國立宜蘭大學自然資源學系 碩士論文
Department of Natural Resources National Ilan University
Master Thesis
宜蘭地區水田地景與紅冠水雞生殖生態學研究 Landscape and Breeding Ecology of Common Moorhen
(Gallinula chloropus) in Rice Paddies in Ilan
指導教授:林亞立博士
Ya-Lih Lin Ph. D.
袁孝維博士
Hsiao-Wei Yuan Ph. D.
研究生:黃仲霙
Jung-Ying Huang
摘要
紅冠水雞(Gallinula chloropus)是一種適應力很好且世界分布廣泛的秧 雞科鳥類,在台灣的水田內也常見其棲息活動。因此,為瞭解水田的地景 變化及人為經營擾動對於紅冠水雞的族群量與生殖行為的影響,本研究於 2006 年 7 月至 2007 年 10 月進行調查,樣區位於宜蘭縣冬山鄉永美社區,
面積約 247.47 公頃。調查項目包括紀錄樣區內每個區塊的水位與植被高 度,紅冠水雞成鳥及幼鳥族群量,測量鳥巢的巢形值與巢位環境因子,並 觀察生殖行為。
將水田棲地變化依照區塊類別及稻作活動產生的不同干擾程度將一整 年的時間區分成蓄水休耕期、翻土插秧期、稻作生長期、收割休息轉作期 及田菁翻田期等五個稻作經營分期。族群量變化及分布在各稻作經營分期 呈現顯著差異,其中稻作生長期提供足夠生殖棲息的植被環境且干擾低,
族群量最高;蓄水休耕期因無人為干擾,少量植被便足夠提供群聚數量生 存棲息,族群量次高。由此可知,族群數量受稻作經營的活動影響呈現明 顯的消長。生殖季節時,各稻作經營分期間的成鳥數及幼鳥數也呈現顯著 差異。成鳥在高植被的稻作生長期數量最高,為偏好之生殖環境,但其他 分期明顯因稻作活動及地景改變而數量降低;幼鳥的數量則主要受生殖季 節的影響。因此,水田的地景變化與干擾對紅冠水雞生殖行為的影響,主 要在於生殖個體的生殖成功率及族群量。
研究期間發現孵卵巢的成功與失敗巢數分別為 33 及 23 個,生殖成功 率 58.9%。成功巢與失敗巢的巢形值或巢位選擇皆無顯著差異。生殖失敗的 原因主要為人為因素,其次為巢掠食者。此外,並紀錄紅冠水雞孵卵用的 孵卵巢數,以及親鳥於夜間協助幼鳥保暖使用之育幼巢數,分別 61 個與 70 個。在不同棲息地的兩種巢數呈現顯著差異,顯示蓄水田內紅冠水雞對育 幼巢的需求增加,推測缺少足夠植被進行夜棲。此外,兩種巢的巢形值無
顯著差異,但比較水田內兩種巢的巢位選擇,孵卵巢較育幼巢具有高植被 的需求,推測是避敵考量。再進一步分別與隨機點的巢位環境因子進行比 較,發現兩種巢的巢位選擇因使用不同產生不同的避敵策略,孵卵巢較隨 機點具有高植被需求,且離田埂較遠,而育幼巢離道路與田埂較遠。但兩 者都比隨機點的水深較淺,推測皆為了降低築巢時的負擔。
由此可知,棲息在水田上的紅冠水雞族群與生殖皆受稻作活動影響,
在生殖季節則因稻作活動造成的不同地景產生不同的生殖策略。
關鍵字:紅冠水雞、生殖行為、築巢行為、水田、宜蘭
Abstract
Common moorhen (Gallinula chloropus) is an adaptable and widely distributed member of the rail family around the world. The common moorhen in Taiwan often inhabits in the rice paddy. In order to understand the population and the breeding behaviors of the common moorhen rendered by the variation of landscape and the artificial disturbance of paddy field management , a survey was conducted on a 247.47-hactare site located in the Yong-Mei area, Dong-Shan Shang in Ilan county from July 2006 to October 2007.
The deliberative parameters included were the presence and appearance of vegetation, the water depth of each paddy patch, the numbers of the adult and the young in the moorhen population, the feature of nests and the environment factors around the nest, the particular reproduction behaviors of the moorhens and data of other related information.
Based on the impacts occurred on each rice paddy patch and the disturbance degree of cultivating management, the yearly variation of rice paddy was divided into the following five periods: 1. water immerging and fallow, 2.
plowing and rice seedlings transplanting, 3. rice growing, 4. rice harvesting, and sesbavia planting, and 5. plowing and water flooding
The results reveal that the moorhen population fluctuation and distribution depended highly on the management period stated above. During the rice growing period, vegetation provided sufficient coverage for moorhen inhabitation, which rendered the highest population amount. The water immerging and fallow period performed the least landscape variation and human disturbance of the patches in the study area, hitherto, presented the second highest population amount. Furthermore, the numbers of the adult varied with the management periods during breeding. Each of the five management periods created its special inhabitations to common moorhens, and affected the numbers of the adult profoundly. The numbers of the young moorhen seemed
indicates the moorhen population was highly affected by the cultivating activities in the rice paddy.
During the survey period, 33 reproductive success nests and 23 failure nests were recorded. The reproductive success rate is 58.9%. No significant correlation among the features of the nests and the environment factors around were observed. The key factors in accordance with reproductive failure are the cultivating activities and nest predators.
In addition, there are two types of nest built by the moorhen studied. 61egg nests and 70 brood nests were recorded, the rate number of two type nests between five management period is significant difference, and there are more brood nests in water immerging paddies. But no significant difference between on the nest feature of these two types was noticed. Possibly owning to the anti-predator strategy, the egg nest was significantly more in need of surrounding vegetation in both coverage and height than the brood nest did.
Comparing the environment factors measured around the egg nest site and the randomly selected point in the same patch. The egg nest site appeared to be in need of more vegetation shelter and farther away from the levee, which may be the evidence of anti-predator strategy, while the shallower water level observed on the egg nest site may be due to the energy conservation consideration of the parents during nest building works. Results from similar comparison between the brood nest and the random point shows that the brood nest has the same requirement with the egg nest on the site factors of farther distance from the levee and shallower water level.
In conclusion, the variation of landscape and the artificial disturbance along with rice paddy management affect mainly on the individual reproductive success rate of the moorhen and the numbers of the adult in the population, and common moorhens in the rice paddy altered their breeding strategies to be in coincidence with the five cultivating management period.
Keywords:Moorhen, Breeding behavior, Nesting behavior, Rice paddy field, Ilan.
