行政院國家科學委員會專題研究計畫 期中進度報告
探討主宰阿滋海默症之毒性機制--阿滋海默症蛋白質與細
胞膜間交互作用之研究(1/2)
計畫類別: 個別型計畫 計畫編號: NSC93-2214-E-002-033- 執行期間: 93 年 08 月 01 日至 94 年 07 月 31 日 執行單位: 國立臺灣大學化學工程學系暨研究所 計畫主持人: 王勝仕 報告類型: 精簡報告 報告附件: 出席國際會議研究心得報告及發表論文 處理方式: 本計畫可公開查詢中 華 民 國 94 年 5 月 23 日
探討主宰阿滋海默症之毒性機制--阿滋海默症蛋白質與細胞膜間交互作用之研究 (1/2) 計畫編號 : NSC 93-2214-E-002-033 王勝仕 國立台灣大學化學工程學系 摘要 本研究利用電位敏感性染劑配合螢光光度計,量測細胞膜偶極電位受不同實 驗條件變化之影響;並進一步藉由觀察細胞膜偶極電位之改變,對蛋白質與生物 細胞膜間之交互作用機制加以探討。嘗試尋求分子階層生物物理模式,以對Aβ 與 細胞膜間交互作用之機制作深入之探討與了解。 計畫緣由與目的 大約有二十種與人類有關之蛋白質因進行不正常地摺疊(misfold)而生成一種 稱為amyloid fibril 之物質並導致身體上之病變。這些蛋白質相互間在序列(sequence) 及功能方面並無相似之處;此外,在臨床、病理、及生物化學方面,這些疾病並 不直接相關且各具有其獨特之特性。然而,重要的是,這群疾病不僅具有相同的
發生機制,且造成這群疾病之各 amyloid 蛋白質,卻都能自我組合(self-assemble)
或 自 我 聚 集(self-aggregate) 成 具 有 獨 特 β- 摺 疊 板 (β-pleated sheet) 等 二 級 結 構 (secondary structure)之穩定 fibrils。導致阿滋海默症(Alzheimer’s Disease)之蛋白質 Aβ,即為這些蛋白質中族群中相當重要之一種 (Kelly 1998; Lansbury 1999)。
大 腦 皮 層(cerebral cortex) 中 不 可 溶 解 的 老 人 斑 (senile plaque) 之 逐 漸 凝 聚 (progressive deposition)被視為是阿滋海默症的一個重要病理上之標記(pathological hallmark);一般相信不可溶解之老人斑或 senile plaque 是由 amyloid fibrils 聚集而 成,而此些amyloid fibrils 即為一種稱為 β-amyloid 胜肽(又稱爲 Aβ)之蛋白質成份 所構成。Aβ 是一分子量約為 4.3 kD 之小蛋白質單體分子(monomer),在經過一連 串複雜反應後,能自我聚集成約為數個微米(micron)大之 fibril 分子,而幾個 fibril 分子則結合形成稱為senile plaque 之硬塊。一般發現,環繞在此些 senile plaque 硬 塊周圍之腦神經細胞(neuron)皆會經歷神經退化(neurodegeneration)過程而死亡 (Selkoe 1994)。雖然由遺傳學(Genetics)之研究(Kimberly et al. 2000)與體外毒性(in vitro toxicity)實驗之結果(Seilheimer et al. 1997; Hartley et al. 1999; Ward et al. 2000) 得知,Aβ 在阿滋海默症病理學(pathogenesis)上具有相當程度之重要性;然而由此 Aβ 誘發的神經毒性(neurotoxicity)與神經細胞死亡之分子階層機制則尚未完全清 楚。
關於 amyloid 蛋白質(包含 Aβ)與細胞膜間之交互作用,近來已被視為在 amyloidogenic 疾病發生機制上扮演一相當重要之角色。整體而言,過去文獻研究 之方向不外致力於交互作用與Aβ 之聚集作用的關係,以及交互作用在神經毒性所 扮演的角色。越來越多的文獻證實細胞膜的存在之於Αβ 聚集現象的重要性;一些 結果發現Aβ 能夠與磷脂質類細胞膜作用且此作用為與聚集狀態有關。相關的結果 重 點 分 別 為 :(1) 細 胞 膜 組 成 成 份 物 質 如 膽 固 醇 (cholesterol) 與 神 經 節 糖 苷 (ganglioside)能改變 Aβ對細胞膜之親和性(affinity)(Choo-Smith et al. 1997; Avdulov et al. 1999; Matsuzaki and Horikiri 1999)。 (2)胜肽與細胞膜間之結合(association)取 決於可獲得疏水性表面之含量多寡(Kremer et al. 2000)。(3)胜肽一旦與細胞膜結
合,負電性磷脂質類、膽固醇、與神經節糖苷被發現能提升 β-sheet 之含量及 Aβ
聚集反應速率(Choo-Smith et al. 1997; McLaurin et al. 1998; Avdulov et al. 1999; Del Mar Martinez-Senac et al. 1999; Matsuzaki and Horikiri 1999)。(4)不論體內或體外上, 研 究 發 現 可 溶 解 膽 固 醇 含 量 和/ 或 其 生 化 合 成 之 改 變 能 影 響 APP 之 正 常 processing:抑制膽固醇之合成會降低 Aβ之生成(Howland et al. 1998; Simons et al. 1998; Frears et al. 1999)。
雖然許多研究團隊提出了一些解釋Aβ 毒性作用之可能的分子機制,但目前仍
無統一之說法。幾個在Aβ 與細胞作用後與毒性有關聯的細胞反應(cellular response)
為鈣離子恆定性之改變(destabilization of calcium homeostasis)、細胞膜之去極化 (depolarization)、因自由基(free radical)之產生而造成細胞膜通透性(permeability)之 上升、細胞膜流動性之改變、鉀離子通道(potassium channel)之阻擋或功能喪失、 細胞膜整體性之直接破壞等。
一些研究結果發現,藉由改變細胞膜之靜電電位(electrostatic potential)
(Hertel et al. 1997);或降低細胞膜中膽固醇及神經節糖苷之含量(Wang et al. 2001); 或經由Congo red 或 rifampicin 與 Aβ 作用以降低 Aβ 與細胞膜間之疏水性交互作用 (hydrophobic interaction)(Burgevin et al. 1994; Lorenzo and Yankner 1994; Tomiyama et al. 1994; Sadler et al. 1995),皆可減輕神經毒性。另外一些研究者則觀察到 Aβ與磷 脂質細胞膜間之交互作用是與靜電作用力有關(Terzi et al. 1994; 1995; 1997)。在 circular dichrosim、titration calorimetry、及 ultracentrifugation 等測量方法輔助下,
