中國醫藥大學機構典藏 China Medical University Repository, Taiwan:Item 310903500/41330
273
0
0
全文
(2) 目錄 謝誌............................................................................................................................I 縮寫表 .....................................................................................................................II 中文摘要 ................................................................................................................III Abstract ....................................................................................................................V 第一章. 前言.........................................................................................................1. 第二章. 文獻探討.................................................................................................3. 第一節. 樟芝及其主成分介紹.........................................................................4. 第二節. 皮膚癌介紹.........................................................................................8. 第三節. Wnt/-catenin pathway 介紹.............................................................18. 第四節. Wnt/-catenin pathway 與細胞凋亡之相關性.................................27. 第五節. Wnt/-catenin pathway 與細胞週期之相關性.................................35. 第六節. Wnt/-catenin pathway 與細胞轉移之相關性.................................42. 第三章. 研究動機與假說....................................................................................49. 第一節. 研究動機...........................................................................................50. 第二節. 研究假說...........................................................................................51. 第三節. 實驗設計...........................................................................................52. 第四章. 材料方法..................................................................................................55. 第一節. 實驗儀器...........................................................................................56. 第二節. 實驗材料...........................................................................................58. 第三節. 實驗方法...........................................................................................64. 第一部分 AC-10 1. 樟芝發酵液 (AC-10).............................................................................. 64 2. 細胞培養(cell culture).......................................................................64 i.
(3) 3. 細胞形態(morphology)之觀察.........................................................66 4. 細胞存活率分析(cell viability).........................................................66 5. 細胞凋亡試驗(TUNEL assay)..........................................................68 6. 蛋白質萃取(protein extraction)........................................................70 7. 西方墨點(western blotting)分析.......................................................73 8. 流式細胞儀分析(flow cytometry)....................................................78 9. 免疫螢光染色(immunofluenrence)................................................80 10. 轉型作用(transformation)...............................................................82 11. 質體抽取(plasmid DNA extraction)................................................84 12. 轉染作用(transfection)....................................................................88 13. 報導基因活性測定(reporter assay).........................................................90 14. 抑制劑之處理..................................................................................92 15. RNA 萃取(RNA extraction)..............................................................95 16. 逆轉錄反應(RT-PCR)......................................................................95 17. 細胞群落形成分析(colony formation assay)..................................98 18. 細胞遷移試驗(Wound Healing assay)..........................................99 19. 細胞移行試驗(Transwell invasion assay)......................................100 20. 統計分析.........................................................................................101 第二部分 AC-0 1. AC-10 主成分(AC-0)...........................................................................102 2. 細胞培養(cell culture).......................................................................102 3. 細胞形態(morphology)之觀察..........................................................103 4. 細胞存活率分析(cell viability)..........................................................103 5. 細胞凋亡試驗(TUNEL assay)............................................................103. ii.
(4) 6. 蛋白質萃取(protein extraction)..........................................................103 7. 西方墨點(western blotting)分析........................................................103 8. 免疫螢光染色(immunofluenrence)....................................................103 9. 轉染作用(transfection).......................................................................104 10. 報導基因活性測定(reporter assay).........................................................104 11. 抑制劑之處理...................................................................................104 12. 免疫沉澱分析(Immunoprecipitation)..............................................104 13. RNA 干擾(RNA interference, RNAi)...................................................106 14. Annexin V staining..............................................................................108 15. RNA 萃取(RNA extraction)................................................................109 16. 逆轉錄反應(RT-PCR)........................................................................109 17. 細胞群落形成分析(colony formation assay)....................................110 18. 細胞遷移試驗(Wound healing assay)............................................110 19. 細胞移行試驗(Transwell invasion assay)..........................................110 20. 腫瘤異種移植(Tumor Xenograft study)..............................................110 21. 免疫化學組織染色(immunochemistry staining)................................111 22. 組織細胞凋亡分析(tissue TUNEL assay)..........................................114 23. 組織病理分析(H&E staining).............................................................116 24. 統計分析..............................................................................................117 第五章 實驗結果與圖表..................................................................................118 第一部分 樟芝發酵液(AC-10)對小鼠黑色素瘤細胞 B16F1、B16F10 之影響 第一節 樟芝發酵液(AC-10)對 B16F1、B16F10 小鼠黑色素瘤細胞之存活 率與生長情形的影響....................................................120 第二節 樟芝發酵液 (AC-10)對 B16 F1、B16 F10 小鼠黑色素 瘤細胞. iii.
(5) W n t / - c a t e n i n p a t h w a y 之調控....................................121 第三節 樟芝發酵液(AC-10)誘發 B16F1、B16F10 小鼠黑色素瘤細 胞 凋 亡 ......................................................................................125 第四節 樟芝發酵液(AC-10)對 B16F1、B16F10 小鼠黑色素瘤細 胞 週 期 影 響..............................................................................126 第五節 樟芝發酵液(AC-10)對 B16F1、B16F10 小鼠黑色素瘤細 胞 轉 移 影 響.............................................................................. 127 第二部分 AC-10 主成分(AC-0) 對小鼠黑色素瘤細胞 B16F10 之影響 第一節 AC-10 主成分(AC-0)對小鼠黑色素瘤細胞 B16F10 之存活率與生長 情形的影響......................................... .......................162 第二節 AC-10 主成分(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 W n t / - c a t e n i n p a t h w a y 之調控......................................163 第三節 AC-10 主成份(AC-0)誘發高轉移性小鼠黑色素瘤 B16F10 細 胞 凋 亡 ........................................................ ...........................167 第四節 AC-10 主成份(AC-0)對高轉移性小鼠黑色素瘤 B16F10 細 胞 轉 移 之 影響...........................................................................168 第五節 -catenin siRNA 參與 AC-10 主成份(AC-0) 調控 Wnt/-catenin pathway 之細胞凋亡、細胞增生以及細胞轉移作用...........170 第六節AC-10主成分(AC-0)抑制裸鼠皮下植入小鼠黑色素瘤細胞 B16F10之腫瘤生長................................................................171 第六章. 綜合討論.........................................................................................217. 第七章. 結論...........................................................................................228. 第八章. 參考文獻...................................................................................233. iv.
(6) 圖表目錄 圖 2-1.樟芝發酵的各種型態............................................................................................5 圖 2-2.台灣皮膚癌年齡標準化發生率之長期趨勢(依性別),1979-2007...................10 圖 2-3.台灣非惡性黑色素瘤年齡標準化發生率之長期趨勢(依性別),1979-2007...10 圖 2-4.台灣惡性黑色素瘤年齡標準化發生率之長期趨勢(依性別),1979-2007.......11 圖 2-5.黑色素瘤評鑑準則示意圖...................................................................................14 圖 2-6. Canonical Wnt/-catenin pathway.......................................................................19 圖 2-7.泛素化降解作用示意圖.......................................................................................24 圖 2-8.Wnt/-catenin pathway 的突變............................................................................26 圖 2-9.細胞凋亡與細胞壞死的區別...............................................................................28 圖 2-10.細胞凋亡的三種路徑.........................................................................................32 圖 2-11.細胞週期分佈圖.................................................................................................36 圖 2-12.細胞週期各調控蛋白分佈示意圖.....................................................................39 圖 2-13.Wnt/-catenin pathway 可被活化之細胞週期調控示意圖...............................41 圖 2-14.為癌細胞轉移示意圖.........................................................................................42 圖 2-15.為金屬基質蛋白酶分類圖.................................................................................44 圖 2-16.癌症與血管新生作用示意圖.............................................................................47 圖 4-1.蛋白質轉印...........................................................................................................76 圖 4-2.轉型示意圖...........................................................................................................82 圖 4-3.為 Plasmid Extraction kit 操作流程模擬圖.........................................................85 圖 4-4.為 liposome 轉染示意圖......................................................................................88 圖 4-5. MG132 結構式....................................................................................................92 圖 4-6. Cycloheximide 結構式........................................................................................93 圖 4-7. SB216763 結構式...............................................................................................94 v.
(7) 圖 4-8.為模擬細胞早期凋亡之磷脂絲氨酸外翻情形................................................108 圖 5-1.觀察 AC-10 對 B16F1、B16F10 黑色素瘤細胞之細胞形態影響.................129 圖 5-2.觀察 AC-10 對 B16F1、B16F10 黑色素瘤細胞之細胞存活率影響.............130 圖 5-3.樟芝發酵液(AC-10)對小鼠黑色素瘤細胞 B16F1、B16F10 -c a t e n i n 之 影 響.............................................................................................................131 圖 5-4.樟芝發酵液(AC-10)對小鼠黑色素瘤細胞 -c a t e n i n 核 質 蛋 白 表 現 之 影響..............................................................................................................132 圖 5-5.樟芝發酵液(AC-10)對黑色素瘤細胞 -c a t e n i n 入 核 表 現 之 影響.........133 圖 5-6.樟芝發酵液(AC-10)對黑色素瘤細胞中 W nt / -c a t e n i n p a t h w a y 調 控 之 下 游 蛋 白 表 現 影 響 ..........................................................................134 圖 5-7.樟芝發酵液(AC-10)對 -c a t e n i n 蛋 白 的 降 解 調 控 ................................136 圖 5-8.樟芝發酵液(AC-10)對 -c a t e n i n 蛋 白 降 解 調 控 .....................................138 圖 5-9. SB216763 預防 -c a t e n i n 走 向 G S K 3 蛋 白 降 解 調 控 ......................139 圖 5-10.樟芝發酵液(AC-10)對 -c a t e n i n 調 控 之 轉 錄 表 現 影 響 ..................140 圖 5-11.樟芝發酵液(AC-10)對 -c a t e n i n mR N A 表 現 量 的 影 響 ....................141 圖 5-12.樟芝發酵液(AC-10)對 -c a t e n i n 蛋 白 穩 定 性 影 響 ..............................142 圖 5-13.樟芝發酵液(AC-10)誘發黑色素瘤細胞凋 亡 之 影響..................................143 圖 5-14.樟芝發酵液(AC-10)對黑色素瘤細胞內 Bax/Bcl-2 蛋白表現之 影響.........145 圖 5-15.樟芝發酵液(AC-10)對黑色素瘤細胞內 Cytochrome c 蛋白表現 之 影響.............................................................................................................146 圖 5-16.樟芝發酵液(AC-10)誘發黑色素瘤細胞凋 亡 之 蛋白表現..........................147 圖 5-17.樟芝發酵液(AC-10)導致黑色素瘤細胞週期停滯.........................................149 圖 5-18.樟芝發酵液(AC-10)使黑色素瘤細胞週期停滯於 G1 期之蛋白表現..........151 圖 5-19. 樟芝發酵液(AC-10)抑制黑色素瘤細胞轉移表現.......................................153 vi.
