onto each brain section and sections were covered with parafilm and incubated at 55 °C for 16h. After hybridization, parafilm were removed and sections were rinsed in 4 x SSC containing 1 mM DTT and then incubated in 30 ng/μl RNase A (in 0.5 M NaCl, 10 mM Tris-HCl, and 5 mM EDTA) at 37 °C for 30 min. After RNase A treatment, the sections were washed in 2 x and 1 x SSC containing 1 mM DTT at room temperature for 30 min and then finally washed in 0.5 x SSC containing 1 mM DTT at 55 °C for 30 min. The sections were dehydrated through a series of ethanol from 50, 70, 80, 95, to 100% (15 sec each) and exposed to X-ray film at room temperature for 3-5 days. Immunohistochemistry Ten m frozen sections were fixed with 4% paraformaldehyde for 2 hr, and pre-incubated.
(6) with blocking solution (5% normal goat serum and 0.5% Triton X-100 in PBS) for 1 hr, and then incubated with primary antibody mIAP301 (1:500, BD) and TAG-1 (1: 1, HBDS) in the blocking solution overnight. The secondary antibodies Alexa-Flour 555 goat (1:500, Invitrogen) anti-rat and Alexa-Flour 488 (1:400, Invitrogen) in the blocking solution were used. In the condition of double staining, Alexa-Flour 555 was used to interact with mIAP301, before tissue reacting with the TAG-1 antibody. For the identification of nucleus, Hoechst (1 ng/ml in PBS) was used. Slides were mounted with mounting medium (Vector) and observed under fluorescence microscopes (Leica). Gel electrophoretic mobility shift assays (EMSA) The double-stranded oligonucletotide probes were prepared by the fill-in method described in the previous study (Chang and Huang, 2004). The sequences of the two oligonucleotides are: foward, 5'-GAG TGC GCG TGC GCG GCT CT-3'; reverse, 3'-TCA CGC GCA CGC GCC GAG AG-5'. Thirty pmole of each of the forward and reverse oligonucleotides placed in a volume of 23 l of 1 × Klenow (DNA polymerase) buffer were heated at 94 °C for 2 min and annealed at room temperature for 30 min. The annealed double-stranded oligonucleotides were end-labeled by a fill-in reaction using DNA polymerase (Klenow) (Promega). One unit of the DNA polymerase (Klenow) and 40 μCi of [α-32P]dCTP (Perkin-Elmer Life Science) was added into the annealed oligonucleotides and the mixture was incubated at 30 °C for 15 min. The labeled oligonucleotides were purified by Sephadex G-50 columns (Amersham Pharmacia Biotech). The nuclear extracts were obtained from the cerebellum of mice at P1, P7, P14, P21, P28 and the adult of 18-week old. The DNA-binding reaction was conducted at 4°C for 30 min in a mixture containing 3 μg of nuclear extract, 10 mM Tris-Cl (pH 7.5), 50 mM sodium chloride, 0.5 mM dithiothreitol, 0.5 mM EDTA, 1 mM magnesium chloride, 4% glycerol, 0.075 μg poly(dI-dC).poly(dI-dC) (Amersham Pharmacia Biotech), and 2 × 104 cpm of 32P-labeled double-stranded oligonucleotides. The mixture with additional anti-NRF-1 serum (gift kindly from Scapulla) was mixed for 3 hr before adding the probed. Samples were analyzed on a 4% polyacrylamide gel (acrylamide/bisacrylamide 29:1 in 0.5 × Tris-borate-EDTA buffer) at 10 V/cm for 2.5 hr. The gel was dried and analyzed by autoradiography. Primary cerebellar culture and transient transfection Whole brains were removed from mice at postnatal day 3 and placed on ice-cold MEM medium. The cerebella were dissected out and transferred to fresh ice-cold MEM. Cerebellar tissues were triturated with a pipette with 250 l tip at the pole. Dissociated cells (2 × 105) were plated onto 12-mm coverslips coated with 0.1 mg/ml poly-L-lysine and grown for 1 h in MEM supplemented with 10% fetal bovine serum, 50 U/ml penicillin and 50 μg/ml streptomycin. Different plasmid was transfected into cells by calcium phosphate precipitation method (Jordan et al., 1996). Briefly, plasmid DNA (1 μg each) was mixed with 5 μl of 2.5 M calcium chloride in a volume of 50 μl containing 0.1× TE buffer. The mixture of the plasmid and calcium chloride was mixed with 2× HEPES buffer and incubated for 1 min at room temperature. The mixture was added onto the cells for transfection. After 12 h of transfection, the medium was replaced with fresh medium without antibiotics and the cells were cultured for another 48 h for neurite measurement..