研究人員發現磷脂質細胞膜之存在對於 Aβ 構形(conformation)在隨機團與 β-摺疊
片間之轉換(random coil-to-β-sheet transition)有關,且磷脂質帶電性之改變之於 Aβ 之構形有相當程度的影響。當在帶負電荷之磷脂質細胞膜加入系統後,Aβ 之構形 則幾乎全數以β-摺疊片呈現。(Terzi et al. 1994; 1995; 1997)。此外,研究者亦測試 Aβ 磷脂質細胞膜之親合作用以及 Aβ 誘發之神經細胞毒殺性受改變細胞膜性質添 加物質(如 phloretin、exifone)之影響(Hertel et al. 1997),結果發現 Aβ 與細胞膜間之
交互作用是受非專一性交互作用(non-specific interaction)所主宰。諸如此類對真正 Aβ 與細胞膜間交互作力模式之假說,例如:Aβ 與細胞膜間之交互作用否受細胞膜 流 動 性(membrane fluidity) 操 控 ? 此 作 用 力 是 否 為 偶 極 - 偶 極 間 交 互 作 用 力 (dipole-dipole interaction) ? 此 作 用 力 是 否 為 純 靜 電 作 用 力 (electrostatic interaction)?皆尚未有定論,仍待更多的技術與實驗數據加以證明及了解之。
偶極電位(dipole potential)是一種與生物膜(如:細胞膜)有關的局部化電位 (localized electrical potential)。此存在於生物膜間之偶極電位是源自於(1)生物膜中 脂質分子(lipid molecule)偶極極性殘基(dipolar residues)之對齊排列(alignment)以及 (2)介於水─脂質界面區域具方向性水分子之分子偶極(molecular dipole)(Brockman 1994; Clarke 2001)。近年來,許多與偶極電位測量相關之研究中,多採用一些例如 RH421 或 di-8-ANEPPS 等具電位高敏感性之 styrylpyridinium 染劑(voltage-sensitive styrylpyridinium dye);此類染劑之 chromophore 部位可與脂質細胞膜之脂質頭部位 (lipid headgroup)結合(Clarke 2001)。此類電位高敏感性之 styrylpyridinium 染劑乃是 應用其 pKa 之改變或螢光激發光譜(fluorescence excitation sprectrum)之偏移(shift) 會隨局部電位(local electric field)變化之原理間接地來量測生物膜中偶極電位之變 化(Clarke 2001)。
雖此偶極電位對疏水性離子(hydrophobic ion)穿越細胞膜之程序(translocation) 之影響性已廣為知(Ando et al. 1984);然而,此偶極電位在大部分牽涉巨分子物質 (macromolecule)與脂質雙層(lipid bilayer)之生化程序(biological processes)所扮演之 角色在最近才開始為人所注意。過去的研究結果顯示,偶極電位大小之變化會影 響胜肽或蛋白質與模式細胞膜(model membrane)間之交互作用,此類之作用包括: (1)細胞膜與訊息胜肽(signal peptide)間交互作用以及訊息胜肽構形(conformation)改 變(Cladera and O'Shea 1998; Voglino et al. 1998);(2)免疫缺陷病毒融合生胜肽 (immunodeficiency virus fusion peptide)其融合誘導能力(fusion induction capability) 及其與細胞膜之交互作用(Cladera et al. 1999);(3)短桿菌素(gramicidin)與細胞膜之 交互作用(Rokitskaya et al. 1997; Shapovalov et al. 1999);(4)分子偶極對人類表皮穿 透性之影響(Cladera et al. 2003);以及(5)磷脂酶 A(phospholipase A)與細胞膜之交互 作用等(Maggio 1999)。由上述可知,藉由觀察偶極電位之改變及變化,可提供吾 人對胜肽或蛋白質與生物細胞膜間之交互作用機制之進一步了解。 結果與討論 由之前的文獻回顧中,我們可以得知細胞膜表面、或是細胞膜之組成對於類 澱粉 β 蛋白質的毒性機制具有相當大的影響;而其他的研究提出了細胞膜的偶極 性會影響到細胞膜上的兩性蛋白質。所以在此提出一個可能影響類澱粉 β 蛋白質
與細胞膜間交互作用的因素-細胞膜的偶極電位;並且在研究中進一步探討細胞 膜組成成分對細胞膜偶極電位之影響。 為了研究細胞膜上的電位在類澱粉 β 蛋白質與細胞膜間交互作用上所扮演的 角色,於本研究中採用一種廣為使用的脂醇(sterol),phloretin,來降低細胞膜的偶 極電位;而6kc(6-ketocholestanol)則被用來增加細胞膜的偶極電位。 欲探測細胞膜偶極電位的變化,本研究中採用了雙波長螢光比值檢測法 (dual-wavelength ratiometric fluorescence method),利用加入各種化學物質引起細胞 膜電位變化,再運用電位敏感性染劑來標定細胞膜在不同物種影響下之電位變 化。此電位變化可藉由在固定 emission wavelength 下激發光譜(excitation spectra) 中,螢光讀值在460nm 與 520nm 的比值 R 表示。由實驗觀察得知,於固定 emission wavelength 下,在可降低細胞膜偶極電位化學物種(例如:phloretin)之激發光譜 (excitation spectra)中,會發現紅移現象(red shift);而可增強細胞膜偶極電位物種(像 是:6kc)之激發光譜中,則會呈現藍移現象(blue shift)。而藉由此兩種產生不同效
應的物種為基準,使得定義出的 R 值具有相對應的減小及增大結果;如此可免去
許多確切探測偶極電位的複雜方法,僅藉由觀察 R 值變化即可推知某物種對細胞
膜偶極電位的影響。
上述兩種最具代表性的影響偶極電位物種,並不僅僅改變偶極電位;也會改 變細胞膜的表面電位(surface potential)、穿膜電位(transmembrane potential)…等;但
由於所定義的R 值對此兩種電位並無相對之對應性,而偶極電位對於 R 值之對應
性則非常顯著;故在此實驗中僅選定此雙波長螢光比值檢測法來測定細胞膜偶極 電位的變化。
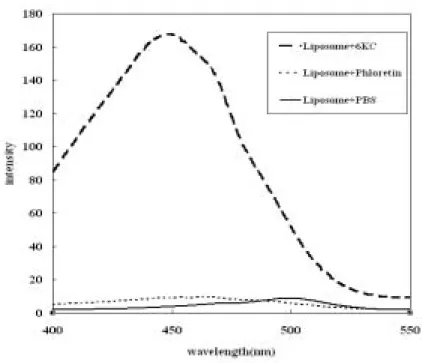
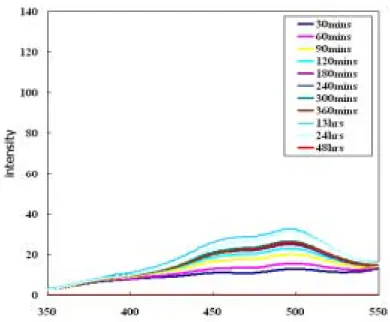