致謝
畢業後對賞鳥產生的濃厚興趣,在山區海邊趴趴走賞鳥的同時,一頭 栽進了研究所,將興趣延伸研究,是當初始料未及的事,卻開啟了我另一 扇"賞鳥的窗口"。一年多調查的日子,無論颳風下雨、烈日高照,常因 為觀察到鳥類的有趣行為而忘卻疲憊,才發現動物行為的奧秘及趣味都必 須親臨挖掘才能體會感受。
本篇論文的誕生,最先感謝的人是林亞立老師,在我研究的過程中循 循善誘、悉心導引,給予極大的支持與鼓勵,且在我論文修整期間不厭其 煩地耐心逐字斧正。並十分感謝袁孝維老師百忙中常撥冗細心指導,懇切 給予建議提醒。對兩位恩師的支持與關懷,致上最誠摯的敬意與謝意。
研究期間,謝謝毛俊傑老師提供調查方面的諮詢,陳子英老師提供的 參考資料。並蒙兩位老師與丁宗蘇博士在口試期間費心審閱並提供寶貴意 見,致深摯謝忱。
也謝謝研究室內學妹們的加油打氣,謝謝小萍、微微幫忙計算面積,
秀容協助植被調查。還有許多鳥友同事們的友情相挺,謝謝吳永華大哥提 供的建議,靜芬、昌鴻、熙梅、馨儀、定郎協助野外調查,祐薰提供的資 料,國凌、國梁主任協助將影片轉檔製片,游先生提供稻作經營活動的訪 談資料,其中更感謝季蓉在進行鳥巢調查時,不畏辛勞地陪伴調查。還有 許多經常給予鼓勵的友人們,衷心銘感。此外,也十分感謝永美社區許多 農夫們經常受我打擾,卻還是熱心的有問必答。
最後感謝陪伴我一路走來的家人們,不僅在調查研究中隨時準備出人 出力,且調查研究後給予最溫暖支援與容忍,讓我順利完成學業。
黃仲霙 謹誌
中華民國 97 年 1 月 31 日
目錄
英文摘要……… i
中文摘要……… iii
誌謝……… vi
目錄……… vii
表目錄……… x
圖目錄……… xi
附表目錄……… xii
壹、前言……… 1
貳、稻作經營活動之地景變化……… 4
一、研究地點……… 4
(一)環境概述……… 5
(二)溫度雨量……… 7
(三)稻作之經營活動……… 8
二、材料與方法……… 11
(一)地景變化調查……… 12
(二)人為稻作經營活動的干擾程度……… 14
(三)資料分析……… 15
三、結果……… 16
(一)地景變化與稻作經營活動及分期……… 16
(二)稻作經營分期與干擾程度……… 18
四、討論……… 19
參、水田棲息地與紅冠水雞族群數量之關係……… 21
一、水田生態系與鳥類關係……… 21
二、材料與方法……… 24
(一)族群與成幼鳥數量調查……… 24
( 二 ) 資 料 分 析 … … … 2 5 三、結果……… 26
(一)水田地景變化與紅冠水雞族群數量的關係……… 26
(二)稻作地景變化與紅冠水雞成幼鳥數的關係……… 35
四、討論……… 37
(一)稻作活動影響族群數量及分佈……… 37
(二)水田與生殖棲息地的關係……… 40
肆、紅冠水雞的生殖生態學……… 43
一、生殖成敗與巢位選擇……… 43
(一)生殖築巢行為……… 43
(二)生殖成功率與失敗因素……… 48
(三)巢位選擇與避敵策略……… 53
二、材料與方法……… 55
(一)生殖情況調查……… 56
(二)巢形值與巢位調查……… 56
(三)資料分析……… 58
三、結果……… 59
(一)生殖成功率及失敗原因……… 59
(二)巢生殖成功率與失敗因素……… 63
(三)築巢行為……… 6 6 (四)育幼巢的使用情況……… 69
(五)稻作經營分期與築巢行為之關係……… 71
(六)孵卵巢與育幼巢的比較……… 72
(七)孵卵巢與育幼巢的巢位選擇……… 74
四、討論……… 75
(一)生殖成敗的討論……… 75
(二)巢位選擇……… 76
(三)育幼巢的功能……… 78
伍、結論……… 81 參考文獻……… 8 3
表目錄
表1 樣區內的稻作經營過程……… 10
表2 區塊類別的覆水、植被及調查情況……… 13
表3 機械干擾與人為撫育的活動項目與分級……… 15
表4 稻作經營活動的資料分析……… 15
表5 年度中各個稻作經營分期之稻作活動所產生的干擾程度級數…… 19
表6 水田棲息地與紅冠水雞族群數量關係之資料分析……… 25
表7 紅冠水雞冬季群聚活動之周邊植被的植物組成及覆蓋度………… 34
表8 有關紅冠水雞的生殖生態學整理……… 44
表9 紅冠水雞的生殖生態學之資料分析……… 59
表10 紅冠水雞在不同棲息地的巢之生殖成功或失敗的數量………… 63
表11 紅冠水雞孵卵巢生殖失敗的因素……… 66
圖目錄
圖1 研究樣區的地理位置圖……… 4
圖2 研究樣區的地景類型面積分布圖……… 5
圖3 調查樣區內不同地景類型的每區塊面積的平均值……… 6
圖4 研究樣區之氣候生態圖,2006年7月至2007年10月月均溫與平均月雨 量……… 8
圖5 研究樣區內的所有區塊……… 11
圖6 樣區內每次調查之不同區塊類別的總面積比值與各個稻作經營分期 的時段……… 17
圖7 各個稻作經營各期內紅冠水雞的平均族群數量……… 26
圖8 蓄水休耕期之紅冠水雞分布圖……… 29
圖9 翻土插秧期之紅冠水雞分布圖……… 30
圖10 稻作生長期之紅冠水雞分布圖……… 31
圖11 收割休息轉作期之紅冠水雞分布圖……… 32
圖12 田菁翻田期之紅冠水雞分布圖……… 33
圖13 不同稻作經營分期內的成鳥及幼鳥的平均數……… 36
圖14 紅冠水雞不同棲息地的鳥巢……… 68
圖15 2007年間紅冠水雞在研究地區築巢的巢位……… 71
圖16 稻作經營分期調查所得之孵卵巢與育幼巢的巢數……… 72
圖17 紅冠水雞鳥巢的巢形值的四分位圖……… 73
附表目錄
附表 1 影響紅冠水雞族群數量的區塊類別結果……… 90 附表 2 比較成鳥數量與幼鳥數量在各個的稻作經營分期的差異之分析結
果……… 90 附表 3 影響紅冠水雞成鳥數量之區塊類別結果……… 91 附表 4 影響紅冠水雞幼鳥數量的區塊類別結果……… 91 附表 5 比較生殖成功巢與失敗巢的各項巢形值之分析結果………… 91 附表 6 比較生殖成功巢與失敗巢的各項巢位選擇之分析結果……… 92 附表 7 不同棲息地上的孵卵巢與育幼巢的巢數……… 92 附表 8 不同棲息地中孵卵巢與育幼巢的巢形值比較……… 93 附表 9 不同棲息地中孵卵巢與育幼巢的巢位選擇因子比較之分析結果 93 附表 10 孵卵巢與隨機採點的巢位選擇因子比較之分析結果……… 94 附表 11 育幼巢與隨機採點的巢位選擇因子比較之分析結果……… 94
壹、前言
水田因人類操縱故呈現稻畦阡陌、溝渠縱橫的地景,加上因人類栽種 經濟作物,地景不僅具有週期性植被與水深的高度變化,且經營活動時產 生的干擾,對於以此類環境為主要生殖棲息地的動物而言,都是一種極大 的擾動。
紅冠水雞(Gallinula chloropus)屬於鶴形目、秧雞科,為廣泛分布於世界 的鳥類,通常分布的地理範圍多於低地接近水域的環境。台灣的紅冠水雞 首次紀錄於1861 年 Robert Swinhoe 在高雄市柴山發現,分佈的範圍從平地 到中海拔的水域環境都可見其蹤跡。
紅冠水雞是Gallinula屬中最能適應不同棲息地的鳥種,因此普遍分布於 各種具有水域的棲地間,如人工或自然水域環境、靜止或流動的河流等棲 息環境(Wood, 1974;Cramp and Simmons, 1980)。許多研究指出,紅冠水雞 覓食活動區域或棲息領域都需要足夠的植被用以逃避危險(McRea, 1997;
Ritter and Savidge, 1999;Forman and Brain, 2004)。紅冠水雞的覓食棲息地 屬於茂密、挺水性的草叢地、淺水域、泥灘岸,常發現在浮葉植物上方行 走,並從水中或植物表面取食,也會以涉水或是潛水方式覓食,食物類型 包含種子、草本與水生植物的嫩芽、昆蟲、軟體動物、兩棲類幼蟲以及小 魚等(Wood, 1974;Forman and Brain, 2004;Beedy, 2005)。台灣首篇紅冠水 雞的正式(論文)報告是陳忠應(2005)研究高雄的洲仔濕地及台南縣的六個鄉 鎮內的族群,指出不同個體胃的內容物呈現多變化,包括水藻、種籽、禾 本科或浮葉植物的葉子、椿象、幼蟲、菱角等,是屬於雜食性的鳥類,且 研究顯示紅冠水雞日間活動(6:00AM-6:00PM)的行為模式,有74%的時間在 進行覓食活動,可知該鳥種選擇棲息地時,最佳覓食地應該是很重要的考 量因素。除了覓食外,安全的考量亦十分重要,Ritter and Savidge(1999)研 究馬尼拉群島上的馬尼拉紅冠水雞(Gallinula chloropus guami)族群數量與
自然濕地的減少成正相關,且Ritter and Savidge(1993)較早的研究報告指出 同地的紅冠水雞分布數量與挺水植物的覆蓋度成正相關。
紅冠水雞的雌雄外型十分相近不易分辨(王嘉雄等;1991),陳忠應(2005) 研究指出雌雄細部的不同,如雄鳥的翅膀、趾、跗蹠骨、嘴喙的長度都顯 著高於雌鳥。紅冠水雞成鳥體長約30-38cm,雄鳥平均體重339g,雌鳥平均 體重約245g。生殖期時的成鳥,頭頸黑色,前額板紅色,嘴喙紅色,嘴尖 黃色,身體上半部暗橄欖綠色,下腹部灰色,翼側兩脅有白斑,尾部白色,
