國立臺灣大學醫學院暨工學院醫學工程學研究所 碩士論文
Graduate Department of Biomedical Engineering College of Medical and College of Engineering
National Taiwan University Master Thesis
受電場而極化的脂膜筏引導纖維母細胞之方向性移動 Electric Field-induced Lipid Raft Polarization Guide
Fibroblast Directional Migration
林栢江 Bo-Jiang Lin
指導教授:趙本秀 博士
Advisor: Pen-Hsiu Grace Chao, Ph.D.
中華民國 102 年 7 月
July, 2013
口試
I
試委員審定 定書
II
序言
工業技術與醫學知識的對撞,激盪出宣示未來新希望的煙火,也散發著那使
我著迷的炫彩,而臺灣大學醫學工程所給予了我一個契機,讓我得以參與這歷史
的進程,最要感謝的人,是指導我何謂科學家思維的教授─趙本秀老師,「要對data
忠心」正闡述著趙老師對於假說的嚴謹,也是其要求麾下研究生應有的態度,兩
年碩士班生涯走過,也因有宋承修和曾筱筠的陪伴才能如此順遂與歡樂,好夥伴
的相互扶持,已畢業的鄭祐甄、蔡承憲、馮嘉襄、徐向儀、沈守謙和張維仁的教
導,讓我學習到待人處事上的眉角,也感謝博士班學長溫新民的坐鎮,讓 lab501
運轉率百分百;Jeff 除了是個有品味的飯友也是英文會話的家教;已提早退席的游
婉婷著實地讓平凡生活增加不少趣事,此外,也感謝王兆麟老師麾下的崔哲豪、
王彥凱、陳志維以及徐于鈞等人平日增添本實驗室風采之外,也讓我跟隨順道拜
訪夏威夷的國際會議團,一路上除了驚險萬分也是收穫滿分,本論文得以完成則
有泰半來自生命科學院核心實驗室莊以君的體諒,才能有著高品質的 confocal 螢
光照片,還有醫學院第一共同研究室許華蔓的操作,才得順利在ORS 上發表不同
種類電場如何影響細胞的論文,也感謝暑期研究生何玉樺辛苦量化 lipid raft 的分
佈和Alex 熬夜撰寫的 lipid raft 模擬程式,使本論文更加完整,因為有了過去的你
們,才成就了現在的我,由衷感謝。
III
摘要
當微生物或細胞受到電場影響而展現出順著電場方向移動,此為趨電性。許
多研究指出膜蛋白的分佈狀況會影響趨電性的行為,在先前的研究中指出 α2β1
integrin 受電場影響而有極化的現象,且其在交流電與直流電刺激中有著相反的
分佈情形,因此本研究想探討細胞膜上的微結構─脂膜筏,在纖維母細胞受交流
電或直流電影響後,如何控制細胞的移動性,結果中發現,脂膜筏受電場影響而
聚集,且跟 α2β1 素蛋白有著相似的分佈,此外,九成的 α2β1 素蛋白坐落在脂膜
筏上,如果破壞脂膜筏會抑制纖維母細胞的趨電性行為,另外,抑制caveolin-1 的
基因表現也會抑制往負極向移動和 RhoA 的分佈,因此從本研究的研究中,可以知
道脂膜筏會藉由 caveolin-1 影響纖維母細胞趨電性的表現。
關鍵字:脂膜筏、素蛋白、細胞移動、電場、趨電性
IV
Abstract
Galvanotaxis is a phenomenon in which microorganisms migrate in response with
the electric current. Most studies indicate that the redistribution of plasma membrane
proteins guides cell directional motility. The previous study showed that α2β1 integrin
polarizes with AC and DC electric fields. This study shows when fibroblasts are
stimulated by an electric field, lipid rafts polarize and the polarization coincides with
asymmetrically-distributing α2β1 integrins. Disruption of lipid rafts inhibits EF-induced
directional migration. The caveolin-1 knockdown inhibits cell directional motility and
RhoA polarization. The results indicate that lipid raft is a mechanosensor to EF
stimulation and lipid raft polarization lead to integrin and caveolin-dependent
directional migration.
Key Words: lipid raft, integrin, migration, electric field, galvanotaxis
V
Contents
口試委員審定書 ... I
序言 ... II
摘要 ... III
Abstract ... IV
List of Tables ... VII
List of Figures ... VII
Chapter 1. Introduction ... 1
Chapter 2. Materials and Methods ... 5
2.1 Primary Porcine Fibroblasts Culture ... 5
2.2 Microfluidic channel Fabrication ... 5
2.3 RNA interference ... 6
2.4 Pharmacological Treatment ... 7
2.4.1 Cholesterol Depletion ... 7
2.4.2 Plasma Membrane Solidification... 7
2.4.3 Integrin functional block ... 7
2.5 Electric field stimulation ... 7
2.6 Fibroblast Behavior Quantification ... 8
2.7 Lipid Raft Labeling ... 8
VI
2.8 Reverse Transcription Polymerase Chain Reaction... 8
2.9 Immunofluorescence staining ... 9
2.10 Image Analysis ... 9
2.11 Statistics ... 10
Chapter 3. Results ... 11
3.1 EF-induced lipid raft redistribution ... 11
3.2 Caveolin-1 signaling pathway ... 12
Chapter 4. Discussion ... 14
Reference ... 34
VII
List of Tables
Table 1. Primers used in this study (Primer-BLAST, NCBI) ... 17
Table 2. siRNA for caveolin-1 (Qiagen) ... 18
List of Figures
Figure 1. Galvanotaxis Chamber (Cannizzaro et al. 2009). ... 19Figure 2. Microfluidic Channel. ... 20
Figure 3. Directionality and Asymmetry Index ... 21
Figure 4. Lipid raft polarization ... 22
Figure 5. Coincide between lipid raft and α2β1 integrin ... 23
Figure 6. α2β1 integrin riding lipid raft ... 24
Figure 7. Lipid raft polarization disrupt by Ch and MβCD ... 26
Figure 8. Ch and MβCD impair galvanotaxis ... 27
Figure 9. Cav1 polarize under electric field stimulation. ... 29
Figure 10. Cav1 knockdown by RNAi. ... 30
Figure 11 Caveolin-1 knockdown disrupt α2β1 integrin distribution ... 31
Figure 12. Cav1 knockdown reduce directionality. ... 32
Figure 13. RhoA polarization induced by EF is impaired by RNAi. ... 33
1
Chapter 1.