(7) Immunoflurorescence microscopy and measurements of neurite parameters The cells transfected with different plasmids were washed with PBS and fixed with 4 % paraformaldehyde for 10 min. After a 1-h block with 10% normal goat serum and 0.1% Triton X-100 in PBS, cells were incubated with monocolonal anti-MAP 2a,b antibody diluted 1:400 in blocking solution at 4 °C for overnight. The cells were washed with PBS and incubated for 1 h with the Alexa-Flour 594 conjugated with goat anti-mouse antibody (1:400) and Alexa-Flour 488 conjugated anti-GFP antibody (1:400, Molecular Probes) diluted in blocking solution. Following a final wash, the dishes were mounted with mounting medium (Vector) and sealed with nail polish. The cells were observed under fluorescence microscopes. To determine the number and length of the neurites, GFP/MAP 2a,b-positive cells were observed under the fluorescent microscope (Leica). The length of the process was determined by the software of Metamorph. The process longer than 4 m from soma is defined as a neurite.. 結果及討論 In situ hybridization analysis of IAP gene expression during development To investigate the role of IAP during development, we examined at first the expression of IAP mRNA by in situ hybridization. Whole embryo from embryonic day 15 (E15), E17, and E19, and whole brain from postnatal day 1 (P1), P7, P14, P21, P28, and adult (about 18 weeks) were examined. IAP transcripts were widely expressed in the whole brain at E15, especially in telencephalon, diencephalon, and rhombic lip (Fig. 1A). These structures are the origins of cerebral cortex, thalamus, and cerebellum. At E17, IAP was expressed in similar regions. The signals in the telencephalon at this stage were concentrated in the ventricular zone (Fig. 1B), where the neuroepithelium surrounds the lateral ventricle. At E19, the IAP mRNA was mainly expressed in differentiated layers of the cerebral cortex and the external granular layer (EGL) of the cerebellum (Fig. 1C). In the peripheral tissues, strong expression of IAP was found in the liver at E15 (Fig. 1A) and was gradually decreased from E17 to E19 (Fig. 1B & C). The high abundance of IAP mRNA expression in the liver was also found in a previous study, in which IAP was reported to be expressed in the stromal cells of fetal liver (Furusawa et al., 1998). At postnatal stages, the IAP mRNA was detected in most brain regions including cortex, thalamus, hippocampus and cerebellum (Fig. 1D to 1I). The expression level of IAP were systemically increased gradually from P1 to P28 (Fig. 1D to 1H), but slightly decreased in the adult brain (Fig. 1I). The highest expression level of IAP was found in the cerebellum from P7 to adult (Fig. 1E to 1I). At P7, IAP was highly expressed in the EGL of cerebellum (Fig. 1E). However, the signals of IAP in the EGL at P14 were almost disappeared, and higher abundance was found in the internal granular layer (IGL) of cerebellum (Fig. 1F). The highest expression level of IAP in the cerebellum was found at P14 and gradually decreased from P21 to adult (Fig 1F to 1I). Strong signals of IAP in the telencephalon and in its differentiated regions of cortex and hippocampus suggest that IAP was involved in the development of cortex and hippocampus, consisting with recent findings that IAP promotes neurite outgrowth in the cortical and hippocampal cultured neurons (Numakawa et al., 2004; Ohnishi et al., 2005). The IAP sense probes gave almost no signal (Fig. 1J) revealed the specificity of detection..
