國立臺灣大學生命科學院生命科學系 博士論文
Department of Life Science College of Life Science National Taiwan University
Doctoral Dissertation
鈣網蛋白與血管內皮生長因子於神經母細胞瘤分化調控 之研究
Study of Calreticulin and VEGF-A on the Regulation of Neuronal Differentiation in Neuroblastoma
翁妏謹 Wen-Chin Weng
指導教授:李心予 博士 Advisor: Hsinyu Lee, Ph. D .
中華民國 104 年 1 月
January 2015
誌謝
在鍵盤上敲下誌謝的這一刻,才真實感受到博士班的生涯即將告一段落,回 首過去,研究的生涯一度面臨家庭與工作必須同時兼顧的痛苦掙扎,也曾萌生放 棄的念頭,但我很慶幸這一路上有許多的貴人相助,讓我能夠堅持到這一刻,在 此由衷獻上我最誠摯的感謝。
首先最要感謝的是李心予老師對於我的諄諄教誨與悉心關懷。李老師不僅在 實驗上給予專業的指導與充分的支援,更在我人生遇到各種困境與挫折時不斷鼓 勵我,幫助我,常讓我在沮喪之餘,再度重燃信心。老師對於研究與教育的熱忱 與風範,都讓我永誌不忘,深刻影響著我日後的工作和生活。此外,還要感謝李 旺祚醫師,他在臨床工作與研究上對於我的指導與建議,讓我少走許多冤枉路,
讓我這些年來無論在工作或研究上都獲益匪淺。另外,要特別感謝許文明醫師,
他對於晚輩們總是慷慨且毫不保留地提供重要的意見與實質的協助,在辛苦工作 之餘仍不忘與我討論實驗的方向,是這篇論文能夠完成的重要推手。再者,很榮 幸可以邀請到廖永豐老師和黃敏銓老師擔任我的口試委員,給予我寶貴的建議與 指導,讓我的論文可以更臻完善。
李老師的研究室是一個溫暖且歡樂的大家庭,雖然我是研究室中年齡最大的 學姊,但在許多實驗的技術上常常需要學弟妹的幫忙。特別感謝貫浤、沛翊、柏 堅、Milky、易謙、予農,以及許多曾經一起打拼的學弟妹們,謝謝你們在實驗 上的協助,也很喜歡跟你們相處的日子,讓我常常覺得自己年輕了十歲,也讓我 以身為實驗室的一分子為榮。
最後,最要感謝的是在背後支持我的家人們。謝謝老公這幾年來的陪伴,在 我人生的低潮,你的鼓勵與支持一直是讓我繼續努力的最大動力,還有我一對最 可愛的寶貝,每天回家看到寶貝們的笑容,聽到寶貝們的聲音,所有的辛苦都煙 消雲散。有你們相伴,即使面對再大的難關,我也會鼓起勇氣繼續前進,謹將此 成果與你們共享,也將此榮耀獻給我辛苦的母親,與在天上來不及參與此刻的最 疼愛我的父親。謝謝 !!
中文摘要
神經母細胞瘤是幼兒期最常見的惡性腫瘤,其腫瘤形成的分子機制依然 不清楚,有可能是胚胎交感神經系統發育異常使神經母細胞無法分化或無法 凋亡而造成。我們過去的文獻回顧發現內質網伴護蛋白包括鈣網蛋白 (Calreticulin, CRT)與葡萄糖調節蛋白(GRP)為神經系統的胚胎發過育過程所
必需。我們的斑馬魚研究亦顯示 CRT 在神經系統的胚胎發育過程極具重要
性。過去的研究發現CRT 是神經母細胞瘤一項重要的預後指標,CRT 的大
量表現與神經母細胞瘤的分化程度呈正相關性,病人並有較佳的預後表現。
因此,CRT 也在神經母細胞瘤的分化上扮演了重要的角色。
血管內皮生長因子(VEGF-A)與其引導的血管新生現象亦被證實在神經
母細胞瘤的形成具有重要的角色。而在胃癌研究發現CRT 與 VEGF-A 具有
相關性。因此,本研究希望藉由細胞實驗與動物實驗探討 VEGF-A 是否參
與在CRT 對於神經母細胞瘤的分化調控,並進一步探討 VEGF-A 對於人類
神經母細胞瘤的分化調控與臨床意義。我們過去的研究結果發現在三種不同 的神經母細胞瘤細胞株實驗,CRT 的大量表現皆會正向調控 VEGF-A 與其上 游調控分子缺氧誘導因子 HIF-1的表現量,並增加 VEGF-A 的蛋白質分泌。
反之,利用 shRNA 抑制 CRT 的表現亦會造成 VEGF-A 及 HIF-1的表現下降。
而在本研究中,我們進一步發現 CRT 的大量表現不會影響細胞凋亡,但會促
進細胞分化並抑制細胞增生。此外,我們利用 VEGF 接受器抗體去抑制 VEGF-A 的作用,則神經母細胞瘤的神經分化指標包括 GAP43、NSE、NFH 及 TrkA 亦 會受到抑制。以上結果顯示 VEGF-A 的確在 CRT 誘導的神經母細胞分化扮演
重要角色。然而,我們利用神經母細胞瘤細胞株進行實驗發現 CRT 大量表
現會促進細胞之神經分化無法增生,因此經由四環黴素誘導 CRT 表現之
stNB-V1 神經母細胞瘤細胞株來進行動物實驗。我們接著利用腫瘤異體移植
實驗進一步證實CRT 對於 VEGF-A 及 HIF-1的正向調控,實驗亦發現誘導
CRT 的表現可以顯著抑制腫瘤體積並促進腫瘤分化。除此之外,我們發現
在病人腫瘤裡CRT 的表現和 VEGF-A 的表現呈現顯著正相關。更重要的是,
從病患的病理切片染色發現具有 VEGF-A 表現的病患其預後亦較佳。
VEGF-A 的表現與腫瘤的分化程度呈正相關,與 MYCN 的表現呈負相關,
但與內皮血管形成無相關性,顯示 VEGF-A 可能透過與內皮血管新生無關
之機轉來調控神經母細胞瘤的形成與分化。
總之,本研究證實在神經母細胞瘤中,CRT 可正向調控 VEGF-A 表現並
促進腫瘤的神經分化,且 VEGF-A 確實參與此神經分化調控。我們亦首次
證實VEGF-A 是神經母細胞瘤的一個獨立預後因子,具有 VEGF-A 表現的病
患其預後亦較佳。我們的研究為神經母細胞瘤的腫瘤形成開啟嶄新的機轉,
同時也有助於對神經母細胞瘤新治療的發展。
關鍵詞:鈣網蛋白、血管內皮生長因子、神經母細胞瘤、神經分化、血管新生
Abstract
Neuroblastoma (NB) is the most common malignant tumor of infancy. The tumorigenesis of NB could be a divergence of the embryonic development of
sympathetic nervous system. ER chaperones including calreticulin (CRT) and GRP78 are suggested to participate during embryonic development in our previous review.
Our present study in zebrafish also revealed that CRT is essential for embryonic and neuronal development. Previous study has identified CRT as an independent favorable prognostic factor which is related to differentiated histologies in NB. Taken together, CRT could play an important role in neuronal differentiation of NB. Recently
evidence has suggested that vascular endothelial growth factor (VEGF)-A, a key regulator of physiological and pathologic angiogenesis, participates in the behavior of NB. Furthermore, recent studies have found a correlation between CRT and VEGF-A in gastric cancers. In the present study, we aimed to determine whether the CRT expression in NB was associated with the VEGF-A pathway and to determine the role of VEGF-A in regulating NB behavior focusing on angiogenesis and neuronal
differentiation in vitro and in vivo. Our previous study clearly demonstrated that in different NB cell lines, CRT over-expression increases the expression and secretion of VEGF-A and HIF-1, a major positive regulator of VEGF-A. In contrast, knockdown
of CRT decreases VEGF-A and HIF-1 expression. In the present study, we further
demonstrated that NB cell apoptosis was not affected by CRT over-expression in stNB-V1 cells. Nevertheless, over-expression of CRT suppressed cell proliferation and enhanced cell differentiation in stNB-V1 cells, whereas blockage of VEGFR-1 markedly suppressed the expression of neuron specific markers including GAP43, NSE and NFH as well as TrkA, a molecular marker indicative of NB cell
differentiation. These results indicate an essential role of VEGF-A in CRT-related neuronal differentiation in NB. However, constitutive over-expression of CRT led to NB cell differentiation without proliferation. Thus, we used an inducible-CRT stNB-V1 cell line by a tetracycline-regulated gene system for further animal
experiments. The mice xenograft models further confirmed the positive regulation of CRT on VEGF-A and HIF-1, as well as the role of CRT in enhancing neuronal
differentiation and suppressing tumor growth in NB. Furthermore, we have demonstrated a significantly positive correlation between CRT and VEGF-A expression in human NB tumors. Most important of all, we verified that VEGF-A expression predicts a favorable outcome in NB patients and are associated with differentiated histology and normal MYCN expression, both of which are favorable prognostic factors. On the other hand, there was no correlation between the expression of VEGF-A and CD34, a marker of endothelial cells, suggesting a novel mechanism
of VEGF-A participating in NB formation through angiogenesis-independent pathway.