腳黃色。非生殖期時,與生殖期相近,但是嘴喙到前額板呈暗紅色。亞成 鳥頭頸灰色,喉部有黑色細斑,下半部灰色,胸部則有一塊白斑,嘴暗紅 色,腳則暗綠色(Gillmor et al., 1983;王嘉雄等,1991)。
Forman and Brain(2004)研究紅冠水雞的生殖行為,指出英國康橋郡內 族群的生殖季節開始前,先產生領域的競爭行為,並構成生殖配偶或生殖 組合,整個生殖季節內生殖組合的個體會共同防禦領域。此外,幾篇英國 境內紅冠水雞的研究都指出,生殖求偶期約1、2月開始,且生殖配偶或組 成的方式多樣,大都為一雄一雌的單配偶型(monogamous),也有一雄多雌 (polygynous)、一雌多雄(polyandrous)、多雌多雄(polygyandrous)等類型,以 及生殖配偶與無生殖成鳥或幼鳥的合作生殖組成(Wood, 1974;Petrie, 1984;
Gibbons, 1987;McRae, 1995, 1996;Beedy, 2005;陳忠應,2005)。生殖個 體會共同孵卵和育雛。雛鳥為早成鳥,一經孵化後,即全身披茸毛且可立 即活動(Gillmor et al., 1983;Forman and Brain, 2004)。紅冠水雞的巢通常築 於濱臨水域或是水體上,選擇巢位棲息地也偏好高植被覆蓋度的環境 (Cramp and Simmons, 1980;Taylor, 1984; Gibbons, 1989)。此外,Wood(1974) 與Gibbons(1987)亦同時指出生殖個體通常在生殖季結束後,會形成冬季群 聚行為並在同一區域覓食活動。
雖然有許多報告研究紅冠水雞生殖的相關行為,但是多數的研究環境
是濕地棲息地,較少是針對棲息於水田中的紅冠水雞。而水田棲息地因人 為的稻作經營活動產生諸多不同變因,勢必對於棲息於此類型棲息地上的 紅冠水雞造成影響。
宜蘭縣的平原地景以水田為主,因地形氣候上的限制,土地不利作物 生長,稻作產量偏低,民國70年代政府推廣稻米生產及稻田轉作時,便以 全年休耕或一期稻作為主(簡文憲,1990)。此外,為因應國人飲食習慣改變 及稻米進口的衝擊,與配合行政院農委會推廣稻田輪作、休耕等措施(行政 院農委會,2003),縣內稻田有經營耕作都以一期稻作,二期轉作綠肥作物 為主,冬季則以蓄水休耕的方式至次年春耕,因此地景便產生年度週期性 的變化。這些變化是否會造成棲息於水田內的紅冠水雞產生不同的生殖策 略,而紅冠水雞的族群是否會隨著稻作經營而產生不同的變化或分布情 況。且在7、8月水田收割翻土後的蓄水田中,經常可以發現紅冠水雞在無 植被掩蔽的環境下築巢,且常發現許多鳥巢並非使用於孵卵,故紅冠水雞 在選擇築巢的巢位是否有特別意義,及何種原因促使紅冠水雞築非孵卵用 巢。
基於以上之因素,本研究於2006年7月至2007年10月針對宜蘭縣冬山鄉 永美社區內的水田進行紅冠水雞的研究,監測生殖族群數量的變化、成鳥 與幼鳥的數量變化,並調查生殖成敗的原因、巢的類型,及巢位周邊的環 境因子。由此調查結果可藉以深入探討動態環境下,紅冠水雞適應後是否 會產生對應的變異行為。此外,紅冠水雞在台灣本島是普遍分佈的留鳥,
故本研究藉由監測水田棲息環境內之族群的生殖行為,不但可反應出受稻 作活動影響的情況,可更深入認識該物種的生殖生態學,並且期望透過對 紅冠水雞因應人為經營棲息地的生殖與活動模式的探討,未來可做為生殖 或棲息水田上的物種之保育與經營的參考。
貳、稻作經營活動之地景變化
稻作經營產生地景改變與活動干擾,故本章目的除對調查樣區進行描 述外,並紀錄樣區內的整年稻作經營所呈現之不同地景類型與活動情況,
再進行分期,做為瞭解年周期環境變動中紅冠水雞產生適應行為之對應依 據。
一、研究地點
本研究樣區位於宜蘭縣冬山鄉(24°38’N 121°46’E),台九線、永美路、
義成路與冬山河等四個邊界所圍成之區域,樣區面積約247.47 ha(圖1)。本 區東側約5 km處為境內著名之五十二甲濕地,冬季常聚集大批水鳥於該處 棲息;東南方約10 km處為民國84年農委會公告之無尾港水鳥保護區。自 2006年7月起至2007年10月底進行水田棲息地上的紅冠水雞調查監測。調查 的內容包括地景區塊變化、紅冠水雞族群組成及分布的變化,及紅冠水雞 築巢生殖行為監測等工作。
圖1 研究樣區的地理位置圖 Fig.1 Location of Study site.
註:圖片來源 google earth
(一)環境概述
傳統水田多以田埂為界且呈不規則狀,經農地重劃後大多長約100 m、寬約 35 m 的長方形,於本研究中稱之為區塊(patch)。本樣區以水 田為主要地景類型,另有房舍、溪流、灌溉溝渠、荷花田、茭白筍田 等不同地景類型。
以2006 年的資料為準,研究樣區面積約 247.47 ha。水田區塊(patch) 共計 873 塊,面積約 186.93 ha,佔總地景面積 75%;其他非建物區塊,
包括茭白筍田 1 塊、荷花水池 4 塊、菜圃 3 塊、給排水渠道(超過 2 m 寬的河流),及灌溉溝渠,共計 1.81 ha,佔總地景面積 1%;建物區塊(包 括建築物、石礫空地)207 塊及道路,面積計 59.54 ha,佔總地景面積 24% (圖 2)。
24%
75%
1%
建物 59.54 ha 稻田 186.93 ha 其他 1.81 ha 圖2 研究樣區的地景類型面積分布圖
Fig.2 Contrbution of different landscape categories in the study site.
比較本樣區內不同地景類型區塊的平均面積與標準差,發現建物 區塊的平均面積較高於水田與其他,區塊間變異大;而水田的個別區
著一條灌溉溝渠,及一條約3 m寬可供車輛行駛的道路,加上本區為一 般稻作型態,並無其他不同之稻作經營方式在內,如有機稻作或合鴨 米稻作等,因此每區塊受人類的干擾情況一致,且相似經營模式的棲 息環境,於棲地內提供物種存活的可能食物質量,應屬於均質分佈的 狀態。
0 1000 2000 3000 4000 5000 6000 7000 8000
建物 水田 其他
平均面積(㎡)
圖3 調查樣區內不同地景類型的每區塊面積的平均值 Fig.3 The means +SD of area in each landscape category.
註:〝其他〞只計算菜圃、荷花田、茭白筍田的區塊
note:others include a vegetable paddies, a lotus paddies and a wild rice paddies.
近年來,因為交通便利、觀光產業發展,農耕地急遽轉變為農舍,
2006 年 7 月至 2007 年 10 月調查期間,4 塊原為稻作田變更為建築用 地並興建房舍。近年來因政策性的推行,土堤田埂被水泥田埂取代,
灌溉溝渠也多數改建成水泥溝渠,這些轉變都將原本連續性的生態環 境切割成零碎化的地景地貌,對原有豐富植被或土壤的微棲地環境造 成影響。本區內有三條超過 2 m 寬的給排水渠道(溪流)流經,引水至灌 溉溝渠內,溝渠內幾乎終年有水,以因應稻作經營時所需。
本樣區內的茭白筍田僅有一區塊且有人為經營管理,其餘零星的 茭白筍則成排狀分布於水稻邊的溝渠內。人為的經營管理開始於春 季,通常整地後植株開始萌芽分蘗,9 - 11 月為採收期,秋冬為茭白筍 的休眠期 (李文汕,2000)。本區內的 4 塊荷花田皆無人經營管理,其 中 2 塊於 2006 年秋天翻土蓄水休耕,隔年不再耕作,另 2 塊荷花田則 於秋冬進入休眠期後,水上植株會逐漸腐壞,露出水域面積,直至隔 年春天才會重新生長荷葉至水面上。
(二)溫度雨量
依據中央氣象局蘇澳站資料顯示2006 年 7 月至 2007 年 10 月的平 均溫度24.0 ℃,平均雨量 372 ㎜(2006 年 11 月至 2007 年 10 月年均溫 23.1 ℃,平均年雨量 386 ㎜),屬於終年較潮濕的氣候環境(圖 4)。總計 2006 年 7 月至 2007 年 10 月,台灣地區共發佈 10 個颱風警報(2006 年,
碧利斯、凱米、桑美、寶發、珊珊;2007 年,帕布、梧提、聖帕、韋 帕、柯羅莎),無論暴風圈有無覆蓋或路經宜蘭地區,都為宜蘭帶來許 多雨量,造成水田內水位升高。颱風在八月份較多,這期間稻米收成 完畢後轉作綠肥,也造成綠肥作物植物體因風雨而傾倒。
圖 4 研究樣區之氣候生態圖, 2006 年 7 月至 2007 年 10 月月均溫與平均 月雨量
Fig.4 The climate ecology diagram, the average temperature and rainfall per mouth in July 2006 to October 2007.
註:資料來源中央氣象局宜蘭地區蘇澳氣象觀測站 note:data form Suao station of the Central Weather Bureau.