首先,先使用微脂粒(liposome)進行初步的實驗,以確定 phloretin 與 6kc 能夠 確實的作用在本實驗的系統中。在初步的實驗中,以DPPC(dipalmitoyl phosphatidyl choline)配置 SUV 大小的微脂粒,加入適量的 phloretin 與 6kc,以 2:1 的比例混合, 先置入恆溫培養箱中靜置 30 分鐘,再加入 di-8-ANEPPS(1-(3-sulfonatopropyl) -4-[[2-(di-n-octyl-amino)-6-naphthyl]vinyl]-pyridinium betaine)之電位敏感性染劑,最 後靜置於恆溫培養箱中2 小時,再以螢光光度計測得在 emission wavelength 580nm 下之激發光譜(圖 1);所得之 R 值如圖 2。
圖1 微脂粒與 6kc 及 phloretin 之激發光譜;微脂粒為 SUV 大小,濃度為 1.035mM、 6kc 及 phloretin 濃度為 3mM;PBS 為 Control 組,即 Liposome 加入 PBS。(6kc 及 phloretin 為溶解在甲醇中)
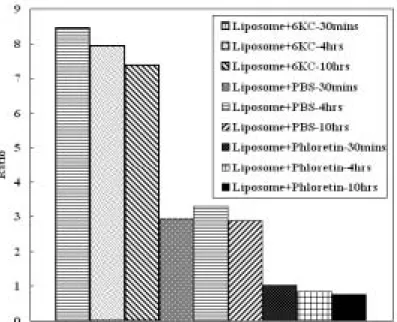
圖2 微脂粒與 6kc 及 phloretin 之 Ratio 圖;微脂粒為 SUV 大小,濃度為 1.035mM、 6kc 及 phloretin 濃度為 3mM ;PBS 為 Control 組,即 Liposome 加入 PBS。(6kc 及 phloretin 為溶解在甲醇中) 由初步的實驗發現,相對於對照組(不加入其他物質,僅加入原微脂粒所用之 0 2 4 6 8 10 1 Ratio 6KC PBS Phloretin
溶劑PBS)時,6kc 與 phloretin 皆造成 R 值顯著的增加與下降;所以此兩種物種確 定可使用於本實驗系統。而在實驗過程中,發現隨著不同的靜置時間,其 R 值也 會產生相對應的變化,並且影響明顯;為了找出實驗的最適靜置時間,故重新設 計實驗,希望可以找到靜置時間對R 值的影響。 針對靜置時間的影響之實驗步驟如下:配置足量的微脂粒,並將6kc、phloretin 與 PBS 分別和微脂粒混合均勻後,靜置於恆溫培養箱,並控制此段靜置時間分別 為30 分鐘、4 小時、及 10 小時;接著,將染劑(di-8-ANEPPS)加入,混合均勻後, 同樣將其靜置於恆溫培養箱中經過兩小時的靜置時間,再以螢光光度計測得分別 之激發光譜(如圖 3、4、及 5),並計算出其 R 值 (如圖 6) 由實驗結果顯示,第一段靜置時間雖然對於 6kc 及 phloretin 的激發光譜有明 顯的影響,但從R 值之觀點來看,phloretin 導致之 R 值變化幅度不大;而即使 6kc 的R 值,僅具一定程度的變化;但由於其 R 值本來就偏高,所以相對的變化率也 就不甚明顯。此外,PBS 不管激發光譜或是 R 值變化都不大,也符合了對照組不 會有所影響的觀點。 由實驗結果發現第一段靜置時間對 R 值的變化並不大,所以在之後的實驗可 以選取一長一短的第一段靜置時間,以重複確定此部分實驗結果。 圖3 微脂粒與 6kc 在不同時間混合靜置後,再加入染劑靜置第二段時間,之後使用
螢光光度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 0 20 40 60 80 100 120 140 160 180 350 400 450 500 550 wavelength(nm) inten sity
大小,濃度為1.035mM、6kc 濃度為 3mM 。(6kc 為溶解在甲醇中)
圖 4 微脂粒與 phloretin 在不同時間混合靜置後,再加入染劑靜置第二段時間,之
後使用螢光光度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂 粒為SUV 大小,濃度為 1.035mM、phloretin 濃度為 3mM 。(phloretin 為溶解在甲 醇中)
用螢光光度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小,濃度為 1.035mM,6kc 及 phloretin 濃度為 3mM ;PBS 為 Control 組, 即Liposome 加入 PBS。
圖6 微脂粒與 6kc、phloretin 及 PBS 在不同時間混合靜置後,再加入染劑靜置第二
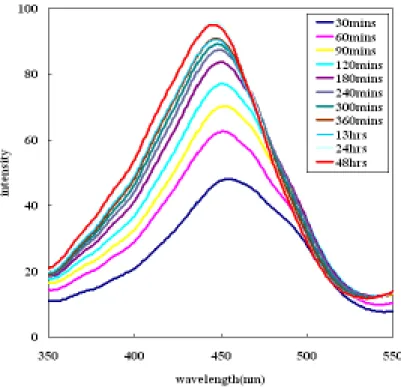
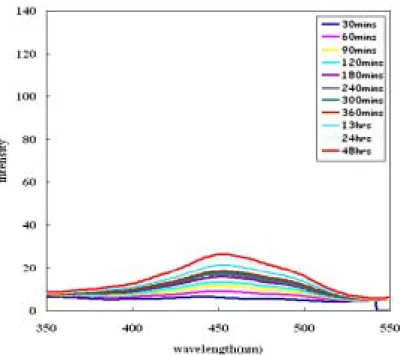
段時間,之後使用螢光光度計在emission wavelength 為 580nm 情況下所得之 Ratio 圖;微脂粒為SUV 大小,濃度為 1.035mM,6kc 及 phloretin 濃度為 3mM。(6kc 及 phloretin 為溶解在甲醇中) 關於靜置時間對R 值的影響,實驗步驟如下:配置足量的微脂粒,並將 6kc、 phloretin 與 PBS 和微脂粒混合均勻後,靜置於恆溫培養箱,並固定此段靜置時間 為30 分鐘(後文中以 t1代替);緊接著,將染劑(di-8-ANEPPS)加入,並將其再靜置 於恆溫培養箱中經過第二段靜置時間(後文中以 t2代替),以螢光光度計測得隨著不 同t2所得的激發光譜(如圖 7、8、9);並計算出 R 值隨著 t2的變動(如圖 10)。 由實驗結果可以發現,t2對 R 值的影響是相當明顯的:在長時間的情況下, 各種不同特性的物種對 R 值的影響都趨向一個平衡穩定值;像是 6kc 此類會使 R 值增大的物種就會隨著 t2的增加而使 R 值趨於一個最大值;反之,像是 phloretin 這類會減小 R 值的物種則是趨向一個最小值。以此觀之,之後的實驗設計在 t1的 選定並不重要,重要的是 t2最好選擇較長的時間點,因為除了時間長會得到越大 的R 值差異外,在短時間的測定時可由圖 4-2-8 發現其變動性很大,所以在測試未 知物種時,選擇較長的時間也會更加容易分辨出其特性。
圖7 微脂粒與 6kc 在混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2使用螢光光 度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小, 濃度為1.035mM,6kc 濃度為 3mM。(6kc 為溶解在甲醇中)
圖8 微脂粒與 phloretin 在混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2使用螢
光光度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小,濃度為1.035mM,phloretin 濃度為 3mM。(phloretin 為溶解在甲醇中)