In conclusion, our study indicated that CRT-dependent VEGF-A up-regulation is critical for NB differentiation and VEGF-A is involved in CRT-related neuronal differentiation in NB. For the first time, we have demonstrated that VEGF-A is an independent prognostic factors and predicts favorable outcomes in NB patients with tight relationship with differentiated histology and MYCN status. Our findings also delineate a novel mechanism of VEGF-A in the biology of NB. This study provides important information that is needed for deciphering the crucial role of CRT and VEGF on the regulation of NB differentiation. Furthermore, our findings will shed light to a novel therapeutic strategy to improve the outcome of NB patients in the future.
Keywords: Calreticulin, VEGF-A, neuroblastoma, neuronal differentiation,
angiogenesis
Table of contents
口試委員會審定書 ... i
誌謝 ... ii
中文摘要 ... iii
Abstract ... v
List of Tables ... x
List of Figures ... xi
Chapter I. Introduction ... 1
1.ER stress and ER-resident chaperone proteins ... 2
1.1 ER stress and UPR... 2
1.2 Multifunctional roles of ER-resident chaperone proteins ... 3
1.3 Calreticulin ... 3
1.3.1 Fundamental information of calreticulin ... 3
1.3.2 Regulation of calreticulin expression ... 4
2.Roles of calreticulin in neuroblastoma ... 5
2.1ER chaperones in cancer development ... 5
2.2Calreticulin in regulating cancer cell proliferation ... 7
2.3 The biology of Neuroblastoma ... 8
2.4 ER chaperones and calreticulin in neuroblastoma ... 10
3.Roles of calreticulin in embryonic and neuronal development ... 11
3.1ER chaperones in embryonic and neuronal development ... 11
3.2 Roles of calreticulin in neuronal development and neuronal differentiation ... 12
3.2Zebrafish as a model in studying embryonic development ... 13
4.VEGF-A and angiogenesis in neuroblastoma ... 13
4.1 Fundamental information of VEGF-A ... 13
4.2 Angiogenesis in neuroblastoma ... 14
4.3 VEGF-A in neuroblastoma ... 15
5.The relationship between calreticulin and VEGF-A-driven angiogenesis ... 16
6.Rationales ... 17
Chapter II. Materials and Methods ... 18
Chapter III. Results ... 31
Chapter IV. Discussions ... 44
Chapter V. Concluding remarks and future perspectives ... 53
References ... 57
Tables ... 67
Figures ... 68
Appendix I: Role of glucose-regulated Protein 78 in embryonic development and neurological disorders ... 98
Appendix II. Calreticulin Regulates VEGF-A in Neuroblastoma Cells ... 108
List of Tables
Table 1. VEGF-A expression and clinicopathologic and biologic
characteristics of neuroblastomas... 67
List of Figures
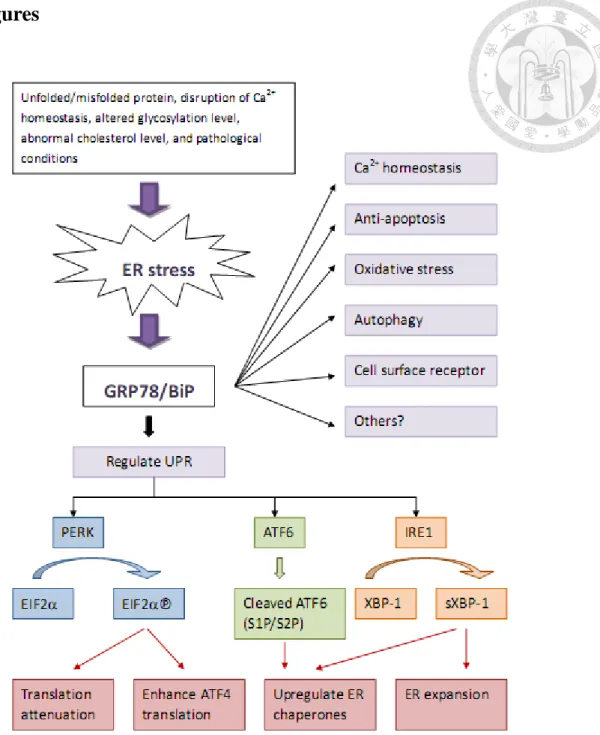
Figure 1. Summary of physiological functions of glucose-regulated protein 78. ... 68
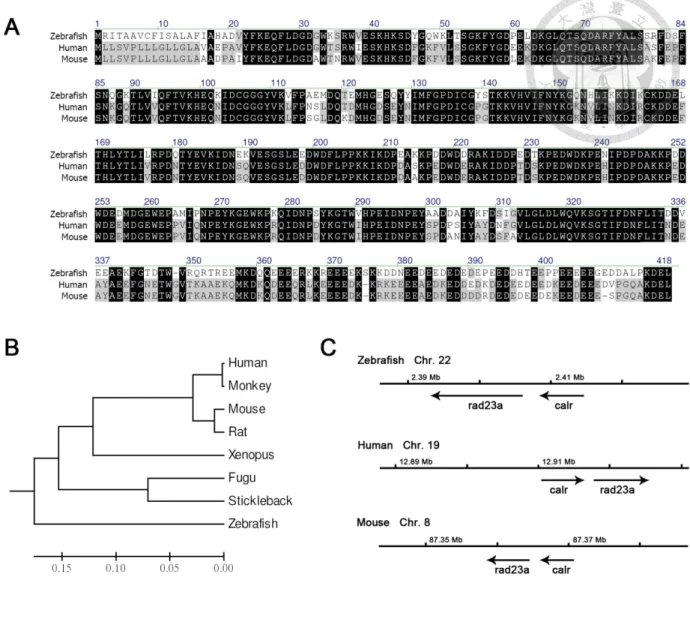
Figure 2. Sequence conservation analysis of zebrafish CRT. ... 69
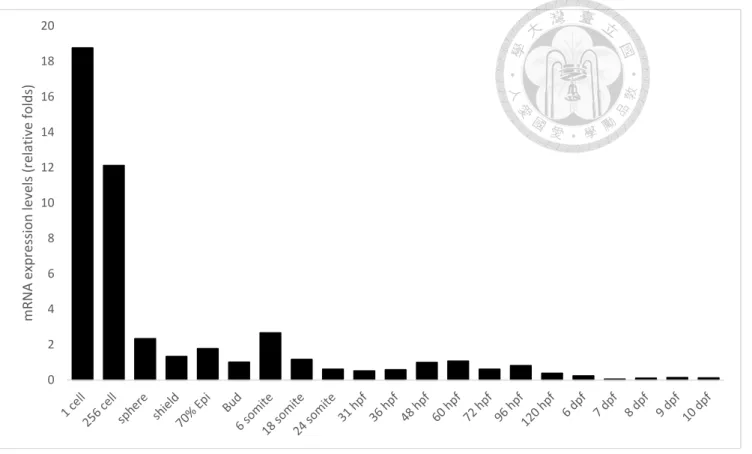
Figure 3. Temporal expression pattern of CRT in developing zebrafish embryos. ... 70
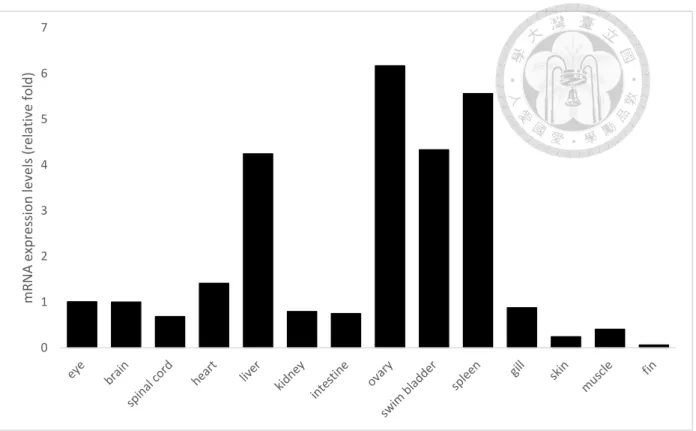
Figure 4. Spatial expression patterns of CRT in adult fish. ... 71
Figure 5. The zCRT-ATGMO morphants at 2dpf showed severe developmental retardation with concentration-dependent effects. ... 72
Figure 6. CRT over-expression up-regulated VEGF-A and HIF-1 in SK-N-DZ and SH-SY5Y NB cells. ... 73
Figure 7. Knockdown of CRT down-regulated VEGF-A and HIF-1 in SK-N-DZ and SH-SY5Y NB cells. ... 75
Figure 8. VEGF-A protein expression and secretion level were up-regulated in SK-N-DZ and SH-SY5Y NB cells with CRT over-expression, but decreased by the knockdown of CRT in SK-N-DZ and SH-SY5Y NB cells. ... 77
Figure 9. The VEGF-A and HIF-1 expressions were regulated by CRT expression induced by tetracycline treatment in stNB-V1 cells. ... 79
Figure 10. The VEGF-A and HIF-1 mRNA expressions were lower in CRT knockdown stable stNB-V1 cells. ... 81
Figure 11. Apoptosis was not affected by CRT expression induced by tetracycline
treatment in stNB-V1 cells. ... 83
Figure 12. Over-expression of CRT reduced the proliferation rate of stNB-V1 cells. 84
Figure 13. CRT-induced neuronal markers expression in NB cells is VEGF-A
dependent. ... 85
Figure 14. CRT expression suppressed NB tumor growth in mice xenograft models.