(8) The expression pattern of IAP mRNA during development was summarized in Table 1. There are waves of the expression levels of the IAP mRNA in most of the brain regions during development. IAP mRNA levels decreased from E15 to P1, increased from P7 to P28, and decreased again in the adult brain. The highest IAP mRNA level was found in the cerebellum after P7, compared to other brain regions, suggesting that the IAP gene plays an important role in the development of cerebellum. In situ hybridization analysis of IAP gene expression during cerebellar development To investigate more detail the expression of IAP mRNA during cerebellar development, 6 stages of the brain including postnatal day 6, 7, 8, 10, 12, and 14 were examined. At P6 and P7, IAP mRNA was found only in the EGL of the cerebellum (Fig. 2A & B), and the signal of IAP mRNA first appeared in the IGL at P8 (Fig. 2C). The intensity of IAP mRNA signals became stronger in the IGL from P8 to P12. In the contrary, lower and lower expression level of IAP was found in the EGL from P8 to P12 (Fig. 2D to 2E). At P14, IAP mRNA was almost disappeared in the EGL (Fig. 2F). It is known that the granular cell precursors (GCPs), which are differentiated from rhombic lip, proliferate in the EGL in the developing mouse cerebellum. At P6, the GCPs begin to migrate radially along the Bergmann glial cells into the IGL and become the matured cerebellar granular cells. The cerebellum completes maturation at about 3 weeks after birth in mice (Komuro and Yacubova, 2003). Our results suggest that the transition of IAP mRNA expression from EGL to IGL may correlate with the migration of GCPs from the EGL to the IGL during cerebellar development. Immunohistochemistry analysis of IAP protein expression during cerebellar development To further characterize the role of IAP during cerebellar development, immunohistochemistry was used to examine IAP protein expression in the cerebellum from P6 to P14. At P6, IAP-positive signals were very week in the cerebellum. At this stage, Hoechst 33258 signals revealed that the EGL is much thicker than the molecular layer (ML) (Fig. 3A). From P7 to P14, strongest signals were detected in the ML. The intensity of IAP signals was correlated with the increase of width of the ML and decrease of width of the EGL (Fig. 3B to F). The IAP signals were also detected in the IGL. The intensity was increased from P7 to P14 (Fig. 3B to F). In the EGL, we noticed that the IAP signals were detected in the inner layer, surrounding the cells. IAP was localized in the migrating granular cell precursors In the EGL, the granular cell precursors proliferate in the outer layer of EGL, enter the inner layer of EGL (iEGL) and stop proliferation, and begin to migrate radially along the Bergmann glia cells into the IGL (Choi et al., 2006). We proposed that the expression of IAP in the inner layer of EGL is correlated to the migration of granular cells from EGL to IGL. TAG-1 is a unique maker of migrating GCPs in the iEGL and was used to identify this region (Pickford, 1989). Immunohistochemistry demonstrated that the IAP expression from P6 to P8 (Fig. 4A, D & G) in the inner layer of EGL is colocalized with TAG-1 (Fig. 4B, E & H), suggesting that expression of IAP in the GCPs may initiate the migration of granular cells from EGL to IGL..
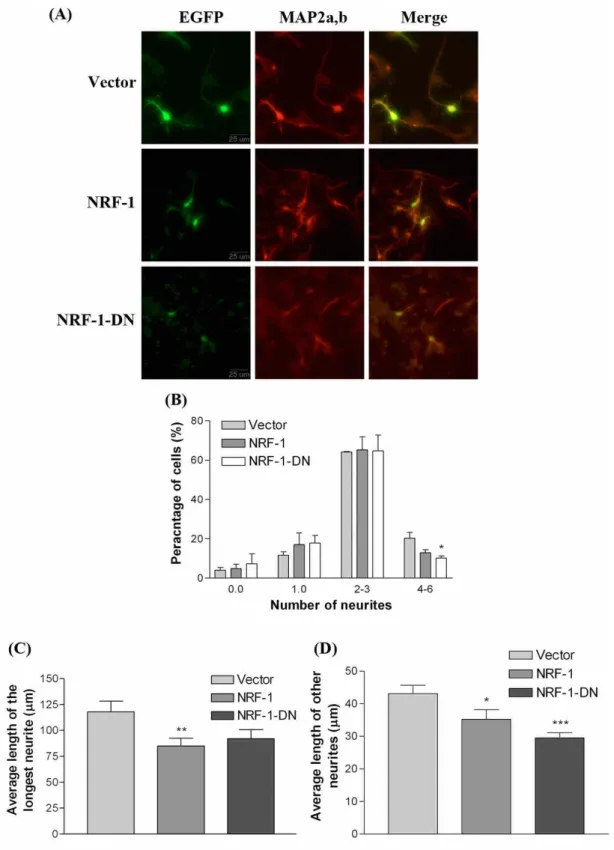
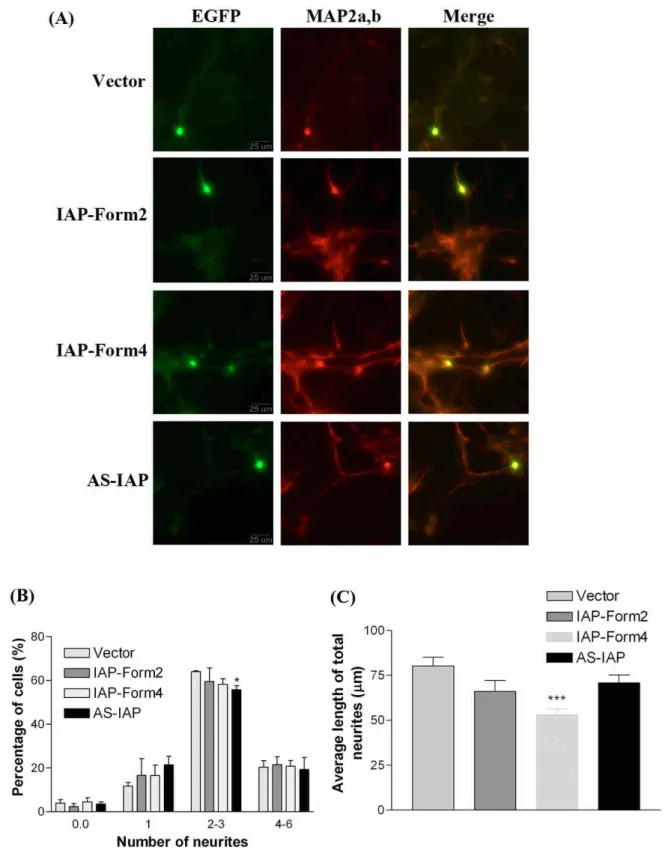