(8) 圖 5-20. 樟芝發酵液(AC-10)抑制黑色素瘤細胞侵襲之表現..................................155 圖 5-21. 樟芝發酵液(AC-10)抑制黑色素瘤細胞群聚之表現..................................158 圖 5-22. 樟芝發酵液(AC-10)抑制黑色素瘤 MMP-9、MMP-2 蛋白表現..............159 圖 5-23. 樟芝發酵液(AC-10)抑制黑色素瘤 VEGF 蛋白表現.................................160 圖 5-24. 觀察 AC-10 主成分(AC-0)對不同類型之皮膚細胞之細胞存活率影響...173 圖 5-25. 觀察 AC-0 對高轉移性小鼠黑色素瘤細胞 B1610 細胞型態之影響....174 圖 5-26.AC-10 主成分(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 之 Wn t / -c a t e n i n p a t h w a y p r o t e i n 之 影響........................................175 圖 5-27. AC-10 主成分(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 之 -c a t e n i n 入 核 表 現 影響....................................................................178 圖 5-28. AC-10 主成分(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 -c a t e n i n 入 核 表 現 之 影響..................................................................180 圖 5-29. AC-10 主成分(AC-0)對 W n t / - c a t e n i n p a t h w a y 調 控 之 下 游 蛋 白 表 現 影 響 .................................................................................181 圖 5-30. AC-10 主成分(AC-0)對 -c a t e n i n 蛋 白 的 降 解 調 控 ...........................183 圖 5-31. MG132 預防 AC-0 對 -c a t e n i n 蛋 白 的 降 解 表 現 ..............................184 圖 5-32. AC-10 主成分(AC-0)對降解 -c a t e n i n 之 G S K 3 依 賴 型 調 控 ........185 圖 5-33. AC-10 主成分(AC-0)對 -c a t e n i n 、 A x i n 、 G S K 3 蛋 白 複 合 體 的 調 控 .........................................................................................................186 圖 5-34. AC-10 主成分(AC-0)抑制 -c a t e n i n 調 控 的 轉 錄 能 力 影 響 .............187 圖 5-35. AC-10 主成分(AC-0)對 -c a t e n i n mR N A 表 現 量 的 影 響 .................188 圖 5-36. AC-10 主成份(AC-0)對 -c a t e n i n 蛋 白 穩 定 性 影 響 ...........................189 圖 5-37. AC-10 主成份(AC-0)誘發 B16F10 細胞凋 亡 之 影 響 ..............................190 圖 5-38. AC-10 主成份(AC-0)誘發 B16F10 晚期細胞凋 亡 之 表 現 ......................191 vii.
(9) 圖 5-39. AC-10 主成份(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 Bax/Bcl-2 蛋白表現之 影響..........................................................................193 圖 5-40. AC-10 主成份(AC-0)對高轉移性小鼠黑色素瘤細胞 B16F10 誘發黑色素瘤細胞凋 亡 之 蛋白表現..........................................................194 圖 5-41. AC-10 主成份(AC-0)對高轉移性小鼠黑色素瘤細 胞 B 1 6 F1 0 細 胞 遷 移 之 影響.........................................................................................196 圖 5-42. AC-10 主成份(AC-0)對高轉移性小鼠黑色素瘤細 胞 B 1 6 F1 0 細 胞 侵 襲 之 影響.........................................................................................198 圖 5-43. AC-10 主成份(AC-0)抑制高轉移性小鼠黑色素瘤細 胞 B1 6 F1 0 細胞群聚之表現.............................................................................................200 圖 5-44. AC-10 主成份(AC-0)對 Wnt/-catenin pathway 調控之轉移相關因子 蛋白表現影響.................................................................................................201 圖 5-45. RNAi 參與 AC-10 主成份(AC-0) Wnt/-catenin pathway 調控 之細胞凋亡、細胞增生以及細胞轉移作用.................................................202 圖 5-46.AC-10 主成份(AC-0)對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 腫瘤抑制結果..................................................................................................205 圖 5-47.控制組裸鼠腫瘤組織病理切片(H&E stain) ..................................................207 圖 5-48.實驗組裸鼠腫瘤組織病理切片(H&E stain) ..................................................208 圖 5-49.AC-10 主成份(AC-0)對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 腫瘤蛋白分析..................................................................................................209 圖 5-50. AC-0 對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 免疫組織化學分析..........................................................................................211 圖 5-51.AC-10 主成份(AC-0)對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 腫瘤細胞凋亡分析..........................................................................................213. viii.
(10) 圖 5-52. AC-0 對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 腫瘤蛋白分析.............214 圖 5-53. AC-0 對裸鼠皮下植入小鼠黑色素瘤細胞 B16F10 腫瘤 細胞凋亡蛋白分析..........................................................................................216 表 2-1.CDK inhibitor 抑制 CDK 活性...........................................................................37. ix.
(11) 謝誌 在中國醫藥大學度過兩年的研究生活,回首這段時間,歡笑淚 水與我常在。除了學習到許多實驗技術之外,在這裡也學到很多待人 處事的態度,讓我對於事情的思考面變為多元思考。在失敗的實驗中 吸取經驗以及面對更多的挑戰使我貫徹對事情的責任感。兩年的研究 生活,除了專業態度上,環境的挑戰也無時無刻上演,對此,我感謝 學姐盈禎、男友靖壹、同學俐伶、育瑄的開導,每當在實驗室面臨挫 折時,容忍我的不穩情緒及低潮,給予我許多的安慰與鼓勵,讓我有 勇氣繼續面對許多挫折以及這充滿挑戰的環境。 在論文完成的同時,感謝指導老師楊新玲教授與共同指導老師 許游章教授這些日子的指導,他們培養我獨立思考的精神,並以李遠 哲博士的故事作為一個勵志題材,不斷的鼓勵我向上。另外,也特別 感謝口試委員呂鋒洲老師、林凱元老師以及陳師慶老師對此論文的指 導,使本篇論文的內容更加完整。 另外,感謝學妹佩君、蓉瑩、曉涵在實驗上的幫助以及傾聽心 事;學弟書緯常伴我於夜深人靜的實驗室奮鬥,也是很好的宵夜夥 伴;印度籍博士後研究員 kumar 在實驗上給予意見以及英文會話的練 習;特別要感謝同學可婷在這段日子裡與我的點滴,直到最終,讓我 了解到和我個性有極大差異的她,其實也具有溫柔面所附帶的優點包容力以及對人性的樂觀面。 最重要的,非常感謝我的父母,不僅讓我無後顧之憂的順利完成 學業,也支持我去完成我想做的事情,僅以碩士論文獻給所有曾經幫 助過我的人,感謝!. 鄒曉彤謹致. . 中國醫藥大學 營養系碩士班. I. 中華民國一百年七月.
(12) 縮寫表 AC : Antrodia camphorata APS : ammonium persulfate APC: adenomatous polyposis coli β-TrCP: β-Transducin repeat Containing Protein Dvl: Disheveled DAPI : 4',6-diamidino-2-phenylindole DMSO : dimethylsulfoxide Fzd: Frizzled FBS:fetal bovine serum FITC : fluorescein isothiocyanate GSK3β: glycogen synthase kinase 3β HRP : horseradish peroxidase LRP 5/6:lipoprotein receptor-related protein 5/6 MMP: matrix metalloproteinase NSAID: non-steroidal anti-inflammatory drugs NEAA:non-essential amino acid PARP : Poly (ADP-ribose) polymerase PBS:phosphate-buffered saline PBST : phosphate buffered saline containing-Tween-20 PI : propidium iodide SDS : sodium dodecyl sulfate TEMED : N,N,N,N-tetramethyl-ethylenediamine WB : Western Blotting VEGF: vascular endothelial growth factor. II.