... 87 Figure 15. The VEGF-A, HIF-1 and GAP43 expressions were up-regulated by CRT
expression in mice xenograft models. ... 89
Figure 16. VEGF-A mRNA expressions were positively correlated with CRT
expression in primary NB tumors. ... 91
Figure 17. The expressions of CRT and VEGF-A are enhanced in primary NBs with
differentiated histology. ... 92
Figure 18. Immunohistochemical study of CRT, VEGF-A and CD34 expression in
NB tumors. ... 94
Figure 19. VEGF-A protein expression is correlated with favorable outcome in NB
patients. ... 96
Figure 20. Schematic illustration summarized the role of VEGF-A in CRT-related
neuronal differentiation in NB. ... 97
Chapter I.
Introduction
1. ER stress and ER-resident chaperone proteins
1.1 ER stress and UPR
Endoplasmic reticulum (ER) is the principle cellular organelle in which secretory and membrane proteins are properly folded and modified. ER also functions as a major intracellular calcium store and is responsible for biosynthesis of steroids and cholesterol. Moreover, ER is the site for N-linked glycosylation, which is also important for correct protein folding [1]. Proteins that cannot be correctly folded would be eradicated via proteasome-mediated ER-associated degradation (ERAD) pathway [2]. The accumulation of unfolded or misfolded proteins in the ER lumen has referred to as ER stress and in eukaryotes it would induce adaptive coordinated
responses, which are described as unfolded protein response (UPR) [3]. A number of cellular stress conditions, such as disruption of calcium homeostasis, altered
glycosylation level, secretory protein mutations, and abnormal cholesterol level, can activate the UPR and contribute to a wide range of human diseases. Three ER-resident transmembrane proteins function as stress sensors and are involved in the UPR: the inositol requiring kinase 1 (IRE1), activating transcription factor 6 (ATF6) and PKR-like endoplasmic reticular kinase (PERK) transducers, which transduce the unfolded proteins signal cross the ER membrane and lead to the activation of UPR [4]
(Fig. 1). If these adaptive coordinated responses cannot eliminate inappropriate
folding proteins during prolonged and severe ER stress, the UPR would elicit pro-apoptotic pathway and trigger apoptotic cell death [5].
1.2 Multifunctional roles of ER-resident chaperone proteins
To assist and regulate the correct folding of proteins, ER contains abundant proteins known as molecular chaperons including the 78-kDa glucose-regulated protein 78/immunoglobulin-binding protein (GRP78/BiP), GRP75 and calreticulin (CRT). These chaperone proteins contain ER stress elements (ERSE) in their
promoters and are upregulated during ER stress [6]. Other than as regulators of UPR, several ER chaperones are involved in many cellular processes For example, GRP78, a member of heat shock protein 70 (HSP70) family, also referred to as BiP or Hspa5, has been found to regulate important biological functions including calcium
homeostasis, anti-apoptosis, autophagy and as cell surface receptor [4] (Fig. 1).
1.3 Calreticulin
1.3.1 Fundamental information of calreticulin
Calreticulin (CRT) is another important chaperone protein primarily localized to the endoplasmic reticulum and highly conserved in diverse species [7]. It is composed of three structurally and functionally distinct domains, including N-, P-, and
C-domain [8]. The N-terminal domain is responsible for chaperon functions of CRT [9]. The proline-rich P-domain contains two sets of three repetitive regions [10].
These repeated amino acid sequences form the lectin-like chaperone structures which are responsible for protein-folding function of CRT. The C-domain of CRT is a highly acidic region which is important for Ca2+-buffering functions. It is known that Ca2+
binding to this region play a critical role in the interaction with other chaperone proteins in ER [8, 11].
Apart from highly enriched within the ER lumen, CRT has also been identified in cytosol [12] and on cell surface [13]. Cell-surface CRT is found to be associated with phagocytic uptake and immunogenicity of cells [14]. The evidence provides more insights for CRT as a multifunctional protein. The multi-functional roles of CRT in protein chaperoning, Ca2+ homeostasis, cell transduction, modulating cell adhesion, and gene expression disclose its major involvement in various physiologic and pathologic conditions [7, 15].
1.3.2 Regulation of calreticulin expression
The human calreticulin gene (CALR) is located on chromosome 19p13.2 with nine exons. Calreticulin promoter region contains several binding site for reputed transcriptions factors and many of these factors have been identified as important modulators of CRT expression including NKx2.5, MEF2C, COUP-TF1, GATA6,
Evi-1, and PPAR factors [16]. In addition, calcium depletion and ER stress were shown to be important activators of CALR transcription [17]. Recently, studies have also revealed that nerve growth factor (NGF) can also up-regulate CRT expression in both ovarian cells and neuronal differentiation [18, 19]. These results further
suggested an involvement of CRT expression in various biological and pathological processes.
2. Roles of calreticulin in neuroblastoma
2.1 ER chaperones in cancer development
More and more evidences demonstrated that ER-resident chaperone proteins play critical roles in many pathological illnesses and a variety of human diseases [20].
Whether the involvement of ER-resident chaperone proteins is the primary cause or a secondary consequence in diseases is yet to be determined. Most findings of ER chaperones in human diseases have been focused on carcinogenesis and tumor progression. For example, our recent reviews showed that GRP78 is involved in cancer progression and drug resistance [4].
Moreover, the correlation between CRT expression levels and tumorigenesis has been extensively studied in various cancers and most reports have revealed that tumor tissues express significant higher levels of CRT compared to normal tissues [21].
Up-regulation of CRT presented in a variety of cancer cell lines, solid tumors, and human cancer cells, including oral cancer [22], breast ductal carcinoma [23, 24], colorectal cancer [25], prostate cancer [26], and vaginal carcinoma [27]. Tumor invasion, metastasis and recurrence were also positively correlated to increased CRT expression while the patients’ survival rate were inversely correlated to increased CRT expression in gastric cancer [28], breast cancer [29], bladder cancer [30], pancreatic cancer and esophageal squamous cell carcinoma [31, 32]. These results indicate that increased CRT expression might play a crucial role in cancer progression.
Another important role for CRT exposed on the cell surface, which is relevant for destruction of cancer cells, is via induction of the immune response [14, 33]. Results from several laboratories have demonstrated that cell-surface CRT facilitates the phagocytic uptake of apoptotic and cancer cells [34-36]. Obeid et al. demonstrated that drug treatments (anthracyclins) caused tumor cell to expose a surface pro-phagocytic protein, CRT, which induced immunogenic cell death [37]. It is becoming clear that surface exposure of CRT is required for phagocytosis on dying tumor cells. CRT expressed on the cell surface is considered as an “eat-me” signal and promote phagocytic uotake of cancer cells by immune system for multiple human cancers.
On the other hand, the roles of CRT in ovarian cancer progression are
inconclusive. Compared with primary tumors and solid metastases, reduced CRT expression was observed in malignant effusions of high-grade ovarian carcinoma along disease progression [38]. Besides, CRT expression levels may be associated with better response to chemotherapy while the survival was not related to CRT expression [38]. Very recently, mutations in calreticulin gene (CALR) were detected in a majority of myeloproliferative neoplasms [39-41]. Furthermore, in neuroblastoma, increased CRT expression is found to be associated with better prognosis and differentiated histologies [42, 43]. Therefore, the impact of CRT on tumor formation and progression may depend on different cell types and clinical stages.
2.2 Calreticulin in regulating cancer cell proliferation
Cancer formation and progression is characterized by rapid proliferation of mutant cells. Increasing evidence have revealed that manipulation of CRT expression had obvious effects on cell proliferation in various cancers. In pancreatic cells, overexpressed CRT enhanced cell growth, while knockdown of CRT had the opposite effect on cell growth [31]. In addition, reduction of CRT caused cell cycle arrest at the G0/G1 phase which resulted in significantly suppressed growth rate, colony-formation capacity, and anchorage-independent growth in oral cancer cell [22]. In bladder cancer, knockdown of CRT is also found to suppress cell growth
[15]. Although most studies have suggested that CRT expression positively regulates cell growth, other studies provided conflicting evidence on this issue. A recent study demonstrated that prostate cancer cells with higher CRT levels produced fewer colonies as well as inhibition of tumor growth both in vitro and in vivo [44].
According to these reports, whether CRT promotes or suppresses cell proliferation may rely on different cell types.
2.3 The biology of Neuroblastoma
Neuroblastoma (NB) is a common childhood tumor and the most frequently diagnosed malignancy in infancy, with more than 96% of patients diagnosed at the age of <10 years old [45, 46]. It is derived from the sympatho-adrenal lineage of embryonic neural crest cells [47]. Children with NB have a broad spectrum of clinical diversity that is highly associated with age at onset and genetic, biological, and pathological characteristics [47]. As a result, 50% of the NB patients die from this disease that continues to be one of the most challenging tasks among pediatric tumors.
The mechanism underlying its tumorigenesis remains obscure, despite the identification of several clinically relevant prognostic markers. Previous studies suggest that incomplete development and failure of differentiation or apoptosis of neuroblastic cells is critical in its development [48]. Previous studies have shown that
NB cells exhibit a capacity of differentiating into mature cells or spontaneous
regression by apoptosis [49, 50]. Studies also demonstrated that NB can be forced to differentiate upon the treatment of retinoic acid [51]. On the other hand, NB with better prognosis often express molecular markers indicative of cell differentiation, such as TrkA [52]. Furthermore, the expressions of apoptosis-related genes including p53, Bcl-2, and Bax have been demonstrated in NB and are correlated with favorable prognosis [53]. However, the factors contributing to the regulation of NB cell
differentiation or apoptosis are still unclear.