(9) The functional activity of NRF-1 during cerebellar development In the previous study, we have found that the transcription factor NRF-1 regulates the human IAP promoter (Chung and Huang, 2004). To investigate functional activity of NRF-1 during cerebellar development, gel mobility shifted assay (EMSA) was used to examine the binding activity of NRF-1 in the cerebellum from P1 to adult. As shown in Fig.5, the NRF-1 shifted band at P1 and P7 showed low signals and higher signals appeared at the P14 and P21, but slightly decreased at P28 and the adult. The tendency of NRF-1 binding activity was correlated with the IAP mRNA expression level in the cerebellum from P1 to adult (Fig. 1D to 1I; Table. 1). NRF-1 regulates neurite outgrowth in primary cerebellar neurons To investigate whether NRF-1 is involved in the differentiation of cerebellar neurons, the effects of overexpression of NRF-1 on neurite outgrowth in primary cerebellar neurons were examined. Primary cerebellar cells were transfected with pCMS-EGFP plasmid DNA containing NRF-1 cDNA or dominant negative NRF-1 cDNA which lacks the trans-activation domain to compete for the function of endogenous NRF-1. The neuronal marker MAP2a.b was used to identify neurons, and GFP expressing cells represented transfected cells. The morphology of MAP2a,b and GFP positive cells were observed under a fluorescent microscope (Fig. 6A). The process protruded from soma longer than 4 m was identified as a neurite. The transfected neurons were divided into several groups of cells containing 0, 1, 2 to 3, and 4 to 6 neurites. Overexpression of dominant negative NRF-1 significantly decreased the percentage of neurite bearing cells containing 4 to 6 neurites (Fig 6B), but no effects was observed on the full length NRF-1 group. On the other hand, overexpression of full length NRF-1 reduced average length of longest neurites and other neurites, and dominant negative NRF-1 also had tendency to reduce the average of longest neurite, and significantly decreased the other neurite length. (Fig.6 C & D). These results showed that NRF-1 is involved in the neurite outgrowth of cerebellar neurons. However, the inhibition of neurite elongation caused by overexpression of NRF-1 suggested that there should be a critical level of NRF-1 to regulate neurite elongation in cerebellar neurons. The neurite outgrowth of primary cerebellar neurons was regulated by IAP The plasmid vector pCMS-EGFP vector containing full length IAP form 2, or form 4 cDNA, and antisense IAP form 4 cDNA was transfected into primary cerebellar cells. Form 4 IAP is the major isoform expressed in the brain. GFP-positive and MAP2a,b-positive transfected cells were chosen to measure the neurite number and length (Fig. 7A). Overexpressing antisense IAP significantly reduced the percentage of cells containing 2 to 3 neurites (Fig. 7B). There were no effects of overexpressing IAP form 2 or antisense IAP on the average length of neurites, but cells with overexpression of IAP form 4 cDNA showed significant reduction in average length of neurites (Fig 7C). Theses results indicate that IAP also regulates the neurite differentiation of cerebellar neurons. The inhibition of neurite elongation by overexpressing NRF-1 and IAP, which is contrast to the results in the neuroblastoma cells and primary cortical neurons reported in the previous study (Chang et al., 2005), suggesting that different mechanism is involved in different cell type. 結論.