Introduction
Most studies indicate that endogenous electric currents advanced the nerve
regeneration and facilitate the wound healing [1-3]. While epithelia are injured,
trans-epithelial potentials (TEP, [4]) on the wound drops and the potential differences
drive the electric current toward the wound center. For example, an electric field
measured 42mV/mm in cornea wound and an electric current generated 10 μA/cm2 at
peak, persisted at 4 μA/cm2 in human skin [4, 5]. That migration of cell in response to
an electric field is a phenomenon, called galvanotaxis or electrotaxis. Galvanotaxis was
first observed on leukocytes by Dineur in 1891. The displacement of a cell depends on
current density, voltage, and solute in the medium. Besides that, cell morphology will
change under electric field. For example, fibroblasts not only increased speed and
directionally migrated toward anode but also rearranged cytoskeletons perpendicular to
electric currents and elevated the ECM-related gene expression, like type I collagen,
while exposed on electric field [1, 6]. Consequently, many studies have attempted to
delineate the mechanisms how cells sense an electric field.
Due to the high impedance of plasma membrane, how electric field sensing
mechanism is following on four physical hypotheses: asymmetric ion flow,
2
mechanical force sensor, electro-osmosis and electrophoresis [7]. Asymmetric ion
flow is that the ion redistribution induced by electric field. Mechanical force sensor is
that charge membrane channels open gate by electric field. Electro-osmosis is water
potential owing to plasma membrane attracting ion. Electrophoresis is that charge
particles move along the electric field. Recently studies reveal that the behavioral
response to electric field is attributed to the polarization of plasma membrane
molecules [8]. Some plasma membrane proteins, like integrin and EGF receptor, are
founded to asymmetrically distribute under electric field [9, 10]. Impaired the function
of integrin can disrupt the directional motility of cells [9]. Integrin, a heterodimer
containing an α and a β subunit, is a major families of cell adhesion. Its function can
be divided into two parts. One is to assembly with cytoskeleton and signal molecules,
like PI3 kinase and FAK [7, 11]. The other is to combine with extracellular matrix as a
focal adhesion, a main component of cell adhesion and migration [12, 13]. Therefore,
spatial downstream signaling pathway activated by integrin is a hub to reveal the
intrinsic mechanism of galvanotaxis.
Rho family of small GTPase, including Rho, Rac1 and Cdc42, is G-protein
which transits the extracellular message as a cellular signal molecule [14]. While
migrating, cell protrude their actin-based lamellipodium, where integrin-based
adhesion activate RhoA [15]. RhoA-related contract leads cells to the orientation into
3
which lamellipodia project [16]. Polarized distribution of α2β1 integrin has guided
ligament fibroblasts via RhoA in our previous study. However, fibroblasts also exhibit
polarization in applied AC fields, despite the asymmetrical nature of the alternating
current. It implies that a small structure with fast response time (half the applied 60Hz
duration) can initiate the mechanism to polarize integrin and direct cell migration.
“Membrane rafts are small (10–200 nm), heterogeneous, highly dynamic, sterol- and sphingolipid-enriched domains that compartmentalize cellular processes. Small rafts can sometimes be stabilized to form larger platforms through protein-protein and protein-lipid interactions” report by Pike [17]. Lipid raft is a key structure for
endocytosis [18], membrane traffic [19] and cell migration [20]. The property of lipid
raft is to cluster into huge while stimulated [21]. Some reports show that integrin
activating by ECM cluster on lipid raft [19]. The impairment of the lipid raft inhibits
the polarization of integrin [22]. Lipid raft contains caveolin-1, which interact with
different membrane protein-related signaling pathway and form a caveolae [23]. For
example, caveolin-1 bind with β1 integrin to activate RhoA though inactivation of
p190RhoGTPase [24]. The caveolin-1 also control the β1 integrin activation though
Cbp-Csk-Src pathway while forming the caveolin-1/β1 integrin complex [25]. Studies
indicate that caveolin-1 is crucial for directional migration by activation RhoA [26,
27]
4
Therefore, we hypothesize that lipid raft taking α2β1 integrin clusters on the
leading edge of migration direction and guides cells to respond to electric fields
though caveolin-1. In this thesis, we aim to figure out the role of lipid raft in
galvanotaxis and understand the function of lipid raft on the integrin-dependent RhoA
signaling pathway. The results showed that depletion of lipid raft resulted in the loss
of oriented motility but no significant change of speed. It also impaired the polarized
distribution of integrin. The knockdown of caveolin-1 disrupted galvanotaxis. We
delineated a brief EF-induced cell signaling pathway, which lipid raft was an electric
currents sensing unit and a complex combined with integrin to activate RhoA.
5
Chapter 2.
