國 立 成 功 大 學
化 學 研 究 所
碩 士 論 文
餵食半乳糖大白鼠水晶體蛋白醣化位置之研究
An in-vivo Study For The Site Specific Glycation
of Galactosemic Rat Lens Crystallins
研 究 生:廖 郁 儒
指導教授:黃 福 永
中 華 民 國 一 百 零 二 年 六 月
中文摘要
本實驗利用蛋白質體學的分析技術對餵食半乳糖誘使產生白內
障的大白鼠之水晶體作研究,得到結果如下:由快速蛋白質液相層析 儀分析水晶體內的水溶性水晶體蛋白,發現隨著得到白內障的時間增 長大白鼠水晶體內的 β 水晶體蛋白和 γ 水晶體蛋白明顯的減少。另 外,尿素可溶性蛋白中主要的成份β-A3、β-B1 和 β-B3 的多胜肽片段 出現被修飾的情形,如組胺酸(Histidine)、色胺酸(Tryptophan)、甲硫 胺酸(Methionine)被氧化(Oxidation)、丙胺酸(Alanine)接有乙醯基、親 水性的半胱胺酸(Cysteine)出現 Oxalate Monoalkylamide,而水溶性蛋 白成分中的β-A3、β-B1 和 β-B3 多肽片段也有被氧化、乙醯化,但沒 有出現 Oxalate Monoalkylamide 的修飾,這個差異可能就是造成 β-A3、β-B1 和 β-B3 水晶體蛋白從水溶性蛋白質變化成非水溶性蛋白
質的原因之一,而這些蛋白質受到的修飾可能讓水晶體蛋白聚集形成 非水溶性蛋白質造成水晶體混濁。
關鍵字:大白鼠水晶體蛋白,半乳糖血症,白內障,轉譯後修飾 糖基化
Abstract
Both proteomics and LC-MS/MS have been used to intensively investigate the protein post translation. In this study, two-dimensional gel electrophoresis followed by LC-MS/MS analysis have been employed to study the protein glycation of galactosemic cataract rat lenses. It was found that after feeding galactose, the rat lens started to show cortical nucleus cataract at week three. And the water soluble fraction of the lens showed that the contents of βH and γ crystallins decrease significantly with the increase of feeding time. It was also found that β-A3、β-B1 and β-B3crystallin in urea soluble fraction of the cataract lens had undergone oxidative modification at Histidine, Trptopha, and Methionien residues, acetylation at Alanine residue, and oxalate monoalkylamide modification at Cysteine residue. Interestingly, the water soluble fraction of cataract rat lens also underwent oxidation and acetylation; however, no oxalate monoalkylamide modification was observed for β-A3、β-B1 and β-B3 crystallin. The difference in oxalate monoalkylamide modification in water soluble fraction may suggest that this modification is the main cause to cause the denaturation of rat lens crystallins as to cause the formation of cataract.
Key Words: Rat lens crystallins, Galatosemic, Cataract, post translation, Glycation
誌
謝
我終於完成論文取得碩士學位了!!能夠在充滿文化氣息、歷史
悠久的成功大學裡作研究,並且實現人生夢想之一,實在是一件相當 幸運、令人稱羨的事!感謝恩師 黃福永博士在學業上的細心指導與 平日的關心照顧,讓我能夠順利完成學業,師恩浩蕩,永存心頭。
化學所 蕭世裕博士、嘉義大學 張文興博士對本文的肯定,以及
在口試期間提出許多寶貴意見使本文的內容更加充實,在此由衷感謝 兩位老師的指導。
感謝實驗室好友詣婷、杜杜、雅雅、邱張、王品、老邱、黑人,
大家不僅是平日一起作實驗、出遊的夥伴也是和我一起分享研究生活 的喜怒哀樂的好朋友,感謝你們,有你們真好!以及感謝醫學院的助 理園堤、學長捷修,謝謝園堤借我使用儀器和軟體,也謝謝學長捷修 給我在實驗上的寶貴意見。謝謝好友馮姐、育德、許樂、蒼哥、阿龍、
政志、金門、浩羽、龍哥、TONY、阿寬、稚展、任良、學長啟敬、
學弟育群、子賢、學妹文馨,伴我渡過充實的兩年時光。
特別感謝我最親愛的爸爸、媽媽,感謝你們支持我想念化學系的
夢想,因為你們的努力辛勞我才能無憂無慮地繼續念書,無須煩惱生 活,因為有了你們的用心栽培才有今天的我,在此把這小小的研究成 果獻上。謝謝爸爸、媽媽、蔚綸、鎮、鎧的支持和關心,我愛你們!
目錄
頁次 中文摘要 ... III 英文摘要 ... IV 誌 謝 ... V 表目錄 ... IX 圖目錄 ... XII
第一章 緒論
1-1 眼球構造的介紹 ... 1
1-2 水晶體的介紹 ... 4
1-2-1 水晶體的位置 ... 4
1-2-2 水晶體的形成與組成 ... 4
1-3 水晶體蛋白質的介紹………..…..5
1-4 水晶體蛋白的介紹 ... 7
1-4-1 alpha(α)水晶體蛋白 ... 7
1-4-2 beta(β)水晶體蛋白 ... 9
1-4-3 gamma(γ)水晶體蛋白... 11
頁次
1-5 白內障簡介 ... 13
1-5-1 白內障的形成原因 ... 13
1-5-2 糖基化的介紹 ... 14
1-5-3. 糖基化與白內障的關係 ... 15
1-6 研究目的 ... 19
第二章 實驗 2-1 材料 ... 20
2-2 實驗器材與儀器設備 ... 20
2-3 藥品 ... 21
2-4 水晶體蛋白的均質化 ... 23
2-4-1 萃取水溶性水晶體蛋白 ... 23
2-4-2 萃取尿素可溶性水晶體蛋白 ... 24
2-5 水晶體蛋白之濃度測定 ... 25
2-5-1 水溶性水晶體蛋白之濃度測定 ... 25
2-5-2 尿素可溶性水晶體蛋白之濃度測定 ... 27
2-6 水晶體蛋白之快速蛋白質液相層析 ... 28
2-7 SDS-PAGE 電泳分析 ... 30
2-8 二維電泳分析 ... 32
頁次
2-9 質譜分析 ... 35
2-9-1 In-gel Digestion ... 35
2-9-2 全蛋白質譜分析 ... 39
第三章 結論與結果 3-1 水晶體蛋白之快速蛋白質液相層析分析 ... 41
3-2 一維電泳之結果分析 ... 43
3-3 二維電泳之結果分析 ... 44
3-3-1 水溶性水晶體蛋白之二維電泳結果分析 ... 45
3-3-2 尿素可溶性水晶體蛋白之二維電泳結果分析... 47
3-4 質譜分析結果 ... 49
第四章 結論 ... 52
第五章 參考文獻...126
表目錄
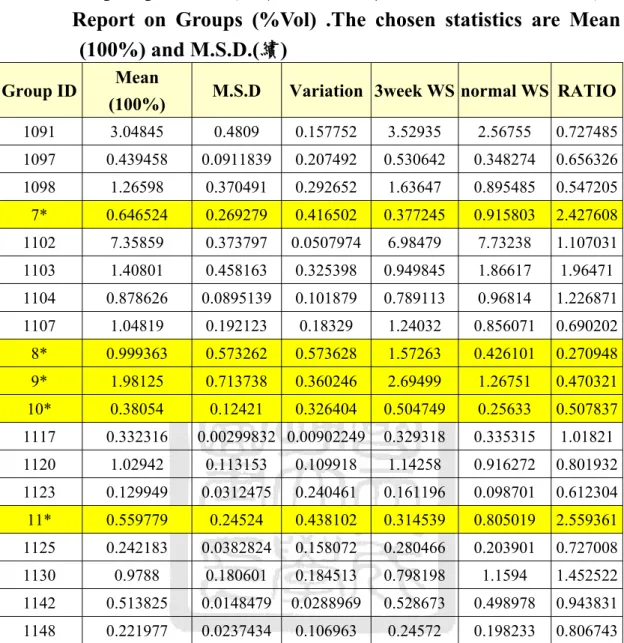
頁次 表一 BRADFORD蛋白質濃度測定之檢量線的配製 ... 26 表二 BRADFORD蛋白質濃度測定之檢量線的配製 ... 27 表三 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第一週):
REPORT ON GROUPS (%VOL).THE CHOSEN STATISTICS ARE MEAN
(100%) AND M.S.D ... 63 表四 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第二週):
REPORT ON GROUPS (%VOL).THE CHOSEN STATISTICS ARE MEAN
(100%) AND .S.D ... 68 表五 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第三週):
REPORT ON GROUPS (%VOL).THE CHOSEN STATISTICS ARE MEAN
(100%) AND .S.D ... 73 表六 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第四週):