(10) IAP is expressed in the telencenphalon, diencenphalon, and rhombic lip in embryonic stages. In the postnatal brain, IAP is widely expressed in the cortex, hippocampus, thalamus, and cerebellum with a specific pattern of wave form. The strong expression of IAP in cerebellum suggests it plays a role in cerebellar development. Colocalization of IAP and TAG-1, the marker of migrating granular cells, suggests that expression of IAP may involve in the migration of granular cells from external granular cell layer to inner granular cell layer. High correlation between NRF-1 binding activity and IAP mRNA expression suggests that NRF-1 is also involved in the cerebellar development. Finally, both NRF-1 and IAP affect neurite initiation and elongation in the primary cerebellar neurons. These results suggest that NRF-1 and its downstream gene IAP play a role in the cerebellar development and neuronal differentiation. 參考文獻 Becker TS, Burgess SM, Amsterdam AH, Allende ML, Hopkins N (1998) not really finished is crucial for development of the zebrafish outer retina and encodes a transcription factor highly homologous to human Nuclear Respiratory Factor-1 and avian Initiation Binding Repressor. Development 125: 4369-4378. Brown, EJ, and Frazier, WA (2001) Integrin-associated protein (CD47) and its ligands. Trends Cell Biol. 11: 130–135. Chang WT, Chen HI, Chiou RJ, Chen CY, Huang AM (2005) A novel function of transcription factor alpha-Pal/NRF-1: increasing neurite outgrowth. Biochem Biophys Res Commun 334:199-206. Chang WT and Huang AM (2004) Alpha-Pal/NRF-1 regulates the promoter of the human integrin-associated protein/CD47 gene. J Biol Chem 279: 14542-14550. Chau CM, Evans MJ, Scarpulla RC (1992) Nuclear respiratory factor 1 activation sites in genes encoding the gamma-subunit of ATP synthase, eukaryotic initiation factor 2 alpha, and tyrosine aminotransferase. Specific interaction of purified NRF-1 with multiple target genes. J Biol Chem 267: 6999-7006. Choi Y, Borghesani PR, Chan JA, Segal RA. (2005) Migration from a mitogenic niche promotes cell-cycle exit. J Neurosci 25:10437-45. Chung, J., Gao, A.G., and Frazier, W.A. (1997). Thrombospondin acts via integrin-associated protein to activate the platelet integrin II3. J. Biol.Chem. 272: 14740–14746. Cooper, D., Lindberg, F.P., Gamble, J.R., Brown, E.J., and Vadas, M.A. (1995) Transendothelial migration of neutrophils involves integrin-associated protein (CD47). Proc. Natl. Acad. Sci. USA 92: 3978–3982. Efiok, B. J. S., Chiorini, J. A., and Safer, B. (1994) A key transcription factor for eukaryotic initiation factor-2 is strongly homologous to developmental transcription factors and may link metabolic genes to cellular growth and development. J. Biol. Chem. 269:18921-18930. Efiok BJ and Safer B (2000) Transcriptional regulation of E2F-1 and eIF-2 genes by alpha-pal: a potential mechanism for coordinated regulation of protein synthesis, growth, and the cell cycle. Biochim Biophys Acta 1495: 51-68. Furusawa T, Yanai N, Hara T, Miyajima A, Obinata M (1998) Integrin-associated protein (IAP, also termed CD47) is involved in stroma-supported erythropoiesis. J. Biochem. 123: 101-106..
(11) Gomez-Cuadrado A, Martin M, Noel M, Ruiz-Carrillo A (1995) Initiation binding repressor, a factor that binds to the transcription initiation site of the histone h5 gene, is a glycosylated member of a family of cell growth regulators [corrected]. Mol Cell Biol 15:6670-85. Huang AM, Wang HL, Tang YP, Lee EH. (1998) Expression of integrin-associated protein gene associated with memory formation in rats. J Neurosci 18: 4305-4317. Komuro H and Yacubova E (2003) Recent advances in cerebellar granular cell migration. Cell. Mol. Life Sci 60: 1084-1098. Miyashita M, Ohnishi H, Okazawa H, Tomonaga H, Hayashi A, Fujimoto TT, Furuya N, Matozaki T (2004) Promotion of neurite and filopodium formation by CD47: roles of integrins, Rac, and Cdc42. Mol Biol Cell 15: 3950-3963. Numakawa T, Ishimoto T, Suzuki S, Numakawa Y, Adachi N, Matsumoto T, Yokomaku D, Koshimizu H, Fujimori KE, Hashimoto R, Taguchi T, Kunugi H (2004) Neuronal roles of the integrin-associated protein (IAP/CD47) in developing cortical neurons. J Bio Chem 279: 43245-43253. Ohnishi H, Kaneko Y, Okazawa H, Miyashita M, Sato R, Hayashi A, Tada K, Nagata S, Takahashi M, Matozaki T (2005) Differential localization of Src homology 2 domain-containing protein tyrosine phosphatase substrate-1 and CD47 and its molecular mechanisms in cultured hippocampal neurons. J Neurosci 25: 2702-2711. Parkos, C.A., Colgan, S.P., Liang, T.W., Nusrat, A., Bacarra, A.E., Carnes, D.K., and Madara, J.L. (1996) CD47 mediates post-adhesive events required for neutrophil migration across polarized intestinal epithelia. J. Cell Biol. 132: 437–450. Pickford LB, Mayer DN, Bolin LM, Rouse RV. (1989) Transiently expressed, neural-specific molecule associated with premigratory granule cells in postnatal mouse cerebellum. J Neurocytol 18:465-78. Reinhold, M.I., Lindberg, F.P., Kersh, G.J., Allen, P.M., and Brown, E.J. (1997) Costimulation of T cell activation by integrin-associated protein (CD47) is an adhesion-dependent, CD28-independent signaling pathway. J. Exp. Med. 185:1–11. Scarpulla RC (2002) Nuclear activators and coactivators in mammalian mitochondrial biogenesis. Biochim Biophys Acta 1576: 1-14. Virbasius, C. A., Virbasius, J. V., and Scarpulla, R. C. (1993) NRF-1, an activator involved in nuclear-mitochondrial interactions, utilizes a new DNA-binding domain conserved in a family of developmental regulators. Genes Dev. 7: 2431-2445..