(13) 中文摘要 黑色素瘤(melanoma)為 皮 膚 惡 性 腫 瘤 的 一 種 , 惡 性 度 最 高 , 容 易 轉 移 , 對 西方人而言是皮膚疾病中發病率佔第一位的死亡 原因,因黑色素瘤對化學治療及放射線治療的效果不佳,又伴隨著高度 的轉移性;再者,目前對於治療黑色素瘤 (melanoma) 藥物的研究尚未 完善; 若 能 尋找到輔助癌症治療的天然食材,是當前重要的課題。 Wnt/-catenin 訊 息 途 徑 可 調 控 許 多 細 胞 的 進 程 , 包 括 增 生 (proliferation)、分化(differentiation)、存活(survival)、細胞凋亡(apoptosis) 及細胞的移行(motility)。約有30%的黑色素瘤可觀察到Wnt/-catenin訊 息途徑異常活化的現象,因此抑制Wnt/-catenin 訊息途徑也成為治療黑 色素瘤的策略之一。 樟芝(Antrodia camphorata;AC) 為台灣常見的傳統中藥,這種特有 的蕈類生長於牛樟樹上,先前文獻指出樟芝具有抗癌、抗發炎及調節免 疫等功效。在第一部分的實驗利用樟芝發酵液(AC-10),作用在小鼠黑色 素瘤細胞B16F1及B16F10,其IC50皆約為80g/mL。在細胞凋亡分析上, AC-10具有劑量效應誘發細胞凋亡的能力,並伴隨p53的蛋白表現增加, cytochrome c 釋 出 至 細 胞 質 、 BAX/Bcl-2 的 ratio 提 高 以 及 caspase-3 、 caspase-9以及PARP的裂解。在Wnt/-catenin調控路徑之下,利用免疫螢 光染色法發現AC-10可抑制-catenin入核表現,並且降低核質-catenin 的蛋白表現;再同時給予AC-10及MG132(proteasome inhibitor),則會使 -catenin 的 蛋 白 表 現 量 回 復 , 並 利 用 SB216763(GSK3 抑 制 劑 ) 得 知 AC-10降解-catenin為GSK3依賴型(GSK3dependent)。但由RT-PCR 實 驗 得 知 , AC-10 並 不 改 變 -catenin mRNA 表 現 。 而 同 時 給 予 Cycloheximide (CHX, protein synthesis inhibitor)及AC-10可縮短-catenin. III.
(14) 蛋白質降解的半衰期,此現象說明了AC-10造成-catenin的減少原因為 加速蛋白質的降解作用。同時也利用Reporter assay觀察到AC-10可抑制 -catenin調控的轉錄活性,導致下游標的基因如c-myc、cyclin D1、 survivin、MMP-9及VEGF的減少。Flow cytometry分析AC-10會使黑色素 細胞瘤皆停滯在G1期,並連帶的抑制細胞週期蛋白cyclin D1、CDK4; 增加p27、p21的表現。在細胞轉移部分上,細胞刮傷試驗(wound healing assay)以及細胞侵襲試驗(invasion assay)測定發現AC-10可降低癌細胞遷 移的表現,並連帶的使MMP-9、MMP-2及VEGF表現量減少。 在第二部分的實驗中,另以AC-10純化後產物AC-0進行體外試驗觀 察到AC-0也具有抑制-catenin及促進細胞凋亡表現,由RT-PCR實驗得 知,AC-0同樣不會改變-catenin mRNA表現,而同時給予CHX及AC-0 卻無法加速-catenin蛋白質的降解,因此推測AC-0雖無法加速降解,但 令-catenin減少仍為蛋白降解作用。利用SB216763(GSK3抑制劑)得知 AC-0降解-catenin為GSK3非依賴型(GSK3-independent);並利用免疫 沉澱法(IP)來探討GSK3與-catenin之交互作用,同樣也證明了GSK3 的非依賴型。 又以體 內實 驗(裸鼠 腫瘤 模式 )證 實 AC-0確 實可 以抑 制裸 鼠植入 B16F10誘發之腫瘤大小、重量、及體積,並且經免疫組織染色分析、腫 瘤蛋白分析得知,-catenin及其target gene表現也被抑制。 總結本研究得知AC-10及其主成分AC-0可藉由抑制Wnt/-catenin訊 息路徑,而達到抑制腫瘤生長及轉移的效用。. 關鍵字: AC-10;Wnt/-catenin pathway;細胞凋亡;細胞週期;轉移. IV.
(15) Abstract Melanoma is the most serious form of skin cancer. Aberrant activation of Wnt/β-catenin signaling cascade has been observed in approximately one-third of melanomas, indicating that modulation of Wnt/β-catenin activation might be a novel strategy for melanoma treatment. The downstream targets of Wnt/β-catenin signaling pathway including c-Myc, cyclinD1, MMPs, and survivin, which are regulating number of cellular functions, such as proliferation, differentiation, survival, apoptosis and invasion. Antrodia camphorata, a well known medicinal mushroom in Taiwan that has been used as Chinese folk medicine for many years. Previous studies have shown that A. camphorata (AC) possessed greater anti-tumor activity against a variety of tumor cells. However, the anti-tumor efficacy of AC against melanoma was poorly understood. In addition, the currently employing treatment for melanoma is a tough topic, due to the high resistance. to. radio-and. chemotherapy. and. most. the. synthetic. chemotherapeutic drugs are volnarable to non-melanoma skin cells. Therefore, the present study, we aimed to investigate the anti-tumor efficacy of fermented culture broth extracts of A. camphorata (AC-10) and it derived pure compound (AC-0) in murine melanoma cells. The first set of experiment, we observed AC-10 treatment significantly decreased murine melanoma B16F1 and B16F10 cell viability with an IC50 value of 80g/mL. The reduction of cell viability is directly correlated with the inhibition of -catenin and its downstream protein expression. Immunofluorescence analysis confirmed that AC-10-treatment markedly reduced -catenin translocation into the nucleolus, and also downregulates -catenin-mediated. transcriptional. activity.. Furthermore,. MG132. a. proteosomal inhibitor that prevent proteosomal degradation of -catenin,. V.
(16) conversely, GSK3. inhibitor SB216312 also suppressed -catenin. degradation which strongly suggest that -catenin degradation is GSK3dependent.. In. a. similar. way,. AC-10-treatment. attenuate. GSK3expression, suggesting that AC-10 may regulate the proteasomal degradation of -catenin by GSK3-dependentmechanism. Furthermore, flow cytometry analysis showed that AC-10-treatment significantly arrest G1 to S-phase transition followed by the suppression of cyclin D1 (wnt/-catenin target gene), CDK4 expression and increased in p27, p21 levels. Moreover, TUNEL assay revealed that AC-10-treatment induce apoptosis in a dose-dependent manner, followed by the disregulation of BAX/BCL-2 ratio, and the down-regulation of pro-caspase-9, pro-caspase-3 and pro-PARP. Migration and invasion assay shows AC-10 could abate melanoma metastatic ability, through the inhibition of MMP-9, MMP-2, VEGF expression, which are also wnt/-catenin target genes. In the second part, AC-0 also found to effectively inhibit Wnt/-catenin pathway cascades, and suppressed -catenin-mediated transcriptional activity. Similar with AC-10, AC-0 treated cell found to decreased melanoma metastasis and augmented apoptotic induction. MG132 and AC-0 treatment prevents -catenin expression, however, SB216312 failed to prevent GSK3β expression, which suggesting that AC-0-treatment may regulate. the. proteasomal. degradation. of. -catenin. via. GSK3-independentmechanism. This phenomenon also demonstrated with immunoprecipition assay that AC-0 treated cells decreased the interaction between -catenin and GSK3 In vivo study, AC-0 decreased the growth of B16F10-derived tumors development in the athymic nude mice. The decreased B16F10-derived. VI.
(17) tumor growth was associated with a down-regulation of Wnt/-catenin target genes such as c-myc, cyclin D1, MMP-9. AC-0 treatment also induced B16F10-derived tumors apoptosis in nude mice. In conclusion, our data demonstrated that AC-10 and its major compound AC-0 appreciably modulate Wnt/-catenin pathway in melanoma cells. Therefore, we believe AC-10/AC-0 might be a potential chemo-preventive agent for melanoma treatment.. Keywords: Antrodia Camphorata; Wnt/-catenin pathway; melanoma; metastasis; Apoptosis; cell cycle. VII.
(18) 第一章 前言. 1.
(19) 癌症,又稱為惡性腫瘤。惡性腫瘤的發生是由許多外在與內在因素所 造成的,外在因素不外乎為飲食、生活習慣、以及環境等;又內在因素有 性別、年齡、遺傳、種族等誘因,會造成細胞異常生長而影響人體內正常 的恆定。癌症的可怕之處是因癌細胞具有不正常生長、建構新生血管的能 力和癌細胞不老的特性,使得持續進行增生,而導致健康的惡化。 台灣行政院衛生署統計資料顯示,民國九十九年國人的十大死亡原因 之首仍然為惡性腫瘤。其中皮膚癌雖不為國人發生率高的癌症之一,但年 年增加的發生率以及其預後差、化學治療的效果低,仍必須加以注意。而 皮膚癌中最惡性的一種-惡性黑色素瘤,很容易由皮下蔓延至淋巴結,轉移 至全身,包括各個臟器,因此導致高死亡率發生。臨床上而言,在發現惡 性黑色素瘤病灶之後,首次進行手術治療時,大約已有20-40%的患者已發 生淋巴轉移,所以淋巴結是否已受侵犯對未來之預後有相當大之影響力。 影響黑色素瘤分期之最重要的兩個關鍵因素,一是腫瘤的大小,一是侵犯 轉移的程度。在臨床研究上,若能抑制黑色素瘤的大小或轉移能力,就表 示有抗癌之功效。 樟芝(學名 Antrodia camphorata) 為台灣特有的蕈類,又被稱為牛樟 芝、樟芝、牛樟菇或是樟菇,樟芝生長在台灣山區的牛樟樹上,具生理機 能的部位有:子實體、菌絲及孢子。目前已有文獻證實樟芝具有抗腫瘤、 抗氧化、抗發炎等效用,但樟芝對於黑色素瘤的抗腫瘤效用尚未探討完善, 因此本實驗將使用樟芝發酵液(AC-10)以及其主成分(AC-0)來探討對黑色 素瘤之抗腫瘤效用。. 2.