圖9 微脂粒與 PBS 在混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2使用螢光光 度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小, 濃度為1.035mM。
圖10 微脂粒與 6kc、phloretin 及 PBS 分別混合靜置 30 分鐘後,再加入染劑靜置, 並隨著t2使用螢光光度計在emission wavelength 為 580nm 情況下所得之 Ratio 圖;
微脂粒為SUV 大小,濃度為 1.035mM,6kc 及 phloretin 濃度為 3mM。(6kc 及 phloretin 為溶解在甲醇中) 除了t2的影響外,實驗中亦探討6kc 及 phloretin 初始溶劑對 R 值之影響(如圖 11、圖 12 及圖 13)。由於此 R 值的改變不見得是因為溶劑本身的影響,有可能是 溶解度不同所造成的結果,故之後的實驗設計仍然採取有較佳溶解度的溶劑。 圖11 微脂粒與 6kc 在混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2使用螢光光
度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小, 濃度為1.035mM,6kc 濃度為 3mM。(6kc 為溶解在 PBS 中)
圖12 微脂粒與 6kc 在混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2使用螢光光 度計在emission wavelength 為 580nm 情況下所得之激發光譜;微脂粒為 SUV 大小, 濃度為1.035mM,6kc 濃度為 3mM。(phloretin 為溶解在 PBS 中)
圖13 微脂粒與 6kc、phloretin 及 PBS 分別混合靜置 30 分鐘後,再加入染劑靜置, 並隨著t2使用螢光光度計在emission wavelength 為 580nm 情況下所得之 Ratio 圖; 微脂粒為SUV 大小,濃度為 1.035mM,6kc 及 phloretin 濃度為 3mM。(6kc 及 phloretin
由上述的實驗決定實驗之基本條件如溶解溶劑、t1及t2;為了能夠觀察到較為 明顯的 R 值影響,所以選擇了固定 t1而仍隨著 t2以螢光光度計測量隨 t2變化之 R 值;其中,仍以甲醇作為6kc 及 phloretin 的溶劑。 除了上述這些控制條件外,在本研究中還另外探討了微脂粒本身濃度對 R 值 的影響;因為合成之微脂粒本身為球狀,而用來測量膜偶極電位變化的染劑是以 嵌入的方式與微脂粒作用,更進一步影響其偶極電位;所以假設:微脂粒濃度提 高,相對應的染劑用量應該也提升,如此所測得之 R 值才會實際地反映出該物種 所造成的變化。 由上述之假設,進一步分別以不同濃度(0.5mM、1mM 及 2mM)的微脂粒,重 複前述的實驗步驟,所得的結果如圖 14;由圖中可發現在相同的 t1及 t2下,高濃 度的微脂粒會有較佳的R 值變化,所以之後對於未知物種的性質測定,將採取 2mM 的微脂粒為主。 圖 14 微脂粒在不同濃度下(0.5mM、1mM 及 2mM) 分別與 6kc、phloretin 及 PBS 混合靜置 30 分鐘後,再加入染劑靜置,並隨著 t2 使用螢光光度計在 emission
wavelength 為 580nm 情況下所得之 Ratio 圖;微脂粒為 SUV 大小,6kc 及 phloretin 濃度為3mM。(6kc 及 phloretin 溶解在甲醇中)。
經過上述的實驗可以決定實驗的基本條件:微脂粒濃度、溶劑、t1及 t2; 而
值影響,所以選擇了2 小時及 13 小時為實驗的 t1;而t2為13 小時;仍以甲醇作為 6kc 及 phloretin 之溶劑;微脂粒濃度則選擇 2mM 為實驗用濃度。
在決定好所有控制條件後,一開始先測試類澱粉β 蛋白質(Aβ 25-35)在本系統
中是會對膜偶極產生增加或減少的影響。所以一方面經由加入新鮮備製及溶解於 不同溶劑下(TFA 或 DMSO)的類澱粉 β 蛋白質(Aβ 25-35)代替原本 6kc 及 phloretin 所扮演的角色;另一方面仍然同時間測量6kc、phloretin、PBS、PBS(D)及 PBS(T)(即 PBS 中含有少部分的 TFA 或 DMSO,其比例等同於欲溶解類澱粉 β 蛋白質所用之 劑量)分別與微脂粒混合後的 R 值作為對照(如圖 15)。 由實驗結果顯示,類澱粉 β 蛋白質(Aβ 25-35) 溶解在不同的溶劑對似乎影響 並不大;而且類澱粉β 蛋白質(Aβ 25-35)本身也沒有對 R 值產生變大或變小的趨勢 (與添加 PBS 之對照組相比)。所以可以由實驗結果推論,新鮮備製(fresh)的類澱粉 β 蛋白質(Aβ 25-35)不論溶解於何種溶劑,本身並沒有改變細胞膜偶極電位的能 力。除此之外,t1對R 值的影響在 t2的拉長下,也產生了些微的影響;不過與整體 之R 值比較下,此改變並不重要,與之前實驗所得的結果吻合。 圖15 微脂粒分別與 6kc、phloretin、PBS、PBS(T)、PBS(D)、Aβ25-35(T)、Aβ25-35(D) 混合靜置2 小時或 13 小時後,再加入染劑靜置於恆溫培養箱中 13 小時,之後使用
螢光光度計在emissionwavelength 為 580nm 情況下所得之 Ratio 圖;微脂粒為 SUV 大小,濃度為2mM,6kc 及 phloretin 濃度為 3mM,Aβ 25-35 濃度為 50µM,標示
(T)及(D)表示於配置過程中分別先溶於 TFA 及 DMSO 中(同第三章驗步驟所述), 之後符號亦同,將不再解釋。(6kc 及 phloretin 溶解在甲醇中)。 雖然新鮮備製的類澱粉β蛋白質 Aβ(25-35)本身並沒有影響細胞膜偶極電位的 能力,但是隨著類澱粉β蛋白質 Aβ(25-35)的培養時間拉長,其構造會逐漸轉變, 而此結構的轉變是否會改變細胞膜的偶極電位呢?為了要試驗此假設,於是又再 試驗了類澱粉 β 蛋白質 Aβ(25-35),並將其分為兩組,分別為新鮮備製(fresh)與備 製後旋轉培養(rotation) 7 天(aged),希望藉著旋轉培養時間的拉長而看出其對細胞 膜偶極電位的影響,而其結果如圖16。 由圖中可以看出即使拉長旋轉培養時間,類澱粉β 蛋白質 Aβ(25-35)對 R 值似 乎仍沒有顯著的改變(與對照組相比);而這是否意味著類澱粉 β 蛋白質 Aβ(25-35) 對 R 值就完全沒有影響呢?還是旋轉培養的時間仍然不夠長,而使得類澱粉 β 蛋 白質Aβ(25-35)本身的構造變化不夠完整?為了解決這個問題,於是繼續設計了之 後的實驗。 圖16 微脂粒分別與 6kc、phloretin、PBS、PBS(T)、PBS(D)、Aβ(25-35)(T)、Aβ(25-35)(D) 混合靜置2 小時或 13 小時後,再加入染劑靜置於恆溫培養箱中 13 小時,之後使用