REPORT ON GROUPS (%VOL).THE CHOSEN STATISTICS ARE MEAN
(100%) AND .S.D ... 78 表七 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第五週):
REPORT ON GROUPS (%VOL).THE CHOSEN STATISTICS ARE MEAN
(100%) AND .S.D ... 83 表八 GROUP REPORT分析(水溶性蛋白:控制組對照實驗組第六週):
REPORT ON GROUPS (%VOL) .THE CHOSEN STATISTICS ARE MEAN
(100%) AND .S.D ... 87
頁次 表九 Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第一 週):Report on Groups (%Vol) . The chosen statistics are Mean
(100%)andM.S.D ... 92
表十 Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第二 週):Report on Groups (%Vol) . The chosen statistics are Mean (100%)andM.S.D ... 98
表十一Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第三 週):Report on Groups (%Vol) . The chosen statistics are Mean (100%)andM.S.D ... 103
表十二Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第四 週):Report on Groups (%Vol) . The chosen statistics are Mean (100%)andM.S.D ... 109
表十三Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第五 週):Report on Groups (%Vol) . The chosen statistics are Mean (100%)andM.S.D ... .114
表十四Group Report 分析(尿素可溶性蛋白:控制組對照實驗組第六 週):Report on Groups (%Vol) . The chosen statistics are Mean (100%)andM.S.D ... 118
表十五 水溶性水晶體蛋白之全蛋白質譜分析(αA)...121
表十六 水溶性水晶體蛋白之全蛋白質譜分析(αB)...122
表十七 水溶性水晶體蛋白之全蛋白質譜分析(βB3)...122
頁次
表十八 水溶性水晶體蛋白之全蛋白質譜分析(βB1)...123
表十九 水溶性水晶體蛋白之全蛋白質譜分析(βA4)...123
表二十 水溶性水晶體蛋白之全蛋白質譜分析(βA3)...123
表二十一 尿素可溶性水晶體蛋白之全蛋白質譜分析(βB3)...123
表二十二 尿素可溶性水晶體蛋白之全蛋白質譜分析(βB1)...124
表二十三 尿素可溶性水晶體蛋白之全蛋白質譜分析(βA3)...124
表二十四 尿素可溶性水晶體蛋白之全蛋白質譜分析(γD)...125
表二十五 尿素可溶性水晶體蛋白之全蛋白質譜分析(γF)...125
圖目錄
頁次
圖一 人類的眼球構造 ... 3
圖二 Γ水晶體蛋白的三級結構 ... 12
圖三 蛋白質胺端與葡萄糖進行糖基化示意 ... 18
圖四 糖基化過程 ... 18
圖五 蛋白質濃度測定之檢量線 ... 26
圖六 蛋白質濃度測定之檢線 ... 28
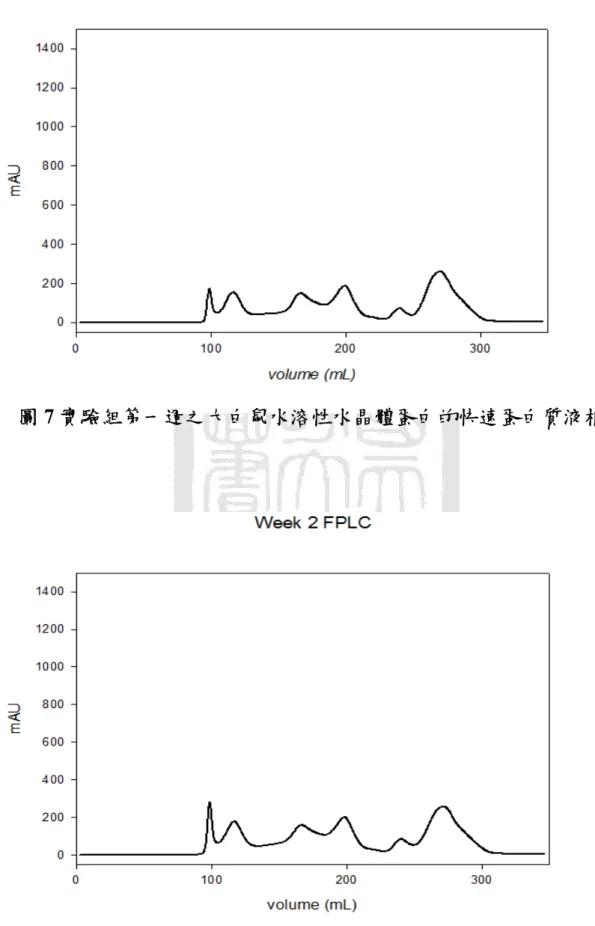
圖七 實驗組第一週之大白鼠水溶性水晶體蛋白的快速蛋白質液相 層析...55
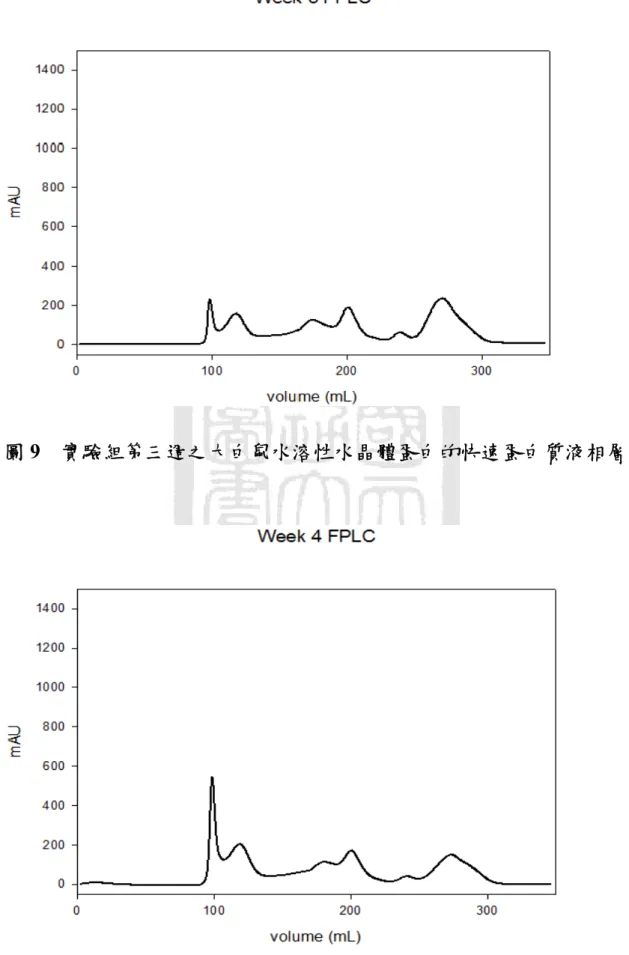
圖八 實驗組第二週之大白鼠水溶性水晶體蛋白的快速蛋白質液相 層析...55
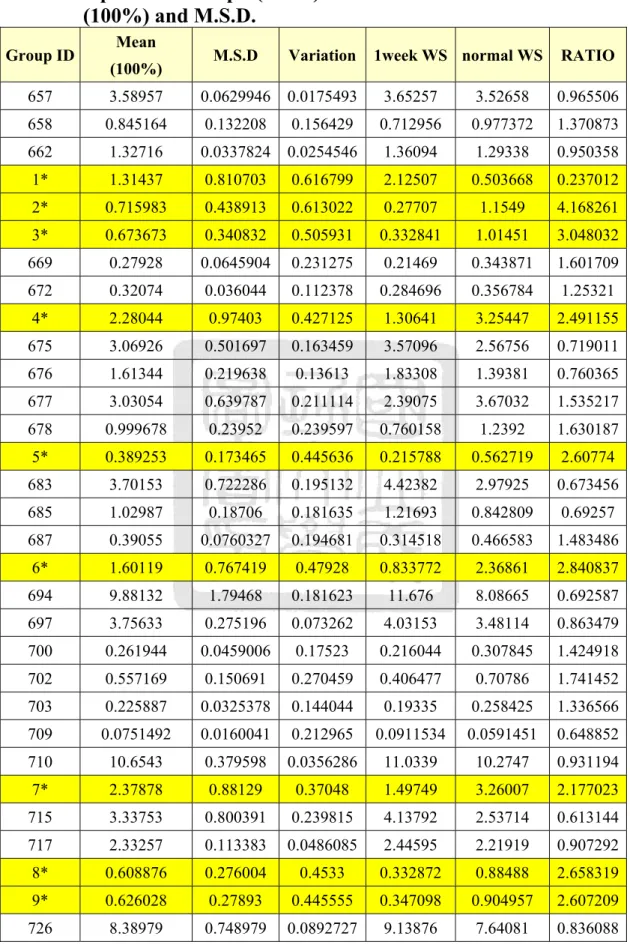
圖九 實驗組第三週之大白鼠水溶性水晶體蛋白的快速蛋白質液相 層析...56
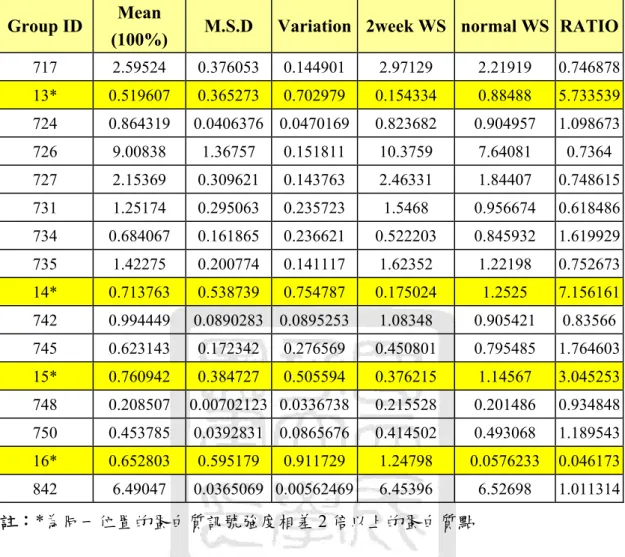
圖十 實驗組第四週之大白鼠水溶性水晶體蛋白的快速蛋白質液相 層析...56
圖十一 實驗組第五週之大白鼠水溶性水晶體蛋白的快速蛋白質液相 層析...57
頁次 圖十二 實驗組第六週之大白鼠水溶性水晶體蛋白的快速蛋白質液
相層析...57
圖十三 實驗組第一至第六週之大白鼠水溶性水晶體蛋白的快速蛋 白質液相層析的綜合比較...58 圖十四 大白鼠之水溶性水晶體蛋白 SDS 一維電泳分析...59 圖十五 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實驗組
第一週)...60 圖十六 大白鼠控制組與實驗組第一週水溶性水晶體蛋白之二維電 泳的結果比對圖...61
圖十七 大白鼠實驗組第一週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...62
圖十八 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實驗組 第二週)...65
圖十九 大白鼠控制組與實驗組第二週水溶性水晶體蛋白之二維電 泳的結果比對圖...66
圖二十 大白鼠實驗組第二週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...67 圖二十一 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實
頁次 驗組第三週)...70
圖二十二 大白鼠控制組與實驗組第三週水溶性水晶體蛋白之二維電 泳的結果比對圖...71
圖二十三 大白鼠實驗組第三週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...72 圖二十四 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實驗
組第四週)...75 圖二十五 大白鼠控制組與實驗組第四週水溶性水晶體蛋白之二維電 泳的結果比對圖...76
圖二十六 大白鼠實驗組第四週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...77 圖二十七 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實驗
組第五週)...80 圖二十八 大白鼠控制組與實驗組第五週水溶性水晶體蛋白之二維電 泳的結果比對圖...81
圖二十九 大白鼠實驗組第五週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...82
圖三十 大白鼠水溶性水晶體蛋白之二維電泳分析比較結果(實驗
頁次
組第六週)...84 圖三十一 大白鼠控制組與實驗組第六週水溶性水晶體蛋白之二維電 泳的結果比對圖...85
圖三十二 大白鼠實驗組第六週與控制組水溶性水晶體蛋白之二維電 泳分析結果比對圖...86
圖三十三 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果 (實驗組第一週)...89
圖三十四 大白鼠控制組與實驗組第一週尿素可溶性水晶體蛋白之二 維電泳的結果比對圖...90
圖三十五 大白鼠實驗組第一週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...91
圖三十六 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果 (實驗組第二週)...95
圖三十七 大白鼠控制組與實驗組第二週尿素可溶性水晶體蛋白之二 維電泳的結果比對圖...96
圖三十八 大白鼠實驗組第二週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...97 圖三十九 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果
頁次 (實驗組第三)...100
圖四十 大白鼠控制組與實驗組第三週尿素可溶性水晶體蛋白之二
維電泳的結果比對圖...101
圖四十一 大白鼠實驗組第三週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...102
圖四十二 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果 (實驗組第四週)...106
圖四十三 大白鼠控制組與實驗組第四週尿素可溶性水晶體蛋白之二 維電泳的結果比對圖...107
圖四十四 大白鼠實驗組第四週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...108
圖四十五 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果 (實驗組第五週)...111