(20) 第二章 文獻探討. 3.
(21) 第一節樟芝及其主成分介紹. 一、樟芝概述 樟芝(學名: Antrodia camphorata;簡稱AC) ,為台灣特有的真菌,僅 生長於牛樟樹(Cinnamomum kanehirae)上的蕈類。牛樟樹是大型的長青 闊葉樹木,只生長於台灣,其分布於海拔200-2000公尺的闊葉森林中 (Chang et al.,1995),分布範圍從桃園角板山、苗栗南庄、南投竹山、高雄 六龜、台東到花蓮,生長季節從六月至十月(Patwardhan et al., 2005)。在台 灣,樟芝亦有別名為牛樟芝、牛樟菇或是樟菇,當地居民也將樟芝稱為蕈 菇界的紅寶石(Wu et al., 2006)。自古以來,樟芝被原住民當作為一種傳統 的處方,來治療飲酒過量或是疲憊不適感。因此,人們相信攝取樟芝具有 保健與延年益壽的功效,可以預防或是治療的疾病包括:肝病、食物或是 藥物中毒、腹瀉、腹痛、高血壓、皮膚癢及腫瘤疾病等(Tsai et al., 1985; (Chen et al., 2007)。 因牛樟樹為台灣稀有且珍貴的樹種,而導致樟芝的高價格與盜採濫 伐。因此目前有替代方式來培養樟芝,分別為子實體或是菌絲體。子實體 具品質好的生物活性,但培養條件較嚴苛且耗時,因此培養菌絲體的方式 較為經濟。菌絲體培養分為固態培養法以及液體發酵法;前者可培養出類 似子實體的紅色菌絲,而後者可得到白色菌絲及紅色菌絲,優點為培養時 間短,缺點為無法取得野生牛樟芝特有且最有療效的三帖類化學物質。目 前針對樟芝的研究仍著重在三萜類與多醣體,目前已知多醣體之研究多集 中在抗腫瘤 (Wasser et al; 1999)、調節免疫 (Wang et al; 1997)、抗泡疹、抗. 4.
(22) 病毒 (Eo et al; 1999)等功能上。三萜類則為抗癌、保護肝臟機能、消除疲 勞以及調解血壓等等。. (B). (A). (C). (D). 圖2-1. 樟芝發酵的各種型態. (Geethangili and Tzeng, 2009). (A) 子實體年齡約12個月。. (B) 子實體年齡約18個月。. (C) 發酵培養AC-10。. (D) 培養後之發酵液。. 5.
(23) 二、樟芝的生物活性 樟芝目前已被探討出具有抗癌、抗發炎、抗B肝病毒複製、抗氧化、保 肝、預防肝臟纖維化、保護神經作用、抗高血壓以及血管舒張的效應。先 前文獻證實樟芝子實體以及菌絲體皆具有抗腫瘤增生的效用,目前已被探 討的癌細胞如:肝癌細胞(Hep3B、J5、J2、G2及SK-Hep-1)、小鼠腫瘤細 胞(Sarcoma 180)、血癌細胞(HL-60) (Hseu et al., 2004)、乳癌細胞(MCF-7) (Yang et al., 2006)、膀胱癌細胞(RT4, TSGH-8301, T24) (Peng et al., 2007)。 在抗發炎表現上,樟芝可以抑制LPS誘發NO、TNF-、IL-1以及PGE2的產 生,並且於NF-kB途徑中誘發iNOS及COX-2的生成(Hseu et al., 2005)。 樟芝也被探討出在生物體內具有抗 B 肝病毒複製的能力,且不具細胞 毒性(Hsiao et al., 2003);加上樟芝成份具有 polyphenol、curde triterpenoids 及多醣體,在保護肝臟效用上具有抗氧化效力(Song and Yen, 2002)。在抗 氧化研究上,樟芝子實體及菌絲體甲醇萃取物均具有清除自由基及螯合金 屬離子能力(Huang et al., 1999);也具有提升肝臟中 glutathione peroxidase 及 SOD 酵素活性(劉; 2002)。在保護神經作用上,樟芝菌絲體萃取液可使 PC-12 細 胞 經 由 PKA-dependent 途 徑 及 抑 制 JNK 、 P38 活 性 來 預 防 apoptosis(Lu et al., 2008)。在抗高血壓效用上,樟芝萃取液中的 Methanol(MS) 萃取具有抗高血壓的效用(Wang et al., 2003)。樟芝在許多生理功能上具有 調控功能,在毒性試驗上,樟芝發酵液的服用並不會造成急性死亡或是畸 胎(江等; 2001);即使在慢性試驗上,乾燥菌絲體對生物體同樣不會造成毒 性(林等; 2000)。. 6.
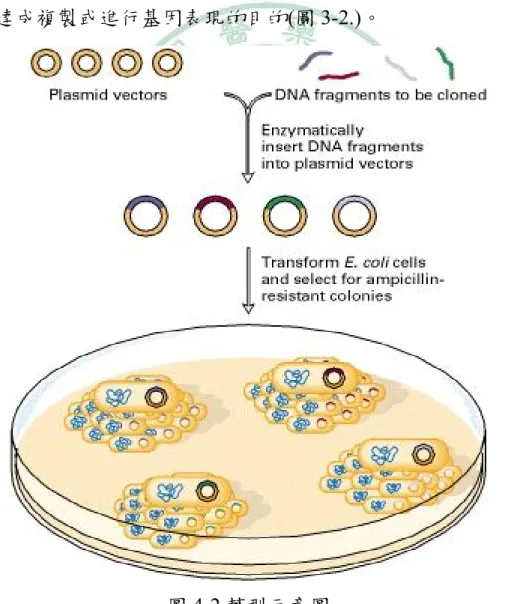
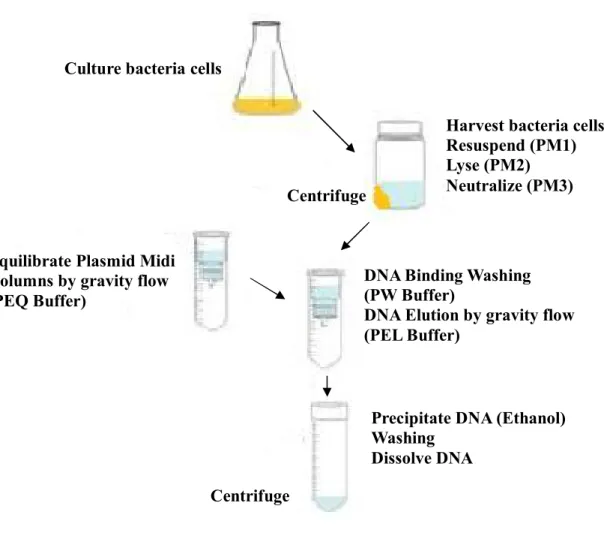
(24) 三、樟芝發酵液(AC-10)之主要成分 利用 HPLC 分析出 AC-10 之各種成分,本實驗第二部分則利用其中之 主要成分(命名為 AC-0),同樣探討惡性黑色素瘤之抗腫瘤效果,且以體內 試驗以及體外試驗進行。. 7.
(25) 第二節. 皮膚癌介紹. 一、皮膚癌背景 皮膚,是人體最分佈最廣的器官,包括表皮層、真皮層、及皮下脂肪 組織。不論任何皮層的各種細胞皆可發生癌病變。皮膚癌好發於裸露部位 如頭、面、頸及手背,也見於口腔粘膜、唇、舌、外陰等部位。長期過度 陽光照射被認為是造成皮膚癌最基本因素,所以大部分的皮膚癌都發生在 臉部、頸部、耳朵、前臂和手背等暴露於陽光的部位。因此,較容易罹患 皮膚癌的人大多為農夫、室外工作者、運動員,或是一些必須暴露於陽光 的工作;以及某些危險因子,如 X 光、焦煤和砷,都比較容易使皮膚受到 UV 照射的損害或是化學因子的傷害,而造成不同的變異,甚至轉變為皮 膚癌。另外,有皮膚癌家族史者以及先天性基因疾病,如色素性乾皮症、 母斑性基底細胞癌症候群患者都有罹患皮膚癌的危機。 皮膚癌好發於西方人,其中澳洲更是全球發生率第一名的國家,因其 陽光日照時間長,其中每十萬人就有三十五位皮膚癌患者。從 1950 年起至 今,黑色素瘤的發生率提升了十倍(Bataille, 2003),同時也使惡性黑色素瘤 成為美國十大癌症之一;再者據美國國家癌症研究所(NCI)近三十年研 究指出,美國每年平均皮膚癌確診的婦女約 62,000 例,死於惡性黑色素瘤 (Malignant Melanoma)的女性超過 8,400 人,死亡平均年齡從 15 歲至 34 歲不等,此研究數據每年仍持續增加。英國皮膚協會(British Skin Foundation) 指出皮膚癌是英國成長最快的疾病,每日約新增 200 起皮膚癌案例。在台 灣,皮膚癌雖無排名在十大癌症之內,但其年年增加的趨勢以及全球氣候. 8.