螢光光度計在emission wavelength 為 580nm 情況下所得之 Ratio 圖;微脂粒為 SUV 大小,濃度為2mM,6kc 及 phloretin 濃度為 3mM,Aβ 25-35 濃度為 50µM 並於製 備後旋轉七天。(6kc 及 phloretin 溶解在甲醇中)。
由過去本研究室的結果顯示,細胞膜表面或是細胞環境的組成對於類澱粉 β 蛋白質與細胞膜的交互作用也有相當大的影響,其中又以膽固醇及神經節甘脂的 存在會增加類澱粉β 蛋白質中 β-sheet 的成分及聚集速率;再配合上述實驗之結論, 所以進一步探討微脂粒中膽固醇的存在,是否會影響類澱粉 β 蛋白質 Aβ (25-35) 與微脂粒之R 值變化。 實驗步驟如前面所述,選擇 2 小時及 13 小時為實驗的 t1;而t2為13 小時;以 甲醇作為6kc 及 phloretin 的溶劑;而微脂粒濃度則選擇 1mM 及 2mM 為實驗用濃 度;此外,在配置微脂粒的同時加入了 30%的膽固醇(Cholesterol);另外類澱粉 β
蛋白質Aβ(25-35)皆備製了 fresh 及 aged 兩種不同的實驗組。因膽固醇本身就會增
強細胞膜偶極電位,也就是使R 值增大;可由觀察加入膽固醇後,各物種的 R 值 皆會明顯增加證實。為了減去膽固醇本身對 R 值所造成的干擾,在此定義出一個 比值(公式如下):為對照組所測得之 R 值減去加入某物種所測得之 R 值,再除以對 照組之R 值,最後乘以 100%;而藉由這樣的定義,可以比較出,實際上各物種實 際對R 值的影響,更進一步推測該物種是增強細胞膜偶極電位,或是相反 (如表 1 所述)。 R(%):代表新定義出的 R 值。 RatioControl:代表對照組所測出的R 值。 RatioChemical:代表各物種所測得的R 值。 表1 加入膽固醇後,各物種利用公式所算出的 R(%) (C)2hr_R_% (C)12hr_R_% A-beta(25-35)(T)_0.053 g/l 9.095435013 12.67875874 A-beta(25-35)(T)_0.053 g/l_A 18.1270354 24.49888866 PBS(T)_70 -1.71912725 3.05075805 計畫成果自評 整體而言,此研究計畫之目標在於鑑別與了解amyloid 胜肽與細胞膜間之交互
作用。此amyloid 胜肽與細胞膜間之交互作用已知與胜肽聚集(peptide aggregation) 和細胞毒殺性(neurotoxicity)具密不可分之關係。研究中利用電位高敏感性之 Control Chemical Control
Ratio
- Ratio
R(%) =
100%
Ratio
×
styrylpyridinium 染劑配合螢光光譜激發光譜量測技術檢視及鑑定 amyloid 胜肽之聚 集狀態(aggregation state)和胜肽胺基酸順序(amino acid sequence)對胜肽與數種磷脂 質類模型細胞膜(model membrane) 間交互作用之影響,並進行系統化之探討。本年 度完成(1)電位高敏感性之 styrylpyridinium 染劑配合螢光量測系統最適化操作條件 之探討;(2)以電位高敏感性之 styrylpyridinium 染劑配合螢光量測 Aβ(25-35) (fresh or aged)與磷脂質模型細胞膜間交互作用後之細胞膜偶極電位之變化;(3)探討細胞
膜組成如膽固醇對amyloid-磷脂質模型細胞膜偶極電位之影響等。擬將現有成果延
伸至下年度,除進行Aβ(1-40)及其他 amyloid 蛋白質之探討,與膽固醇對 amyloid-磷脂質模型細胞膜偶極電位之影響進一步深入研究外,亦嘗試著手進行神經細胞 培養實驗,尋求細胞培養神經毒性資料與分子階層生物物理模式間之關聯性,以
對Aβ−細胞膜間交互作用之機制與 Aβ 誘發之神經毒性作進一步之探討與了解。
參考資料
Ando, H.Y., Schultz, T.W., Schnaare, R.L., and Sugita, E.T. 1984.
Percutaneous-Absorption - a New Physicochemical Predictive Model for
Maximum Human Invivo Penetration Rates. Journal of Pharmaceutical Sciences 73: 461-467.
Avdulov, N.A., Chochina, S.V., Igbavboa, U., Warden, C.S., Schroeder, F., and Wood, W.G. 1999. Lipid binding to sterol carrier protein-2 is inhibited by ethanol. Biochim. Biophys. Acta. 1437: 37-45.
Brockman, H. 1994. Dipole Potential of Lipid-Membranes. Chemistry and Physics of Lipids 73: 57-79.
Burgevin, M.C., Passat, M., Daniel, N., Capet, M., and Doble, A. 1994. Congo red protects against toxicity of beta-amyloid peptides on rat hippocampal neurones. Neuroreport 5: 2429-2432.
Choo-Smith, L.P., Garzon-Rodriguez, W., Glabe, C.G., and Surewicz, W.K. 1997. Acceleration of amyloid fibril formation by specific binding of Abeta- (1-40) peptide to ganglioside-containing membrane vesicles. J. Biol. Chem. 272: 22987-22990.
Cladera, J., Martin, I., Ruysschaert, J.M., and O'Shea, P. 1999. Characterization of the sequence of interactions of the fusion domain of the simian immunodeficiency virus with membranes - Role of the membrane dipole potential. Journal of Biological Chemistry 274: 29951-29959.