Materials and Methods
2.1 Primary Porcine Fibroblasts Culture
Anterior cruciate ligament fibroblasts were harvested from young porcine knee
joints. Anterior cruciate ligament (ACL) wash in PBS then is torn into pieces. The
explants were cultured in 3cm dishes with 4.5 g/L D-glucose DMEM with 10% FBS
(Invitrogen), 1% penicillin/streptomycin (Gibco) and 1% HEPES (Biowest), under 37
℃ incubator with 5% CO2. Growth media were changed every 3 dayuntil confluence.
For electric field stimulation studies, ACL fibroblasts were resuspended by 2 min
incubation with 0.25% trypsin-EDTA (Gibco) and seeded on the microfluidic channel
at 4 × 10 cells/cm2 or on sterile glass slides at 4 × 10 cells/cm2 for 2 hours in
incubator. The slides combined with a galvanotaxis chamber for electric field
stimulation (Figure 1).
2.2 Microfluidic channel Fabrication
The straight microchannels made of polydimethylsiloxane (PDMS) by soft
lithography procedures. SU8 photo resist (Gersteltec Sarl) spread on silicon wafer by
a spinning coater then exposed with UV lights though mask to develop patterns.
6
PDMS was molded over the SU8 master for two days then separated. PDMS channels
were then cured at 70 ℃ overnight. After cooling, the PDMS channels were
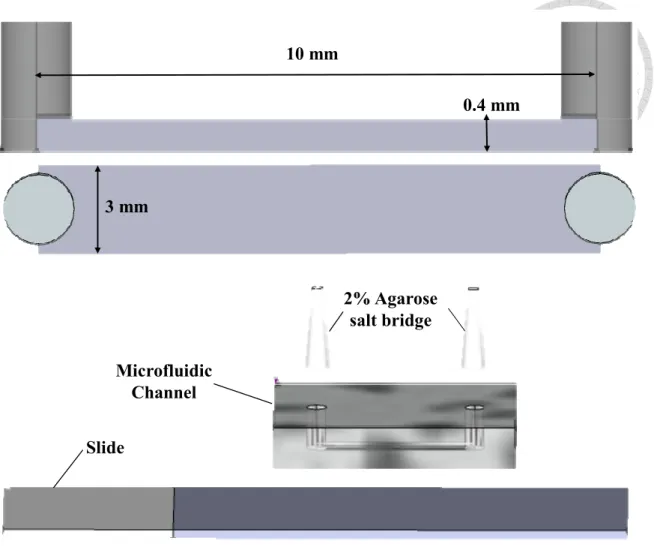
punctured for inlets and outlets and attached to glass substrate [28]. Channels are 3 mm
wide, 400 μm high and 10 mm long (Figure 2). Fresh DMEM were added into
microchannel every hour in case of evaporation and connect the agarose salt bridge on
the poles of chamber after 2 hours incubation.
2.3 RNA interference
The RNA interference for caveoline 1 (Cav1) was performed by small interfering
RNAs (siRNA) and AllStars negative siRNA modified with alexa flour 488
(Cat#1027284, Qiagen) was used as control. The siRNA, is complement to Cav1, was
designed by Qiagen and the sequence is the following Table 2. ACL fibroblasts were
seeded on 6-well dishes at 2 × 10 cells/well. siRNA diluted on 100μl serum-free
DMEM then were mixed with 6μl HiPerFect® transfection reagent (Cat#301702,
Qiagen) by vortexing 10 sec. After 10 minutes on siRNA misture standing, every well
was added with 150 ng si-Cav1 or 75 ng Negative Control siRNA for 2 days then
washed and reseed on the sterile glass for electric field stimulation.
7
2.4 Pharmacological Treatment
2.4.1 Cholesterol Depletion
Cells were washed with PBS and incubated for 1 h at 37°C with serum-free
DMEM of 5 mM methyl β-cyclodextrin (MβCD, Cat#C4555, sigma,[29]).
2.4.2 Plasma Membrane Solidification
Cholestryl hemisuccinate (Ch, Cat#C6512, sigma), an analog of cholesterol,
were dissolved in dimethylsulfoxide and diluted with PBS at 0.5% final
concentration. Cells were preincubated with1 mM ChH for 30 minutes then
washed [30].
2.4.3 Integrin functional block
Anti–porcine integrin α2β1 antibody (Cat#mab1998, Invitrogen, 1 mg/ml)
diluted with PBS at 2.8 μg/ml. Cells were treated with it for 30 min then
washed [9].
2.5 Electric field stimulation
Constant direct current (DC) EF was applied at a field strength of 6 V/cm
parallel to the microchannel direction and galvanotaxis chamber using a Keithley
Source Meter and alternative current (AC) sinusoid waves (sin) was applied at a peak
8
intensity of 1.2 V at 50 Hz using a custom stimulator (Dynaprog, MingQuo, Taiwan).
2.6 Fibroblast Behavior Quantification
Cells were captured by Cannon EOS every 15 minutes and calculated their
position by determining the centroid at the initiation and the end. The displacement and
orientation were measured and represented as velocity and directionality. The
directionality was descripted by cosine θ, where θ was the angle between the EF field
axis and the cell translocation vector. θ = 0˚ was assigned to the cathode and 180˚ was
assigned to the anode. Therefore, the range of cosine θ was between +1 and -1 and an
index of directed migration (Figure 3.a).
2.7 Lipid Raft Labeling
Alexa Fluor® 555 conjugate-cholera toxin B (CTxB, Cat#C-22843, Molecular
Probes), binding to the GM1 gnagliosides, was diluted with chilled complete growth
medium at 1μg/ml. After the EF Stimulation, channels were filled with chill complete
growth medium. It was replaced with CTxB for 10 minutes at 4℃ then wash. Finally,
4% formaldehyde rinse it for 30 minutes and maintain the temperature on 4℃.