圖四十六 大白鼠控制組與實驗組第五週尿素可溶性水晶體蛋白之二 維電泳的結果比對圖...112
圖四十七 大白鼠實驗組第五週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...113 圖四十八 大白鼠尿素可溶性水晶體蛋白之二維電泳分析比較結果
(實驗組第六週)...115 圖四十九 大白鼠控制組與實驗組第六週尿素可溶性水晶體蛋白之二 維電泳的結果比對圖...116 圖五十 大白鼠實驗組第六週與控制組尿素可溶性水晶體蛋白之二 維電泳分析結果比對圖...117
第一章 緒論
本研究第一章為緒論,共有六部分,分別為眼球構造的介紹、水 晶體的介紹、水晶體蛋白質的介紹、水晶體蛋白的介紹、白內障疾病 簡介,研究目的,分別敘述如下。
1-1 眼球構造的介紹
人類的眼球由眼球內容物以及眼球壁所構成,其中眼球壁可以分 成內、中、外共三層。外層:由前 1/6 透明的角膜和後 5/6 瓷白色的 鞏膜,共同構成眼球完整、封閉的外壁,起到保護眼內組織、維持眼 球形狀的作用,包含了角膜(Cornea)、鞏膜(Sclera)、前房腳(Anterior Chamber Angle)。角膜:位於眼球前極(Anterior Pole)中央,呈略向前 凸的透明橫橢圓形組織。由水及透明膠質(Collagen)構成,主要功能 為保護水晶體、接受外界光線,無血管分佈,提供三分之二的物體在 眼內成為影像所需的光度,透過淚液及房水獲取養份及氧氣並排除廢 物。 鞏膜:覆蓋眼後 80%的表面,為眼球壁最外一層,鞏膜可保護 眼球內部,並維持眼球的形狀。其質地堅韌,呈乳白色,主要由緻密 且相互交錯的膠原纖維組成。鞏膜幾乎無血管,但表層鞏膜有緻密的 血管結締組織。前房角:位於周邊角膜與虹膜根部的連接處。中層包 含了虹膜(Iris)、睫狀體(Ciliary Body)、 脈絡膜(Choroid)。虹膜:為
一圓盤狀膜,位於水晶體前方,中央形成瞳孔。虹膜自睫狀體伸展到 晶狀體前面,將眼球前部腔隙隔成前、後房,由括約肌(Sphincter Muscle)及擴張肌(Dilator Muscle)調控瞳孔的大小以便控制進入眼內 的光線,其中的色素含量可決定瞳孔顏色。睫狀體:主要由睫狀肌和 睫狀上皮細胞組成。睫狀肌由外側的縱行、中間的放射狀和內側的環 形三組肌纖維構成,而睫狀上皮細胞層由外層的色素上皮和內層的無 色素上皮二層細胞所組成。睫狀體位於虹膜與脈絡膜之間,經由懸小 帶調節水晶體曲度及由睫狀突可分泌水樣液體即為房水。房水可營養 角膜,並維持眼內壓。脈絡膜:介於視網膜與鞏膜之間,與鞏膜粘著 緊密。內層:為視網膜(Retina),位於眼球壁最內層,佈滿感光細胞
及神經纖維,其血液由脈絡膜及網膜小動脈供應。視網膜中心區域,
稱為“黃斑部",是一層透明的膜,位於脈絡膜的內側。
眼球內容包含房水(Aqueous Humor)、水晶體(Lens) 和玻璃體 (Vitreous Body) 三種透明的物質,是光線進入眼內到達視網膜的通
道。房水:為眼內透明液體,充滿前房(Anterior Chamber)與後房 (Posterior Chamber)。前房指角膜後面、與虹膜和瞳孔區晶狀體前面 之間的眼球內腔,後房為虹膜後面、睫狀體內側、晶狀體懸韌帶前面 和晶狀體前側面的環形間隙。水晶體:水晶體位於虹膜與玻璃體之 間,是一個大約 9 毫米直徑,4 毫米厚度的雙凸型組織,由水晶體懸
韌帶與睫狀體聯繫固定。形狀像凸透鏡,可以折射光線,經過水晶體 折射後的影像會投映在視網膜上,就像照相機的鏡頭使光線聚焦在底 片一樣(元新眼科, 2007)。
水晶體的厚度會隨年齡增長而緩慢增加,前晶體上皮細胞不斷增 生(Harding & Dilley et al., 1976),並且分化成水晶體纖維細胞。水晶 體纖維細胞不斷生長、延長且合成大量的水晶體蛋白(Mörner, 1894),
並將舊的纖維擠向中心,逐漸硬化而形成水晶體,核水晶體核外較新 的纖維稱為水晶體皮質,水晶體富有彈性,隨年齡增長水晶體核逐漸 濃縮、增大,彈性逐漸減弱,玻璃體:為透明的膠質體,充滿於玻璃 體腔內,占眼球內容積的4/5 (如圖 1 所示)。
圖 1 人類的眼球構造圖
資料來源:“中華民國視網膜色素病變協會" (2007)
1-2 水晶體的介紹
1-2-1 水晶體的位置
水晶體(Lens)是眼球(Eyeball)的主要曲光(Refraction)結構,也是 唯一有調節能力的曲光間質;形狀為一個雙凸形的凸透鏡似的構造。
水晶體位在角膜與虹膜之後、玻璃體與視網膜之前,其周緣部被環狀 的水晶體韌帶(Lens Ligament, Zonule)繫於周圍的睫狀體,懸吊於前玻 璃體(Anterior Virtreous Humor)與虹膜之間,以固定位置;使其不會隨 著眼球的轉動而移位,並且讓入射光適當的呈像在視網膜上。
1-2-2 水晶體的形成與組成
水晶體為透明、略帶一點點淡黃色的構造,成人水晶體的前凸曲
率半徑為 10mm,後凸曲率半徑為 6mm,厚度約為 5mm,直徑約為 9-10mm。是 一個雙凸形的透明構造。水晶體內沒有神經、也缺 乏 血 管,水 晶 體 細 胞 所 需 的 養 份 與 氧 氣,則 是 經 由 周 圍 液 體 的 擴 散 作 用來 供 應。 水晶體終其一生都在生長(Berman, 1991),它的 大小、重量會隨著年紀越大而增加,水 晶 體 細 胞 在 成 長 、 進 行 細 胞 分 裂 的 過 程 中,會 持 續 往 水 晶 體 的 中 央 移 動,為 了 要 達 成 全 部 透 明,水 晶 體 細 胞 在 分 裂 成 長 的 過 程 中 啟 動 了 自 我 摧 毀 的 程 式 , 消 除 各 種 胞 器 如 細 胞 核 、 粒 線 體 、 高 基 式 體 與 內 質 網 等 ,
最 後 僅 剩 下 細 胞 質,即 含 有「 晶 狀 蛋 白 」(Crystalline)的濃稠溶 液 。
然 而 , 與 一 般 「 細 胞 凋 亡 」 不 同 的 是 , 一 般 細 胞 死 亡 的 自 我 摧 毀 程 式 一 旦 發 生,最 終 目 標 是 將 細 胞 徹 底 摧 毀 成 碎 片;但 水 晶 體 細 胞 的 程 式 卻 不 一 樣,最 後 將 留 下 完 整 的 細 胞 膜、還 保 有 蛋 白 質 構 成 的 細 胞 骨 架 , 而 能 承 擔 水 晶 體 細 胞 應 有 的 工 作 。 眼球中的水晶體,是 人體唯一的透明組織。過去幾年,專 家 已 確 知 聚 光 所 必 需 的 透 明 度,大 部 份 源 於 水 晶 體 細 胞 的 獨 特 能 力,它 們 可 在 發 育 即 將 完 成 時 啟 動 自 我 摧 毀 程 式,變 成 中 空 但 能 續 存 的 細 胞,能 讓 可 見 光 通 過(張孟媛 與 侯育致, 2012)。
1-3 水晶體蛋白質的介紹
在水晶體細胞發育成熟的過程中,逐漸消除了細胞內胞器,最
後 僅 剩 下 細 胞 質 、 完 整 的 細 胞 膜 、 還 有 蛋 白 質 構 成 的 細 胞 骨 架。因 此 水 晶 體 絕 大 部 分 都 是 由 蛋 白 質 所 組 成,且 水 晶 體 的 蛋 白 質 含 量 佔 了 約 35%,比 其它的身體組織所含的蛋白質含量都 還 要 更 高(Kamp&Hoenders, 1973)。
組 成 水 晶 體 的 蛋 白 質 包 含 了 細 胞 骨 架 蛋 白 (Cytoskeletal Protein) 、 細 胞 膜 蛋 白 (Membrane Protein) 、 和 水 晶 體 蛋 白
(Crystallin),其中超過 90%的蛋白質組成是屬於水晶體蛋白,
而 細 胞 骨 架 蛋 白 和 細 胞 膜 蛋 白 占 水 晶 體 內 所 有 蛋 白 質 總 量 的 10% , 因 此 水 晶 體 蛋 白 是 構 成 水 晶 體 的 主 要 蛋 白 質 (Kopylova et al, 2011)。
而 水 晶 體 的 組 成 蛋 白 質 依 照 性 質 的 不 同 可 以 分 成 三 大 類 :(1)水 溶 性 蛋 白 質 (Water-Soluble Fraction), 指 的 是 蛋 白 質 中 能 夠 使 用 水 溶 性 緩 衝 溶 液 或 是 水 作 萃 取 得 到,水 溶 性 蛋 白 質 約 佔 水 晶 體 所 有 蛋 白 質 的 90%以上,主 要是由水晶體蛋白所構 成,(2)尿素可溶性蛋白質 (Urea-Soluble Fraction),無法用水或 水 溶 性 緩 衝 溶 液 萃 取 的 蛋 白 質 。 非 水 溶 性 部 份 可 以 使 用 6-8M 高 濃 度 尿 素 或 是 鹽 酸胍(Guanidine-HCl)萃 取 得 到 , 約 佔 水 晶 體 內 所 有 蛋 白 質 含 量 的 5%,大部份為細胞骨架蛋白、細胞膜蛋 白 ,(3)介 面 活 性 劑 可 溶 性 部 份 (Detergent-Soluble), 為 無 法 溶 解 於 水 和 尿 素 的 蛋 白 質 , 約 佔 水 晶 體 所 有 蛋 白 質 含 量 的 2% , 主 要 由 細 胞 膜 蛋 白 構 成。其 餘 的 就 是 少 數 無 法 被 溶 解 的 酵 素 和 一 些 與 水 晶 體 蛋 白 以 非 共 價 鍵 結 作 用 力 結 合 在 一 起 的 細 胞 膜 蛋 白(Horwitz, 1992)。
1-4 水晶體蛋白的介紹