Cladera, J., and O'Shea, P. 1998. Intramembrane molecular dipoles affect the membrane insertion and folding of a model amphiphilic peptide. Biophys J 74: 2434-2442. Cladera, J., O'Shea, P., Hadgraft, J., and Valenta, C. 2003. Influence of molecular dipoles
on human skin permeability: Use of 6-ketocholestanol to enhance the transdermal delivery of bacitracin. Journal of Pharmaceutical Sciences 92: 1018-1027. Clarke, R.J. 2001. The dipole potential of phospholipid membranes and methods for its
detection. Advances in Colloid and Interface Science 89: 263-281.
Del Mar Martinez-Senac, M., Villalain, J., and Gomez-Fernandez, J.C. 1999. Structure of the Alzheimer beta-amyloid peptide (25-35) and its interaction with negatively charged phospholipid vesicles. Eur. J. Biochem. 265: 744-753.
Frears, E.R., Stephens, D.J., Walters, C.E., Davies, H., and Austen, B.M. 1999. The role of cholesterol in the biosynthesis of beta-amyloid. Neuroreport 10: 1699-1705. Hartley, D.M., Walsh, D.M., Ye, C.P., Diehl, T., Vasquez, S., Vassilev, P.M., Teplow, D.B.,
and Selkoe, D.J. 1999. Protofibrillar intermediates of amyloid beta-protein induce acute electrophysiological changes and progressive neurotoxicity in cortical neurons. J. Neurosci. 19: 8876-8884.
Hertel, C., Terzi, E., Hauser, N., Jakob-Rotne, R., Seelig, J., and Kemp, J.A. 1997. Inhibition of the electrostatic interaction between beta-amyloid peptide and membranes prevents beta-amyloid-induced toxicity. Proc Natl Acad Sci U S A 94: 9412-9416.
Howland, D.S., Trusko, S.P., Savage, M.J., Reaume, A.G., Lang, D.M., Hirsch, J.D., Maeda, N., Siman, R., Greenberg, B.D., Scott, R.W., et al. 1998. Modulation of secreted beta-amyloid precursor protein and amyloid beta- peptide in brain by cholesterol. J. Biol. Chem. 273: 16576-16582.
Kelly, J.W. 1998. The alternative conformations of amyloidogenic proteins and their multi- step assembly pathways. Curr Opin Struct Biol 8: 101-106.
Kimberly, W.T., Xia, W., Rahmati, T., Wolfe, M.S., and Selkoe, D.J. 2000. The transmembrane aspartates in presenilin 1 and 2 are obligatory for
gamma-secretase activity and amyloid beta-protein generation. J. Biol. Chem. 275: 3173-3178.
Kremer, J.J., Pallitto, M.M., Sklansky, D.J., and Murphy, R.M. 2000. Correlation of beta-amyloid aggregate size and hydrophobicity with decreased bilayer fluidity of model membranes. Biochemistry 39: 10309-10318.
Lansbury, P.T., Jr. 1999. Evolution of amyloid: what normal protein folding may tell us about fibrillogenesis and disease. Proc Natl Acad Sci U S A 96: 3342-3344. Lorenzo, A., and Yankner, B.A. 1994. Beta-amyloid neurotoxicity requires fibril
formation and is inhibited by congo red. Proc Natl Acad Sci U S A 91: 12243-12247.
Maggio, B. 1999. Modulation of phospholipase A(2) by electrostatic fields and dipole potential of glycosphingolipids in monolayers. Journal of Lipid Research 40: 930-939.
Matsuzaki, K., and Horikiri, C. 1999. Interactions of amyloid beta-peptide (1-40) with ganglioside-containing membranes. Biochemistry 38: 4137-4142.
McLaurin, J., Franklin, T., Fraser, P.E., and Chakrabartty, A. 1998. Structural transitions associated with the interaction of Alzheimer beta-amyloid peptides with
gangliosides. J. Biol. Chem. 273: 4506-4515.
Rokitskaya, T.I., Antonenko, Y.N., and Kotova, E.A. 1997. Effect of the dipole potential of a bilayer lipid membrane on gramicidin channel dissociation kinetics. Biophys J 73: 850-854.
Sadler, II, Smith, D.W., Shearman, M.S., Ragan, C.I., Tailor, V.J., and Pollack, S.J. 1995. Sulphated compounds attenuate beta-amyloid toxicity by inhibiting its association with cells. Neuroreport 7: 49-53.
Seilheimer, B., Bohrmann, B., Bondolfi, L., Muller, F., Stuber, D., and Dobeli, H. 1997. The toxicity of the Alzheimer's beta-amyloid peptide correlates with a distinct fiber morphology. J. Struct. Biol. 119: 59-71.
Selkoe, D.J. 1994. Alzheimer's disease: a central role for amyloid. J Neuropathol Exp Neurol 53: 438-447.
Shapovalov, V.L., Kotova, E.A., Rokitskaya, T.I., and Antonenko, Y.N. 1999. Effect of gramicidin A on the dipole potential of phospholipid membranes. Biophysical Journal 77: 299-305.
Simons, M., Keller, P., De Strooper, B., Beyreuther, K., Dotti, C.G., and Simons, K. 1998. Cholesterol depletion inhibits the generation of beta-amyloid in hippocampal neurons. Proc. Natl. Acad. Sci. U. A. 95: 6460-6464.
Terzi, E., Holzemann, G., and Seelig, J. 1994. Alzheimer beta-amyloid peptide 25-35: electrostatic interactions with phospholipid membranes. Biochemistry 33: 7434-7441.
Terzi, E., Holzemann, G., and Seelig, J. 1995. Self-association of beta-amyloid peptide (1-40) in solution and binding to lipid membranes. J Mol Biol 252: 633-642. Terzi, E., Holzemann, G., and Seelig, J. 1997. Interaction of Alzheimer beta-amyloid
peptide(1-40) with lipid membranes. Biochemistry 36: 14845-14852.
Tomiyama, T., Asano, S., Suwa, Y., Morita, T., Kataoka, K., Mori, H., and Endo, N. 1994. Rifampicin prevents the aggregation and neurotoxicity of amyloid beta protein in vitro. Biochem Biophys Res Commun 204: 76-83.
Voglino, L., McIntosh, T.J., and Simon, S.A. 1998. Modulation of the binding of signal peptides to lipid bilayers by dipoles near the hydrocarbon-water interface. Biochemistry 37: 12241-12252.
Wang, S.S., Rymer, D.L., and Good, T.A. 2001. Reduction in cholesterol and sialic Acid content protects cells from the toxic effects of Beta -amyloid peptides. J Biol Chem 276: 42027-42034.