2.8 Reverse Transcription Polymerase Chain Reaction
Cells were lysed by TRIzol and reverse transcription was performed by using
superscriptTM III transcriptase in a thermal cycler (Bioner) and amplification was
9
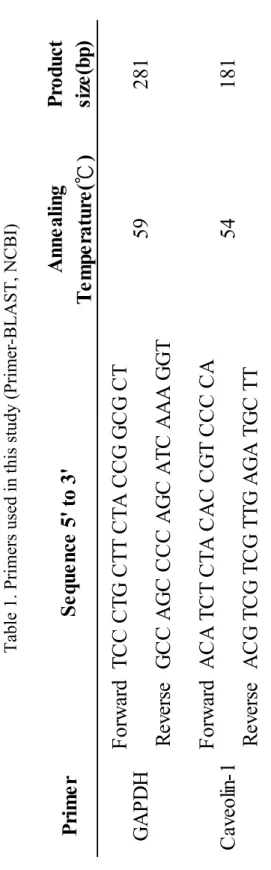
performed by GoTaq® green master mix (Promega). The primers of caveolin 1 and
GAPDH are as following Table 2 (Primer-BLAST, NCBI). Amplified cDNA
fragments were resolved by electrophoresis on 2% agarose gel staining ethidium
bromide. The intensity of band was detected by ultraviolet illumination, quantified by
ImageJ and normalized against that of GAPDH.
2.9 Immunofluorescence staining
Upon completion of fixation, cells were permeabilized by 0.3% triton X-100 for 5
mins and blocked with 5% FBS in PBS for 1 hour after wash trice. Cells were incubated
overnight in 4℃ with primary monoclonal anti-α2β1 integrin antibody (1:200,
Invitrogen) or primary monoclonal anti-caveolin 1 antibody (1:200, Cytoskeleton) or
primary monoclonal anti-RhoA antibody (1:200, Cytoskeleton). Goat anti-mouse alexa
flour 488-conjugated antibody (1:250, Invitrogen) supplemented with 5% FBS for 1
hour and so was goat anti-rabbit alexa flour 555-conjugated antibody (1:250,
Invitrogen). SlowFade® gold antifade reagent (Invitrogen) was used to mount the
coverslips onto the glass slides. Slides were observed on confocal microscopy (Leica
TCS SP5).
2.10 Image Analysis
Each cell was divided into four equal quadrants and the mean fluorescence
10
intensity was measured by an image process program developed from LabVIEW 2011
SP1 (National Instruments). Asymmetry Index (AI) was calculated by subtracting the
normalized intensity of the region facing cathode from the region facing anode, and
normalized to the overall average intensity of the cell. A positive value of AI indicates
cathodal distribution and a negative value of AI indicates anodal distribution (Figure
3.b).
2.11 Statistics
R 3.0 (The R Foundation for Statistical Computing) was used to perfume
one-way ANOVA with Tukey’s HSD post hoc test under α = 0.05. Data represent
mean ± standard error.
11
Chapter 3.
Results
3.1 EF-induced lipid raft redistribution
In our previous study, we found that α2β1 integrin showed different distribution
between DC and AC. It is possible that lipid raft, a larger size microdomain than protein,
floating on the plasma membrane carry these parallel to EF. To investigate whether
lipid raft response to EF, we quantified the fluorescence intensity of cholera toxin
B-labeled lipid raft distribution with time. The lipid raft polarized and coincided with
α2β1 integrin (Figure 4 and Figure 5). The α2β1 integrin polarization is consistent with
previous study. It indicated that lipid raft not only reacted to EF but also correlated with
α2β1 integrin.
To delineate the relationship between lipid raft and α2β1 integrin, we observed the
α2β1 integrin distribution while lipid raft deconstruction and the lipid raft distribution
while functional block on α2β1 integrin. We found raft impairment caused the α2β1
integrin randomly distribute but integrin block did not affect the lipid raft polarization
(Figure 6). It indicated that α2β1 integrin were shipped by lipid raft.
Based on our hypothesis that lipid raft can move on the plasma membrane with
low-drag, we investigate the role of lipid raft distributed between on cell migration.
12
Pharmaceutical treatments used cholesteryl hemisuccinate to reduce membrane fluidity
and methly β-cyclodextrin to impair the lipid raft structure. Significantly, lipid raft
distribute asymmetrically when exposed to electric field and randomly when treated
with these chemicals (Figure 7).
We measured the displacement and direction of fibroblasts with cholesteryl
hemisuccinate and methly β-cyclodextrin (Figure 8.a). Lipid raft disruption and
membrane solidification did not vary the velocity; however, they did inhibit the
directional migration induced by EF (Figure 8.b).
3.2 Caveolin-1 signaling pathway
We further investigated whether or not caveolin-1 played a role in α2β1 integrin
and lipid raft with electric field stimulation. Caveolin-1 polarized while DC electric
field stimulation (Figure 9). Knockdown of caveolin-1 expression by RNAi impair
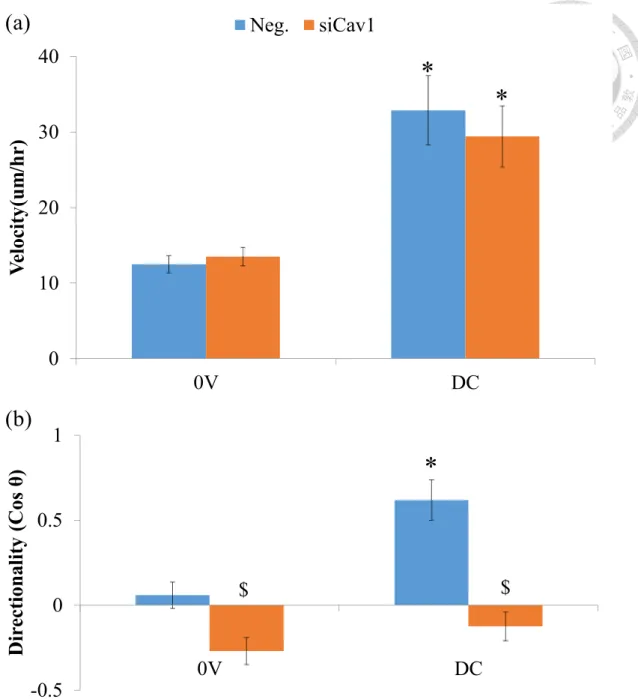
cell directionality but not impact on cell motility (Figure 10 and Figure 12).