哺乳類動物(Mammalian)的水晶體蛋白質的主要成份為水溶性水 晶體蛋白,而其種類又可以分成α、β、γ-三大類(Van Der Ouderaaw, de Jong and Bloemendal, 1977)、且每個家族內的水晶體蛋白之間具有高 度的同源性(Homology)(Kopylova et al., 2011)。另外除了分成三大 類之外,又可以再細分成αA、αB,βA1到βA4和βB1到βB3,γA到γF 還有γS(Jungblut et al., 1998)。
1-4-1 Alpha(α)水晶體蛋白
α 水晶體蛋白占全部水晶體蛋白含量的 40%,是脊椎動物 (Vertebrate)水晶體內主要的蛋白質成份,高濃度的 α 水晶體蛋白基質 (Mmatrix)也被認為是能讓水晶體具有透明度和折射性質的主要原因 (Rao et al., 1993)。α 水晶體蛋白包含了兩類單體,分別是 αA 水晶體
蛋白和αB 水晶體蛋白,莫耳比約為 3:1,分子量大約為 20kDa,且 αA 和 αB 水晶體蛋白兩者之間有 55%的基因同源性(Homologous Gene)(Ouderaa et al,. 1973),但是在蛋白質的二級結構和三級結構上 卻有所不同(Sun, Das & Liang, 1997)。α 水晶體蛋白在自然(Native)的 情況下為單體 αA 和 αB 水晶體蛋白以非共價作用力的方式聚集成巨 大的水溶性集結體,分子量範圍 300-1200kDa,平均分子量約為
800kDa(Horwitz,2003),代表了 α 水晶體蛋白可以由 15 至 60 個單體
所構成。因此,判定在自然狀態下 α 水晶體蛋白的實際分子量並不容 易,因為集結體的分子量會受到溫度、離子強度、pH 值和水晶體的 年紀…等因素影響而不同(Jong, Caspers & Leunissen, 1998; Rao et al., 1993;Horwitz, 2009)。
在功能方面,αA 水晶體蛋白和 αB 水晶體蛋白的氨基酸序列被 發現和一些小熱休克性蛋白質(Small Heat Shock Proteins, SHPSPs)有 40%的胺基酸序列同源性,特別是在 C 端的區域(Ingolia & Craig, 1982)。αB 水晶體蛋白在基因表現上可以被歸類為小熱休克性蛋白 質,之後又證實了αB 水晶體蛋白確實為小熱休克性蛋白質,此外研 究指出α 水晶體蛋白在環境改變的壓力之下,例如:溫度升高(Horwitz, 1992)、輻射(Borkman, Knight & Obi, 1996)、還原劑(Wang & Spector, 1995),幫助其他蛋白質防止受到外力影響而產生不正常的堆積 (Aggregate)、沉澱或是變性(Denature)的功能(Khanova et al., 2005),和
小熱休克性蛋白質的伴護活性有相似的作用,稱之為類伴護活性 (Chaperone-Like Activity),這代表了 α 水晶體蛋白能夠維持水晶體的 透明度的重要功能;也就是保護β 水晶體蛋白和 γ 水晶體蛋白避免因 環境壓力的影響而發生不正常的摺疊,進而生成集結、沉澱造成水晶 體的混濁(Wang et al., 2011)。但與分子伴護功能不同的是,α 水晶體
蛋白會與部分展開(Partial Unfolding)或是變性(Denature)的其它蛋白 質互相結合成複合物(Complex)的形式(Rao et al., 1993)。例如,α 水晶 體蛋白在加熱引發集結(Heat-induced Aggregation)的情況之下,會以 其疏水端(Hydrophobic)和變性的蛋白質交互作用,而且加熱 α 水晶體
蛋 白 則 此 能 力 會 增 加 , 在 60℃ 時 α 水 晶 體 蛋 白 會 作 Thermal transition,結果使部分未折疊的蛋白質的分子量和體積增加為兩倍,
但不會再繼續集結了(Khanova et al., 2005)。
另外,α 水晶體蛋白也存在於非水晶體的組織中,例如 αA 水晶
體蛋白除了主要在水晶體內,還可以在脾臟、胸腺中發現有其微量存 在(Srinivasan, Nagineni & Bhat, 1992);而 αB 水晶體蛋白被發現存在 於其他器官,如心臟、肺、腎臟、腦、骨骼肌之中(Bhat & Nagineni, 1989),可能是 α 水晶體蛋白具有生物上的重要功能,因此會存在於 這些組織中。
1-4-2 Beta(β)水晶體蛋白
β 水晶體蛋白是三大類水晶體蛋白中最複雜的一類,約占水晶體 蛋白總量的 35%,是由六種單體所組成的集結體,這些單體分別是 三個酸性βA1、βA3、βA4 以及三個鹼性 βB1、βB2、βB3 (Mach et al., 1989),酸性與鹼性單體之間的同源性相似度為 45%,酸性和鹼性單
體之中的同源相似度為 58%,而其聚集的形式和程度會受到溫度、
酸鹼等因素的不同而有不一樣的組合,其集結的方式可能是使用酸性 和鹼性各一個單體結合成二聚體後,再以此為架構做更高程度的聚集 (Bax et al., 1990)。β 水晶體蛋白之單體分子量約為 23-32 kDa,含有 205-242 個氨基酸殘基,而 β 水晶體蛋白的分子量範圍從 46-200kDa,
其中分子量較大的部分稱為βH水晶體蛋白,分子量約為210 kDa,而 分子量較小的部分稱作βL水晶體蛋白,分子量約為52 kDa。
β 水晶體蛋白和 γ 水晶體蛋白之間的同源性約為 25%(Berbers et al., 1984),其一級結構與二級結構與 γ 水晶體蛋白類似,但是 β 水晶
體 蛋 白 多 了 N- 端 延 伸 鏈 (N-terminal extension) 和 C- 端 延 伸 鏈 (C-terminal Extension),此點被認為是 β 水晶體蛋白集結時不可或缺 的因素(Wistow, Slingsby & Blundell, 1981)。
β 水晶體蛋白會隨著白內障的生成而發生沉澱,是水晶體蛋白中 尿素可溶解的主要成份,這可能是和β 水晶體蛋白的集結特質有關。
另外,在某些 β 水晶體蛋白的集結體中,曾發現有分子內雙硫鍵 (Intra-molecular Disulfide Bond)的存在,這可能是導致非水溶性蛋白 生成的主要原因(Mach et al., 1989)。
1-4-3 Gamma(γ)水晶體蛋白
γ 水晶體蛋白含量占水晶體蛋白的 25%,包含了七種單體,分別
為γA、 γB、 γC、γD、 γE、 γF、γS,分子量皆約為 20kDa,γ 水 晶體蛋白不以集結體的方式存在(Mach et al., 1990),因此 γ 水晶體蛋 白是分子量最小的水晶體蛋白。
γ 水晶體蛋白由 173-174 個胺基酸組成,其一級結構的胺基酸序
列為已知,二級結構為β-平板組成,三級結構由 X-ray 解得,主要是 由兩個區域(Domain)構成,分別是 N-末端區域和 C-末端區域,每一 個區域由兩區段(Motif)構成,而每一區段又稱作一迴紋鎖(Greek Key),由四條反平行的 β-平板所構成,其中一號、二號平板位在 N- 末端區域,三號、四號在C-末端區域,γ 水晶體蛋白的結構又可以描 述成二區域-四迴紋鎖(2-Domain 4-Greek key)結構(Norledge et al., 1997) 。在每一區段中有三條多肽鏈與另一個區段內的一條多肽鏈有
分子間氫鍵的交互作用,形成完整的平板結構,藉由氫鍵纏繞成安定 的三級結構(如圖 2 示)。
β 水晶體蛋白和 γ 水晶體蛋白兩者在單體之分子結構和基因序列上都 很相似,被認為是由同一始祖基因(Ancestral Gene)演化而來,因此被 歸類在同一個家族,稱作βγ 超級家族(βγ-superfamily)(Lubsen, Arts &
Schoenmakers, 1988)。考慮水晶體蛋白的二級結構和三級結構、電荷
的穩定性、硫氫基的位置分佈和分子間的作用力時,β 水晶體蛋白和 γ 水晶體蛋白具有相似性質;兩者皆具有高比例的 β-平板結構。但是 在水溶液中 β、γ 水晶體蛋白的自身的結合能力不一,使得 β 水晶體 蛋白為聚合的分子,而γ 水晶體蛋白以單體方式存在,雖然源自於同 一基因但在複製時產生了選擇性以及變異性(Arts et al., 1989)。γ 水晶
體 蛋 白 具 有 多 量 含 硫 殘 基 , 如 半 胱 胺 酸(Cysteine) 和 甲 硫 氨 酸 (Methionine) , 以 及 芳 香 族 殘 基 , 如 酪 胺 酸 (Tyrosine) 、 色 氨 酸 (Trytophan),而這些殘基很容易被氧化;如半胱胺酸(Cysteine)的硫氫 基氧化後會產生雙硫鍵使蛋白質產生集結;甲硫胺酸可能被氧化成硫 氧化物,這些殘基的變異很可能會影響到整個蛋白質的性質。許多關 於白內障的研究結果發現,在水晶體的中心有γ 水晶體蛋白的交聯聚 集(Cross-Linked Aggregation)情形產生,因此這些變異殘基被認為是 造成水晶體蛋白集結和形成白內障的原因之一(Arts et al., 1987)。
圖 2 γ水晶體蛋白的三級結構圖
1-5 白內障疾病簡介
1-5-1 白內障的形成原因
白內障是一種眼睛水晶體產生雲霧狀(Clouding)或是不透明 (Opacity)的疾病,這些不透明物體的出現原因為水晶體蛋白不正常的 堆積、聚集形成集結體(Clump),而阻擋了部份或全部的光線進入視 網膜,造成視覺模糊。