(26) 異常所帶來的暖冬效應、臭氧層的破壞,使台灣日曬時間逐年增加,而提 高了皮膚癌的發生率,從台灣癌症登記小組的統計資料得知,1979 年男性 皮膚癌粗發生率為 1.71,2007 男性皮膚癌粗發生率則提高至 11.32,漲幅 為 9.61(每十萬人口);1979 年女性皮膚癌粗發生率為 0.99,2007 女性皮膚 癌粗發生率則提高至 9.81,漲幅為 8.88(每十萬人口)(圖 2-2)。在非惡性黑 色素瘤粗發生率上,男性由 1.64 提升至 10.41,漲幅為 8.77(每十萬人口); 女性由 0.91 提升至 9.05,漲幅為 8.14(每十萬人口)。而惡性黑色素瘤粗發 生率,男性由 0.08 提升至 0.91,漲幅為 0.83(每十萬人口),女性由 0.08 提 升至 0.76,漲幅為 0.68(每十萬人口) (圖 2-3、4)。由以上現象得知,即使 非惡性黑色素瘤之發生率漲幅要惡性黑色素留明顯,但因惡性素瘤之高侵 襲性與高轉移性,仍要密切注意且防範。. 9.
(27) 10. 每 十 萬 人 口 年 齡 標 準 化 發 生 率. 9 8 7 6 5 4 3 2 1. 男性. 女性. 0 1979 1980 1981 19821983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000 2001 2002 2003 2004 2005 2006 2007. 年 度 (年). 圖 2-2. 台灣皮膚癌年齡標準化發生率之長期趨勢(依性別),1979-2007 資料來源:台灣癌症登記小組 10. 每 十 萬 人 口 年 齡 標 準 化 發 生 率. 9 8 7 6 5 4 3 2. 男性. 1. 女性. 0 19791980 1981 19821983 19841985 19861987 1988 19891990 19911992 199319941995199619971998 19992000 20012002 20032004 2005 20062007. 年 度 (年). 圖 2-3.台灣非惡性黑色素瘤年齡標準化發生率之長期趨勢(依 性別)1979-2007 年. 資料來源:台灣癌症登記小組. 10.
(28) 0.9. 每 十 萬 人 口 年 齡 標 準 化 發 生 率. 0.8 0.7 0.6 0.5 0.4 0.3 0.2 0.1. 男性. 女性. 0.0 197919801981198219831984 198519861987198819891990 19911992199319941995 199619971998199920002001 200220032004200520062007. 年 度 (年). 圖 2-4.台灣惡性黑色素瘤年齡標準化發生率之長期趨勢(依性別), 1979-2007 年. 資料來源:台灣癌症登記小組. 二、皮膚癌的分類及診斷 皮膚癌的表現是多變化的,皮膚癌如果在初期被發現是容易治癒的,但往 往因為其病灶不痛不癢而使人忽視它的存在,使得死亡率增加。皮膚癌的 種類繁多,最常見的三種如下: 1.基 底 細 胞 癌 基底細胞癌是台灣地區最常見的皮膚癌,大約佔所有皮膚癌的 45~50%。 基底細胞癌大多發生在曝曬部位如臉部,手背及前臂,與長期紫外線的 傷害有關。其病灶看起來表皮周圍光亮凸出、呈現膚色伴隨有微血管擴. 11.
(29) 張的小腫塊,中間偶爾有潰瘍傷口。雖然其惡性度是皮膚癌中最小且轉 移機率也較低的,但可能因為忽視其重要性而侵犯局部神經或造成局部 組織的破壞。 2.鱗 狀 細 胞 癌 為第二常見的皮膚惡性腫瘤。可發生於與長期紫外線傷害的曝曬部位, 也會發生於慢性的潰瘍傷口或是工作上常須接觸到化學物質的人,有時 會發生在腿或腹壁等受到慢性傷害或輻射的部位。鱗狀細胞癌通常會發 展成大而蕈樣狀的腫塊,而且可能經由淋巴腺散播出去,造成淋巴結腫 大,更可能轉移到其他內臟,如:肺部、肝臟、大腦或骨骼。 3.黑色素細胞癌 為第三常見的皮膚惡性腫瘤,所有皮膚癌中發生率小於 5%,但惡性度最 高,容易轉移,致死率為三種皮膚癌中最高,達到 80% (Tsao et al., 2004)。 黑色素細胞癌在亞洲人身上較常出現在肢端,如手指腳趾、手掌腳掌、 趾甲等。臨床上,可能以黑色或色澤不均的斑塊出現,也可能呈現凸起 的腫塊,必須和良性色素性細胞痣或母斑加以辨別。黑色素細胞癌恐怖 的地方在於其可以轉移到任何器官,除了可以轉移到他處的皮膚、皮下 組織以及淋巴節外,最常轉移的器官是肺臟、肝臟、腦部、骨骼、腸道, 而導致預後不佳。黑色素瘤又可分成四型:(1)結節狀黑色素瘤(Nodular melanoma):病灶呈現半球狀隆起,大約佔病例的 15-20%。生長迅速, 是侵襲性較高的一種,也是較常見的一種。(2)表面擴散型黑色素瘤 (Superficial spreading melanoma ):佔 2/3 的病例,進程緩慢,往往要好. 12.
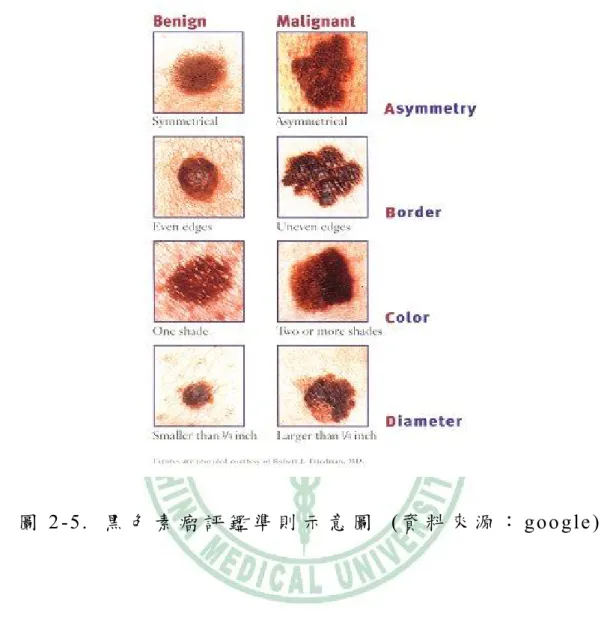
(30) 幾年,外型有如一塊不規則的扁平黑痣,好發在上背部。 (3)小痣樣黑 色素瘤(Acral lentigous melanoma):好發在慢性日曬部分,外觀上像是形 狀奇特不規則的黑斑。 (4) 小痣黑色素瘤(Lentigo maligna melanoma): 最少見的一種,通常在鼻子或是臉頰發生,常見於老人。這是指任何發 在足底、手掌及指甲周圍的黑色素瘤。一般惡性黑色素瘤表現不規則的 黑色斑塊或凸出,但有些則呈現紅色的凸出腫塊,易被錯認為慢性發炎。. 目 前 皮 膚 癌 的 診 斷 方 式 仍 以 目 前 皮 膚 切 片 為 主,但 平 時 可 依 據 以 下 準 則 ( A BC D E ) 來 區 分 良 性 的 痣 或 惡 性 的 黑 色 素 細 胞癌,做定期自我檢查: A ( A s y m me tr y ) : 痣 的 外 觀 是 否 出 現 不 對 稱 性 B( Border) : 邊 緣 是 否 呈 現 不 規 則 或 模 糊 不 清 的 現 象 C ( C o lo r ) : 色 調 上 是 否 呈 現 不 均 勻 、 深 淺 不 一 D ( D ia me t e r ) : 大 小 是 否 超 過 六 公 釐 以 上 或 變 大 的 情 形 E ( E le v a t i o n ) : 表 面 是 否 有 不 規 則 的 隆 起 現 象. 13.
(31) 圖 2 -5 . 黑 色 素 瘤 評 鑑 準 則 示 意 圖 ( 資 料 來 源 : g o o g le ). 三、皮膚癌的分期 1.基底細胞癌及鱗狀細胞癌的分期 第 1 期 腫瘤直徑小於 2 公分 第 2 期 腫瘤直徑大於 2 公分 第 3 期 腫瘤已經侵犯軟骨、肌肉、骨骼或附近的淋巴結 第 4 期 腫瘤已經轉移到其他的器官. 14.
(32) 2.黑色素細胞癌的分期須考量侵犯的深度,比較複雜,簡化如下: 第 0 期 惡性細胞侷限在表皮,沒有侵犯真皮層 第 1 期 腫瘤厚度小於 1 公厘伴有表皮潰瘍或腫瘤厚度 1 公厘到 2 公 厘,沒有表皮潰瘍 第 2 期 腫瘤厚度 1 公厘到 2 公厘,伴有表皮潰瘍,沒有轉移到淋巴 結或腫瘤厚度超過 2 公厘,沒有表皮潰瘍,無轉移到淋巴結 第 3 期 腫瘤已經侵犯鄰近的組織或鄰近的淋巴結 第 4 期 腫瘤已經轉移到其他的器官或遠處淋巴結. 四、皮膚癌的治療與預後 對於基底和鱗狀細胞癌來說,早期發現的治癒率很高;而黑色素瘤可 以快速的擴散到身體其他部位,致死率也高,加上其外觀常被誤認為痣, 而使得發現更晚,導致預後狀況更差。皮膚癌侵犯的部位深度和惡性度都 決定不同的治療方式。目前治療皮膚癌的方式包括外科手術切除、放射療 法、電燒刮除、冷凍療法、局部化學療法、光動力治療、生物治療(免疫治 療)等等;但因黑色素瘤對於化學治療效果具有高度的阻抗性,因此研究具 抗氧化特性的營養素來預防皮膚癌也是具潛力的發展方向之一(Katiyar and Mukhtar, 1997),目前較為研究探討之營養素抑制皮膚癌與其功能性分別條 列如下:. 15.