Knockdown caveolin-1 show no effect on caveolin-1 polarization but impair integrin
distribution; however, integrin functional block show no effect on lipid raft
polarization but impair caveolin-1 distribution (Figure 9 and Figure 11). RhoA
polarization induced by electric field also disrupt while RNAi treatment and lipid raft
deconstruction (Figure 13). It seems that the polarization of caveolin-1 and α2β1
integrin coincide with quantity of caveolin-1 and function of α2β1 integrin. The data
13
imply that caveolin-1 on the lipid raft could control galvanotaxis though RhoA and
cooperation with α2β1 integrin.
14
Chapter 4.
Discussion
It is evident from the results that overall, lipid rafts polarization induced by
electric field guide cell migration though caveolin-1. That electric field causes the
lipid rafts polarization correspond with that lipid raft can merge into large and stable
structure under stimulation. The polarization of lipid raft shares a similar pattern with
these of α2β1 integrin. These consist with previous finding, which show that α2β1
integrin polarize under electric field stimulation [9]. As previous research by Leitinger
indicated, lipid raft localization are relevant to integrin cluster [22]. The α2β1 integrin
colocalize with lipid raft. The deconstruction of lipid raft impairs the redistribution of
α2β1 integrin; however, the function inhibition of α2β1 integrin shows no change on
the polarization of lipid raft. It is indicated that lipid raft regulate the distribution of
α2β1 integrin with electric field stimulation.
According to Hart’s formula, the force from membrane, environment viscosity
can affect the electric force exerting on the membrane molecules [31]. The drag force
and viscous force could influence the lipid raft polarization. The larger size of lipid
raft causes the higher drag force from membrane. Furthermore, the huger lipid raft
can bear more plasma membrane proteins. The possible mechanism on AC sin wave
15
electric field is that the merge of lipid raft induced by electric field could reduce their
diffusion rate. Lipid rafts gather with each other at beginning then stay behind at
reverse electric field. It is reasonable that the larger lipid rafts become huger due to
the possibility of their contacting with small one. As hypothesized, lipid rafts polarize
at the cell area to which the initial 16ms current orient under AC sin wave electric
field stimulation. The opposite initial currents compared to DC electric field
determine the orientation of lipid raft redistribution. This finding reveals that lipid raft
can polarize at briefly electric field exposure time.
As expected, the increase of membrane viscosity by cholestryl hemisuccinate can
inhibit the lipid raft polarization not only on AC but also on DC. Consequently, the
results show that the lipid rafts size and the membrane viscosity affect lipid raft
diffusion rate. Lipid raft clustering is dependent on the initial orientation of current
due to the change of diffusivity.
Our results show that knockdown of caveolin-1 can impair galvanotaxis. How
caveolin-1 affect cell migration is that Rey-Barroso delineates that caveolin-1 control
focal adhesion and migration though activating FAK and β1 integrin [25]. Moreover,
the knockdown of caveolin-1 affect integrin polarization and functional block of α2β1
integrin influence caveolin-1 distribution. It imply that integrin and caveolin
cooperate with each other [24].
16
It also indicated that RhoA polarization induced by electric field is impaired by
caveolin-1 knockdown and lipid raft disruption. RhoA cause actin-based contraction
and lead to cell migration. RhoA is located on fibroblasts podosomes, which rich in
F-actin related to cell adhesion [32]. Therefore, the RhoA location affects the cell
motility. Our results show that lipid raft can affect directional migration though
controlling RhoA polarization.
We find that lipid raft polarize under electric field stimulation. The polarization
guides cell directional migration though caveolin-1. The initial orientation of electric
current determines the area at which lipid raft distribute and the direction to which
cell migrate. In future study, we would investigate the signaling mechanism of
caveolin-1 and RhoA. For instance, previous research indicates that
integrin/caveolin-1 complex could activate RhoA though p190 RhoGAP [24].
17
17
Table 1. Primers used in this study (Primer-BLAST, NCBI)
Pr im er An ne al in g Te m pe ra tu re (℃ ) P ro duc t si ze (bp) F or w ar d T CC CT G CT T CT A CCG G C G CT R ev er se G CC A G C CCC A G C A T C A A A G G T F or w ar d A CA T C T CT A CA C CG T CCC CA R ev er se A C G TC G TC G TTG A G A TG C TT
GAP D H Ca ve ol in -1
S eque nc e 5 ' t o 3 ' 59 281 54 181
18
18
Table 2. siRNA for caveolin-1 (Qiagen)
N am e S eq ue nc e 5' t o 3' Co nc . (n m ol /tu be ) T ar ge t C T GGAAT A AAT T C A AAT T C T T S en se s tr an d GGAAUAAAUUC A AAUUC UUUU An tis en se s tr an d AAGAAUUUGAAUUUAUUC C A G 20
Figu
Cell
cham
thou
med
brid
ure 1. Galva
ls are depos
mber. The o
ugh the gla
dium during
dge [33].
anotaxis Ch
sited on the
objectives o
ass window
g EF stimul
hamber (Can
center of th
of the phase
w. The chan
lation and c
19
nnizzaro et
he glass slid
e contrast m
annels filled
connect to s
al. 2009).
de, which is
microscope
d with DM
saline reserv
s fitted into
observe the
MEM mainta
voirs with 2
o the transpa
e cell behav
ain cells u
2% agarose arent
viors
under
e salt
20
10 mm
0.4 mm
3 mm
Slide
Microfluidic Channel
2% Agarose salt bridge
Figure 2. Microfluidic Channel.