(三)稻作之經營活動
本人於研究期間訪問農作經營者,所得可知本樣區的稻作經營活 動約在 2 月中旬至 3 月開始,農夫在耕作插秧前先進行翻土整地,再 使用插秧機栽種秧苗。插秧至分蘗生長時期,農夫通常會施灑選擇性 除草劑(顆粒狀)於水田中,待水田內的雜草除去後改灑肥料,直至水稻 於分蘗期生長階段,此時約 1 個星期巡視田間一次,若天候不佳或是 有病蟲害則增加巡田的次數。水稻於最高分蘗生長期至收穫前,即插 秧後的第 2 個月,此時水田內不再蓄水且水稻植高茂盛,農作活動改 以田埂巡視,並噴灑農藥或施灑肥料。稻子結穗時期至 6 月中旬稻穀 成熟之前,農夫再視病蟲害情況噴灑農藥,並增加巡視田間的次數。
6 月底開始使用機械收割,收割後部分稻田會以焚燒稻梗以改變地
力,部分保留收割後的稻梗,大部分水田都會翻田蓄水,此期約 1 個 半月。爾後水田陸續開始進行二期轉作的綠肥作物施灑,主要綠肥作 物為田菁(Sesbania sesban),地景相貌呈現另一種植被類型,美觀且增 加土地肥力。約 9 月底,當田菁生長約 1 個月半月後,便又開始翻土 蓄水,然後度過整個冬天,整個地貌主要呈現蓄水樣貌,且才真正少 有農作活動產生的干擾。而本樣區的主要水田地貌則因耕作模式而具 有年度週期及規律性的經營活動(表 1)。
與黃如鈺(2001)應用衛星遙測影像及影像處理方式歸納台灣地區 的第一期水田的土地覆蓋情況及水稻的生長週期進行對照,2 月中至 3 月初為插秧期,土地覆蓋包括水體與植被;3 月初至 6 月初為分蘖期,
土地覆蓋則為植被;6 月初至 8 月初為收穫期及休息期,土地覆蓋為裸 露地。結果與本樣區呈現的稻作經營時間與情況相近。
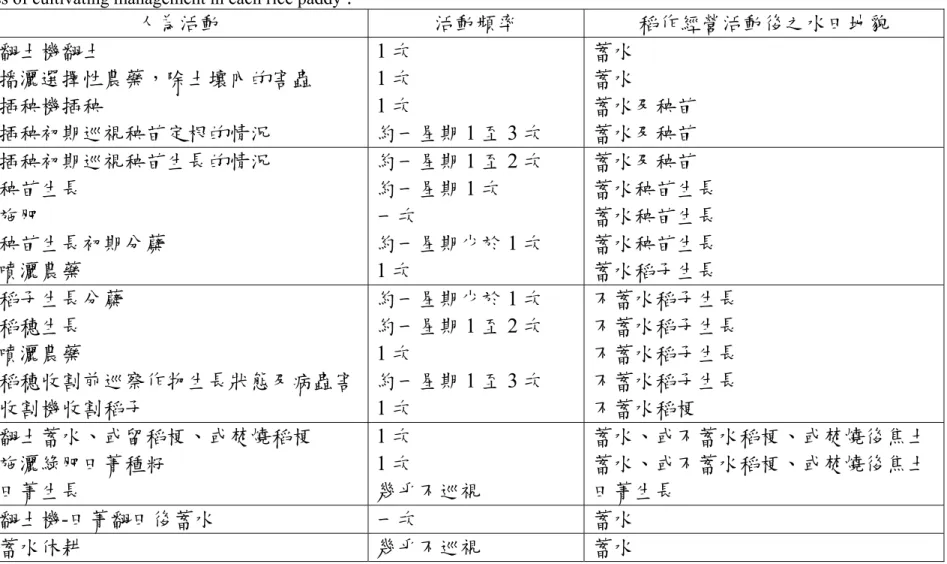
表1 樣區內的稻作經營過程
Table 1 The process of cultivating management in each rice paddy .
時間 人為活動 活動頻率 稻作經營活動後之水田地貌
2 月底 3 月初
翻土機翻土
播灑選擇性農藥,除土壤內的害蟲 插秧機插秧
插秧初期巡視秧苗定根的情況
1 次 1 次 1 次
約一星期1 至 3 次
蓄水 蓄水
蓄水及秧苗 蓄水及秧苗
3 月至 4 月底
插秧初期巡視秧苗生長的情況 秧苗生長
施肥
秧苗生長初期分蘗 噴灑農藥
約一星期1 至 2 次 約一星期1 次 一次
約一星期少於1 次 1 次
蓄水及秧苗 蓄水秧苗生長 蓄水秧苗生長 蓄水秧苗生長 蓄水稻子生長
5 月至 6 月底
稻子生長分蘗 稻穗生長 噴灑農藥
稻穗收割前巡察作物生長狀態及病蟲害 收割機收割稻子
約一星期少於1 次 約一星期1 至 2 次 1 次
約一星期1 至 3 次 1 次
不蓄水稻子生長 不蓄水稻子生長 不蓄水稻子生長 不蓄水稻子生長 不蓄水稻梗 7 月至 8 月
翻土蓄水、或留稻梗、或焚燒稻梗 施灑綠肥田菁種籽
田菁生長
1 次 1 次
幾乎不巡視
蓄水、或不蓄水稻梗、或焚燒後焦土 蓄水、或不蓄水稻梗、或焚燒後焦土 田菁生長
9 月 翻土機-田菁翻田後蓄水 一次 蓄水
10 月至隔年 2 月 蓄水休耕 幾乎不巡視 蓄水
二、材料與方法
於調查前,本人先利用Google earth呈現的地景空照圖進行繪製 本樣區之區塊調查圖。區塊調查圖呈現樣區內每塊不同類型的區塊,
區分成白色區塊及黑色區塊,其中白色區塊包含水田、荷花田、茭白 筍田、菜圃、深水域,黑色區塊包括建物(包括住家、庭院、水泥空 地),道路等(圖5)。每一區塊之面積以實際地表與區塊調查圖的比例 尺進行換算,可將每次調查後之每塊區塊地貌類型加以加總計算,結 果不僅用於瞭解本樣區內土地利用面積大小,並可瞭解樣區內稻作活 動與地景變化情況。
圖5 研究樣區內的所有區塊。
Fig.5 Patch formation in study site.
註:白色區塊為水田及水域環境,黑色為建物及道路。
note:rice and water paddy in white, building and road in black.
(一)地景變化調查
於2006 年 7 月底至 2007 年 10 月底,每兩個星期進行一次 調查,紀錄各非建物區塊的植被與覆水情況,做為人為經營稻作 後的地景區塊調查資料。區塊類別描述說明如下:
1.植高狀況
調查時區塊內若植被覆蓋度低於 50%,且有植被部份密 度十分稀疏,約1 m2少於 10 棵植株,則紀錄為無植被。若 50%以上有植被覆蓋,且植被不屬於十分稀疏,則為有植被 狀態。再將有植被狀態,以15 cm 為測量,據以將植被狀況 分為45 cm 以下為低植被,45 cm 至 90 cm 為中植被,90 cm 以上則為高植被。
2.覆水狀況
水田內無水,紀錄為無水狀態;水深超過土面,則記為 有水狀態。
3. 地景狀態的區塊類別
將以上植被、覆水狀況合併,將區塊狀態區分成八個類 別,包括無植被無水區塊、無植被有水區塊、低植被無水區 塊、低植被有水區塊、中植被無水區塊、中植被有水區塊、
高植被無水區塊、高植被有水區塊(表 2)。
每個區塊呈現的類別視為地景狀態的表現,每次地景狀 態因土地變化的情況而有所改變,據以反應樣區的稻作活動 時期與觀察紅冠水雞時的調查情況。
表2 地景狀態的區塊類別
Table 2 Critaria for each type of landscape category.
區塊類別 覆水狀況 植高狀況 調查情況
無 植 被 無 水 區塊
土地呈無蓄水狀態,
土 壤 含 水 歸 於 此 類 型。
無植被的田地,或有植被但覆蓋面積達到該區塊面積的 50%以 下,且植被密度十分稀疏(1 m2少於10 棵植株)。
可清楚觀察到區塊上鳥類 個體活動情況。
無 植 被 有 水
區塊 土地呈有蓄水狀態。 無植被的田地,或有植被但覆蓋面積達到該區塊面積的 50%以 下,且植被密度十分稀疏(1 m2少於10 棵植株)。
可清楚觀察到區塊上鳥類 個體活動情況。
低 植 被 無 水 區塊
土地呈無蓄水狀態,
土壤含水歸於此類。
植被覆蓋面積達到區塊面積的50%以上,且植被密度不屬於十分 稀疏者歸於有植被狀態。且植被的平均高度低於45 cm 以下。
可清楚觀察到植被間的鳥 類個體活動情況。
低 植 被 有 水
區塊 土地呈有蓄水狀態。 植被覆蓋面積達到該區塊面積的50%以上,且植被密度不屬於十 分稀疏者歸於有植被狀態。且植被的平均高度低於45 cm 以下。
可清楚觀察到植被間的鳥 類個體活動情況。
中 植 被 無 水 區塊
土地呈無蓄水狀態,
土 壤 含 水 歸 於 此 類 型。
植被覆蓋面積達到該區塊面積的50%以上,且植被密度不屬於十 分稀疏者歸於有植被狀態。且植被的平均高度範圍為45 cm 至 90 cm 之間。
僅能觀察個體於植被內接 近路邊約1 m 左右的活動 情況。
中 植 被 有 水
區塊 土地呈有蓄水狀態。
植被覆蓋面積達到該區塊面積的50%以上,且植被密度不屬於十 分稀疏者歸於有植被狀態。且植被的平均高度範圍為45 cm 至 90 cm 之間。
僅能觀察個體於植被內接 近路邊約1 m 左右的活動 情況。
高 植 被 無 水 區塊
土地呈無蓄水狀態,
土 壤 含 水 歸 於 此 類 型。
植被覆蓋面積達到該區塊面積的50%以上,且植被密度不屬於十 分稀疏者歸於有植被狀態。且植被的平均高度高於90 cm 以上。
無法清楚觀察到鳥類個體 在植被內活動的情況。
高 植 被 有 水
區塊 土地呈有蓄水狀態。 植被覆蓋面積達到該區塊面積的50%以上,且植被密度不屬於十 分稀疏者歸於有植被狀態。且植被的平均高度高於90 cm 以上。
無法清楚觀察到鳥類個體 在植被內活動的情況。
(二)人為稻作經營活動的干擾程度
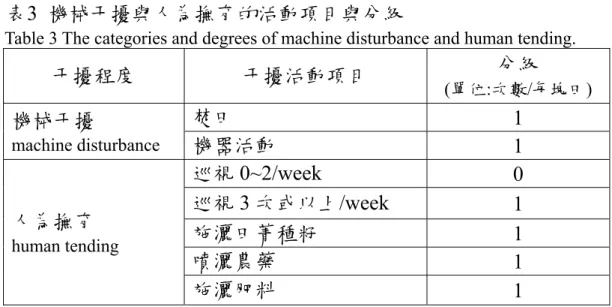
訪問宜蘭縣目前正在從事稻作的經營者,並從大約的日期 來歸納區分稻作經營的人為活動,進而分析人為活動所產生的 干擾程度。干擾程度區分為機械干擾與人為撫育。
1.機械干擾
此區使用機械進行的稻作活動通常為合資租賃機械共 同連續進行稻作活動,加上機械活動造成地景改變,故對 紅冠水雞干擾高於人類活動,因此界定為機械干擾。而焚 田的稻作活動僅於收割後,也是連續性且改變地景,因此 與機械活動同為機械干擾的項目。
2.人為撫育
以人類活動為主,包括巡視田間、施肥、噴灑農藥等活 動。因為短暫巡視不至於影響紅冠水雞回原來棲息地,而 較頻繁的人類活動可能造成個體離開原棲息地或棄巢等行 為,故將巡視田間每週3次或以上之頻度才歸於人為撫育。
而灑田菁種籽、施肥、噴灑農藥都因為存留於田中,故視 為對紅冠水雞棲息具有影響力,因此也界定為人為撫育。
而機械干擾與人為撫育的計算方式僅以活動項目的級數各 設定為1(表3),再將各個時期發生的次數加總做為各個時期所 產生的機械干擾與人為撫育的級數。
表3 機械干擾與人為撫育的活動項目與分級
Table 3 The categories and degrees of machine disturbance and human tending.