Genetic studies have postulated MYCN as one of the most important molecules that is closely related to the pathogenesis including differentiation, proliferation and apoptosis of NB. MYCN is a proto-oncogene normally expressed in the developing central and peripheral nervous systems [54]. Its expression is regulated by several signaling pathways, such as E2F1 [55]. As a bHLH transcription factor, MYCN dimerizes with MAX and binds to the E-box sequence (CACGTG) of the promotor region of target genes [56]. The major gene family that is directly regulated by MYCN is called the N-MYC Downstream Regulate Gene (NDRG) [57]. Through the transcriptional regulation of these genes, MYCN regulates the proliferation, growth, differentiation and survival of cells in the developing nervous system. In NB tumors, amplification of MYCN is closely associated with metastasis, advanced disease stages
and poor outcome [58].
2.4 ER chaperones and calreticulin in neuroblastoma
Though extensive exploration of ER chaperones in tumorigenesis, the roles of ER chaperones in neuroblastoma have not carefully addressed. Recently, Hsu et al.
have demonstrated that several chaperones including CRT, GRP75 and GRP78 are independent favorable prognostic markers in NB [42, 59, 60]. Hsu et al. found that the percentage of positive GRP78 immunostaining increased as the tumor histology of NB became differentiated (p=0.001) [60]. GTP78 expression as a significant factor for predicting favorable outcome has also been found in other studies of olfactory NB [61]. In addition, Hsu et al. have also found that positive GRP75 immunostaining is strongly correlated with differentiated histologies (p<0.001) and as an independent favorable prognostic factor [59].
Moreover, evidences also suggest that CRT may play an essential role in the biology of NB. Previous studies reveal that CRT is on the surface of NB cells and is essential for neurite formation when NB cells are induced to differentiate [62, 63]. Another in vitro study using NB cell line reveals that increased CRT expression is correlated with the differentiation of NB cells [64]. Recently, CRT has been identified by Hsu et al. as an independent favorable prognostic marker in
NB [42]. They have found that increased CRT expression in NB positively correlates with tumor differentiation and therefore predicts favorable outcome. In conclusion, CRT may play an essential role in the differentiation of NB cells, as well as in the differentiation of other neural progenitor cells or neural stem cells.
However, how CRT affects the differentiation of NB warrants further clarification.
3. Roles of calreticulin in embryonic and neuronal development
3.1 ER chaperones in embryonic and neuronal development
A number of molecules normally expressed during embryonic development, including HNK-1, neuropeptide Y, tyrosine hydroxylase, TrkA and CD44, are found in NB [48, 65], suggesting that the tumorigenesis of NB could be a divergence of the embryonic development of the sympathetic system. During embryogenesis a number of coordinated events occur to ensure orderly generation of the diverse cell types leading to proper organ system development including nervous system development.
One of these events is apoptosis that has been shown to widely occur during central nervous system development [66]. Apoptosis may be induced by a number of different stimuli including ER stress [67]. Recently, a study found evidence that ER stress involved in the development of the central nervous system in the mouse models
[68]. Several ER chaperones, including CRT and GRP78 were expressed at higher levels in embryonic brain and retina than in adult tissues. Apart from the roles in human diseases, converging evidences suggested that ER-resident chaperone proteins also participate during embryonic development and physiological conditions [4, 20].
3.2 Roles of calreticulin in neuronal development and neuronal differentiation
Neuroblastoma is derived from the sympathoadrenal lineage of embryonic neural crest cells and results from incomplete development and differentiation of precursor cells [45]. A recent study using CRT knockout (KO) mouse model exhibited lethality with significant defects in heart, brain and body wall, suggesting that CRT is essential in the embryonic development of nervous system [69]. Moreover, a recent study using PC-12 cells expressing mutant CRT lacking a Ca2+-buffering domain (C domain), suggested that the Ca2+-regulating capacity of CRT is essential for NGF-elicited neuronal differentiation [19]. In NB, previous studies have demonstrated that the expression levels of CRT positively correlated with the differentiation of NB both in
vitro and in vivo [42, 43]. In addition, it has been reported that surface CRT is crucial for neurite formation when NB cells are induced to differentiate [62]. Accordingly, these findings suggest that CRT may play an important role in nervous system
differentiation) of NB.
3.2 Zebrafish as a model in studying embryonic development
The zebrafish is vertebrae with transparent embryos and identifiable, stereotyped neurons. As a consequence of the widespread use of zebrafish in developmental biology studies, an extensive array of experimental tools and techniques has also been assembled in the analysis of human neurodegenerative and neurodevelopmental diseases.
4. VEGF-A and angiogenesis in neuroblastoma
4.1 Fundamental information of VEGF-A
Vascular endothelial growth factor (VEGF)-A (also referred to as VEGF), which belongs to a family of secreted homodimeric disulfide-bound glycoproteins, is a key regulator of physiologic and pathologic angiogenesis and highly conserved between animals [70, 71]. In mammals, this family includes VEGF-A (VEGF), VEGF-B, VEGF-C, VEGF-D, and placental growth factor (PlGF). The human VEGF-A gene contains eight exons and alternative splicing of VEGF-A gene gives rise to at least six different transcripts [72].
The biological functions of VEGF-A in regulating endothelial cell proliferation, migration, vascular permeability, secretion and other endothelial functions were
mediated by two related receptor tyrosine kinase (RTK), VEGFR-1(Flt-1) and VEGFR-2 (KDR/Flk-1) [73]. Both VEGFR-1 and VEGFR-2, which differ in signaling properties, contain seven immunoglobulin-like extracellular domains, a single transmembrane region and a consensus tyrosine kinase sequence. The VEGF-A also binds to two non-tyrosine kinase receptors of the neuropilin (NRP) family, NRP1 and NRP2, which are thought to be modulators for the VEGF-VEGFRs signaling.
VEGF-A expression is mainly regulated by oxygen tension [74]. The VEGF-A gene contains hypoxia-responsive enhancer elements (HREs) and the mRNA expression of the VEGF-A gene induced by hypoxia is mediated by binding of the transcription factor hypoxia-inducible transcription factor 1 (HIF-1) to the HRE.
HIF-1 is a heterodimer composed of HIF-1 and HIF-1 subunits. Under hypoxic situations, the HIF-1 is up-regulated because the proline hydroxylation is inhibited
and then enhanced the transcription of VEGF-A [71]. In addition, several growth factors and inflammatory cytokines, including TGF-, TGF-, platelet-derived
growth factor, fibroblast growth factor, insulin-like growth factor-1, interleukin-1 and interleukin-6 can also upregulate VEGF-A expression [70].
4.2 Angiogenesis in neuroblastoma
NB tumors are highly vascular and a large variety of pro-angiogenic factors,
such as VEGF, FGF-2,IL-8, TGF-, PDGF-A, angiopoietins and erythropoietin have
been found to play crucial roles in the induction of angiogensis in advanced-stage NB tumors [75, 76]. Although it is still debatable, the high vascular index, as expressed by numbers of vessels per square millimeter of tissue area, has been shown to correlate with the adverse prognosis of NB patients, suggesting a close relationship between aggressive tumor behavior and active angiogenesis [77, 78]. However, another study reveals that tumor vascularity is not correlated with prognosis and tumor stage in NB patients [79]. Moreover, it has been shown that there is no tumor growth difference in NB xenograft mice treated with angiogenesis inhibitor [80]. In a recent study,
investigators have explored the essential role of angiogenesis in the maturation phase of NB [81]. Therefore, the role of angiogenesis in NB tumorigenesis and outcome is complicated and needs further investigation.
4.3 VEGF-A in neuroblastoma
Apart from its role in embryonic vascular development, more and more studies reveal that VEGF-A plays an important role in the progression and metastasis of cancer cells [82]. In NB, angiogenesis is an essential mechanism regulating NB tumorigenesis, while VEGF-A-driven angiogenesis plays a critical role in the pathogenesis of NB formation and metastasis [83-85]. Both in human and in
experimental NB, VEGF-A over-expression has been demonstrated and correlated with a high-risk phenotype [86, 87]. In contrast, recent studies have found that VEGF-A is not related to tumor progression and metastasis in NB [88]. Moreover, it has been shown that there is no tumor growth difference in NB xenograft mice treated with or without Anti-VEGF antibody [89]. Thus, the roles of VEGF-A in the
tumorigenesis of NB remain obscure and need further clarification.
5. The relationship between calreticulin and VEGF-A-driven angiogenesis
Some studies have found a correlation between CRT and VEGF-A in human cancers. In gastric cancers, CRT has been found to up-regulate VEGF expression and enhance angiogenesis, leading to poor prognosis [28]. However, it has also been reported that exogenous CRT and the CRT fragment, vasostatin, are considered as anti-angiogenic factors and inhibit VEGF-induced endothelial cell proliferation [90, 91]. According to these reports, whether CRT promotes or suppresses
VEGF-A-driven angiogenesis remains elusive and needs further exploration.
6. Rationales
The tumorigenesis of NB could be a divergence of the embryonic development of sympathetic nervous system. ER chaperones including CRT and GRP78 are found to participate during embryonic development. Studies in CRT knockout mice which demonstrated the essential role of CRT for embryogenesis have suggested the important role of CRT in nervous system development as well as the biology of NB.
As we mentioned above, calreticulin (CRT) has been previously correlated with the differentiation of NB tumors, implying a favorable prognostic factor. However, how CRT affects the neuronal differentiation and of NB remains unclear.