It is made of MEMS. The channel is 0.4 mm height, 10 mm length and 3 mm width.
Two pores are punctured as an inlet and an outlet. The channel combines with glass
slide and connects to power supply with 2% agarose salt bridge.
Figu
(a)
cell
dire
quan
cell
asym
in th
side
ure 3. Direc
The cell di
migration
ectionality. (
ntified. Q1
. The differ
mmetric ind
he cathodal
e.
a.
○
b.
ctionality an
isplacement
and theses
(b) Cell is d
represent th
rence betwe
dex (AI). Th
l side and t
○
+nd Asymme
t from initia
s projecting
divided into
he cathodal
een Q1 and
he positive
the negative
Q3
21
etry Index
ation to the
g on the e
o four areas
l side of cel
d Q3 is a sy
value of AI
e value of A
θ
Q2
Q4
e end of sti
electric fiel
s then the m
ll and Q3 re
ymbol of the
I means the
AI means th
Q1
mulation is
d axis are
mean fluores
epresent the
e target dist
majority of
hese cluster
○
-s represente
represente
scence inten
e anodal sid
tribution, ca
f protein clu
rs in the an ed as
d as
nsity
de of
alled
uster
nodal
Figu
Lipi
sign
vs. 0
ure 4. Lipid
id raft labe
nificantly til
0V, n = 12-3
d raft polariz
led with CT
ll electric fi
33)
zation
TxB polariz
eld stimulat
22
zed gradual
tes fibrobla
lly at catho
sts for 60 m
dal side of
minutes. (* P
f fibroblasts
P < 0.05 Gro and
oups
Figu
IntR
repr
colo
loca
ure 5. Coinc
Raft means
resenting α
ocalization
ate on α2β1
cide betwee
s α2β1 int
α2β1 integri
coefficient,
integrin.
en lipid raft
egrin locat
in colocaliz
92% α2β1
23
and α2β1 in
ted on lipi
ze with red
1 integrin lo ntegrin
id raft, vic
d, lipid raft
ocate on lip
ce reversa.
ft. Accordin
pid raft and
. Green si
ng to Mand
d 87% lipid ignal
der’s
d raft
24
Figure 6. α2β1 integrin riding lipid raft
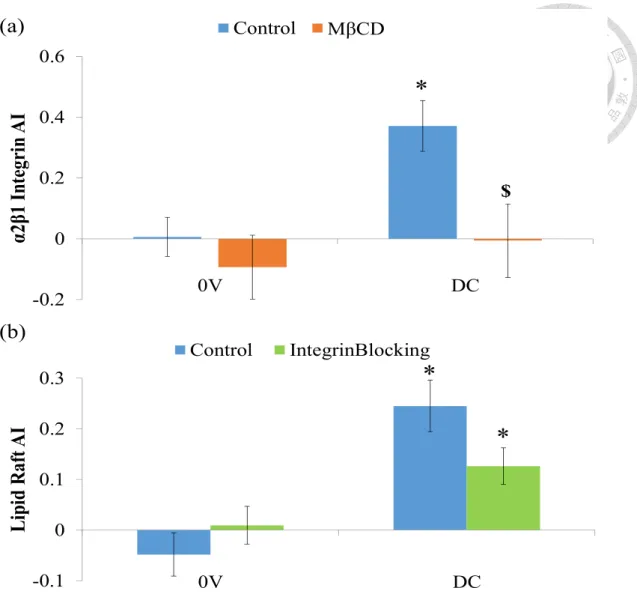
(a) Electric field induces the AI increase of α2β1 integrin and MβCD impair that. (b)
Anti-α2β1 integrin antibody blocks the integrin function but does not affect the AI of
lipid raft. (* P < 0.05 DC vs. 0V, $ P < 0.05 Groups vs. Control, n = 21-50) (a)
(b) -0.2
0 0.2 0.4 0.6
0V DC
α2β1 Integrin AI
Control MβCD
*
$
-0.1 0 0.1 0.2 0.3
0V DC
Lipid Raft AI
Control IntegrinBlocking
*
*
25
26
Figure 7. Lipid raft polarization disrupt by Ch and MβCD
The asymmetric distribution (AI) of lipid raft results from DC or AC. Significantly,
the AI of DC drop while treated with Ch and the AI of AC approximate to 0 while
treated with MβCD (* P < 0.05 Groups vs. 0V , $ P < 0.05 Groups vs. Control , &
P<0.05 Groups vs. DC, n = 17-118).