干擾程度 干擾活動項目 分級
(單位:次數/每塊田)
焚田 1
機械干擾
machine disturbance 機器活動 1
巡視0~2/week 0 巡視3 次或以上/week 1
施灑田菁種籽 1
噴灑農藥 1
人為撫育 human tending
施灑肥料 1
(三)資料分析
使 用SAS statistical software (SAS Institute Inc. 1999) 與 Microsoft Office Excel分析資料。所有資料的顯著水準皆以α=0.05 為準則。稻作經營活動之統計方法,討論變因與討論目的,如表 4。
表4 稻作經營活動的資料分析
Table 4 Methods applied on data analysis of cultivating management activities.
項目 討論目的 統計方法
計算每次調查之八種區塊類別的面積,依 照地景與人為經營活動的狀態將全年區 分成五期的稻作經營分期,做為劃分時間 的依據,便於進行其他部分的討論。
階層式群集法之完 全連鎖法(Hierarchical cluster-complete linkage method)
八 種 區 塊 類 別 與 稻 作 經 營 分 期的關係
利用稻作經營分期與各階段的區塊類別 之總面積進行探討,檢測每個分期內的地 景區塊類別是否顯著差異,檢測集群的穩 定度。
多變量變異數分析a (one-way MANOVA)
干 擾 程 度 與 地 景 區 塊 類 別 面 積 的 關 連 性
因人為經營活動對地景地貌產生變化,同 時也形成干擾程度,故使用稻作經營分期 界定干擾程度。干擾程度區分為機械干擾 與人為撫育。分析機械干擾與人為撫育與 地景區塊類別面積進行探討,藉以找出地 景變化與干擾程度的相關性。
典型相關分析 (canonical correlation)
a
三、結果
(一)地景變化與稻作經營活動及分期
本研究自2006年7月底至2007年10月底為止,共計調查33次 地景變化的情況。依據無植被水區塊、無植被無水區塊、低植被 無水區塊、低植被有水區塊、中植被無水區塊、中植被有水區塊、
高植被無水區塊、高植被有水區塊等8 種區塊類別,將每次調查 地景資料將各個區塊劃分8種區塊類別外,並計算每次調查之區 塊類別的總面積,再進行統計分類(階層式群集法),將整年稻作 經營活動分成五期。
分析結果將人為經營土地的耕作活動情況分為稻作經營五 期(圖6),每一期呈現不同的主要地景類型。第一期收割休息轉作 期,時間約7、8月(20060730-20060901、20070714-20070925),
此期稻作收割後,除部份以翻田蓄水外,地景以低植被無水區塊 的稻梗田為主;第二期田菁翻田期,時間約於9月(20060903- 20061005、20070926-20071007),此期地景主要為低、中植被的 田菁田為主,且幾乎所有田皆有蓄水狀態;第三期蓄水休耕期,
時 間 約10 月 到 隔 年 2 月 (20061006-20070309 、 20071008-2007 1028),此時地景以無植被有水區塊為主;第四期翻土插秧期,
時間約3、4月(20070310-20070503),此時大部分的田都已經翻土 並插秧,故地景以低植被有水區塊為主;第五期稻作生長期,時 間約5、6月(20070504-20070713),此時稻穀已生長較高,故地景 以高植被無水區塊為主。
利用多變量變異數分析(one-way MANOVA)檢測集群的穩 定度,即為檢測稻作經營的分期是否合適,結果顯示稻作經營各 期間的8種區塊類別總面積呈現顯著差異(λ<0.0001),即代表每一
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
2006/7/30 2006/8/6 2006/8/13 2006/8/20 2006/8/26 2006/8/30 2006/9/3 2006/9/16 2006/9/23 2006/9/30 2006/10/6 2006/10/15 2006/10/21 2006/10/29 2006/11/4 2006/11/11 2006/11/20 2006/11/25 2006/12/2 2006/12/9 2006/12/16 2006/12/23 2006/12/30 2007/1/7 2007/1/14 2007/1/20 2007/1/29 2007/2/5 2007/2/11 2007/2/19 2007/2/23 2007/3/4 2007/3/10 2007/3/17 2007/3/25 2007/4/1 2007/4/8 2007/4/14 2007/4/22 2007/4/28 2007/5/4 2007/5/12 2007/5/20 2007/5/27 2007/6/1 2007/6/9 2007/618 2007/6/24 2007/7/1 2007/7/8 2007/7/14 2007/7/22 2007/7/28 2007/8/5 2077/8/11 2007/8/19 2007/8/24 20079/2 2007/9/8 2007/9/15 2007/9/23 2007/9/29 2007/10/8 2007/10/14 2007/10/21 2007/10/28
各區塊類型對總面積的比值
無植被無水區塊 無植被有水區塊 低植被無水區塊 中植被無水區塊 高植被無水區塊 低植被有水區塊 中植被有水區塊 高植被有水區塊 圖6 樣區內歷次調查8種區塊類型對總面積的比值與對照各個稻作經營分期
Fig.6 Ratios of the 8 landscape categories in accordance to the 5 cultivating management periods.
稻作經營分期內的地景呈現顯著不同的主要地景類型,故適合以 不同稻作經營分期進行討論。
(二)稻作經營分期與干擾程度
本研究除了發現人為經營之稻作活動與地景變化呈現顯著關係 外,再進一步探討稻作經營活動與干擾程度的關係。
本樣區內共有873塊水田,人為經營稻作活動的時間並不一致,故 區分稻作經營分期內的稻作活動,每一稻作經營分期依據機器運作及 人為操作的產生的級數進行加總,區分成不同分期的干擾程度(表5)。
由上一節結果可瞭解,地景類型可區分不同稻作經營分期,且稻 作經營分期產生不同干擾程度,故將干擾程度與區塊類別進行分析。
結果發現2種干擾程度(機械干擾、人為撫育)與8種的區塊類別總面積 (無植被無水區塊、無植被有水區塊、低植被無水區塊、低植被有水區 塊、中植被無水區塊、中植被有水區塊、高植被無水區塊、高植被有 水區塊)有極顯著的關連性(典型相關分析,ρ=0.970855,λ<0.0001),表示 干擾程度與區塊類別之間有極大的相關性。故可判斷稻作經營分期呈 現不同的地景變化,也等同於有不同等級的干擾程度。
以上可知,人為經營稻作活動可從地貌變化加以分期,也可將活 動產生的干擾程度加以分類。因此,從稻作經營分期探討紅冠水雞族 群數量與生殖組成的族群數量,便等同於可以反應各期主要的地景狀 況,且可解釋各期不同的干擾程度。
表5 年度中各個稻作經營分期之稻作活動所產生的干擾程度級數 Table 5 Disturbance degrees of each cultivating management period in a year.