白內障的生成原因有很多種,其中包含了:(1)老化:老年退化 性眼疾,是最常見的白內障生成原因。隨著年齡的增長,水晶體老化 而失去彈性以及變得混濁,原因為水晶體內的水溶性水晶體蛋白會逐 漸減少而非水溶性水晶體蛋白會增加,並堆積在水晶體內,形成會使 光線散射的大分子,造成水晶體混濁;(2)遺傳:由於遺傳性、染色 體變異、胎內感染等原因所引發的白內障;(3)代謝:由於血液中糖 份上升,導致醛糖還原酶(Aldose Reductase)的活性增加,葡萄糖被 還原成使血液為高滲透壓的山梨醇(Sorbitol)並且在水晶體內堆積,造 成細胞內的環境為高滲透壓(Hypertonicity),使水分子向胞內流入,
造成纖維腫脹、變性最後產生混濁,此種白內障發生在老年,與老年 性白內障相似,發生在糖尿病人身上稱為糖尿病性白內障。這種高滲 透壓的失調在高血糖症(Hyperglycemia)和半乳糖血症(Galactosemia)
的併發症當中佔了重要的角色;(4)外傷:水晶體受傷使水晶體囊破
裂,房水由水晶體囊的裂縫進入水晶體,而水晶體在受傷部位混濁之 後,形成液泡、水腫影響到水晶體的周邊,最後導致整個水晶體的混 濁(楊宜涵, 2012);(5)藥物:長期使用含有類固醇的藥物所導致的 白內障;(6)併發性引發的白內障:因為眼睛內部的疾病所引發的白 內障;像是青光眼、眼內炎症、出血、因眼內手術引發的白內障;(7) 輻射:暴露在紫外光、短波長紅外線、微波、X 射線、γ 射線及高能 量的β 射線,照射水晶體之後所形成的白內障(Sankeshi et al., 2013)。
1-5-2 糖基化的介紹
糖基化(Glycation)為發生在生物體內的非酶催化反應,發生在含 有游離胺基的分子例如:胺基酸、胜肽(Peptides)、蛋白質、脂質、核 酸。作用方式為糖的醛(酮)基與蛋白質的游離胺基互相結合,形成穩 定、不易降解的糖基化最終產物(Advanced Glycation Endproducts;
AGEs)並且堆積在組織中,對生物產生不好的影響。糖基化是造成許 多老化疾病的主因,包含動脈硬化(Atherosclerosis)、心肌老化、白內 障、阿茲海默症(Alzheimer's Disease)、糖尿病併發症、和皮膚鬆弛…
等症狀。
1-5-3 糖基化與白內障的關係
葡 萄 糖 (Glucose) 是 新 陳 代 謝 中 主 要 的 糖 類 ; 單 糖 (Monosaccharides) 像 是 葡 萄 糖 (Glucose) 半 乳 糖 (Galactose) 果 糖 (Fructose)和甘露糖(Mannose)通常以兩種形式存在:開環(Open-Chain) 或是閉環形式(Closed Ring; A Haworth Projection Form)。 對於葡萄
糖、半乳糖、和甘露糖而言,開環的形式在第一個碳上會露出一個具 有反應性的醛基(Aldehyde Group)。在生理條件中,這些糖被歸類成 還原糖(Reducing Sugars)因此通常具有反應性,醛基與游離的胺端 (E-Amino Group)像是賴胺酸(Lysine)和精胺酸(Argine)的殘基特別有 反應性(如圖 3 所示)。
當還原糖的 CO 雙鍵和一個游離的胺基端作用,會先形成席夫鹼
(Schiff Base, SB)此步驟為可逆反應(Reversible)。席夫鹼接下來再進
行 自 發 性(Spontaneous) 的 重 排 , 稱 為 阿 瑪 多 里 重 排 (Amadori Rearrangement)形成較為安定的阿瑪多里產物(Amadori Product),為一 種糖-胺基複合物(Sugar-Amino Group Complex),此為生成糖基化最
終 產 物 的 中 間 物(Intermediate) 。 之 後 , 阿 瑪 多 里 產 物 會 作 脫 水
(Dehydration)和重排,最後生成互相交連的蛋白質(Kattaet al., 2009)。
還原糖為開環結構時才能生成席夫鹼,但葡萄糖在正常情況下只 有0.002%為開環結構;甘露糖 0.005%為開環結構;半乳糖開環結構
佔了全部的 0.02%;而果糖有 0.7%是開環結構。因此葡萄糖與其他 的糖類相較之下不容易和蛋白質作用,是比較“安全"的糖類。
糖基化作用可以發生在細胞內、外,蛋白質也能與不同的糖類進 行糖基化反應,而隨著與不同的糖類反應,重排的反應機構
(Mechanism)、產物形式也不盡相同。三種常見的糖基化最終產物的 形式如下:(1)具有螢光結構(Fluorescent Structure)且使 Lysine 與 Arginine 互相交連,如 Pentosidine。其生成來自於五碳醣(Pentose)或
是醣類碎片先與胺基酸殘基結合形成阿瑪多里產物,再進行美拉德反 應(Maillard Reaction)氧化所生成,此反應只發生在含有高濃度蛋白質 的組織當中。Pentosidine 被視為糖基化最終產物的一種,在患有白內 障的人類水晶體內發現含有高濃度的Pentosidine;(2)非熒光結構 ( Non-Fluorescent Structure )與氨基酸互相交連,生成如 Imidazole Ring 結構。Imidazole 環狀結構通常存在於組安酸(Histidine)上,但 Lysine
與Arginine 的殘基也能與甲基乙二醛(Methylglyoxal)自發反應產生 Imidazole Ring 破壞蛋白質的功能與結構。Arginine 的側鍊上有兩個 氮,能與甲基乙二醛形成Imidazole Ring,Arginine 與甲基乙二醛結 合的堆積物在老化的人類水晶體內的含量很高;而Lysine 的側鍊上 只有一個氮,因此要形成環狀架橋就需要兩個Lysine,但是一般蛋白 質當中Lysine 之間都相隔遙遠或是位在兩個完全不相關的蛋白質
上,因此由兩個 Lysine 產生的連結將會使得蛋白質結構扭曲或是蛋 白質被黏在另外一個蛋白質上,而無法進行正常的生理功能。這種交 纏在一起的蛋白質團也是糖基化最終產物的一種類型;(3)不具有交
互 連 結 的 集 結 物(Non-Cross-Linking Adduct) , 而 是 與 Lysine 或 Arginine 以共價鍵的方式直接連結,例如 Carboxymethyl Lysine;
CML。其生成方式是由葡萄糖與 Lysine 直接縮合、重排成阿瑪多里 產物,之後直接進行氧化反應所得到(如圖 4 所示)。
糖基化反應如果發生在水晶體蛋白上時,將會破壞水晶體蛋白的
結 構 , 使 得 原 本 穩 定 且 緊 密 排 列 的 水 晶 體 蛋 白 之 間 互 相 交 連 (Protein-Protein Crosslinks),生成非水溶性的聚集體,進而影響水晶 體的透明度、曲光性(Ulrich & Cerami, 2001),改變光線進入水晶體的 折射係數,造成光線散射(Kopylova et al., 2011)。
圖 3 蛋白質胺端與葡萄糖進行糖基化示意
圖 4 糖基化過程
(資料來源為 R&D System Technical Information) 葡萄糖
席夫鹼
亞胺正離子 1,2-烯胺醇
Amadori 重排產物
N-葡萄糖基胺
Amadori 重排反應
1-6 研究目的
本實驗以餵食含有 50%半乳糖的飼料的大白鼠作為實驗組,並
且與餵食正常飼料的大白鼠互相對照,以模擬體內含有高濃度糖分的 環境,誘使大白鼠得到白內障。因此本實驗欲探討產生白內障時水晶 體 內 的 水 晶 體 蛋 白 質 的 成 份 變 化 , 利 用 二 維 電 泳 分 析(Two- Dimensional Gel Electrophoresis, 2-DE)技術,分別對水溶性水晶體蛋 白、尿素可溶性蛋白進行蛋白質分離(Protein Separation),並使用 ImageMaster 比對軟體分別找出水溶性蛋白及尿素可溶性蛋白具有強 度差異的蛋白質點,以瞭解隨著白內障隨著時間增長變得越嚴重時,
水晶體內蛋白質成份的變化情形。再進一步作切膠挖點,經由液相層 析(Waters nanoACQUITY UPLC System)以及二維線性離子阱式傅立 葉轉換電場軌道多次質譜系統(LTQ Orbitrap XL, Thermo-Fisher) 分 析有差異的蛋白質點,以鑑定蛋白質的身份。
綜合上述實驗分析的結果,即可得知有哪些水晶體蛋白被糖基化
(Glycation)以及水晶體蛋白受到何種修飾(Modification)和發生修飾 的位置,則可進一步了解白內障的生成機制。
第二章 實驗
2-1 材料
離乳品系雌性大白鼠(Sprague-Dawley rats)
2-2 實驗器材與儀器設備
冷凍離心機(Centrifuge;Hettich Universal 16R rotor#1614)。
冷凍乾燥機(Freeze Dry System;LABCONCO LYPH LOCK 4.5)。
酸鹼度量計(PH Meter, ORION 420A;SUNTEX SP-701)。
超音波震盪洗淨器(Ultrasonic Cleaner;BRANSONIC B3200R-4)。
快速蛋白質液相層析系統(Fast Protein Liquid Chromatography, FPLC, AKTA basic 10 system, Pharmacia)。
膠體粒子過濾管柱(Gel Giltration Column, HiPrep 26/60 Sephacryl S-300 HR)。
自動收集器(GradFrac collector, Pharmacia;SF-2120, Advantec)。
紫外光/可見光光譜儀(UV/VIS Spectrophotometer;JASCO V-550)。