27
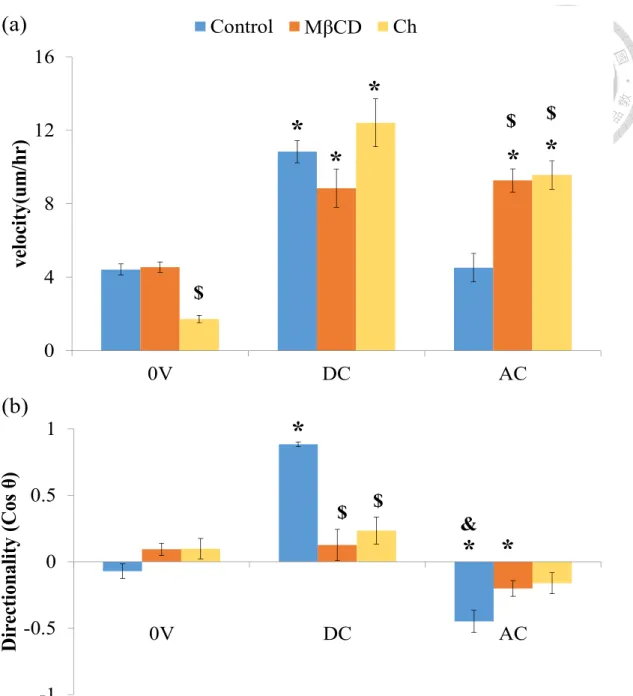
Figure 8. Ch and MβCD impair galvanotaxis
(a) DC EF stimulation enhances the fibroblasts’ migration speed but AC shows not
difference with 0V. No significant difference between control and the groups treated
with MβCD or Ch when fibroblasts exposed on 6V/cm. However, MβCD or Ch
improve the velocity simulated by AC. It seems that MβCD or Ch could enhance cell
performance induced by AC. (b) The directionality simulated by DC is higher than 0V
(a)
(b)
0 4 8 12 160V DC AC
velocity(um/hr)
Control MβCD Ch
*
* *
$ $
$
*
*
-1 -0.5 0 0.5 1
0V DC AC
Directionality (Cos θ)
*
*
$
*
$
&
28
and by AC is more negative than 0V. It represents that the majority of fibroblasts
migrate toward cathode while DC electric field stimulation and toward anode while
exposed on AC. The MβCD and Ch could reduce the DC directionality and increase
the AC directionality. Therefore, pharmaceutical treatments inhibit directional
migration induced by DC or AC. (* P < 0.05 Groups vs. 0V, $ P < 0.05 Groups vs.
Control, & P<0.05 Groups vs. DC, n = 42-171)
Figu
The
The
cave
n =
ure 9. Cav1
e IF images
e functional
eolin-1 show
14-42)
polarize un
show the m
block on α
w no affect
nder electric
majority of r
α2β1 integri
t. (* P < 0.0
29
c field stimu
red signal, c
in decrease
05 Groups v
ulation.
caveolin-1,
caveolin-1
vs. 0V, $ P <
distribute a
AI but the
< 0.05 Grou
at cathodal
knockdow
ups vs. Con side.
n on
ntrol,
Figu
Gen
targ
P <
ure 10. Cav
ne expressio
geting to cav
0.05 Group
v1 knockdow
on level repr
veolin-1, de
ps vs. Contr
wn by RNA
resents the l
ecrease a ha
rol)
30
Ai.
light intens
alf expressio
ity of band
on level of
using by PC
caveolin-1
CR. The siR
than contro RNA,
ol. (*
Figu
Fibr
(* P
ure 11 Cave
roblasts wit
P < 0.05 Gro
eolin-1 knoc
th caveolin-
oups vs. 0V,
ckdown disr
1 knockdow
V, $ P < 0.05
31
rupt α2β1 in
wn show low
Groups vs.
ntegrin distr
wer integrin
. Control, n
ribution
n AI than ne
= 20-26)
egative conttrol.
32
Figure 12. Cav1 knockdown reduce directionality.
(a) Fibroblasts increase their migration velocity whether or not the caveolin-1
knockdown. The expression level of caveolin-1 do not alters cell motility.
(b) Caveolin-1 knockdown inhibit the directionality. (* P < 0.05 Groups vs. 0V, $ P
< 0.05 Groups vs. Control, n = 40-95)
(a)
(b)
0 10 20 30 400V DC
Velocity(um/hr)
Neg. siCav1
* *
-0.5 0 0.5 1
0V DC
Directionality (Cos θ)
$
*
$
Figu
Rho
com
of R
ure 13. Rho
oA polarize
mpared to th
RhoA. ($ P <
oA polarizat
at cathode
he negative c
< 0.05 Grou
tion induced
and AI of R
control. The
ups vs. Cont
33
d by EF is im
RhoA increa
e MβCD an
ntrol, n = 4-3
mpaired by
ase while ce
nd siRNA fo
30)
RNAi.
ll exposed t
or caveolin-
to electric f
1 reduce the field
e AI
34
Reference
1. Guo, A., et al., Effects of Physiological Electric Fields on Migration of Human Dermal Fibroblasts. Journal of Investigative Dermatology, 2010. 130(9): p.
2320-2327.
2. Song, B., Nerve regeneration and wound healing are stimulated and directed by an endogenous electrical field in vivo. Journal of Cell Science, 2004.
117(20): p. 4681-4690.
3. Sun, Y.-S., S.-W. Peng, and J.-Y. Cheng, In vitro electrical-stimulated wound-healing chip for studying electric field-assisted wound-healing process.
Biomicrofluidics, 2012. 6(3): p. 034117-12.
4. Zhao, M., Electrical fields in wound healing-An overriding signal that directs cell migration. Semin Cell Dev Biol, 2009. 20(6): p. 674-82.
5. Zhao, M., et al., Electrical signals control wound healing through phosphatidylinositol-3-OH kinase-[gamma] and PTEN. Nature, 2006.
442(7101): p. 457-460.
6. Chao, P.-h.G., et al., Effects of Applied DC Electric Field on Ligament Fibroblast Migration and Wound Healing. Connective Tissue Research, 2007.
48(4): p. 188-197.