各期的級數 稻作經營
分期 月份時間
機械干擾 人為撫育 稻作經營的活動 蓄水休耕期 10 月 初 -2
月底 0 0 蓄水休耕
翻土插秧期 3 月初-4 月
底 2 3
翻土機翻土、插秧機插秧、插 秧 初 期 巡 視 秧 苗 定 根 的 情 況、施肥、噴灑農藥
稻作生長期 5 月初-7 月
初、中旬 0 3
噴灑農藥、稻穗收割前巡察作 物生長狀態及病蟲害、收割機 收割稻子
收割休息轉 作期
7 月中-8 月
底 2 1 翻土蓄水、或留稻梗、或焚燒
稻梗、施灑綠肥田菁種籽 田菁翻田期 9 月 1 0 翻土機
四、討論
本樣區以水田為主要地景,因重劃而每塊水田呈現相近的面積與干擾
,加上同為一般稻作模式,故樣區內的水田具有均質性。故每塊水田對於 此棲息生殖的紅冠水雞個體則相對影響不大,故排除水域或植被面積影響 個體棲息生殖的選擇,以及排除干擾影響分布情況。而相似經營模式的棲 息環境,可能提供個體的食物質量也相對均質,亦排除因食物資源而產生 的分布不均。故本研究針對整體環境的變化與本樣區內棲息的紅冠水雞個 體進行探討,期望進一步瞭解水田棲息地影響紅冠水雞棲息生殖的主要原 因為何。加上,台灣各地的稻作經營因氣候地理因素或政策宣令而有所不 同,但宜蘭地區的稻作型態較台灣中南部二期稻作的環境更容易造成整年 內具有不同地景類型的變化,對棲息水田的物種不僅需要考量到空間上的 生存,也要考量到時間序列上所造成的影響,故本研究考量兩者的影響 力,一併以時間與空間的概念探討棲息地與紅冠水雞的關係。
本研究將研究樣區內整年的稻作活動區分成五個稻作經營分期,而種 植稻穀(一期稻作)的時間、地貌與黃如鈺(2001)的報告有相近的結果,故
可瞭解稻作經營活動的時間各地差異不大,且地景變化的狀態也會呈現相 似地景面貌。再者,各個稻作經營分期不但呈現相異的主要地景,且因稻 作活動不同也產生不相等的干擾程度,故每分期內的不同地景與干擾不僅 能代表該期的狀態,亦具有解釋稻作經營活動的情況。因此可知水田棲息 地利用稻作經營分期進行時間與空間上的分類,不僅合理且有代表性。故 將進一步利用稻作經營分期探討紅冠水雞數量在各期呈現的結果,及各稻 作經營分期內或各分期間的轉變是否影響紅冠水雞的生殖行為,以做為紅 冠水雞反應該類型棲息環境所形成的適應性行為。
參、水田棲息地與紅冠水雞族群數量之關係
本章主要目的為探討紅冠水雞在水田棲息地上的族群數量與分佈情 況,以調查期間於本樣區內觀察紀錄到的個體情況,與前章的稻作經營分 期進行分析討論,據以瞭解棲息地的空間與時間變動與紅冠水雞的行為之 關係,做為呈現該物種棲息生殖的影響因素,及對應變動後的適應行為依 據。
一、水田生態系與鳥類的關係
所謂生態區位(ecological niche)不僅是表示一個生物體在群落的生態角 色,亦包括如何與其他物種相互作用以及如何受限制的情況(王瑞香,
2005)。而鳥類選擇棲息地的要素在於可以順利覓食且滿足其食物來源,以 及可以充分利用棲息地的型態結構達到避免被掠食,做為其具有偏好棲息 地的選擇行為(Martin, 1985)。尚玉昌(2003)也指出每一物種對生活地點類型 存在著不同的選擇或偏愛,任何的競爭、覓食或生殖等因素也會影響分布,
想要證明棲地選擇具有演化上的意義,就必須考量最利於生存和生殖的部 分。
現今許多自然棲息地因為人口快速成長而被開墾做為經濟活動的土地 利用,造成許多原本存在自然棲息環境下的動、植物面臨生存的壓力 (Soderstorm et al., 2003)。但是,相反的也有許多動、植物卻選擇以人為經 營的區域環境做為生存繁衍的棲息地。目前全世界上經營農作的土地面 積,約佔了陸域棲息地的 30%,其中許多經營稻作的土地,因產業活動需 要形成水域環境,儼然成為許多濕地動物偏好的棲息環境(Tourenq et al., 2001;Pierluissi, 2006)。
Tourenq et al.(2003)及 Zhijun et al.(2004)研究發現藉著瞭解棲息於稻田 內鳥類之生態系統,除了可深入瞭解其族群動態及生物學,亦可進一步發
覺土地利用的變化情況。陳炤杰與盧惠敏(2004)研究指出以淺水域型態為主 的稻田棲息地,存在著許多以此類型環境為棲息地的物種,而此環境因人 類種植經濟作物的經營方式,土地常遍布道路與給排水溝渠系統,故形成 作物棲息地與週遭之溪流、水路及產業道路系統自成獨特的生態體系,甚 至農作物與野生動、植物間亦構成食物網的組成型態。
棲息於水田的多樣物種中,就食物鏈的角色及調查的方便性,動物中 以鳥類較容易成為主要研究的對象。且鳥類是生物多樣性研究中,最多研 究報告的一個類群,因此也是最容易應用為環境指標的一個類群(Bibby, 1999;Herkenrath, 2002)。
許多探討水田環境與鳥類的研究中,亦有探討稻作經營與鳥類的關係 者。Pierluissi(2006)研究路易斯安那州西南部水田中水鳥生殖的使用情況指 出,生殖水鳥個體與稻作生長時期有依存關係。且稻作經營所形成的灌溉 系統與田埂系統提高的土地利用,因溝渠與田埂足以有效的控制水田內水 位及每個區間 (section)的土地利用,並且控制區間中的水量與植物生長的 方式,因此生殖季節時只要植物覆蓋度愈高便會吸引愈多的水鳥進行生 殖,但相對地,也增加哺乳類掠食者的掩蔽性,而容易造成鳥類生殖失敗;
此外,田埂與溝渠通道同時可成為逃離掠食者或稻作收割時鳥類躲避的路 徑。陳炤杰與盧惠敏(2004)的研究也指出稻田因為耕作活動的改變,會產生 鳥種的變化,報告顯示稻田分成秧苗期、成長期、結穗期及割後期,而每 期在覓食或生殖部分都有不同的鳥類多樣性及歧異度。故棲息地的轉變將 改變鳥類生存的棲息環境,即棲息環境條件的優劣會造成鳥類族群數量消 長。欒曉峰等(2004)研究上海農耕地的鳥類群落架構指出,農耕地中以水鳥 (如夜鷺、白鷺等)較多,約佔三分之一,其中又以留鳥居多。
也有報告也顯示稻田是鳥類的生殖棲息地,並探討稻作或水田與鳥類 生 殖 的 關 係 。 羅 柳 墀(2004) 研 究 指 出 台 灣 南 部 地 區 彩 鷸 (Rostratula
benghalensis)的族群數量及生殖成功率與農民的耕作活動有直接且密切的 關係,在休耕半荒廢的農田中,彩鷸生殖族群密度與生殖成功率最高,主 要原因則為農民踩踏或盜取鳥蛋等的人為干擾,可造成高達41.8%的生殖失 敗率。Soderstorm et al.(2003)研究非洲西部地區耕作活動與生殖鳥類或遷徙 鳥類的群聚關係發現,耕作時間與鳥類數量呈現負相關。其他有關人為經 營活動影響鳥類族群數量的報告也有很多。Henrik and Schmidt(2003)研究丹 麥東方的Klydesø 保護區內的濕地牧草經營活動與小辮鴴(Vanellus vanellus) 和反嘴鴴(Recurvirostra avosetta)生殖數量呈現顯著相關,牧草收割後會增加 生殖的築巢棲息地。Atkinson et al.(2005)研究英國濕地與鳥類覓食的關係發 現,鳥類常因植物體掩蔽力降低而增加被掠食的可能性。Borad et al.(2001) 研究報告中也指出,棲息於印度稻田內赤頸鶴(Grus antigone antigone)數量 及分布與水田及灌溉溝渠數量成正相關,研究亦發現赤頸鶴巢的微棲地與 生殖成敗無關,在此類型棲息地上生殖失敗的主要原因為農夫活動、淹水 及巢掠食。