(33) (1) 纖維素(Fibrate):在惡性黑色素瘤中扮演促進細胞凋亡角色,並具有使 細胞週期停滯之抗增生能力(Mossner et al., 2002; Placha et al., 2003)。 (2) 沒食子酸酯化兒茶素(EGCG):具有 photo-protection(Einspahr et al., 2002; Elmets et al., 2001)、抗氧化(Hsu, 2005)、促進小鼠高轉移性黑色素瘤 B16F10 細胞凋亡(Malafa et al., 2002)、使兩株高轉移性黑色素瘤 A375 及 Hs-294T 細胞週期停滯之抗增生能力(Nihal et al., 2005),以及抗血管新生效用 (Tosetti et al., 2002)。 (3) 維生素 E(Vitamin E):具有 photo-protection(Anstey, 2002; Gensler et al., 1996; McVean and Liebler, 1997)、抗氧化(Keller and Fenske, 1998)、促進細 胞凋亡、使細胞週期停滯之抗增生能力(Malafa et al., 2002)。 (4)胡蘿蔔素(-carotene):具有 photo-protection 及抗氧化功能(Wertz et al., 2005)。 (5) 茄紅素(Lycopene):具有 photo-protection(Fazekas et al., 2003)及抗氧化 功能(Hwang and Bowen, 2002)。 (6) 類黃酮素(Flavonoids):具有 photo-protection(Wei et al., 2003)、抗氧化、 促進細胞凋亡、使細胞週期停滯之抗增生能力,以及免疫調節作用(Singh and Agarwal, 2002)。 (7) 前花青素(Proanthocyanidins):抗氧化以及抗發炎作用(Li et al., 2001; Singh and Agarwal, 2002)。. 16.
(34) (8)白藜蘆醇( Resveratrol):抗氧化、促進細胞凋亡、使細胞週期停滯之抗 增生能力,以及抗血管新生效用(Aggarwal et al., 2004)。 (9) 硒(Selenium):具有抗氧化(Rotruck et al., 1973)、促進細胞凋亡、使細 胞週期停滯之抗增生能力、抗血管新生、免疫調節以及 DNA 修補作用 (Rotruck et al., 1973)。 (10) 維生素 D(Vitamin D):促進細胞凋亡、使細胞週期停滯之抗增生能力 以及抗血管新生作用(Vandewalle et al., 1995)。 (11) 視 網 素 (Retinoids) : 促 進 細 胞 凋 亡 及 抗 血 管 新 生 作 用 (Zhang and Rosdahl, 2004)。 (12) 人參(Ginseng):使高轉移性小鼠黑色素瘤 B16-BL6 細胞週期停滯之抗 增生能力(Xiaoguang et al., 1998)以及抗血管新生作用(Mochizuki et al., 1995)。. 17.
(35) 第三節. WNT/-catenin pathway 介紹. 一 、 W n t / - c a te n i n p a t h w a y Wnt/-catenin signaling pathway 可調控許多細胞的進程,包括增生 (proliferation)、分化(differentiation)、存活(survival)、細胞凋亡(apoptosis) 及細胞的死亡(motility)。約有 30%的黑色素瘤(melanoma)可觀察到 Wnt/-catenin signaling pathway 異常活化的現象(Tarapore et al., 2010), 目前較為被探討的 wnt pathway 有以下三種,其中 Canonical Wnt pathway 也是目前研究中較為被深入探討的一條訊息傳遞路徑 。 (1) Canonical Wnt/-catenin pathway: 在 Wnt 這個 ligand 和在細胞膜上的 receptor,如 Frizzled/LRP 做結合, 使細胞質中 Dishevelled 持續性磷酸化,而使 GSK3磷酸化而失去活 性。在此時,GSK3、Axin、APC 及-catenin 的 destruction complex 無法聚合產生作用而降解-catenin;細胞質中-catenin 增加,而後進到 細胞核,和 Tcf/LEF 做結合參與轉錄作用,因而啟動下游特定的轉錄 因子,而這些轉錄因子會影響增生、轉移與血管新生,如: cyclin D1、 c-myc 、 MMP-9 、 VEGF,進而造 成腫 瘤 增生或 是轉移到 其他器 官 (Behrens et al., 1996) 。 反之,當 Wnt 訊息傳遞不活化時,GSK3、 Axin、APC 及-catenin 會組合成 destruction complex,導致-catenin 磷 酸化,進而被 TrCP E3 ubiquitin 標記而泛素化(ubiquitinylation),再 經由 26S proteasome 走向 proteasomal degradation (Hart et al.,1999) 。. 18.
(36) (2) Wnt/planar cell polarity pathway(Wnt/PCP pathway): 此路徑為 Non-canonical Wnt pathway 的一種,當 Wnt 結合到 Fz receptor,Dishevelled 透過 Damm1 活化影響下游的訊息傳遞包含 Rac、Rho 以及 JNK 等訊息傳遞,主要是影響細胞的移動(Polakis, 2000 )。 (3)Wnt-Ca2+ pathway: 此路徑也為 Non-canonical Wnt pathway 的一種,當 Wnt (例如 Wnt-5a) 結合至其 receptor(例如 Fz-2),活化 G-protein 後導致鈣離子的釋放。 因此,某些對鈣離子敏感的酵素如 PKC 便會活化而啟動下游反應。 (Kuhl et al., 2000). 圖 2-6. Canonical Wnt/-catenin pathway (資料來源:Roberts et al., 2007). 19.
(37) 二、 調控 Wnt pathway 之相關蛋白 (1)Wnt: Wnt gene為分泌型醣蛋白(Secreted glycoproteins), 家族目前有19 個成員,做為ligand來傳遞訊息調控,可以調控細胞分化、細胞增 生及細胞存活(Chien and Moon, 2007)。Wnt有很多不同的異構物, 目 前 在 Canonical Wnt/-catenin pathway 中 , 有 參 與 的 ligand 如 Wnt-1、Wnt-3a、Wnt7以及Wnt-8 (Delmas, 2006),而Wnt-4、Wnt-5a、 Wnt-5b 及 Wnt-11 會 活 化 non- canonical pathway(O'Connell and Weeraratna, 2009)。 以下介紹目前較被探討的ligand: a.Wnt-1:參與許多癌症的進程,調控細胞增生、移行及胚胎發育 等(Rijsewijk et al., 1987) 。 b.Wnt-2A:參與血管新生,在大腸直腸癌中有特別表現,在黑色素 瘤中也有高度表現。(Kashani-Sabet et al., 2009) c.Wnt-3A:增加Dvl的磷酸化,穩定-catenin及增加TCF4轉錄活性。 d.Wnt-5A:參與Non-canonical Wnt pathway,可在許多高侵襲性的黑 色 素 瘤 中 發 現 , 但 通 常 不 表 現 於 初 始 的 病 灶 (O’Connell and Weeraratna, 2009),主要調控黑色素瘤的細胞增生與侵襲(Bittner et al., 2000; O’Connell and Weeraratna, 2009; Weeraratna et al., 2002)。 e.Wnt-8A:活化canonical pathway 中的正調控因子-casein Kinase 1 (CK1) (Swiatek et al., 2004) 。. 20.
(38) (2) Frizzled/LRP Frizzled(Fz)以及low density lipoprotein receptor-related proteins 5 and 6(LRP5/6)為兩個具穿膜(trans-membrane receptors)功能的接受 器(Bhanot et al., 1996; Wehrli et al., 2000)。Frizzled接受器可被 -catenin 依 賴 型 路 徑 (canonical) 以 及 -catenin 非 依 賴 型 路 徑 (noncanonical ;如 planar cell polarity) 活化 ,因 此 LRP5/6 因 只 在 Canonical Wnt/-catenin pathway上表現,所以較具有特異性(He et al., 2004a)。. (3)Dishevelled(Dvl) 在 Wnt/-catenin pathway 中是扮演著正調控的角色,由細胞膜上的 接受器 接受 訊息 , 再傳 遞至 下游 的 -catenin (Noordermeer et al. 1994),而過度表現或是持續性活化的 Dvl 會提高癌化的能力。Dvl 在脊椎動物中會有表現,家族中有三位成員(Dvl-1、Dvl-2、Dvl-3), 分別位於染色體的 1p36,17p13.2 及 3q27 (Bui et al., 1997)。先前文獻 發現 Dvl-1 及 Dvl-3 在非小球性肺癌 (nonsmall cell lung cancer) 中有 異常表現,也會影響肺癌細胞的轉移與侵襲。(Wei et al., 2008) (4) Glycogen synthase kinase-3 (GSK3) GSK3 為一富含 serine 及 threonine 之 protein kinase;GSK3在 Wnt/-catenin pathway 中扮演負調控角色,作用為使-catenin 磷酸. 21.