(12) 計畫成果自評: 此計畫原本申請為兩年期,審查通過為一年期,所以執行第一年的計畫,主要探討 α-Pal/NRF-1 及 IAP 在小鼠小腦發育過程的表現及可能功能。四個研究目的結果均已達成。 針對第一個目的,我們看到在胚胎第 15 天,IAP mRNA 廣泛分布在腦中,隨後在發育過程 分佈至特定腦區,包括端腦 (telencephalon)、間腦 (diencephalon),和菱腦唇 (rhombic lip)。 針對第二個目的,我們看到在出生後時期,IAP mRNA 表現量明顯的變化發生在發育中的 小腦。從出生後第 6 至第 14 天,IAP mRNA 表現隨著顆粒前驅細胞 (granular cell precursors, GCPs)由 external granular layer (EGL) 遷移至 internal granular layer (IGL)其表現量逐漸增 加。免疫組織染色法顯示 IAP 蛋白表現隨著小腦成熟,在分子層 (molecular layer)與 IGL 表現量亦逐漸上升。另外在 EGL 中,顆粒前驅細胞同時表現 IAP 與 TAG-1,顯示 IAP 參 與在顆粒前驅細胞遷移過程中。 針對第三個目的,我們發現 NRF-1 與 IAP 啟動子結合的活性隨著小腦發育有所變化, 與 IAP 基因表現一致。針對第四個目的,我們發現調控 NRF-1 與 IAP 對小腦神經細胞神經 突生長有顯著的影響。過度表現 NRF-1 減少了神經突的長度,而抑制型 NRF-1 則降低其長 度及數目。IAP form 4 cDNA 抑制了神經突伸長,然而 antisense IAP cDNA 減少了神經突數 目。這些結果顯示 NRF-1 及其下游基因 IAP 對小腦的發育及其神經細胞的分化扮演了重要 的角色。 整體而言,本計畫執行結果符合原先第一年所提的計畫內容。本研究部分成果發表在 第 21 屆生物醫學聯合年會. Reference: Hsien C.P. and Huang, A.M. (2006) Role of the transcription factor NRF-1 in the cerebellar development and neuronal differentiation. The Twentifirst Joint Annual Conference of Biomedical Sciences. Taipei, Taiwan..
(13) Fig. 1. Distribution of IAP mRNA during development investigated by in situ hybridizaiton. 35S-labeled antisense IAP cRNA was used as probes. Ultracryomicrotome sections (10 m) from the mouse whole embryos at embryonic day 15 (E15) (A), E17(B), E19(C), and brain tissues at postnatal day 1 (P1) (D), P7(E), P14(F), P21 (G), P28 (H), and adult (about 18 weeks) (I) were hybridized with probe at 55℃ for 16 hr. The signals were captured by the X-ray film for 3 days. The sense IAP cRNA probe hybridized with the adult brain section was taken as the control (J). (4V: 4th ventricle; CB: cerebellum; CX: cortex; Dien: diencenphalon; EGL: external granular layer; Hi: hippocampus; IGL: internal granular layer; LV: lateral ventricle; RL: rhombic lip; Th: thalamus; VZ: ventricular zone).