VEGF-A and VEGF-A-driven angiogenesis have been reported to participate in the behavior of NB. However, the roles of VEGF-A in NB progression and prognosis are complicated and controversial. Furthermore, recent studies have found a
correlation between CRT and VEGF-A in gastric cancers. Nevertheless, the
relationship between CRT and VEGF-A in NB has never been studied. Thus, in this
study, we proposed to investigate the association of CRT and VEGF-A in
regulating NB tumorigenesis focusing on angiogenesis and neuronal
differentiation in vitro and in vivo.
Chapter II.
Materials and Methods
Ethics statement
All zebrafish and mouse experiments were performed after approval from the Institutional Animal Care and Use Committee at National Taiwan University. The clinical evaluation and use of tumor tissues for this study were approved by the Institutional Review Board of National Taiwan University Hospital. Written informed consent was obtained from the patients before sample were collected.
Fish breeding and embryo collection
Breeding fish will be maintained at 28.5 °C on a 14-h light/10-h dark cycle in a certified fish facility. Embryos will be collected by natural spawning, raised in 0.3X Danieau’s buffer (by diluting 1X Danieau’s buffer: 58 mM NaCl, 0.7 mM KCl, 0.4 mM MgSO4, 0.6 mM Ca(NO3)2, and 5.0 mM HEPES (pH 7.6), with double distilled water) until observation or fixation. Embryos will be staged according to Kimmel et al.
[92], and stages are given as hours post-fertilization.
Sequence analysis of calreticulin
The homologous calreticulin genes of human, mouse and zebrafish were identified from the NCBI database. The genomic sequence alignment and phylogenetic tree were carried out using Ensembl and MEGA4.3, respectively.
Total RNA isolation and RT-PCR in zebrafish
A reverse-transcription polymerase chain reaction (RT-PCR) will be performed on total RNA extracted from embryos at the designated times using the Trizol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. RNA will be subsequently treated with DNase I (Invitrogen), and first-strand cDNA will be synthesized. cDNA will be generated from total RNA of zebrafish larvae using the SMART kit (Clontech, Mountain View, CA) according to the manufacturer’s
instructions.
Morpholino oligonucleotide (MO) microinjections
Antisense MOs will be purchased or custom made by Gene Tools, LLC (Philomath, OR). A standard control MO (stdMO,
5’-CCTCTTACCTCAGTTACAATTTATA-3’) with no sequence homology to any known zebrafish sequences will be used. To knock down the target genes activity, we used two different MOs targeting the translation initiation site and the splicing donor site, respectively. The MOs will be dissolved in sterile double-distilled water to 1 mM and stored at -20 °C.MOs will be diluted to the desired working concentrations in 1X Danieau’s buffer with 0.5% phenol red and stored at 4 °C before being used.
Thin-wall (1 mm (o.d.) × 0.75 mm (i.d.)) glass capillaries with filaments (A-M Systems, Carlsborg, WA) will be pulled using a horizontal puller (P-97, Sutter Instrument, Navato, CA). Embryos at desired stages will be immobilized at an injection trough on a 100-mm 2% agar plate. MOs will be prepared as described at designated concentrations. An injection pipette will be forced into the chorion and the yolk cells to reach the junction between the yolk cells and blastodisc where the solution will be ejected by using a pressure injector (IM-300, Narishige, Japan). After injection, embryos will be recovered from the injection troughs and cultured in 0.3X Danieau’s buffer at 28.5 °C until being examined.
Cell culture
The NB cell lines SH-SY5Y (ATCCH CRL-2266TM) and SK-N-DZ (ATCCH CRL-2149TM) were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA), while the stNB-V1 was kindly provided by Dr. Yung-Feng Liao of Academia Sinica. These cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM)/High glucose medium (Biowest) containing 10% fetal bovine serum (FBS) and 1% penicillin/streptomycin (Invitrogen). The cells were grown in a humidified atmosphere containing 5% CO2 and 95% air at 37°C. Using a
hemocytometer and trypan blue, cells were sub-cultured in 10 cm plates at a density
of 106 cells per plate.
Transfection
The cells were seeded in 3.5 cm plates at density of 3*105 cells and were transfected with construct plasmids using Lipofectamine 2000 (Invitrogen). Every plate was transfected with 4 μg of the plasmid and 10 μL of the Lipofectamine 2000 in serum-free DMEM. After 8 hours of transfection, the medium was changed into DMEM with 10% FBS. The cells were harvested after 48 h.
Cells transfected with a vector, without the insert gene, were used as vehicle control and cells treated with ddH2O were used as negative control. The construction of CRT expression vector pEGFP-C1-CRT and CRT shRNA vector
pCR3.1-CRTshRNA was as previously described [28].
Construction of stable cell lines
Tetracycline-inducible CRT expression vector pLKO_AS3.1-p5CRT-HY was constructed. pLKO_AS3.1 was the control vector of the inducible vector. CRT shRNA expression vector was purchased from the National RNAi Core Facility Platform, Academia Sinica (Taipei, Taiwan). The shRNA target sequence was: 5’-
CCAGTATCTATGCCTATGATA-3’ (shCRTa, TRCN0000019989), 5’-
CGTCTACTTCAAGGAGCAGTT-3’ (shCRTb, TRCN0000019991). pLKO.1 was the control vector of the shRNA plasmid.
Lentiviral stocks were produced by calcium phosphate transfection. Around 30-40% confluent 293T cells in T25 flasks were prepared and transfected with a DNA mixture containing 7.5 g packaged plasmids and 7.5 g lentivector of the target gene for 16 h. The transfected condition medium was replaced with 12 ml fresh DMEM containing 10 mM sodium butyrate. After 24 h, all conditioned medium were harvested and treated with the stNB-V1 cells for infection. Cells were selected by 1 μg/μL puromycin (InvivoGen, USA).
RNA isolation and reverse-transcription (RT)
Total cellular RNA was extracted using the TRIzol reagent (Invitrogen).
Complementary DNA was synthesized with 1 µg total RNA using a Toyobo RT-polymerase chain reaction (PCR) kit (Toyobo, Osaka, Japan).
Quantitative real-time PCR
The real-time PCR with the mixture reagent KAPA SYBR-Green as the fluorescent dye (Bio-Rad) was conducted on a Mini-Opticon real-time detection system (Bio-Rad, Hercules, CA, USA). Gene-specific primers were used and the
specificity was confirmed by single melting-curve after real-time PCR. Cycling
conditions were 95°C for 3 min, followed by 30 cycles of 95°C for 30 s, 60°C for 30 s, and 72°C for 30 s.
For quantification, the target gene was normalized to the GAPDH to act as an internal control for SH-SY5Y and SK-N-DZ cells and heat shock protein 60 (HSP 60) for stNB-V1. P53, Bcl-2 and Bax were used as apoptotic markers [93]. Primers for the real-time PCR were: GAPDH (F-5’-AAG GTG AAG GTC GGA GTC-3’ and
R-5’-TGT AGT TGA GGT CAA TGA AGG-3’); HSP60 (F-5’- CA CCG T AA GCC TTT GGT CAT-3’ and R-5’- CTT GAC TGC CAC AAC CTG AA-3’); CRT (F-5’-CC TCC TCT TTG CGT TTC TTG-3’ and R-5’-CAG ACT CCA AGC CTG AGG AC);
HIF-1α (F-5’-CAT AAT GTG AGT TCG CAT CT-3’ and R-5’-ATA TCC AAA TCA CCA GCA TC); VEGF-A (F-5’-GGC ACA CAG GAT GGC TTG AAG-3’ and R-5’-GGC ACA CAG GAT GGC TTG AAG-3’); p53 (F-5′-TGA CTG TAC CAC CAT CCA CTA-3′ and R-5′-AAA CAC GCA CCT CAA AGC-3′); Bax ((F-5′
-TGC TTC AGG GTT TCA TCC AG-3′ and R-5′-GGC GGC AAT CAT CCT CTG-3′); Bcl-2 (F-5′-AGG AAG TGA ACA TTT CGG TGA C-3′ and R-5′- GCT CAG TTC CAG GAC CAG GC-3′); GAP43 (F-5′-TCC GTC GAC ACA TAA CAA -3′ and R-5′-CAG TAG TGG TGC CTT CTC C-3′); neuron-specific enolase (NSE) (F-5′-TGT CTG CTG CTC AAG GTC AA-3′ and R-5′-CGA
TGA CTC ACC ATG ACC C-3′); neurofilimant-H (NFH) (F-5′-CCG ACA TTG CCT CCT ACC-3′ and R-5′-GAG CCA TCT TGA CAT TGA GCA-3′); TrkA (F-5′-TTG GCA TGA GCA GGG ATA TCT-3′ and R-5′-ACG GTA CAG GAT GCT CTC GG-3′).
Western blot analysis
Total proteins were extracted from cells using lysis buffer (20 mM Tris, pH 8.0, 150 mM NaCl, 1% NP-40, 1 mM Na3VO4, and 10% glycerol) with 10% protease inhibitor cocktail. The cells were lysed for 15 min on ice and then spun at 4°C and 13,000 rpm for 15 min. The supernatant was then collected for Western blotting. A Bio-Rad protein assay kit was used to measure protein concentration.
Concentration-normalized lysates were boiled at 100°C in an SDS sample buffer for 5 min. Proteins were fractionated by SDS-polyacrylamide gel electrophoresis (PAGE) (80 volts for 30 min in stacking gel and 120 volts for 1.5 h in running gel) and transferred to nitrocellulose membranes (100 Volts for 60 min).