除了做為生殖棲息地外,水田也是許多鳥類的活動棲息地。宜蘭縣野 鳥學會(2002)報告指出,宜蘭縣冬山河畔的濕地環境為縣內紀錄最多鳥類數 量的濕地環境,包括大片水田、水澤區,其中又以候鳥為主,冬候鳥如鴨 科、鷸科、鴴科等種類豐富,夏候鳥則以燕鴴為最大量。Tourenq et al.(2003) 研究法國農耕地亦指出,田地淹水時期也會吸引大量的鷸科鳥類聚集。此 外,冬天蓄水的田地是許多冬候鳥重要的活動棲息地(Elphick and Oring, 1998;Tourenq et al., 2001;馬志軍等,2001;Zhijun et al., 2004;Pierluissi, 2006)。另有報告指出,稻田環境不僅容易成為度冬候鳥的活動棲息地,其 中還包括極需保護的物種,如馬志軍等(2001)研究指出,已瀕臨絕種的朱
(Nipponia nippon)冬天主要的覓食區便是水田環境。
由此可知,水田的經營管理雖然改變許多自然生態環境,也創造許多
鳥類棲息的環境。此外,水田可在自然濕地不足時做為物種轉換棲息地的 一種策略(Tourenq et al., 2001)。Zhijun et al.(2004)研究指出,上海崇明島內 一般農耕地因季節性的農耕活動轉換成人工濕地時,可讓許多原本棲息於 自然濕地的物種如紅冠水雞等,在自然溼地喪失後做為補償性棲息地。
此外,亦有研究調查顯示水田的耕作情況對鳥類有影響,並提出重要 的經營管理策略。Elphick(2004)調查結果顯示,冬天的鳥類因不同食性上使 用蓄水或不蓄水農作田做為主要覓食棲息地,故建議冬天採用混合型的農 作經營時可增加種類與數量的使用棲息環境。
由此可知,水田為許多鳥類生殖或活動的棲息環境,故瞭解人為經營 活動中的水田與鳥類族群的活動與生殖之關係,可更深入認識此類棲息地 中鳥類的生態習性,亦可藉以反應環境變動後鳥類的棲息情況。
二、材料與方法
本研究於2006年7月至2007年10月於樣區內使用地區搜尋法(area search) 進行調查。調查方式主要開車或騎車沿著道路以10倍雙筒望遠鏡進行調 查,平均車速為10 km/hr以下。調查路線固定由永美路開始沿著樣區內12條 東西向道路及3條南北向道路開車,沿途進行水田間的搜尋。本樣區的道路 與水田規劃大致利於本調查方法,每兩條可供調查的道路包含兩列水田在 其中,因此調查時可以清楚掌握每塊水田的最遠邊際距離(平均距離約100 m),以便增加調查的精準度。而調查期間若發現紅冠水雞個體有生殖行為,
或可能築巢地之草生地,則改以徒步方式在水田內或茂草區進行巢位的調 查。
(一) 族群與成幼鳥數量調查
以每星期1次的頻率調查族群與成幼鳥數量,本研究為避免稻作生 長期時,因水田內植被茂密無法準確調查到在植被內活動的個體,故
調查次數,以便取得最佳數據,藉以分析瞭解本樣區中紅冠水雞數量 的消長及分布情況。
此外,成鳥與幼鳥的數量主要代表生殖個體數量與當年生殖成功 的個體數量。調查期間紀錄成鳥與幼鳥數量與活動地點,並標示在區 塊調查圖上。成幼鳥的區分以是否具有成鳥羽色而定,成鳥視為生殖 個體,而前額板已有性成熟的外顯象徵(Eens et al., 2000) 但未有成鳥羽 色之亞成鳥,則視該個體是否生殖成功再加以區分為生殖個體或代表 當年生殖成功個體。
(二)資料分析
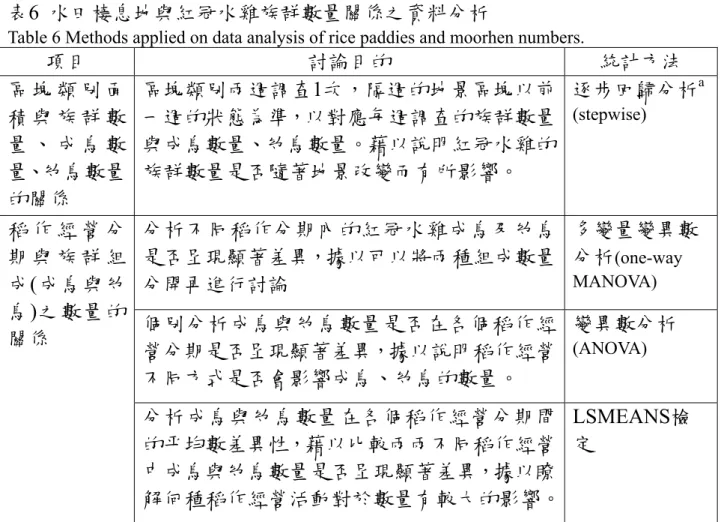
水田棲息地與紅冠水雞族群數量關係之統計方法,討論變因與討 論目的,如表6。
表6 水田棲息地與紅冠水雞族群數量關係之資料分析
Table 6 Methods applied on data analysis of rice paddies and moorhen numbers.
項目 討論目的 統計方法
區 塊 類 別 面 積 與 族 群 數 量 、 成 鳥 數 量、幼鳥數量 的關係
區塊類別兩週調查1次,隔週的地景區塊以前 一週的狀態為準,以對應每週調查的族群數量 與成鳥數量、幼鳥數量。藉以說明紅冠水雞的 族群數量是否隨著地景改變而有所影響。
逐步回歸分析a (stepwise)
分析不同稻作分期內的紅冠水雞成鳥及幼鳥 是否呈現顯著差異,據以可以將兩種組成數量 分開再進行討論
多變量變異數 分析(one-way MANOVA) 個別分析成鳥與幼鳥數量是否在各個稻作經
營分期是否呈現顯著差異,據以說明稻作經營 不同方式是否會影響成鳥、幼鳥的數量。
變異數分析 (ANOVA) 稻 作 經 營 分
期 與 族 群 組 成(成鳥與幼 鳥)之數量的 關係
分析成鳥與幼鳥數量在各個稻作經營分期間 的平均數差異性,藉以比較兩兩不同稻作經營 中成鳥與幼鳥數量是否呈現顯著差異,據以瞭 解何種稻作經營活動對於數量有較大的影響。
LSMEANS檢 定
註:a參考Ritter and Savidge(1999)研究報告
三、結果
(一)水田地景變化與紅冠水雞族群數量的關係 1.稻作經營分期與紅冠水雞族群數量的關係
從稻作經營分期發現紅冠水雞族群數量在不同分期呈現明顯 消長(圖 7)。其中可發現紅冠水雞的族群數量呈現兩個高峰期,各 為稻作生長期(Means+SD=79.30+66.87)與蓄水休耕期(Means+SD
=54.45+23.16)。此兩期的接著稻作經營分期顯著呈現數量上的減 少。
以稻作生長期的紅冠水雞族群數量最高,稻作收割後的收割 休息轉作期之族群數量前期低43%,接著田菁翻田期之族群數量 較前期更低62%。蓄水休耕期的數量次高,但於翻土插秧期的數 量明顯較前期低38 %。
圖7 各個稻作經營各期內紅冠水雞的平均族群數量
Fig.7 Average moorhen number in each cultivating management period 0
10 20 30 40 50 60 70 80 90
收割休息轉 作 期
田菁翻田期 蓄水休耕期 翻土插秧期 稻作生長期
數量(隻次︵
2.區塊類別與紅冠水雞族群數量的關係
本研究為瞭解是否合適棲息地限制紅冠水雞族群數量,故進 一步將 8 種區塊類別與紅冠水雞族群數量進行分析討論(逐步回 歸分析)。結果顯示與紅冠水雞族群數量有顯著相關的地景區塊類 別有3 個區塊類別,分別與無植被有水區塊、高植被無水區塊的 面積呈現正相關,與中植被有水區塊呈負相關(附表 1)。
無植被有水區塊為 10 月至隔年 2 月底的主要地景,亦即是 蓄水休耕時期,而高植被無水區塊為 5 月至 7 月初時的主要地 景,正好與稻作生長期重疊。而族群數量與中植被有水區塊呈負 相關,此類型棲地的出現時間為7 月中旬至 9 月初,正與收割休 息轉作期重疊。
由於影響族群數量的地景明顯不同,故下節以紅冠水雞分布 狀態再加以討論,將各個稻作經營分期的地景狀態與紅冠水雞分 布一併分析討論。
3.稻作分期與紅冠水雞分布的關係
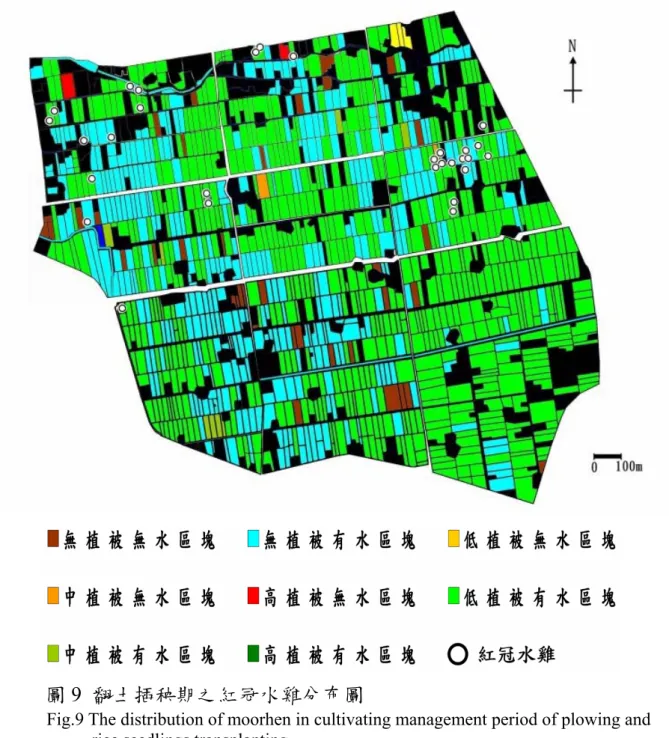