(39) 化而導致降解;然而 GSK3也參與了代謝、結構蛋白以及轉錄因子 的調控(Frame and Cohen, 2001)。 (5)Axin Axin 存在時,-catenin 對 GSK3來說是一個更好的受質(Kishida et al., 1998) , Axin 的過度表現會使得-catenin 失去活性。因 Axin 的作用如 scaffold protein,帶領其他蛋白如 GSK3、APC 等為一複 合體,而降解 -catenin(Behrens et al., 1998; Zeng et al., 1997) 。因 此 Axin、GSK3以及 APC 皆為 Wnt signaling pathway 的負調控因 子。 (6) Adenomatous polyposis coli (APC) APC 在許多癌症中扮演抑癌者的角色,而其突變性與-catenin 的 調控有絕對關係(Polakis et al., 1999);在體外試驗的細胞培養中, Axin 與 APC 接合 位點的 存在對 於 調控 -catenin 是 很 重要 的 (Kawahara et al., 2000)。大約有 90%的大腸癌表現特異性的 Wnt 訊 息傳遞,這大約有 80%的原因是來自於 APC 的突變(Korinek et al., 1997; Miyaki et al., 1994; Powell et al., 1992)。然而,-catenin 的突 變在肝癌、卵巢癌、攝護腺癌、皮膚癌、黑色素瘤似乎比 APC 的 突變要來的普遍,也是造成 Wnt 訊息傳遞異常調控的主因(Polakis, 2000)。. 22.
(40) (7) -catenin -catenin 具有三種功能,第一為在細胞膜上做為細胞黏合功能,可 做為細胞骨架的一個角色;第二為在細胞核內扮演基因的轉錄因 子;第三為在細胞質內做為一個儲備功能,可做為黏合或是轉錄角 色的儲備,最重要的為降解作用。-catenin 的 N 端有被磷酸化位 點,可被 GSK3磷酸化,而大部分的-catenin 突變也是因為磷酸 化位點異常而無法被降解,所以持續活化(Huber et al., 1997)。LRP6 的磷酸化可直接的抑制 GSK3,導致 LRP6 持續性的與-catenin 的降解複合體反應,使得-catenin 的磷酸化被抑制(Cselenyi et al., 2008; Wu et al., 2009)。-catenin 的突變以及其在核內表現的提升皆 為 Wnt 訊息傳遞活化的標的(Haydon et al., 2002);-catenin 轉錄至 核內後與 T-cell factor 4 (Tcf4)結合,並啟動下游 Wnt 特異性的轉錄 因子,包括 cyclin D1、c-Myc、survivin 等,而這些轉錄因子與癌 症的進程皆有正相關(Ito et al., 2000; Liu et al., 2004; Masaki et al., 2003)。. 三、蛋白酶體降解作用(Proteasome degradation) 在細胞質中,無作用的蛋白質會經由泛素蛋白酶體系统 (ubiquitin-proteasome system;UPS) 被降解(Nandi et al., 2006)。過程 中,泛素(ubiquitin) 會需要3個酵素催化,分別為E1、E2、E3而連接到. 23.
(41) 目標蛋白lysine 48,形成polyubiquitin,使目標蛋白得以被28S蛋白酶體 (proteosome) 所 辨 認 , 進 而 將 目 標 蛋 白 分 解 成 小 的 多 胜 肽 (small peptides) 和胺基酸(Hegde, 2004)。E1(又稱為ubiquitinactivating enzyme) 透 過 ATP 與 ubiquitin 結 合 後 , 會 活 化 ubiquitin ; E2( 又 稱 為 ubiquitin conjugating enzyme) 則會被吸引過來與E1相互交換;接著再帶著活化 的ubiquitin和已經與目標蛋白結合的E3 (又稱ubiquitin ligases) 連接在 一起;最後,E3促進E2將ubiquitin 轉換到目標蛋白上。而由以上的步 驟不斷的循環,便形成了目標蛋白的多次泛素化(Polyubiquitination,最 後則被proteosome辨認並分解(Sun, 2006)。人體內約有將近1000種 E3 ligases,β-catenin 的E3 ligases則為βTrCP(Sun, 2006)。. 圖2-7. 泛素化降解作用示意圖 (資料來源:MIT OCW. After W. Hilt, University Matstuttgart). 24.
(42) 四、 Wnt/-catenin pathway in melanoma 惡性黑色素瘤為皮膚癌中高度侵襲與轉移的癌症 ,持續性活化的 Wnt/-catenin signaling pathway 在黑色素瘤中是常發現的異常訊息傳遞 (Reifenberger et al., 2002),其中Wnt-1及Wnt-3A在mouse model中是促進 mammary tumor的因子,會經由-catenin來提升黑色素細胞的分化和腫瘤發 育,而黑色素瘤的細胞質與細胞核中經常有-catenin的表現(Omholt et al., 2001)。 目前文獻指出,Wnt pathway中某些調控蛋白如:APC及-catenin的突 變,造成訊息傳遞持續作用,使得腫瘤持續性的生長 (Rubinfeld et al., 1997);即使許多癌症訊息傳遞中會發現-catenin的突變,但在黑色素瘤中 是較罕見的,大約只有1/65的機率(Rimm et al., 1999);APC的突變在黑色素 瘤中也是罕見的(Worm et al., 2004),因此較不考慮黑色素瘤中突變與Wnt pathway的關係(圖2-7.)。 Wnt-8對於黑色素細胞的分化尚未被深入探討,但其他的 ligand 如 Wnt-2、4、5A、7b、10b則有參與調控。值得注意的是,Wnt-5A及Wnt-2 在目前已知的研究中發現和黑色素瘤有關,Wnt-5A可提高50%黑色素瘤細 胞存活率(Iozzo et al., 1995),也可提高黑色素瘤的細胞移動性及侵襲能力 (Weeraratna et al., 2002)。在高侵襲性腫瘤上也發現高含量的Wnt-5A表現 (Bittner et al., 2000)。因此,本實驗欲探討Canonical Wnt/-catenin signaling pathway中,樟芝發酵液對於訊息傳遞中蛋白的調控影響。. 25.
(43) 圖 2-8.. Wnt/-catenin pathway 的突變. (資料來源:(Barker and Clevers, 2006). 五、Wnt pathway 的藥物發展 從 1990 年開始,Wnt pathway 的異常活化首先由大腸癌和黑色素瘤開 始被關注,因此發展 Wnt pathway 的抑制物在藥物及化學的部分被廣泛注 意(Kinzler et al., 1991; Rubinfeld et al., 1997)。目前有許多藥物已經上市,包 括 non-steroidal anti-inflammatory drugs (NSAIDS)(如 aspirin、sulindac 以及 indomethacin)以及 vitamin 衍生物(如 vitamin A 及 vitamin D)等,這些藥物 可能具有直接或間接抑制 Wnt pathway 的效用。. 26.
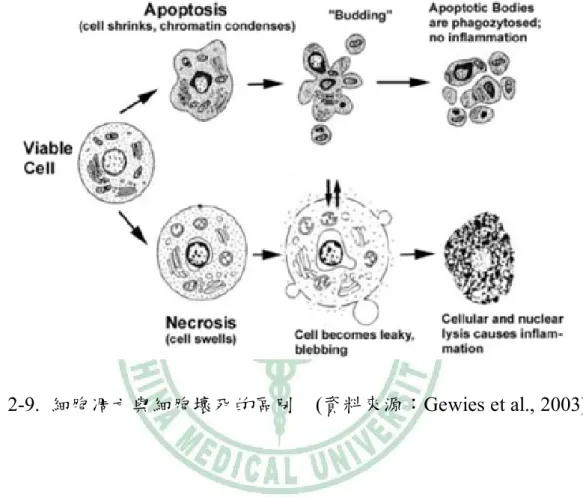
(44) 第四節. Wnt/-catenin pathway與細胞凋亡之相關性. 一、 細胞的死亡(cell death): 細胞的死亡可分為細胞壞死(necrosis)或是細胞凋亡(apoptosis)。細胞壞 死的發生為細胞死亡的一種,造成的原因有感染、毒素或是創傷。與細胞 凋亡相較之下,這是一種非計畫性的死亡方式。細胞壞死通常不會參與訊 息調控,而且細胞壞死後會分泌細胞激素等,會對其他細胞造成影響,也 可能造成其他細胞的傷害。細胞凋亡可以調控細胞的死亡,並且在平衡細 胞恆定上扮演重要角色(Cotter, 2009),通常為細胞受到環境刺激後,引起 訊息傳遞機制來控制其死亡。在正常細胞也可觀察到這種狀況,其中可能 因素為基因突變,為了避免突變的遺傳,而啟動細胞凋亡的機制。細胞凋 亡的發生範圍主要是在單一細胞中,可分為早期(early stage)以及晚期(later stage),在 細胞凋 亡早期 時 ,會出 現染色 質濃縮 (compaction of nuclear chromatin)、細胞質濃縮(condensation of cytoplasm)以及細胞皺縮(shrunken) 等現象;晚期現象則為細胞核裂解並形成凋亡小體(apoptotic bodies)。和細 胞壞死最不同的現象為,細胞凋亡過程中不會危害到周邊組織而引起發炎 反應(圖2-9.)。. 27.
(45) 圖 2-9. 細胞凋亡與細胞壞死的區別. (資料來源:Gewies et al., 2003). 二、調控細胞凋亡之路徑 細胞凋亡的調控主要有三大路徑,分別為粒線體傳遞路徑 (Mitochondrial pathway)、死亡受體路徑(Death receptor pathway)以及內質網 路徑(Endoplasmic reticulum pathway) (圖 2-8.),以下將特別介紹粒線體路徑 及其重要蛋白:. 28.