The membranes were blocked with 5% BSA in TBS-T (0.1% Tween 20 in TBS), followed by overnight incubation at 4°C with appropriate dilutions of primary
antibody in 1% TBS-T. After three washes with TBS-T (5 min each), the membranes were incubated with the appropriate secondary antibody coupled with horseradish
peroxidase. Immuno-complexes were visualized using an enhanced
chemi-luminescence (ECL) kit according to the manufacturer’s instructions.
The antibodies used were as follows: rabbit polyclonal anti-CRT antibody (Upstate Biotechnology, Lake Placid, NY), rabbit monoclonal anti-VEGF-A antibody (Santa Cruz, CA, USA), goat polyclonal anti-β-actin (Santa Cruz, CA, USA), and goat monoclonal anti-GAPDH antibody (Genetex, USA).
Enzyme-linked immuno-sorbent assay (ELISA) for VEGF-A secreted proteins in
the conditioned medium
The cells were seeded in six-well plates at 3x105/well. The conditioned medium was collected after 48 h of transfection and analyzed by ELISA specific for human VEGF (R&D Systems, Minneapolis, MN, USA), following the manufacturer’s instructions. The conditional medium was previously centrifuged to remove cells and other unnecessary particles. Samples and VEGF-A standards were added into
micro-plates that were pre-coated with anti-human VEGF-A capture antibody. The micro-plates were incubated at room temperature on a horizontal orbital shaker for 2 hours. After incubation, samples and standards were discarded and washed with wash buffer 4 times.
After aspiration of conjugates and a further wash, the substrate solution was
added to each well, which were incubated for 30 min. After the reaction was completed, stop solution was added to end the reaction. The optical density of each well was measured with an ELISA plate reader set to a wavelength of 450nm.
Cell Proliferation Assay
Cells were seeded in the 96-well plate at density of 103/100 μL, 5x103/100 μL and 104/100 μL respectively. Cells cultured in serum-free medium were used as
negative control. After tetracycline induction for 48 hours, MTT reagent (Sigma) was added to each well to a final concentration of 0.05% for reaction. After incubation at 37℃for 4 hours, MTT-containing medium were removed and 50 μL dimethyl sulfoxide (DMSO) were added for 20 minutes at 37℃ to dissolve formazan.
Reactions were monitored by 96-well ELISA plate reader at 595nm.
Apoptosis Detection Assay
106 cells were seeded in 10 cm plate. Cells treated with 1 μg/μL
actinomycin-D (Sigma, St.Louis, MO) were used as positive control. After
tetracycline induction for 24 hours, stNB-V1 cells were harvested and washed by cold PBS twice. Cell apoptosis rate was detected by using fluorescein isothiocyanate (FITC) Annexin V apoptosis detection kit (BD, Pharmingen, San Diego, CA). 105
cells were suspended in 100 μL of 1 x binding buffer. Harvested cells were then stained by 10 μL of FITC-conjugated Annexin V antibody and propidium iodide for
15 minutes. The stained cells were analyzed by BD FACSCanto2 cell flow cytometry.
VEGFR-1 Blockade
2x105 stNB-V1 cells were cultured in 6-well plate. Cells were treated with 1μg/mL goat polyclonal anti-human VEGF R1 antibody to block the VEGF-A signaling. Normal goat IgG was used as the negative control.
Patients and sample Preparation
A cohort of histologically proven NB patients with complete follow-up were enrolled in this study. Tumor samples were obtained during surgery and imeditely frozen in liquid nitrogen. The categorization of tumor histology was based on the International Neuroblastoma Pathology Classification scheme [94].
Immunohistochemical staining
A total of 69 tumor specimens collected before chemotherapy were fixed and embedded in paraffin. The expression of CRT, VEGF-A and CD34 was assayed using an avidin–biotin complex immunoperoxidase staining technique on archival
paraffin-embedded tissue specimens obtained before chemotherapy. Tissue sections (5mm) of tumors were deparaffinized and rehydrated in a routine manner. After microwave pretreatment, the CRT, VEGF-A and CD34 antibodies were then applied at a dilution of 1:150 overnight at 48C. The N-Histofine Simple Stain MAXPO (Nichirei, Tokyo, Japan) was then applied for 30 min at room temperature.
Diaminobenzidine was used for visualization and nuclei were counterstained with hematoxylin. One ganglioneuroma tumor with consistent CRT expression by immunohistochemistry was used as a positive control in each staining.
Non-immunized rabbit serum was used as a negative control. Tumors with various differentiating histologies were included in each staining. The immunoreactivity of CRT, VEGF-A and CD34 was assessed by a single pathologist who was blinded to the clinical background of the patients.
Mouse xenograft studies with inducible CRT stNB-V1 cells
5x106 CRT inducible stNB-V1 cells were injected subcutaneously into four-week old female athymic nude mice with matrigel (BD Bioscience). Mice were randomized into two treatment groups. Tumor-bearing mice were treated with doxycycline in their daily drinking water (2g/L) or vehicle alone (sucrose) for 21 days. The growth ability of xenografted tumors on animals was measured according to the metric measurement
of tumor size. Tumor diameters were measured with calipers, and volumes were calculated as LxW2x0.5, where L and W are the tumor length and width in mm, respectively.
Statistical analysis
The correlation between CRT and VEGF-A mRNA expression level were
analyzed using non-parametric Wilcoxon rank-sum test and Spearman’s correlation test. Other data analyses were performed using one-way analysis of variance (ANOVA), followed by Fisher’s protected least-significant difference (LSD) test (StatView; Abacus Concept, Berkeley, CA, USA). Each result was obtained from at least three independent experiments and expressed as mean ± standard deviation.
Statistical significance was set at p<0.05.
Chapter III.
Results
Cloning and gene analysis of zebrafish CRT (zCRT) showed high sequence
homology
To investigate the role of CRT in zebrafish development, we have cloned the full length transcript of zCRT..The zebrafish zCRT are highly homologous to their human and mouse counterparts with protein sequence similarity close to 90% as shown in the calreticulin sequence alignment graph (Fig. 2A). The evolutional distances of zCRT to other species’ CRT are shown in the phylogenetic trees (Fig. 2B). To further
investigate the genetic conservation of CRT in vertebrate including zebrafish, mouse and human, we compared their chromosome location maps from the Ensemble database. The results showed that all CRT genes are located near rad23a. The gene orientation of zCRT is the same as that of mouse CRT but is opposite to that of human CRT (Fig. 2C).
Expression profiles of CRT in embryonic development of zebrafish
To evaluate the temporal and spatial expression analysis of zCRT, the
quantitative RT-PCR (qRT-PCR) was performed. Our studies revealed that zebrafish CRT could be detected very early and reduced gradually during the segmentation and pharyngula stages (Fig. 3). The whole mount in situ hybridization (WISH) analysis of zCRT from ZFIN showed that zCRT was highly expressed before 5 dpf, compatible
with our expression analysis of qRT-PCR. Besides, the spatial analysis from ZIFN revealed that zCRT was expressed mainly in hatching gland, floor plate,
chordo-neural hinge and lateral line in embryos. However, in adult fish (older than three months), the zebrafish CRT were detected in all organs by qRT-PCR analyses (Fig. 4).
Knockdown of zCRT protein expression by morpholinos cause an embryonic
defect and developmental retardation
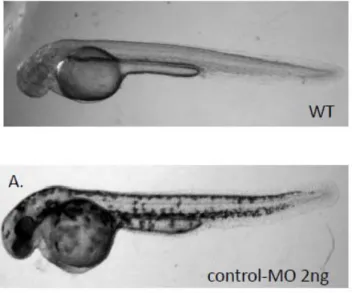
Microinjection of morpholinos targeting the ATG start site into embryos effectively abolished the expression of zCRT protein. Knockdown of zCRT expression resulted in severe embryonic lethality within 3 dpf. The surviving morphants showed developmental retardation, such as slow growth, reduced brain size and heart edema compared to the wild type fish and standard control-MO fish (Fig. 5A). The severe embryonic defect of the zCRT knockdown prevented us from studying its impact on locomotive movement. Both heart edema and phenotype change are morpholino concentration-dependent (Fig. 5B).
CRT positively regulated VEGF-A and HIF-1 expressions
To investigate the relationship between CRT and VEGF-A, CRT was
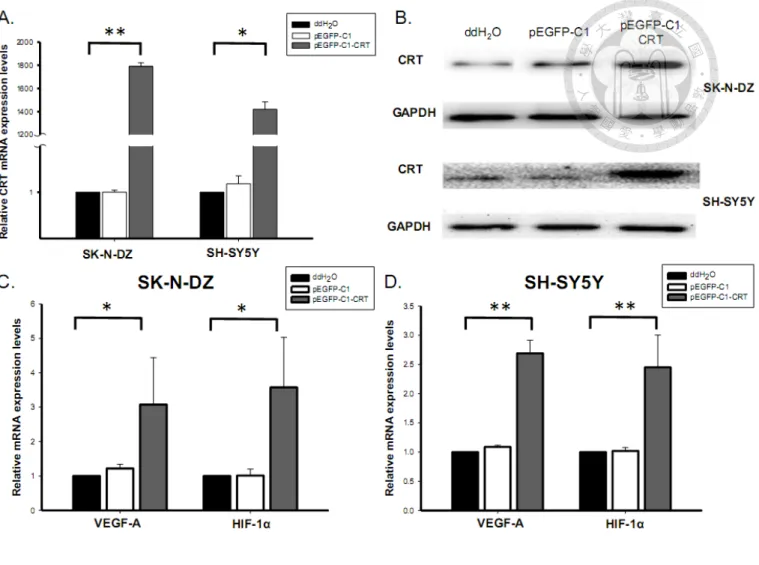
over-expressed using pEGFP-C1-CRT expression vector via Lipofectamine 2000 transfection system in SK-N-DZ and SH-SY5Y cells, which was performed by Kuan-Hung Lin. According to real-time PCR analysis, the expression vector
significantly enhanced CRT mRNA expression in SK-N-DZ and SH-SY5Y to 1800- and 1400-folds higher, respectively, compared to the negative control (none) and vector control (pEGFP-C1) (Fig. 6A). This CRT over-expression at the protein level was also confirmed by western blotting (Fig. 6B).