紫外光檢測器(Monitor UV-1, Pharmacia; UA-6 ISCO)。
透 析 膜(Dialysis Membrane ; SPECTRUM, Specra/Pro 1, MWCO 6-8kDa)。
直立式電泳儀(Vertical Electrophoresis System, Hoefer SE 600 Series
Standard Vertical Units;Amersham Pharmacia Biotch)。
電泳電源供應器(Elecrophoresis Power Supply,EPS 1001, Amersham Pharmacia Biotch)。
等電點電泳儀(IPGphor, Amersham Pharmacia Biotch):13 公分長等電 點電泳膠條陶瓷容槽(Strip Holder)。
四位天秤 (4-Decimal Balance, OHAUS Analytical Pls)。
奈升液相層析搭配二維線性離子阱式傅立葉轉換軌道多次系統(LTQ Orbitrap XL, Thermo-Fisher)
ImageMaster 2D Platinum Software 比對軟體(Version 5.0)
2-3 藥品
三羥甲基氨基甲烷(Trizma Base, J.T. Baker)。
乙二胺四乙酸(Ethylene Diamine Tetraacetic Acid, EDTA;99.5 %, Pharmacia)。
疊氮化鈉(Sodium azide;99.5 %, Aldrich)。
丙烯醯胺(Acrylamide, Pharmacia)。
亞甲基二丙醯烯胺(N,N'-bis-methylene acrylamide, Pharmacia)。
十二烷基磺酸鈉(Sodium Dodecyl Sulfate, SDS, Pharmacia)。
甘胺酸(Glycine, J.T. Baker)。
尿素(Uurea;99.5 %, J.T. Baker)。
碘乙醯氨(2-Iodoac Etamide;99 %,Sigma)。
二硫蘚糖醇(DL-dithiothreitol,DTT;99%, Sigma)。
碘乙酸胺(Iodoacetamide, IAA, Sigma)。
四 甲 基 乙 二 胺 (N,N,N',N'-Tetramethyl Ethylenediamine, TEMED, Pharmacia)。
過硫酸銨(Ammonium Persulfate, Pharmacia)。
三氯醋酸(Trichloroacetic Acid, TCA;95%, MERCK) 葛瑪茜藍(Coomassie Brilliant Blue R-250, TCI)。
乙醇(Ethanol;95 %, ACROS)。
冰醋酸(Acetic Acid;100 %, BDH)。
鹽酸(Hydrochloric acid;37 %, Riedel-de Haen)。
非離子形促溶劑(NP-40, Pharmacia)。
等電點電泳膠條(Immobiline Dry Strip pH 3-10, 13cm, Pharmacia)。
瓊脂糖凝膠(Agarose gel)。
電 泳 分 子 量 標 準 品(PageRuler Prestained Protein Ladder, Thermo Scientific)。
2-4 水晶體蛋白的均質化
2-4-1 萃取水溶性水晶體蛋白
水 晶 體 的 來 源 取 自 三 到 四 週 大 的 雌 性 離 乳 品 系 大 白 鼠 (Sprague-Dawley),分成控制組(Control Group)和實驗組(Treatment Group),分別餵食正常飼料和摻雜50%半乳糖(D-(+)-Galactose)的飼 料,每隔一週時間取出四隻大白鼠以乾冰(CO2)安樂死,並將其眼球 取出,使用生理食鹽水清洗後冷凍保存在-20℃冰箱中。將眼球解凍
後放入研磨缽(Handle Homogenizer)並加入少許緩衝溶液(0.05 M Tris-buffer;50 mM Tris-HCl,0.02 % NaN3,1 mM EDTA, pH 7.4)在4℃
之 下 與4 顆 大 白 鼠 眼 球 一 起 研 磨 , 將 其 徹 底 搗 碎 以 達 到 均 質 化 (Homogenized)的狀態。
收集均質化之後的溶液至1 .5 mL的微量離心管中,在4 ℃之下以 14000 rpm的轉速離心60分鐘,之後觀察微量離心管可以看到溶液分 成上、下兩層,以1 c.c.的附針注射針筒(Syringe with Needle)收集上層
的水溶性水晶體蛋白,通過孔徑為0.22 μm的注射筒專用過濾膜 (Syringe Filter)過濾蛋白質溶液。剩下在微量離心管內的沉澱物為非
水 溶 性 蛋 白 , 則 用 適 量 的Tris-buffer 潤 洗 之 , 以 微 量 離 心 管 杵 (Microcentrifuge Tube)再次均質化,在4 ℃之下離心60分鐘後收集上 層澄清液,重複此步驟約2-3次,最後把剩下的沉澱物保存在-20 ℃冰
箱中。收集全部的上層澄清蛋白質溶液,利用超濾離心管(Vivaspin 6, 3kDa)在4 ℃、轉速為4000rpm之條件下離心,直到蛋白質溶液濃縮至 體積約為1 mL,以Tris-buffer潤洗超濾離心管內壁1-2次,最後收集的 蛋白質濃溶液體積約為2 mL,保存於-20 ℃冰箱內。之後進行蛋白質 濃度的檢測,以定量蛋白質質量。
2-4-2 萃取尿素可溶性水晶體蛋白
萃 取 水 溶 性 水 晶 體 蛋 白 之 後 , 微 量 離 心 管 內 仍 有 不 溶 於 Tris-Buffer 的淡黃色沉澱物,製備以 Tris-Buffer 作為溶劑的 6 M 尿素 溶液,萃取之。加入適量的尿素溶液至微量離心管中,以微量離心管 杵研磨管內沉澱物至溶液呈現淡黃色,在4 ℃之下以 14000 rpm 的轉 速離心60 分鐘之後,以 1 c.c.附針注射針筒吸取上層澄清溶液,並通 過孔徑為0.22 μm 的注射筒專用過濾膜以過濾蛋白質溶液。剩下在微 量離心管內的沉澱物則用適量的尿素溶液潤洗,使用微量離心管杵再 次均質化,離心 60 分鐘後再次收集上層澄清液並且過濾,重複此步 驟約2-3 次,最後剩下在管內的沉澱物則保存在-20 ℃冰箱中。把收 集的上層澄清液利用超濾離心管在4 ℃,轉速 4000 rpm 之條件下離 心,濃縮蛋白質溶液至體積約為 1 mL,加入適量的 6M 尿素溶液潤 洗超濾離心管內壁 1-2 次,最後收集的蛋白質濃溶液總體積約為 1.5 mL,保存於-20 ℃冰箱中。之後進行蛋白質濃度的檢測,以定量蛋白
質質量。
2-5 水晶體蛋白的濃度測定
2-5-1 水溶性水晶體蛋白之濃度測定
以Bradford 方法測定蛋白質的濃度,使用濃度為 2 mg/mL 的牛血 清蛋白(Bovine Serum)加水配置成濃度為 0.1 μg/μL,把稀釋後的 0.1 μg/μL 牛血清蛋白溶液分別取體積為 0、10、30、60、90、100 μL,
個別加入體積為800、790、770、740、710、700 μL 的水中,以製備 蛋白質質量為 0、1、3、6、9、10 μg 的標準蛋白質溶液。而樣品的 配置是經過Tris-buffer 萃取所得到的正常眼球、實驗組之第一週到第 六週的病變眼球的蛋白質濃溶液,各取出 5μL 加到體積為 45μL Tris-buffer 之中使濃度稀釋 10 倍,取不同體積的稀釋後蛋白質溶液加 水使最後總體積為800mL,即完成樣品的配置。全部的溶液配置完成 後,在每管各別加入體積為200μL 的反應試劑,並且充分混合均勻,
避光靜置5-10 分鐘,待反應之後使用分光光度計(Spectro-Photometer) 在波長595 nm 之下檢測蛋白質溶液之吸收度,即可以得到一條檢量 線(如表 1 及圖 5 所示)。
表1 Bradford 蛋白質濃度測定之檢量線的配製 蛋白質質量
(μg)
BSA 體積
(0.1μg/μL)
H2O 體積
(μL)
反應染劑
(μL)
0 0 800 200 1 10 790 200 3 30 770 200 6 60 740 200 9 90 710 200 10 100 700 200
圖5 蛋白質濃度測定之檢量線圖
2-5-2 尿素可溶性水晶體蛋白之濃度測定
以 Bradford 方法測定蛋白質的濃度,使用濃度為 2 mg/mL 的牛 血清蛋白(Bovine Serum)加水配置成濃度為 0.1 μg/μL 的標準蛋白質溶
液,把稀釋後的0.1μg/μL 牛血清蛋白溶液分別取體積為 0、20、40、
60、80、100μL,各別加入體積為 400、380、360、340、320、300 μL
的水中,每管再另加體積為 400μL 的 6M 尿素溶液,使總體積為 800μL,製備成蛋白質質量為 0、2、4、6、8、10 μg 的標準蛋白質溶 液。而尿素可溶性蛋白質的樣品配置,方法為取出未稀釋過的尿素可 溶性蛋白質濃溶液1 μL 加上體積為 399 μL 的 6 M 尿素溶液,每管再 另加入400 μL 的水。待全部溶液都配置完成後,每管加入體積為 200 μL 的反應染劑,並且快速混合均勻,在避光條件之下等待反應時間
約5-10 分鐘。最後使用分光光度計(Spectrophotometer)在波長 595 nm 之下檢測蛋白質溶液之吸收度,即能得到一條檢量線(如表 2 及圖 6 所示)。
表2 Bradford 蛋白質濃度測定之檢量線的配製 蛋白質質量
(μg)