Ward, R.V., Jennings, K.H., Jepras, R., Neville, W., Owen, D.E., Hawkins, J., Christie, G., Davis, J.B., George, A., Karran, E.H., et al. 2000. Fractionation and
characterization of oligomeric, protofibrillar and fibrillar forms of beta-amyloid peptide. Biochem. J. 348 Pt 1: 137-144.
出席國際學術會議心得報告及發表之論文
1.心得報告
計畫主持人王勝仕參加在日本舉行之10-th Symposium of Young Asian Biochemical Engineering Community (2004)及 Asia Europe Biorecognition Symposium 會議。茲將參加會議之議程分列如下:
Conference/Meeting City, Country Date
YABEC (2004) Osaka, Japan 2004, Sept. 23-25 Asia Europe Biorecognition
Symposium
Shimonoseki, Japan 2004, Sept. 26-27 Society of Chemical
Engineers, Japan (SCEJ) meeting
Shimonoseki, Japan 2004, Sept. 27-28
於10-th Symposium of Young Asian Biochemical Engineering Community (2004) 中發表一oral presentation:”Examining the Mechanisms of Alzheimer’s Disease” 及一 poster:”Aggregation Kinetics of β-Amyloid Peptides in Different Solvent
Environments”。於 Third Asia Europe Biorecognition Symposium 中發表一 oral presentation:”Examining the Mechanisms of Alzheimer’s Disease”。相關之邀請函、 大會議程、與摘要見附件。
在此二國際研討會議中,除獲取相關新知與研究進展外,並與各國研究人員 交換心得,收穫頗豐盛。
Examining the Mechanisms of Alzheimer’s Disease
Steven Sheng-Shih WangDepartment of Chemical Engineering National Taiwan University, Taipei, Taiwan 106
ABSTRACT
More than sixteen different human proteins can fold abnormally resulting in the formation of an amyloid fibril and some accompanying pathology. The proteins have little sequence homology; however, they are able to misfold or self-assemble into stable fibrils with a characteristic cross pleated β-sheet secondary structure. Among these proteins, β-amyloid (Aβ) of Alzheimer’s disease is the best characterized. Aβ is the primary protein component of senile plaques associated with Alzheimer’s disease and has been implicated in the neurotoxicity associated with the disease.
The hypothesis that the aggregated Aβ leads to the neurotoxicity associated with AD has been widely accepted. Then one is left with several compelling questions: What are the intermolecular interactions that lead to Aβ aggregates and how might one control them? On a more macroscopic level, one could pose a similar question: What would be the structure of the aggregated toxic species and how might one alter the aggregation pathway to avoid the formation of toxic species? Finally, by what mechanism does aggregated Aβ interact with cells to cause neurotoxicity? In order to answer these questions, we set up experiments to (a) investigate the peptide structure, at the residue level, associated with self-assembly process of Aβ, (b) develop new tools to examine the toxicity of Aβ oligomeric species and characterize the size of toxic oligomers, (c) identify roles of membrane composition (cholesterol and gangliosides) and G protein activation in the mechanisms of the toxic action of Aβ on neurons, and (d) use different amyloid protein(s) to test the feasibility of this structure-function relationship. Achievement of these goals could be used to aid in the further understanding of mechanism of Aβ aggregation, the role of aggregate size in the toxicity, and the mechanism of Aβ toxicity.
The ability to investigate how Aβ aggregates, to characterize the correlation between size of aggregated Aβ species and toxicity, to demonstrate the role of cell membrane composition in Aβ’s toxic effect, and to understand how Aβ interact with cell membranes may lead to better understanding of Αβ-induced neurotoxicity and aid in developing potential treatments for Alzheimer’s disease.
Aggregation Kinetics of
β-Amyloid Peptides in Different Solvent
Environments
Steven S.-S. Wang1 and Josephine W.-J. Wu2*
1
Department of Chemical Engineering
National Taiwan University, Taipei, Taiwan 106
2
Department of Optometry
Chung Shan Medical University, Taichung, Taiwan 402
At least sixteen different human proteins can fold abnormally resulting in the
formation of an amyloid fibril and some accompanying pathology. These proteins
have little sequence homology; however, they are able to misfold or self-assemble
into stable fibrils with a characteristic cross pleated β-sheet secondary structure. β-amyloid of Alzheimer’s disease is the best characterized. β-amyloid (Aβ) is the key protein component of senile plaques associated with Alzheimer’s disease and has
been implicated in the neurotoxicity associated with the disease.
Accumulating evidence indicates that the most crucial factor influencing
Aβ-induced neurotoxicity is the aggregation state of Aβ species. Our previous results also showed the correlation between the type of initial solvent and the
aggregation state of Aβ species. In the present work, we examined the kinetic behavior of β-amyloid peptide during the course of aggregation. Synthetic Aβ(1-40) and its partial fragments Αβ(25-35) were utilized in this study. We believe that this work contributes to our better understanding, from the kinetic perspective, of how Aβ species and its derivatives aggregate.
4-1, Okadayama,Nisinomiya, Hyogo 662-8505, JAPAN
Invitation to YABEC 2004
08. 05. 2004
Dr. Sheng-Shih Wang
Dear Dr. Sheng-Shih Wang
Young Asian Biochemical Engineers’ Community 2004 (YABEC’04) will be held on Sept. 23 – 25, 2004 at the Cosmosquare International Education and Training Center in Osaka. On behalf of the organizing committee of YABEC’04, it is my pleasure to send this invitation for you to attend the forthcoming conference. We are looking forward to discussing your oral presentation entitled “Examining the
Mechanisms of Alzheimer’s Disease”.
This will be the 10th YABEC meeting since the first meeting held in Seoul in 1995. It could be a good opportunity to look back our steps for these 10 years, and think our future collaboration. We would like to hold the conference under a catchphrase “Advancing together for another 10 years!”
We will have oral presentations, poster presentations, and scientific trip. Detailed information can be obtained from our home-page, which will be up-dated frequently, at URL http://web-mff.kobe-c.ac.jp/YABEC04/YB2004.html.
We are looking forward to meeting you at the conference. With best wishes、
Masaaki Terashima, Dr. Eng. Professor
Department of Human Environment Sciences School of Human Sciences
Kobe College
Welcome to YABEC 2004
YABEC 2004 in Osaka
Date: September, 23-25 (2004)
Place: Cosmosquare International Education And Training Center
Homepage:
Home page in English
Information on YABEC 2004
New info!
Schedule of Meeting
(Up-loaded on Aug. 25)
List of all presentations (Keynote Speech, Plenary lectures, Oral
presentations, and Poster presentations) are available.
How to prepare poster
(Up-loaded on Aug. 25)
Techinical Tour
(Up-loaded on Aug. 12)
Info of a Travel Agency -Pre and Post conference
accomodations-Important information to YABECian from China area
Message from Chair.