(14)
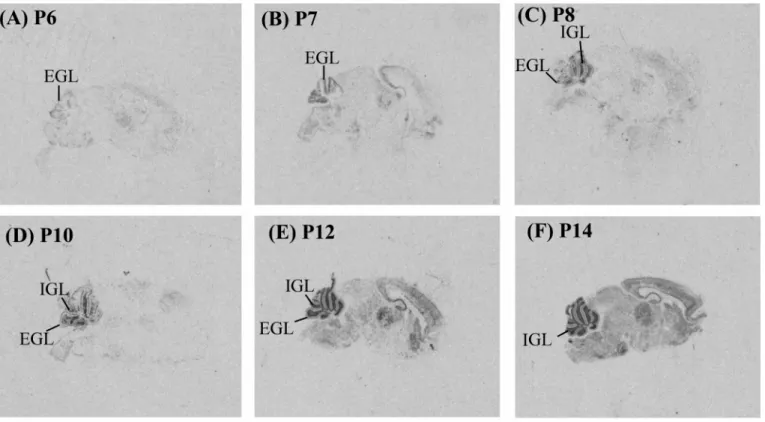
(15) Fig. 2. Distribution of IAP mRNA during cerebellar development investigated by in situ hybridization. Sagittal sections of the mouse brain from P6 to P14 were examined. In the cerebellum, IAP mRNA signal was appeared in the external granular layer (EGL) at P6 (A) and P7 (B). It became weaker in the EGL from P8 to P14 (C to F) and disappeared at P14 (F). IAP mRNA in the internal granular layer (IGL) was appeared at P8 (C) and became stronger from P8 to P14 (C to F)..
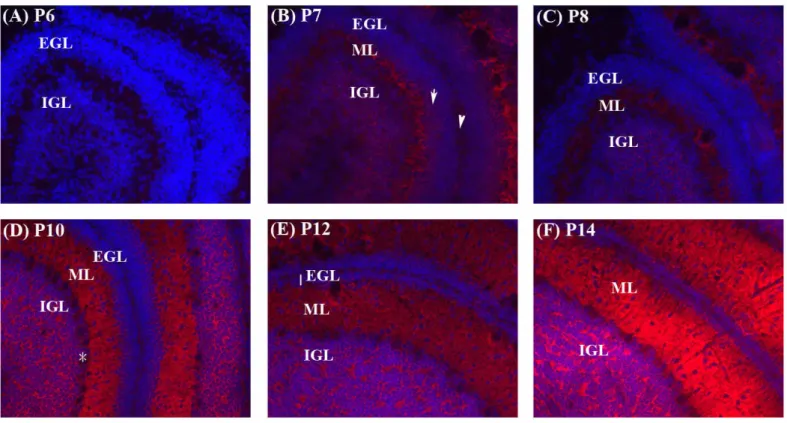
(16) Fig. 3. Distribution of the IAP protein during cerebellar development examined by immunohistochemistry. Sagittal sections (10 μm) of brains from P6 to P14 (A to E) were immunostained with the IAP antibody, mIAP301 (red). Hoechst33258 (blue) was used to identify the nuclei. EGL, external granular layer (arrow); IGL, internal granular layer; ML, molecular layer. The pia is indicated by arrowhead and the Purkinje cell layer was marked by star..
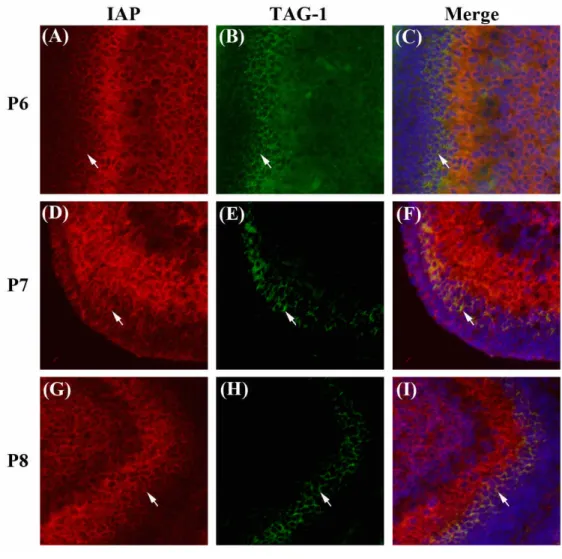
(17) Fig. 4. Expression of IAP and TAG-1 during cerebellar development analyzed by immuohistochemistry. Cerebellum from P6 (A to C), P7 (D to F), and P8 (G to H) were both immunostained with antibody for IAP (red; A, D, G), TAG-1 (green; B, E, H), and Hoechst33258 (blue, C, F, I). TAG-1 is a marker of migrating granular cells in the inner layer of EGL (iEGL)..