BSA 體積
(0.1μg/μL)
H2O 體積
(μL)
尿素溶液
(μL)
反應染劑
(μL)
0 0 400 400 200
2 20 380 400 200
4 40 360 400 200
6 60 340 400 200
8 80 320 400 200
10 100 300 400 200
2-6 水晶體蛋白之快速蛋白質液相層析
膠體粒子過濾管柱(Gel Filtration Column)可以依照分子量的大小 把不同蛋白質分離出來,經過沖堤液沖堤(Eluted),分子量較小的蛋 白質會在膠球中滯留時間久,而較慢被冲堤出來;分子量大的蛋白質 則是較快被沖出。因此,蛋白質樣品中的複雜蛋白質成份就能依照分 子大小,在不同的時間點被冲堤出來,利用這種原理就能把樣品中的 蛋白質作純化與分離,因此選擇適當的膠體粒子孔徑的管柱,就能對 不同分子量範圍的蛋白質有良好的分離效果。本實驗使用的儀器是快 速蛋白質液相層析系統(Fast Protein Liquid Chromatography, FPLC),
管柱為HiPrep 26/60 Sephacryl S-300 HR,屬於對高分子量分子靈敏的 圖 6 蛋白質濃度測定之檢量線圖
管柱。取質量為18500μg、總體積為 2 mL 的水溶性水晶體蛋白溶液,
以針筒緩慢地注入樣品,使用 Tris-Buffer 作為冲堤液,注意使用冲提 液之前必須先經過孔隙為 0.22 μm 的水溶液濾膜過濾,並且除氣 (Degas)一小時。實驗過程中流速為 1mL/min 作等梯度冲提,並以紫 外線偵測器(UV Detector)在波長為 280 nm 之下檢測蛋白質之分離情
形,冲提總體積為管柱體積的兩倍。由結果觀察可以見到出現不同的 波峰(Peak),即是代表不同的蛋白質,把這些不同波峰的蛋白質分別 收集起來,對二次水(De-Ionized Water, D.I. Water)在 4 ℃之下進行透 析(Dialysis)以去除鹽類物質。透析的換水時間間隔為 3 小時、6 小時、
隔夜(Overnight),每次透析使用約 2 L 的二次水。透析完之後收集蛋 白質溶液,將之冷凍乾燥 (Lyophilization),保存在-80 ℃的冰箱中以 待作進一步的分析。
2-7 SDS-PAGE 電泳分析
SDS-PAGE 電泳是目前最常使用來分離蛋白質的技術。其原理是
把SDS 引入聚丙烯醯胺凝膠系統中,介面活性劑 SDS 會附著在蛋白 質的表面上,並且與蛋白質依照重量的比例結合,因此蛋白質-SDS 複合物的表面上帶有大量的 SDS,使得蛋白質失去了原本的電荷狀 態,形成了帶負電荷的團塊。因為蛋白質本身所帶的電荷遠小於附著 在複合物表面上 SDS 分子的負電荷,這樣便能去除蛋白質因為本身 帶有不同電荷而造成在電場中移動速度差異的因素,變成只由蛋白質 本身的分子量大小作為決定在電場中移動速度的因素。如此,就能依 照分子量大小將蛋白質分離,並使用 Marker 來標示蛋白質分子量的 範圍。另外,膠片所含的丙烯醯胺濃度可以依照樣品分子量大小作調 整,本實驗使用的是濃度為12.5%的膠片。
實驗方法為把正常眼球以及實驗組第一到第六週的眼球之水溶 性水晶體蛋白濃溶液以 Tris-Buffer 稀釋 10 倍後,各取出蛋白質質量 為10、15μg 的量,且蛋白質稀釋液與樣品緩衝液(Sample Buffer)以 體積比為 3:1 的比例混合並且在 97℃之下加熱 10 分鐘使蛋白質變 性 。 配 置 7cm 、 12.5 % 十 二 烷 基 硫 酸 鈉 - 聚 丙 烯 醯 胺 凝 膠 電 泳 (SDS-PAGE)於固定電流下進行蛋白質分離,開始電泳後先設定電壓 為75V 並維持此電壓 30 分鐘,待樣品通過上膠進入下膠之後,調整
電 壓 為 100V , 兩 個 小 時 後 電 泳 結 束 並 取 出 膠 片 , 以 葛 瑪 茜 藍 (Coomassie Brilliant Blue )染色膠片 1 小時,最後把膠片浸泡在 Destain Buffer 中直到退染完成。
電泳所需要的溶液及配方如下:
(1) 10% APS (Ammonium Persulfate)
取 5 g APS 加入體積為 50 mL 的二次水中,待其溶解後以注射筒 專用過濾膜過濾。
(2) SDS-PAGE destaining solution
300mL 甲醇、100 mL 醋酸、與 600 mL 二次水混合均勻。
(3) SDS-PAGE 10X gel running buffer
秤取 60 g Tris-Base(248 Mm)與 288 g 甘胺酸(1.92 M)加二次水溶 解,再加入20 g SDS (1% w/v SDS)定量體積為 2000 mL。
不需要除氣及過濾,使用前須稀釋10 倍成 1X 再使用。
(4) SDS-PAGE Coomassie staining solution
1.25 g Comassie R-250 溶解於 225 mL 甲醇、225 mL 二次水、50 mL 冰醋酸中,須要過濾才能使用。
(5) SDS-PAGE gel making buffer (Separating Gel,1.5 M Tris-HCl)
取 18.17 g Tris-Base 和 0.4 g SDS 溶於 80 mL 的二次水,以鹽酸調
整pH 8.8,最後加入二次水使總體積為 100 mL。
(6) SDS-PAGE gel making buffer (Stacking Gel,0.5 M Tris-HCl)
取 6.06 g Tris-Base、0.4 g SDS 溶於 80 mL 的二次水,以鹽酸調 整pH 6.8,最後加入二次水使總體積為 100 mL。
(7) SDS-PAGE marker buffer
取 4.8 mL 二次水 、1.2 mL 1 M Tris-HCl (pH 6.8)、1 mL 100%
醋酸、2 mL 10% w/v SDS、0.5 mL 0.1% w/v bromophenol blue。
2-8 二維電泳分析
第一維電泳,又稱作等電聚焦(Lsoelectric Focusing,簡稱為 IEF)。
原理是蛋白質分子因其組成的胺基酸不同,本身會帶有不同的電荷,
而蛋白質分子所帶有的淨電荷(Net Charge)是取決於環境 pH 值;若環
境 pH 值高於蛋白質的等電點(PI),此蛋白質淨電荷為負電;環境的 pH 值低於其等電點則蛋白質淨電荷為正電;若剛好 pH 值等於其等 電點,則淨電荷為零。
在 IEF 電泳系統中,提供 pH 值梯度的環境,進行電泳時,淨電 荷為負電的蛋白質往正極移動,淨電荷為正電的蛋白質會往負極移 動,淨電荷為零的蛋白質則不易移動,直到蛋白質移動到等於其等電 點的pH 值環境中,也就是蛋白質的淨電荷為零時才會停止移動。因
此樣品中的不同種類蛋白質將會依照其不同的等電點,在具有pH 值 梯度的環境中移動到相對應的位置聚焦,以達到利用pH 值把蛋白質 分離的目的。
本實驗分別使用正常眼球、實驗組第一到第六週的眼球之水溶性 水晶體蛋白溶液與尿素可溶性水晶體蛋白溶液作IEF 電泳分析,水溶 性水晶體蛋白每次實驗的使用量為500μg,尿素可溶性水晶體蛋白的 使用量為1000μg。膠條使用 Immobiline Dry Strip 13cm ,pH 3-10,
線性,Pharmacia。
實驗方法為取出蛋白質樣品,與覆水液(Rehydration Buffer)混
合 , 使 總 體 積 為 375μL , 並 且 加 入 0.0015g 二 硫 蘚 糖 醇 (DL-Dithiothreitol; DTT)並且混合均勻,可以使用超音波震盪使蛋白 質的溶解度增加,待其充分混合後,在4 ℃、轉速為 14000 rpm 之條 件下作離心60 分鐘,取上層溶液體積共 250μL 均勻地注入等電點電 泳膠條陶瓷容槽 (Strip Holder)中,撕除膠條的塑膠保護膜,以膠面朝
下的方式小心地把膠條覆蓋在樣品之上,避免膠條下有小氣泡,確定 無氣泡之後再覆上一層礦物油(Cover Fluid),以壓縮樣品並防止其氧 化,最後蓋上蓋子即可進行一維電泳操作。水溶性水晶體蛋白及尿素 可溶性水晶體蛋白之IEF 電泳操作的梯度皆相同:Rehydration 30 V、
16 Hrs,Step-n-hold 500 V、1 Hrs,Gradient 1000 V、1 Hrs,Gradient
8000 V、11325Vhr,Step-n-hold 8000 V、12000 Vhr。
第一維電泳結束後,接著進行第二維電泳;也就是 SDS-PAGE
電泳。將作用完畢的膠條放入 5mL 含有 0.05g DTT 之平衡液中平衡 20 分鐘進行第二次還原,避免再次產生雙硫鍵;以及 5mL 含有 0.125 g IAA 的平衡液中避光平衡 20 分鐘進行甲基化反應,使之變成硫-甲 基結構,以確保不再產生雙硫鍵,經過兩次平衡之後把膠條置入丙烯 醯胺濃度為12.5 % SDS-PAGE 膠片上方,最後注入濃度為 0.5-1 % 的洋菜凍(Agarose)黏合。利用 SDS-PAGE 膠片將 IEF 膠條上已初步 分離之蛋白質再依照分子量大小不同作第二次分離。進行水溶性水晶 體蛋白及尿素可溶性水晶體蛋白的第二維電泳操作條件皆為:電壓固 定為300 V,開始電泳後的 30 分鐘內電流設定為 25 mA,之後再調 整電流為50 mA,電泳時間總共是 5 小時 10 分鐘,電泳結束後使用 Coomassie Brilliant Blue 染色一小時後,放入 Destain Buffer 中退染。