To elucidate the effects of CRT on VEGF-A and HIF-1, VEGF-A mRNA
expression was analyzed in transiently CRT-over-expressing NB cells. The
over-expression of CRT increased VEGF-A mRNA expression in both SK-N-DZ and SH-SY5Y cells (Fig. 6C&D). The mRNA expression level of HIF-1, a well-known
up-regulator of VEGFs in NB, also positively correlated with CRT level [95].
Elevated HIF-1 expression suggested that HIF-1 might be involved in the
CRT-dependent VEGF-A up-regulation.
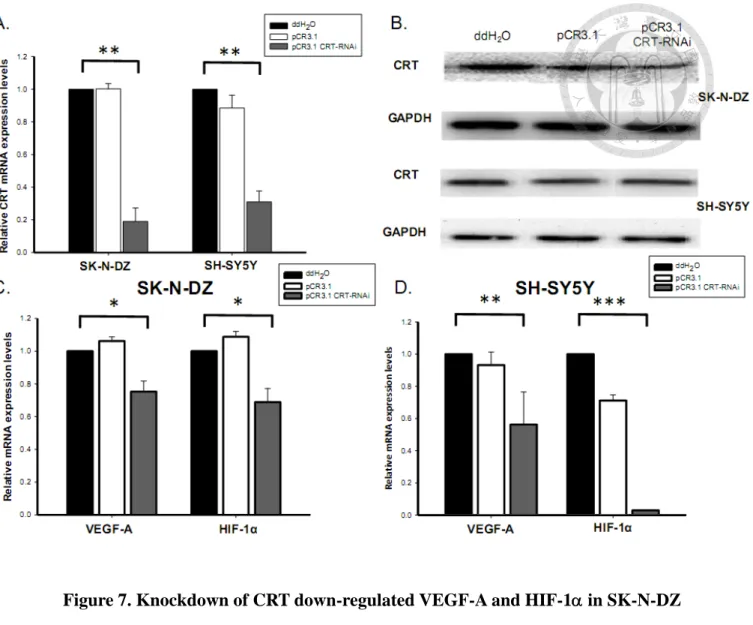
To further clarify the relationship between CRT and VEGF-A, CRT was
transiently knocked-down using shRNA in SK-N-DZ and SH-SY5Y cells. According to real-time PCR analysis, the CRT mRNA expression levels were significantly inhibited by the pCR3.1-CRT-shRNA in both SK-N-DZ and SH-SY5Y cells (Fig.
7A). The knockdown of CRT was further confirmed at the protein level by western
blotting (Fig. 7B).
To examine the effect of CRT knockdown on VEGF-A and HIF-1, VEGF-A and HIF-1 mRNA expressions were analyzed in transiently CRT-knocked down NB cells. The VEGF-A and HIF-1 mRNA expression were lower in SK-N-DZ and
SH-SY5Y cells with CRT knockdown (Fig. 7C&D). The studies about the CRT knockdown were also performed by Kuan-Hung Lin. The results further confirmed that CRT could regulate VEGF-A and its up-regulator, HIF-1 expression in NB cells.
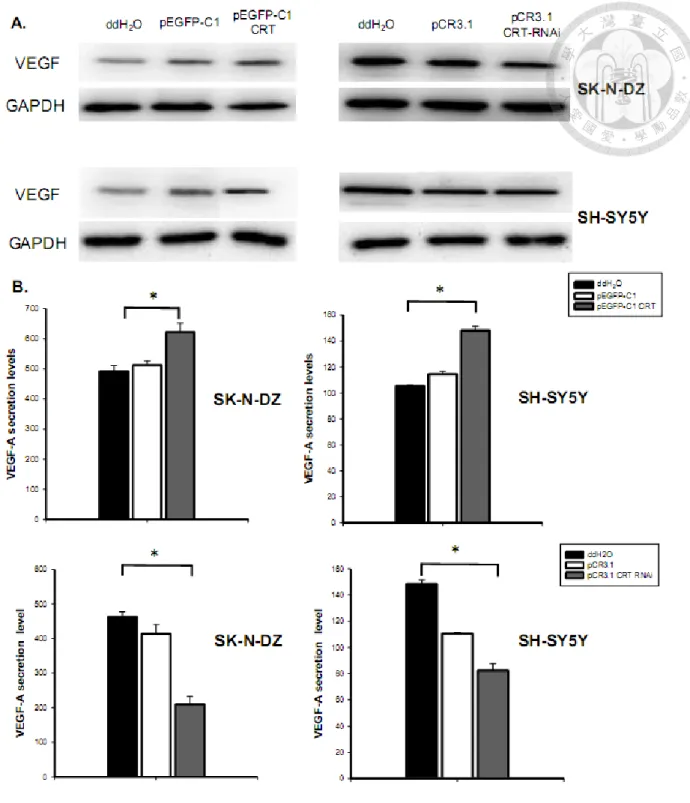
Effects of CRT on VEGF-A protein expression and secretion level in NB cells
Whether the CRT expression affected the VEGF-A protein expression in NB
cells was further determined. Results showed that VEGF-A protein expression was up-regulated by CRT over-expression both in SK-N-DZ and SH-SY5Y cells (Fig. 8A).
Moreover, knockdown of CRT decreased the protein expression of VEGF-A (Fig. 8A).
The results demonstrated that CRT positively regulated VEGF-A both at the mRNA and protein level in different NB cells.
The VEGF family members, including VEGF-A, are known to be secreted polypeptides. The VEGF-A secretion level was then examined in the conditioned media. Results showed that the CRT over-expression up-regulated VEGF-A secretion in the conditioned media of SK-N-DZ and SH-SY5Y cells (Fig. 8B). On the other
hand, CRT knockdown suppressed the VEGF-A protein secretion in conditioned media (Fig. 8B). These results further supported the effects of CRT on VEGF
expression and secretion in different NB cells. The results about the VEGF-A protein expression and secretion level were provided by Kuan-Hung Lin.
Establishment of stable cell lines
In order to further clarify the effects of CRT on NB cell lines, CRT
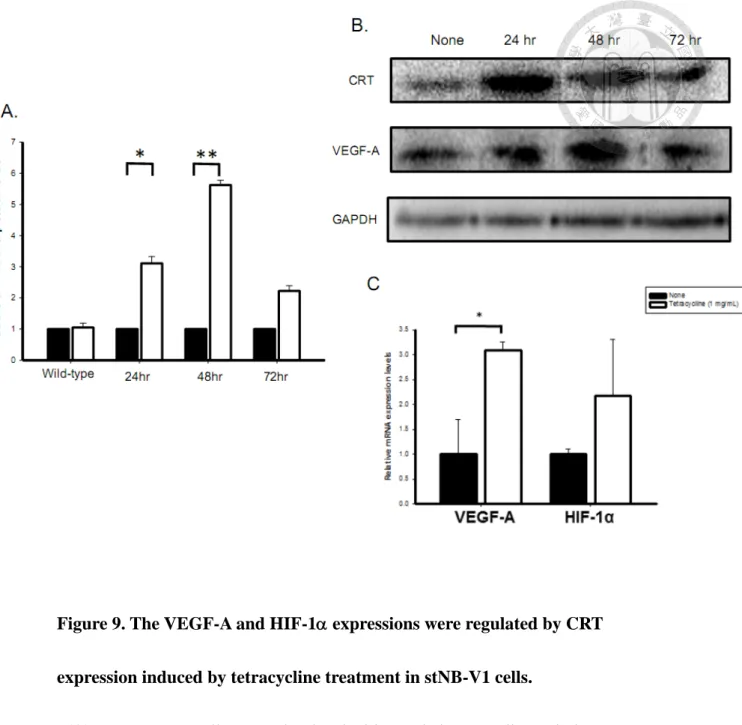
over-expression and knockdown stable cell lines were selected. However, constitutive over-expression of CRT led to NB cell differentiation without proliferation. Thus, kindly provided by Dr. Yung-Feng Liao and Dr. Bo-Jeng Wang, we received an inducible-CRT stNB-V1 cell line by a tetracycline-regulated gene system. To induce CRT expression, cells were treated with tetracycline. After 1 μg/μL tetracycline induction for 24, 48, and 72 h, CRT mRNA expression was elevated by 3-, 6-, and 4-folds higher, respectively, than non-induced cells (Fig. 9A). As such, 48 h induction was chosen as the best condition for subsequent experiments. By western blotting, the protein expression of CRT was also enhanced by tetracycline induction (Fig. 9B).