為了瞭解紅冠水雞在不同稻作經營分期內的分布情況,選擇 各個稻作經營分期的期中調查紀錄繪製紅冠水雞分布圖,呈現出 各期的區塊類型之地貌狀態及紅冠水雞個體的分布情況(圖 8-12)。
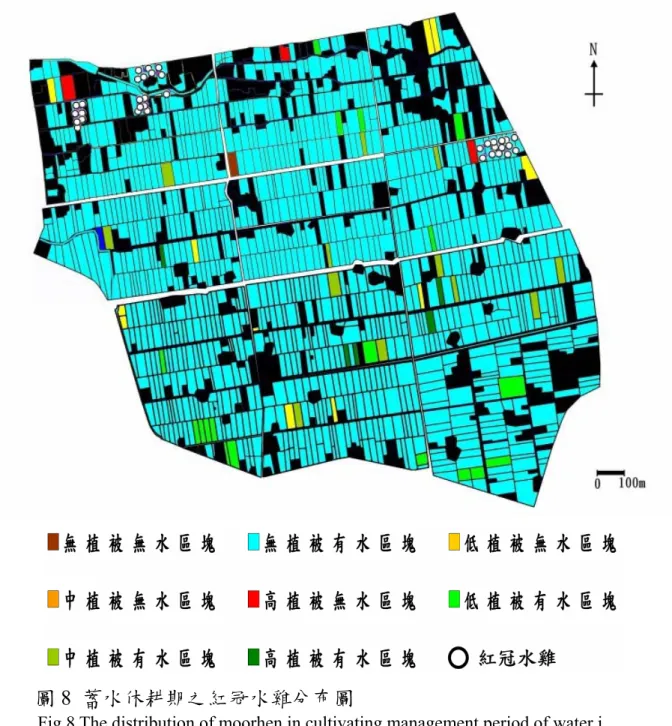
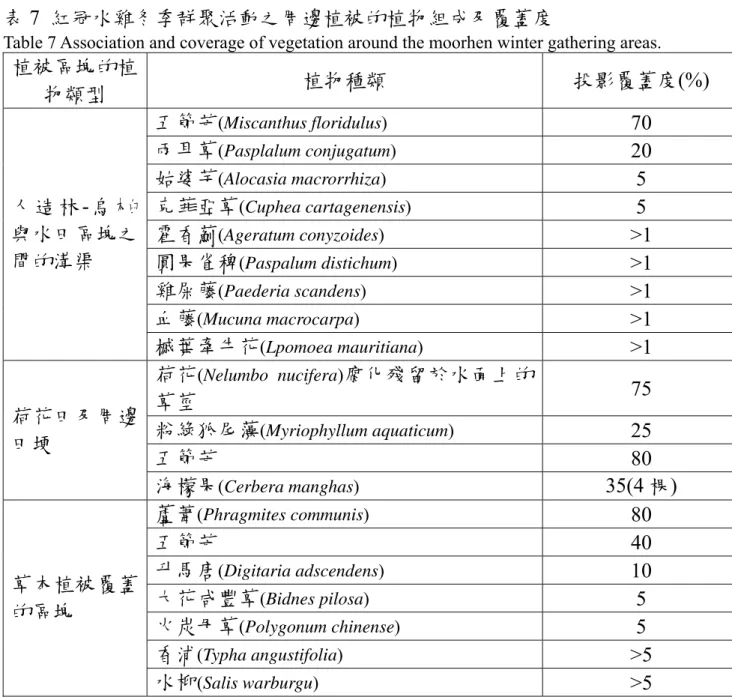
從蓄水休耕期中發現紅冠水雞呈現冬天聚集的現象(圖 8),群 聚的時間約 2006 年 10 月初持續到 2007 年 2 月左右,才開始擴散 開進行生殖。本樣區內有三個主要群聚的地點,周邊都緊鄰著高 覆蓋度的植被區塊。三個群聚點周邊的植被覆蓋呈現不同植物組 成(表 7)。
由翻土插秧期的紅冠水雞分布圖(圖 9)可發現,此時期紅冠水 雞的分布仍呈現聚集的情況,但已有由冬天聚集點向外擴散的現
象。此時地景上陸續有稻作經營活動的擾動及地景上的變化,加 上每次調查與隔次調查的紅冠水雞個體之分布區域不盡相同,因 此推測此時期紅冠水雞雖已經擴散並建立領域,但是受干擾而未 有明顯且固定的棲息領域。
由稻作生長期的紅冠水雞分布圖(圖 10)可發現,地景上紅冠 水雞個體更加分散,每次調查的分布情況則較為固定,因此可以 判定紅冠水雞於此時大致已建立固定領域且進行生殖。
另由收割休息轉作期的紅冠水雞分布圖(圖 11)可發現,此時 水田大都收割完成,紅冠水雞分布呈散佈狀,比較前一期分布圖 (圖 10),發現每個群聚的個體數量較高,而多數是由於親鳥帶著 幼鳥的群聚組織。
再由田菁轉作期的紅冠水雞分布圖(圖 12)來看,此時大量機 械翻土及地貌改變,加上生殖期已結束,可以發現紅冠水雞開始 呈現冬天聚集的情況。
圖8 蓄水休耕期之紅冠水雞分布圖
Fig.8 The distribution of moorhen in cultivating management period of water i mmerging and fallow.
註:以96 年 12 月 16 日紀錄繪製
note:according to the record of 2007/12/16
圖9 翻土插秧期之紅冠水雞分布圖
Fig.9 The distribution of moorhen in cultivating management period of plowing and rice seedlings transplanting.
註:以96 年 3 月 25 日紀錄繪製
note:according to the record of 2007/03/25
圖10 稻作生長期之紅冠水雞分布圖
Fig.10 The distribution of moorhen in cultivating management period of rice growing.
註:以96 年 5 月 20 日紀錄繪製
note:according to the record of 2007/05/20
圖11 收割休息轉作期之紅冠水雞分布圖
Fig.11 The distribution of moorhen in cultivating management period of rice harvesting, and sesbavia planting.
註:以96 年 7 月 14 日紀錄繪製
note:according to the record of 2007/07/14
圖12 田菁翻田期之紅冠水雞分布圖
Fig.12 The distribution of moorhen in cultivating management period of plowing and water flooding.
註:以96 年 9 月 23 日紀錄繪製
note:according to the record of 2007/09/23
表7 紅冠水雞冬季群聚活動之周邊植被的植物組成及覆蓋度
Table 7 Association and coverage of vegetation around the moorhen winter gathering areas.
植被區塊的植
物類型 植物種類 投影覆蓋度(%)
五節芒(Miscanthus floridulus) 70
兩耳草(Pasplalum conjugatum) 20
姑婆芋(Alocasia macrorrhiza) 5
克菲亞草(Cuphea cartagenensis) 5
霍香薊(Ageratum conyzoides) >1
圓果雀稗(Paspalum distichum) >1
雞屎藤(Paederia scandens) >1
血藤(Mucuna macrocarpa) >1
人 造 林- 烏 木 臼 與水田區塊之 間的溝渠
槭葉牽牛花(Lpomoea mauritiana) >1 荷花(Nelumbo nucifera)腐化殘留於水面上的
草莖 75
粉綠狐尾藻(Myriophyllum aquaticum) 25
五節芒 80
荷花田及周邊 田埂
海檬果(Cerbera manghas) 35(4 棵)
蘆葦(Phragmites communis) 80
五節芒 40
升馬唐(Digitaria adscendens) 10
大花咸豐草(Bidnes pilosa) 5
火炭母草(Polygonum chinense) 5
香浦(Typha angustifolia) >5
草本植被覆蓋 的區塊
水柳(Salis warburgu) >5
註:每塊棲地內的植被結構因植物體生長層次不同,故不同層級造成的覆蓋面積加總後 超過100%
由此可知,稻作經營分期內紅冠水雞會呈不同的分布狀態,蓄 水休耕期呈現冬季群聚情況,到了翻土插秧期開始擴散,稻作生 長期分佈更廣,至收割休息轉作期時紅冠水雞個體分佈亦為分 散,但成幼鳥則以生殖族群為單位呈現小群分佈狀態。在田菁翻 田期紅冠水雞的的分布情況再度呈現冬天的群聚行為。
(二)稻作地景變化與紅冠水雞成幼鳥數的關係
本研究為瞭解不同稻作經營活動對於生殖個體是否影響,且是否 因棲息環境條件改變而產生生殖棲息地的不同選擇,故藉由成鳥數(生 殖個體數)與幼鳥數(當年生殖成功個體數)與稻作經營分期進行探討,
以嘗試瞭解紅冠水雞在選擇較佳生殖棲息環境的需求,亦輔助探討紅 冠水雞之巢生殖成敗的不足,以達到完整呈現紅冠水雞在水田棲息地 上整年度的生殖情況。
本人在調查期間發現可以生殖的個體除了成鳥以外,也曾調查到 少部份亞成鳥個體未有成鳥羽色且前額板已有性成熟的外顯象徵,通 常出現在9月份的田菁翻田期,地景上方便觀察該個體是否有生殖築巢 的行為。調查卻僅發現一隻亞成鳥羽色的親鳥有築巢孵卵行為,但最 後棄巢並無生殖成功。故本研究將僅對紅冠水雞的羽色進行區分生殖 組成數量,界定為可生殖個體的成鳥數,以及當年生殖成功個體的幼 鳥數。而生殖季節中幼鳥數通常受成鳥數所影響,即為生殖成功個體 數由生殖個體數所決定。
1.稻作經營分期與生殖組成數量的關係
將紅冠水雞成鳥與幼鳥的數量在不同稻作經營分期內作分 析,發現在不同稻作經營分期內,兩者呈現顯著差異(多變量變異 數分析,λ<0.0001,N=66,圖 13)。顯示兩者呈現顯著差異,故分 別進行兩者數量與稻作經營分期的分析。