丙烯醯胺濃度為12.5 %膠片成份:
30 % Acryamide 15.625 mL 1.5 M Tris-HCl 9.375 mL D.D. Water 12.5 mL 10 % AP 375 μL TEMED 18.75 μL
2-9 質譜分析
利用二維電泳分離完蛋白質的混合物之後,必須利用影像分析 (Image Analysis)方式,才能知道蛋白質的量的變化,但是這樣的分析
只能定量,而不能知道蛋白質的身份,想要知道蛋白質的身份,必需 使用質譜儀(Mass Spectrometry)鑑定膠片上的蛋白質點,再把分析結 果對照資料庫作蛋白質的身份鑑定(Protein Identification, Protein ID)。
本實驗在作完二維電泳分離後,即把膠片掃描成圖檔進行蛋白質
點 的 比 對 工 作 , 使 用 的 比 對 軟 體 為 ImageMaster 2D Platinum Software(Version 5.0),把大白鼠實驗組第一到六週的眼球之水溶性水 晶體蛋白二維電泳膠片共六片,以及實驗組第一到第六週的眼球萃取 尿素可溶性水晶體蛋白的二維電泳膠片共六片,分別與正常眼球水溶 性蛋白、尿素可溶蛋白的二維電泳分析結果作比對,找出訊號強度相 差兩倍以上的蛋白質點作進一步的質譜分析。
另外,針對水溶性水晶體蛋白一維電泳分析結果,取出實驗組第 三週及第六週出現在低分子量範圍(低於 15kDa)的條帶進行膠內酶解 (In-Gel Digest)步驟,再配合 LC/MS/MS 以鑑定蛋白質的身份。
2-9-1 In-gel Digestion
膠片中的蛋白質沒辦法直接取出作質譜分析,因為完整的蛋白 質並不容易由電泳膠片中萃取出來,若是不經過初步的水解,由膠片
中萃取出的蛋白質量會很少。另外,質譜儀雖然可以提供蛋白質分子 量的資訊,但因為蛋白質分子不管在細胞中或是在純化的過程中常會 被修飾(Modification)使蛋白質變得更複雜,所以知道分子量並無法正 確分析出是何種蛋白質;而且 MS 在測量時仍然會產生測量時的錯 誤,愈大團的蛋白質,誤差也會愈大,因此需要使用具有序列專一性 的 蛋 白 脢(Protease) 進 行 水 解 , 把 蛋 白 質 切 成 小 段 的 多 胜 肽 片 段 (Peptide Fragments),經由質譜儀分析這些片段的組合,就能夠和資料
庫的蛋白質資料作比對,鑑定蛋白質身份。
在進行質譜實驗之前必須先把蛋白質從膠片裡取出,並且使用蛋 白脢Trypsin 水解蛋白質,這些步驟稱作膠內酶解(In-Gel Digest),本 實驗使用的是Coomassie Brilliant Blue,步驟如下:
(1)切膠:使用經過 75%乙醇清潔過的刀片切膠,膠塊大小約為邊長 1mm 的立方體,切塊後放入 1.5mL 的微量離心管中並加入 300μL 二
次水在shaker 上搖晃 15 分鐘。
(2)脫色(Destain):加入 300μL HPLC 級的乙腈(Acetonitrile, ACN)清
洗,在室溫下搖晃15 分鐘後丟棄上清液(Supernatant)。加入 300μL 的 100mM NH4HCO3清洗膠塊,在室溫之下搖晃 15 分鐘,把上清液丟 棄。再加入300μL 的脫色液(Destain Solution)( 100mM、50% ACN、
50%NH4HCO3)超音波震盪 5 分鐘之後離心,取出上層澄清液丟棄,
重覆前述步驟共三次直到膠塊變成透明無色為止。
脫色液中具有碳酸氫銨鹽類和含量 30-50%的有機溶劑(通常是 ACN),其中的有機成份可以降低蛋白質和 CBB 之間疏水性的交互作 用(Jin and Manabe, 2005),同時脫色液中的離子成份也會降低蛋白質
帶正電胺端和 CBB 之間的靜電作用力,因此混合有機溶劑和水可以 提高脫色的效果。此外,溫度提高也能幫助脫色(Lloyd, 1996)。
(3)脫水(Shrink):脫色後加入 200uL 的 100% ACN 使膠塊脫水 10 分 鐘之後移除溶液丟棄,此時膠塊收縮並且呈現白色。
(4)乾燥(Dry):使用真空乾冷系統(Dry Vacuum Centrifuge)去除溶劑 10 分鐘。
(5)還原與甲基化(Reduction and Alkylation):第一步驟先加入 50μL 10mM DTT 在 60℃下反應一小時後丟棄上清液,第二步驟為加入 50μL 50mM IAA 在避光條件下於室溫反應 30 分鐘後,吸取並丟棄上 清液。
(6)清洗(Wash):加入 300μL 100mM NH4HCO3,在室溫下搖晃 15 分 鐘,丟棄上清液; 再加入體積 300μL (20mM 50%NH4HCO3和50%
ACN)之溶液在室溫之下搖晃反應 15 分鐘,丟棄上清液。.最後加入 體積 100μL 的 HPLC 級 ACN 在室溫下搖晃 5 分鐘,移除上清液。
以上步驟完成後使用真空乾冷系統(Dry Vacuum Centrifuge)去除溶劑
10 分鐘。
(7) Trypsin:加入 20μL Trypsin 溶液並且在室溫之下反應 60 分鐘,
使蛋白質被水解成胜肽。
(8)Incubate:之後再加入足夠可以蓋過膠塊的 40mM( 90%NH4HCO3
和10% ACN)溶液,在 37℃之下反應 Overnight。
(9)萃取(Extraction):加入 150μL 無菌水與膠塊混合,在室溫之下使用 Shaker 搖晃 10 分鐘。吸取含有多胜肽的上清液放到另一個 1.5mL 的 微量離心管中,用體積 50μL (50% ACN 混合 5% TFA) 於室溫下萃取 膠塊中的胜肽共兩次,每次反應時間為一小時,重覆此步驟並將上清 液集中收集,最後使用真空冷乾60 分鐘以去除溶劑。
試劑配製(Reagent Preparation)分述於下列步驟:
(1)100mM 溶液: 395.3mg NH4HCO3加水至體積為50ML。
(2)100mM NH4HCO3與 50% ACN 混合溶液: 395.3mg NH4HCO3
與體積 25mL HPLC 級的 ACN 混合,加水至體積為 50mL。
(3)10mM DTT:7.71mg Dithiothreitol (DTT)加入 100mM NH4HCO3至
體積為5mL。(使用時再配)
(4)50mM IAA:56mg IAA\ 6.06mL NH4HCO3。(使用時再配) (5)20mM NH4HCO3 與 50% Acetonitrile 混合溶液:
10mL 100mM NH4HCO3混合 25mL HPLC 級 ACN 並且加 5mL 水。
(6)40mM NH4HCO3與 10% ACN 混合溶液:20mL 100mM NH4HCO3
加上5mL HPLC 級 ACN 以及 25mL 水。
(7) Trypsin 溶液:100μL 50mM Acetic Acid 加上 5mL 的 40mM。
NH4HCO3與10% ACN 混合溶液,分裝到 500μL 的離心管中並處存在 -80℃冰箱內保存,每管分裝最多只能冷凍-解凍重覆使用 5 次。
(8)50% Acetonitrile/5% TFA 溶液:25mL HPLC 級 ACN 混合 2.5mL TFA ,加水至體積為 50mL。
2-9-2 全蛋白質譜分析
從大白鼠眼球中萃取得出的水溶性水晶體蛋白以及尿素可溶蛋
白,分別取出150μg 的蛋白質作全蛋白的質譜分析。進行分析之前先 作前處理;三氯乙酸/丙酮蛋白質沉澱法(Trichloroacetic Acid/Acetone Protein Precipitation)。蛋白質沉澱法之目的為濃縮(Concentrate)、沉澱 (Precipitate)蛋白質樣品以及移除汙染物質 (Contaminants),例如鹽 類、清潔劑(Detergent)、核酸(Nucleic Acids)、脂質(Lipids)…等會影響
之後進行分析實驗的物質。處理步驟如下:先把要用的丙酮預先放在 -20℃的冰箱中預冷,分別取出質量為 150μg 蛋白質樣品放入微量離 心管中,加入體積為蛋白質體積四倍的冰丙酮(-20℃),然後放在-20
℃的冰箱中沉澱一小時。之後取出離心管,在 4℃、轉速為 14000
rpm(13000-15000 rpm)的條件下離心 15 分鐘,小心地取出上清液丟 棄,並不要碰到下層的蛋白質沉澱物(Protein Pellet)。接下來使用 1mL
的 冰 丙 酮 清 洗 下 層 蛋 白 質 沉 澱 , 在 4 ℃ 、 轉 速 為 14000 rpm(13000-15000 rpm)的條件之下離心 15 分鐘,並移除上清液。重複 此清洗步驟 2 次,以去除全部的 TCA,最後在室溫之下打開微量離 心管的蓋子 30 分鐘,讓丙酮從蛋白質沉澱中完全蒸發,因為殘存的 丙酮會使樣品不易回溶(Resuspension),但須注意不要讓蛋白質沉澱 過度乾燥(Over-Dry)。之後可以把樣品放入-20℃冰箱內存放,或是之 後要作二維電泳分析,可以用覆水液重覆 Pipetting 直到蛋白質沉澱 溶解。而這裡是再加入 45μL 0.1M NH4HCO3以及5μL 濃度為 1μg/μL Trypsin,反應 18 小時,將蛋白質水解成胜肽。反應結束後利用真空 冷乾系統除去樣品中的水份,並以0.1% 甲酸(Formic Acid)回溶,接 著進行質譜分析。之後的程序由貴儀中心陳明峰先生代為操作,而蛋 白質身份鑑定的資料來源是Mascot software 資料庫。