7. Allen, Greg M., A. Mogilner, and Julie A. Theriot, Electrophoresis of Cellular Membrane Components Creates the Directional Cue Guiding Keratocyte Galvanotaxis. Current Biology, 2013(0).
8. Cho, M.R., et al., Induced redistribution of cell surface receptors by alternating current electric fields. The FASEB Journal, 1994. 8(10): p. 771-6.
9. Tsai, C.-H., B.-J. Lin, and P.-H.G. Chao, α2β1 integrin and RhoA mediates electric field-induced ligament fibroblast migration directionality. Journal of Orthopaedic Research, 2012.
10. Zhao, M., et al., Membrane lipids, EGF receptors, and intracellular signals colocalize and are polarized in epithelial cells moving directionally in a physiological electric field. The FASEB Journal, 2002.
11. Han, J., et al., Integrin beta1 subunit signaling is involved in the directed migration of human retinal pigment epithelial cells following electric field stimulation. Ophthalmic Res, 2011. 45(1): p. 15-22.
12. Jonathan D. Humphries, A.B., Martin J. Humphries, Integrin ligands at a glance. Journal of Cell Science, 2006. 119: p. 3901-3903.
13. Palazzo, A.F., et al., Localized Stabilization of Microtubules by Integrin- and FAK-Facilitated Rho Signaling. Science, 2004. 303(5659): p. 836-839.
14. Ann M. Rajnicek, L.E.F., Colin D. McCaig, Temporally and spatially
35
coordinated roles for Rho, Rac, Cdc42 and their effectors in growth cone guidance by a physiological electric field. Journal of Cell Science, 2006. 119:
p. 1723-1735.
15. Burdisso, J.E., Á. González, and C.O. Arregui, PTP1B promotes focal complex maturation, lamellar persistence and directional migration. Journal of Cell Science, 2013. 126(8): p. 1820-1831.
16. Guilluy, C., R. Garcia-Mata, and K. Burridge, Rho protein crosstalk: another social network? Trends in Cell Biology, 2011. 21(12): p. 718-726.
17. Pike, L.J., Rafts defined: a report on the Keystone symposium on lipid rafts and cell function. The Journal of Lipid Research, 2006. 47(7): p. 1597-1598.
18. Hernandez-Deviez, D.J., et al., Caveolin regulates endocytosis of the muscle repair protein, dysferlin. Journal of Biological Chemistry, 2008. 283(10): p.
6476-6488.
19. Wickström, S.A. and R. Fässler, Regulation of membrane traffic by integrin signaling. Trends in Cell Biology, 2011. 21(5): p. 266-273.
20. Sotobori, T., et al., Bone morphogenetic protein-2 promotes the haptotactic migration of murine osteoblastic and osteosarcoma cells by enhancing incorporation of integrin beta1 into lipid rafts. Exp Cell Res, 2006. 312(19): p.
3927-38.
21. Kusumi, A. and K. Suzuki, Toward understanding the dynamics of membrane-raft-based molecular interactions. Biochimica et Biophysica Acta (BBA) - Molecular Cell Research, 2005. 1746(3): p. 234-251.
22. Leitinger, B. and N. Hogg, The involvement of lipid rafts in the regulation of integrin function. Journal of Cell Science, 2002. 115(5): p. 963-972.
23. Cécile Boscher, I.R.N., Caveolins and Caveolae: Roles in Signaling and Disease Mechanisms, in CAVEOLIN-1: Role in Cell Signaling, Jean-François Jasmin, Philippe G. Frank, and Michael P. Lisanti, Editors. 2012.
24. Yang, B., et al., p190 RhoGTPase-Activating Protein Links the 1 Integrin/Caveolin-1 Mechanosignaling Complex to RhoA and Actin Remodeling. Arteriosclerosis, Thrombosis, and Vascular Biology, 2010. 31(2):
p. 376-383.
25. Rey-Barroso, J., et al., The dioxin receptor controls β1 integrin activation in fibroblasts through a Cbp–Csk–Src pathway. Cellular Signalling, 2013. 25(4):
p. 848-859.
26. Beardsley, A., et al., Loss of caveolin-1 polarity impedes endothelial cell polarization and directional movement. Journal of Biological Chemistry, 2005.
280(5): p. 3541-3547.
27. Arpaia, E., et al., The interaction between caveolin-1 and Rho-GTPases
36
promotes metastasis by controlling the expression of alpha5-integrin and the activation of Src, Ras and Erk. Oncogene, 2012. 31(7): p. 884-896.
28. Feng, C.-h., Y.-c. Cheng, and P.-h.G. Chao, The influence and interactions of substrate thickness, organization and dimensionality on cell morphology and migration. Acta Biomaterialia, 2013. 9(3): p. 5502-5510.
29. Danthi, P. and M. Chow, Cholesterol Removal by Methyl- -Cyclodextrin Inhibits Poliovirus Entry. Journal of Virology, 2003. 78(1): p. 33-41.
30. Carmena, M.J., et al., Cholesterol modulation of membrane fluidity and VIP receptor/effector system in rat prostatic epithelial cells. Regul Pept, 1991.
33(3): p. 287-97.
31. Hart, F.X., et al., Keratinocyte galvanotaxis in combined DC and AC electric fields supports an electromechanical transduction sensing mechanism.
Bioelectromagnetics, 2013. 34(2): p. 85-94.
32. Berdeaux, R.L., et al., Active Rho is localized to podosomes induced by oncogenic Src and is required for their assembly and function. J Cell Biol, 2004. 166(3): p. 317-23.
33. Tandon, N., et al., Electrical stimulation systems for cardiac tissue engineering. Nat. Protocols, 2009. 4(2): p. 155-173.