第 1 頁,共 2 頁 YABEC 2004
2004/9/2 http://web-mff.kobe-c.ac.jp/YABEC04/YB2004.html
OP-II- 3 (15:40 - 16:00)
Jianjiang Zhong (East China University of Science and Technology, Shanghai)
New Elicitors as a Driving Force to Achieve a Highly Productive Bioprocess for Useful Plant Secondary Metabolites
OP-II- 4 (16:00 - 16:20)
Tsong-Rong Yan (Tatung University, Taipei)
Cloning and regulator prediction of Aspergillus niger β-glucosidase II OP-II- 5 (16:20 - 16:40)
Congfa Li (South China University of Tropical Agriculture, Hainan )
Chiral 3-phenyllactic acid preparation by biocatalytic asymmetrical reduction of phenylpyruvic acid OP-II- 6 (16:40 - 17:00)
Jer-Yiing Houng (I-Shou University, Kaohsiung County)
Enhance the Efficiency of the Asymmetric Bioreduction of Achiral Ketone with Statistical Methods
Oral Presentation III
OP-III- 1 (15:00 - 15:20)Zhangfa Tong (Guangxi University, Nanning)
Extraction and Purification of Hirudin from a Blood-Sucking Leech, Hirudinaria Manillensis OP-III- 2 (15:20 - 15:40)
Gyun Min Lee (Korea Advanced Institute of Science and Technology (KAIST), Daejeon)
Enhanced Human Thrombopoietin Production by Sodium Butyrate Addition to Serum-Free Suspension Culture of Bcl-2 Overexpressing CHO Cells
OP-III- 3 (15:40 - 16:00)
Yu-Chen Hu (National Tsing Hua University, Hsinchu )
Baculovirus as a Highly Efficient Gene Delivery Vector for the formation of Hepatitis Delta Virus-like particles in Mammalian Cells
OP-III- 4 (16:00 - 16:20)
Steven Sheng-Shih Wang (National Taiwan University, Taipei) Examining the Mechanisms of Alzheimer s Disease
OP-III- 5 (16:20 - 16:40)
Hiroshi Ueda (The University of Tokyo, Tokyo)
Creation of sensor proteins for specific DNA sequences OP-III- 6 (16:40 - 17:00)
Hea-Yeon Lee (Osaka University, Osaka)
Recognition of nano-well array based on electric gene chip Go back to Conference schedule
第 2 頁,共 2 頁 List of poster presentation
2004/9/2 http://web-mff.kobe-c.ac.jp/YABEC04/oral_list.html
Examining the Mechanisms of Alzheimer’s Disease
Steven Sheng-Shih WangDepartment of Chemical Engineering National Taiwan University, Taipei, Taiwan 106
ABSTRACT
More than sixteen different human proteins can fold abnormally resulting in the formation of an amyloid fibril and some accompanying pathology. The proteins have little sequence homology; however, they are able to misfold or self-assemble into stable fibrils with a characteristic cross pleated β-sheet secondary structure. Among these proteins, β-amyloid (Aβ) of Alzheimer’s disease is the best characterized. Aβ is the primary protein component of senile plaques associated with Alzheimer’s disease and has been implicated in the neurotoxicity associated with the disease.
The hypothesis that the aggregated Aβ leads to the neurotoxicity associated with AD has been widely accepted. Then one is left with several compelling questions: What are the intermolecular interactions that lead to Aβ aggregates and how might one control them? On a more macroscopic level, one could pose a similar question: What would be the structure of the aggregated toxic species and how might one alter the aggregation pathway to avoid the formation of toxic species? Finally, by what mechanism does aggregated Aβ interact with cells to cause neurotoxicity? In order to answer these questions, we set up experiments to (a) investigate the peptide structure, at the residue level, associated with self-assembly process of Aβ, (b) develop new tools to examine the toxicity of Aβ oligomeric species and characterize the size of toxic oligomers, (c) identify roles of membrane composition (cholesterol and gangliosides) and G protein activation in the mechanisms of the toxic action of Aβ on neurons, and (d) use different amyloid protein(s) to test the feasibility of this structure-function relationship. Achievement of these goals could be used to aid in the further understanding of mechanism of Aβ aggregation, the role of aggregate size in the toxicity, and the mechanism of Aβ toxicity.
The ability to investigate how Aβ aggregates, to characterize the correlation between size of aggregated Aβ species and toxicity, to demonstrate the role of cell membrane composition in Aβ’s toxic effect, and to understand how Aβ interact with cell membranes may lead to better understanding of Αβ-induced neurotoxicity and aid in developing potential treatments for Alzheimer’s disease.
International seminar 国際会議
5th Ube bioseparation
seminar
September 25, Saturday,
2004, in Ube
Engineering School,
Yamaguchi University
第5回バイオ分離宇部 セミナー 9月25日(土)山口大学工 学部D11(宇部市)
3rd Asia-Europe Biorecognition Engineering Symposium in Shimonoseki
Sept. 26, Sun and 27, Mon, 2004
at Kaikyo Messe
第3回アジアーヨーロッパ生体認識工学シンポジウム(下関) 下関海峡メッセ8F801 会議室(下関市)26日 (日)27日(月
)
5th Ube bioseparation seminar, Sept.25, Sat,13:00-19:00, at Yamaguchi Univ.,
Ube
"Separation process for antibodies and other protein drugs"
Shuichi Yamamoto, Yamaguchi University, Japan
Introductory remarks: Bioprocess chromatography for antibody
and other protein drug purification.
Alois Jungbauer, IAM, Austria
Characteristics of Protein A chromatography media
Milos Barut, BIA separations, Slovenia
Monolithic support for chromatography of large biomolecules
Oliver Kaltenbrunner Amgen, USA
Application of modeling in the process characterization of
ion-exchange chromatography of proteins
Peter Watler, Vaxgen, USA
Continuous chromatography operation for cost and
capacity optimization of proteins
Testuro Ogawa, Pentax, Japan
Hydroxyapatite for antibody purification
Masayoshi Tsukahara and Takashi Ishihara, Kirin,
Japan
Antibody production and purification processes for
pharmaceuticals
Kazuhisa Uchida, Kyowa Hakko, Japan
A next generation antibody technology: De-fucosylation on improved effector
function of antibodies and the impact for purification.
Yasutami Mitoma, Tosoh, Japan
Development of new chromatography media (Toyopearl) for
antibody purification
Get-acquainted party
-*-*-*-3rd Asia-Europe Biorecognition Engineering Symposium (AEBES)
Sept.26 at
Kaikyo Messe Shimonoseki
in
Shimonoseki 13:30-18:00
第 1 頁,共 3 頁 ube-aebes2004
2004/9/2 http://kako.chem.yamaguchi-u.ac.jp/ube-aebes2004.htm