For knockdown stable cell lines, stNB-V1 NB cells were transfected with a CRT-shRNA plasmid via lentiviral system and then selected by respective antibiotics.
After selection by puromycin, CRT-shRNA cells were generated (19989 and 19991)
and CRT knockdown efficiency was confirmed by real-time PCR (Fig. 10A).
CRT positively regulated VEGF-A and HIF-1 expressions in stable cell lines
The expressions of VEGF-A and HIF-1 were examined in inducible-CRT stNB-V1cells. After 48 h of tetracycline treatment, the mRNA expressions of
VEGF-A and HIF-1 were significantly augmented (Fig. 9C). The positive regulation of CRT on VEGF-A protein expression was also demonstrated in inducible-CRT stNB-V1cells (Fig. 9B), revealing that CRT positively regulated VEGF-A and HIF-1α expressions in different NB cells.
The VEGF-A and HIF-1 mRNA expression levels were further examined in
CRT knockdown stable cell lines (stNB-V1, 19989) and revealed lower mRNA expressions (Fig. 10B). These results further confirmed that CRT regulated the VEGF-A and HIF-1α expression levels in different NB cells.
Apoptotic rate of NB cells was not affected by CRT over-expression
To clarify the impact of CRT over-expression on NB cell apoptosis, we use Annexin V/propidium iodide staining to determine the apoptotic cells in
inducible-CRT stNB-V1 cells. Actinomycin-D, a potent inducer for apoptosis in several cell lines, was used as positive control. After tetracycline treatment, the
fluorescence intensity of apoptotic cells was not different from cells without
tetracycline treatment (Fig. 11A), which indicated that CRT over-expression did not affect apoptosis in stNB-V1 cells. The mRNA expression levels in several apoptotic markers including p53, Bcl-2 and Bax were further determined by real-time PCR [93].
As shown in Figure 11B, no difference in p53, Bcl-2 and Bax mRNA expressions was observed after tetracycline induction in inducible-CRT stNB-V1 cells. These results further confirmed that over-expression of CRT did not induce NB cell apoptosis.
Cell proliferation rate is lower in CRT over-expressing NB cells
We previously found that constitutive over-expression of CRT led to NB cell differentiation without proliferation in SK-N-DZ and SH-SY5Y cells. Thus, we have generated an inducible-CRT stNB-V1 cell line. In order to further confirm the effect of CRT over-expression on cell proliferation, we used MTT assays to evaluate the cell proliferation rate in stNB-V1 cells. Cells cultured in serum-free medium were used as negative control. After tetracycline induction, the MTT readings were significantly decreased (Fig. 12), indicating that up-regulation of CRT suppressed proliferation of NB cells.
VEGF-A is involved in CRT-induced NB cell differentiation
To further clarify the roles of CRT-dependent VEGF-A up-regulation on NB cells differentiation, the expressions of neuron specific markers including GAP43,
neuron-specific enolase 2 (NSE2) and neurofilimant-H (NFH) as well as TrkA, a molecular marker indicative of NB cell differentiation, were examined in
inducible-CRT stNB-V1cells [96-98]. After tetracycline induction, the mRNA
expression levels of GAP43, NSE, NFH and TrkA were significantly augmented (Fig.
13), which is compatible with our previous study suggesting that CRT is critical for NB differentiation.
We next used anti-VEGFR-1 antibodies to examine whether up-regulated
VEGF-A is involved in CRT-related neuronal differentiation in NB. VEGF-A binds to two related receptor tyrosine kinases, VEGFR-1 and VEGFR-2, and regulates
downstream biological signaling. Antibodies against VEGFR-1 could partially block the VEGF-A signaling. As shown in Fig. 13, after treatment with anti-VEGFR-1 antibodies, the mRNA expressions of GAP43, NSE2, NFH and TrkA were suppressed significantly in both CRT over-expression (treated with tetracycline) and controlled (treated with ddH2O) stNB-V1 cells. These results implied the involvement of VEGF-A in neuronal differentiation in NB.
CRT expression suppressed NB tumor growth in mice xenograft model
In previous studies, increased CRT expression was found to be associated with differentiated histologies in NB tumors. Moreover, our present in vitro study using NB cells indicated that CRT could up-regulate VEGF-A and induce neuronal
differentiation in NB cells. To further investigate the roles of CRT-dependent VEGF-A up-regulation on NB tumor growth, we established mice xenograft models of NB using inducible-CRT stNB-V1 cells transplantation. To induce CRT expression, mice were treated with doxycycline in their daily drinking water after tumor cells injection.
During daily doxycycline treatment, the tumor growth was significantly suppressed compared to xenograft mice without doxycycline treatment (Fig. 14). These results further suggested the essential role of CRT in the tumor progression of NB.
VEGF-A is upregulated in CRT-induced NB cell differentiation in mice xenograft
model
Our in vitro study using NB cells indicated that CRT could up-regulate VEGF-A and induce neuronal differentiation in NB cells. In order to confirm the effect of CRT
in vivo, we have established the mice xenograft models. Tumor inoculated mice were
treated with doxycycline in their daily drinking water (2g/L) to induce CRT expression. Tumor inoculated mice were sacrificed after 21-days doxycycline
treatment for CRT induction and the tumor was removed for experiments. The mRNA
expression levels of VEGF-A and HIF-1 were significantly increased, suggesting the
positive regulation of CRT on VEGF-A in xenograft NB tumors (Fig. 15). To further confirm the roles of CRT-dependent VEGF-A up-regulation on NB cell differentiation
in vivo, the expressions of neuro-specific markers, GAP43, was examined. After
doxycycline induction, the mRNA expression level of GAP43 was significantly augmented (Fig. 15), which is compatible with our in vitro study suggesting that CRT is critical for NB differentiation.
VEGF-A expression was positively correlated with CRT and differentiated
histology in human NB tumors
Both our in vivo and in vitro studies have demonstrated the positive regulation of CRT on VEGF-A. To further verify the correlation between CRT and VEGF-A, the expressions of CRT and VEGF-A in 56 primary NB tumor were determined by real-time PCR. The results revealed a significantly positive correlation between CRT and VEGF-A (Fig. 16, Spearman’s = 0.648, p <0.001). Furthermore, from 31
primary NB tumors analysis, we found that differentiated NB exhibited higher CRT and VEGF-A expression levels than undifferentiated NB (Fig. 17A). The CRT and VEGF-A expression were significantly increased in differentiated NB (Fig 17B&C).
These results further supported the critical roles of CRT and VEGF-A in neuronal
differentiation in NB tumorigenesis.
VEGF-A expression and clinicopathologic and biological factors
In order to investigate the clinical significance of VEGF-A as well as its relationship with CRT and other clinicopathologic factors in NB, we examined the protein expression of CRT, VEGF-A and CD34 in 69 NB tumors by
immunohistochemical staining. Both CRT and VEGF-A were commonly found in more differentiated NB than in undifferentiated NB (Fig. 18). Our
immunohistochemical data showed that a positive expression of VEGF-A could be detected in 27 of 69 NB tumors. Positive VEGF-A protein expression was
significantly associated with favorable biomarker of CRT expression (P = 0.001).
Furthermore, we found that positive VEGF-A expression was significantly correlated to differentiated tumor histology (P = 0.013). There was also an inverse association between VEGF-A expression and MYCN amplification (P = 0.034). Interestingly, there was no correlation between the protein expressions of VEGF-A and CD34, a marker of endothelial cells (P = 0.808). We found that CD34, but not VEGF-A, is highly expressed in undifferentiated NB (Fig. 18). Other adverse prognostic factors, including older patient age at diagnosis and advanced clinical stage were not
associated with VEGF-A expression in our study. The relationship between VEGF-A
protein expression and the clinicopathologic and biological variables of NB tumors was summarized in Table 1.
VEGF-A expression in NB tumors predicts a favorable outcome
To further clarify the relationship between the expression of VEGF-A and clinical outcome, Kaplan-Meier analysis was used to predict the five-year survival rate. The survival analysis revealed that patients with positive VEGF-A expression have a significantly higher survival rate compared to patients with negative VEGF-A expression (Fig 19; P = 0.013, Log-rank test), suggesting that positive VEGF-A expression could predict a favorable outcome in NB patients.
Chapter IV.
Discussions
The tumorigenesis of NB could be a divergence of the embryonic development of sympathetic nervous system. ER chaperones including CRT and GRP78 are suggested to participate during embryonic development in our previous review. CRT, a
well-known multi-functional protein, is located not only in the cytosol but also on the cell surface and in the extracellular environment to modulate a number of physiologic and pathologic conditions [7, 16, 63]. Here, we found CRT is highly conserved across species, with an amino acid identity of approximately 90% from the invertebrate to mouse and human [7]. In the human and mouse genomes, CRT is known to be a single-copy gene [10]. In zebrafish, CRT also appears to be a single copy gene. The high sequence homology implies that CRT may be functionally conserved in zebrafish.
Studies in CRT knockout mice which demonstrated the essential role of CRT for embryogenesis have suggested the important role of CRT in nervous system
development as well as the biology of NB [69]. Our temporal and spatial analysis of CRT expression in zebrafish found that CRT is highly expressed in early embryonic development and is mainly in hatching gland, floor plate, chordo-neural hinge and lateral line in embryos, supporting the crucial role of CRT in neuronal development.
Furthermore, our study used anti-sense morpholino oligonucleotide to knock down of zCRT protein expression at different levels. The results showed a