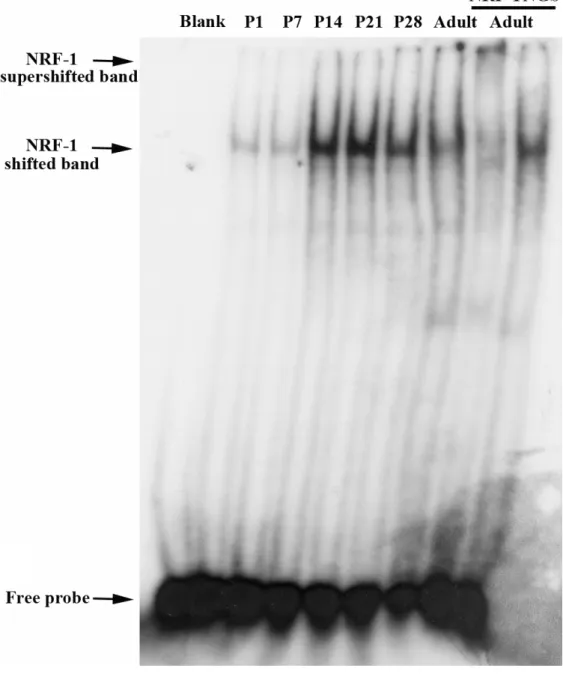
(18) Fig. 5. Electrophoretic mobility shift analysis of the NRF-1 binding activity during cerebellar development. Band shift analysis of the NRF-1 binding activity. Three g of nuclear extracts from cerebellum (CB) at postnatal 1, 7, 14, 21, 28 (P1 to P28) and adult (about 18 weeks) were incubated with 32P-labeled IAP oligonucleotides containing the NRF-1 binding element. Mixtures containing no nuclear extract were used as negative controls (Blank). One l of anti-NRF-1 antiserum were incubated with nuclear extracts for 3 hr prior to the addition of IAP promoter probes for the analysis of the supershifted band. Normal goat serum (NGS) was added as the control. Results showed that the binding activity of NRF-1 increased from P1 to P21, and decreased at P28 and the adult..
(19) Fig. 6. NRF-1 regulated neurite outgrowth in primary cerebellar cells. Cells cultured f r om P3mous ec e r e be l l um we r et r a ns f e c t e dwi t h1μgofe mpt yve c t or ,ve c t orc ont a i n i ng full-length, or dominant negative NRF-1 (NRF-1-DN) cDNA for 12 hr. Cells were fixed and immunostained with antibodies against GFP (green) and neuronal marker MAP2a,b (red) on the following 48 hr. Example of morphology of GFP and MAP2 positive primary cerebellar neurons (A). Left, GFP; middle, MAP2a,b;r i g ht ,me r g e di ma ge s .Ba rs c a l e ,25μm.The percentage of neuron containing different numbers of neurite (B), the length of longest (C) and other neurites (D) of GFP- and MAP2 positive cells were calculated from neurite-bearing GFP-positive cells (~ 100 cells) (n = 3). (*p < 0.05; **p < 0.01; ***P<0.001, unpaired t test.).
(20) Fig. 6. IAP regulated neurite outgrowth in primary cerebellar cells. Cells cultured from P3 mous ec e r e be l l um we r et r a ns f e c t e dwi t h1μgofe mpt yve c t or ,vector containing full-length IAP form2 and form4 cDNA, and antisense cDNA for 12 hr. Cells were fixed and immunostained with antibodies against GFP (green) and neuronal marker MAP2 (red) on the following 48 hr. Example of morphology of GFP and MAP2 positive primary cerebellar granular cells ( A) .Le f t ,GFP;mi ddl e ,MAP2;r i g ht ,me r g e di ma ge s .Ba rs c a l e ,25μm.The percentage of neuron containing neurite numbers (B), the average length of total neurites (C) of GFP- and MAP2 positive cells were calculated by the neurite-bearing GFP-positive cells in about 100 cells (n = 3). *p < 0.05; **p < 0.01; ***P<0.001, unpaired t test..
(21)
數據

+4
相關文件
For academic implementation, the casual relationships of perceived service quality, consumption emotion, perceived value, customer satisfaction, customer complaint
An extremely simple examination paper might generates learner who is high achievement of testing falls out of learning interest, and an extremely difficult examination paper
manufacturing operation in the past and no direct link with customer to get continuous feedback.. Although design chain as well as customer chain has been suggested in recent years
Because there is less information production produced in auctions, the information production theory predicts that auctions in IPOs would have higher volatility and less
The International Conference on Innovation and Management 2012 (IAM 2012) is an annual conference on Innovation and management since 1999, organized and sponsored by the
The core of this research is focusing on Service Innovation, discussing on the composite factors of manufacturing servitization through literatures and questionnaires from
Thus, this study argued the role and function of KIBS through evolution of local innovation system first; and then, this study analyzed interaction between technical firms and
具二年以上中子研究經驗;執行中子散 射、繞射以及台澳中子計畫 SIKA 實驗設施 運轉計畫。2.凝態物理系所博士畢業,具良