(46) I. 粒線體傳遞路徑(Mitochondrial pathway). 1.訊息路徑 此路徑又稱為內在路徑(intrinsic pathway),可被許多外在的壓 力刺激造成,包括 UV 照射、加碼射線、熱或是病毒感染等,大部 分為 DNA damage。當細胞受到外來的刺激後,會活化 BAD 以及 BIM 蛋白,而這兩個蛋白主要功能為抑制 Bcl-2 以及活化 BAX, 造成 BAX/Bcl-2 的比值改變,使粒線體膜電位下降,並且釋放 cytochrome c 及 凋 亡 因 子 Apaf-1 (apoptosis protease-activating factor-1)等。而後 Cytochrome 、APaf-1 與 Procaspase-9 蛋白結合而 形成凋亡複合體(apoptosome)後,會促使 procaspase-9 形成活化態 的 caspase-9,接著開始活化下游的 caspase-3,最後造成 PARP (Li et al., 1997)分解斷裂,使細胞走向凋亡。. 2.重要參與蛋白 (1) BCL-2: BCL-2 家族扮演的角色為「細胞凋亡開關」,調控的對象為粒 線體膜(Giam et al., 2008)。BCL-2 家族成員皆扮演前凋亡或是抗凋 亡的角色,緣由為其 domain 的不同性,BCL-2 家族具有 4 個 BCL-2homology domains,分別為 BH1、BH2、BH3 以及 BH4。抗 凋亡蛋白包括 BCL-2、BCL-XL、MCL1、BCL-W 等,通常含有 3-4 個 BCL-2 homology domains(Adams and Cory, 2007);前凋亡蛋白則. 29.
(47) 包括 BAX、BAK 以及 BCL-XS 等(Reed, 1997),通常也含有 3 個 BH domain。正常情況下,細胞的狀態是由此兩類蛋白的比率來維 持,當 Bax 表現量高於 BCL-2 時,細胞就會走入凋亡;換言之, BCL-2 表現量大於 Bax 時,則會抑制細胞凋亡的情況發生。 (2) Cytochrome c Cytochrome c 又稱為細胞色素 c,是呼吸鏈的傳遞成員之一。 在細胞凋亡過程中,可被 BCL-2 蛋白所調控而開啟滲透性轉移孔 洞(permeabilization transition pore;PTP)後,造成粒線體內膜滲透性 (mitochondrial membrane permeabilization)不穩定,使得粒線體膜電 位下降。在此過程中 Cytochrome c 會釋放到細胞質中,而活化 caspase 3(CPP32) (Zou et al., 1997)。 (3) Caspases家族(蛋白水解酶) 細胞凋亡的起始為由一個蛋白水解酶家族執行切割的功能(Li and Yuan, 2008)。起始者(apoptotic initiators)功能主要在於傳遞細胞 凋亡訊號,成員有 caspase-2、caspase-8、caspase-9 以及 caspase-10; 而執行者(effector caspases)在接收細胞凋亡的訊息傳遞後,執行下 游 蛋 白 水 解 作 用 , 裂 解 下 游 的 蛋 白 質 , 如 poly(ADP-ribose) polymerase(PARP)、 DNA-dependent protein kinase (DNA-PK)等, 而引發細胞凋亡發生,成員有 caspase-3、caspase -6、caspase-7。其 中 PARP 的功能是修復受損的 DNA、調控細胞增殖與死亡的平衡, 和維持基因體的穩定性,當 caspase-3 調控 PARP 裂解時會使 PARP 失去原本的功能(Bellosillo et al., 1997). 30.
(48) II. 死亡受體路徑(Death receptor pathway) 死亡受體路徑又可稱為外在路徑(extrinsic pathway),可被細胞 膜外的死亡受體接受器接收到訊號而活化,例如 TNF 家族(Gallenne et al., 2009; Mahalingam et al., 2009),. 目前已有文獻探討死亡受. 體包括 TNF- 、Fas-L、TNF-related apoptosis inducing ligand 等(Jin and El-Deiry, 2005)。當 Death ligand 與 Death receptor 結合後,會促 使 下 游 的 受 體 傳 導 蛋 白 質 FADD(Fas-associated death domain protein)與 procaspase-8/ procaspase-10 結合而成一 Death Inducing Signaling Complex(DISC),而使 Procaspase-8/10 活化為 caspase-8/ 10(Jin and El-Deiry, 2005)。活化後的 caspase-8/10 可使 pro-casepase3 活化為 caspase-3,而後走向細胞凋亡(Kluck et al., 1997)。此外,死 亡受器傳遞路徑同時也會藉由 Bid 活化 Bcl-2 家族蛋白,進一步促 使 Cytochrome c 的釋放(Eskes et al., 2000)。. III.內質網路徑(Endoplasmic reticulum pathway) 在內質網路徑中發現 caspase-12 專一性地存在於內質網中。當 內質 網受到壓力刺激會造成鈣離子釋出,進而活化 caspase-12,最 後形成細胞凋亡(Nakagawa et al., 2000)。. 31.
(49) 圖2-10. 細胞凋亡的三種路徑. 資料來源:(Ghavami et al., 2009). 32.
(50) VI.其他參與細胞凋亡的重要調控因子. (1) p53 基因調節 p53為一抑癌基因,約在50%的癌症表現上可發現到p53的功能失常 (Oren, 1999),p53會感受到DNA demage並且直接使細胞生長停滯或是 走向細胞凋亡(Vogelstein and Kinzler, 2004);再者,p53的活化不只具 有抑癌基因的功能,其影響層面包含細胞週期、細胞凋亡、細胞分化、 DNA修復以及血管新生(Janus et al., 1999; Sionov and Haupt, 1999)。當 細胞受到外來的壓力時,p53活化p21表現,會參與細胞週期G1期的調 控;而活化GADD45會使G2期停滯;BAX、Fas則會導致caspase的活化 及誘發細胞凋亡(Burns and El-Deiry, 1999; el-Deiry, 1998)。p53蛋白質主 要藉由磷酸化及乙醯化等轉譯後修飾作用來增加p53蛋白質的穩定性 (Susin et al., 1998)。許多逆境因子會誘發p53蛋白質活化,包括核酸缺 失、微小管瓦解、缺氧、一氧化氮、紫外線及化學物質等(Susin et al., 1998)。. (2) IAP家族 IAP 家族為 The inhibitor of apoptosis (IAP) family 的簡稱,為抗細胞凋 亡的家族,也是調控 caspase 能力的重要調控者(Deveraux et al., 1997), 家族成員包含了 cIAPs(cIAP1 及 cIAP2)、NIAP、XIAP、Livin/ML-IAP 以及 survivin 等成員(Tamm et al., 2000)。. 33.
(51) 四、Wnt /-catenin pathway 及細胞凋亡的相關性 Wnt/-catenin pathway所調控的轉錄因子包含了Survivin(Wei et al., 2010)。Survivin為inhibitor of apoptosis (IAP) family中的一員,不僅可調 控細胞週期的有絲分裂期,也可做為抗凋亡因子的角色(Altieri, 2008)。 WT-Survivin 分 享 相 同 的 N 端 , 其 C 端 的 不 同 決 定 了 不 同 的 survivin isotype,其中 Survivin Ex3為抑制粒線體依賴型的細胞凋亡(Fangusaro et al., 2005);Survivin-2可能減弱抗細胞凋亡的能力;Survivin-2B扮演 survivin的對立者(Mahotka et al., 1999)。粒線體中的survivin可以迅速的 持續釋放至細胞質中,以防止細胞凋亡的發生,並且藉由參與細胞凋 亡路徑中蛋白之間的交互作用支持著細胞存活的狀態 (Dohi et al., 2004) 。 p53 在 前 文討 論 過其 扮 演 的角 色 為 抗凋 亡 因 子, 而 p53 及 -catenin pathways 之間也已被探討具有相關性,尤其是在腫瘤新生作 用及DNA demage時,發現穩定的-catenin可藉由抑制p53 降解而導致 p53的穩定(Damalas et al., 2001),用以平衡細胞間的訊息傳遞。在黑色 素以及上皮的皮膚癌中,survivin的表現和腫瘤的侵襲、病人的存活率 降低有關 (Chen et al., 2009; Lo Muzio et al., 2001; Piras et al., 2007)。有 文獻指出,黑色素型的增生病灶表現出高度的survivin和 melanoma的 起始有關(Chiodino et al., 1999; Florell et al., 2005; Ibrahim and Brown, 2008),因此,在melanoma調控中,抑制Wnt調控之survivin對細胞凋亡 的調控是很重要的重點之一。. 34.
(52) 第五節 Wnt/-catenin pathway與細胞週期之相關性 一、何謂細胞週期(cell cycles) 人體是由多個細胞組成,各種細胞皆有其生命期,細胞具有分裂的功 能,也是生物體存活的重要特徵之一。 細胞週期是一個過程會調控細胞的 生長與增生,也牽涉到 DNA 的修復、組織充血性傷害以及疾病,例如: 癌症等(Schafer, 1998)。許多調控細胞週期的蛋白也參與了此過程,例如 cyclin-dependent kinases(CDKs)以及 cyclin 蛋白,會調控細胞的四個進程: G0/G1 phase (Gap 1 phase)、S phase (DNA synthetic phase)、G2 phase (Gap 2 phase)、M phase (Mitotic phase) (Schafer, 1998),使細胞從一次有絲分裂結 束後生長,再到下一次分裂完成的循環過程。G1 期以及 G2 期代表細胞週 期的一個「gap」 ,分別發生在 DNA 合成以及有絲分裂之前。在 G1 期,細 胞準備 DNA 的複製;在 S 期,細胞合成 DNA,由 2N 變成 4N;G2 期為 第二個分裂點,細胞準備有絲分裂;最後一個階段為 M 期,為細胞進行有 絲分裂的時期,由 4N 變成 2N,而得到兩個子細胞。在此還有一個時期為 G0 期,為靜止狀態的休眠期,G0 本指細胞並無在細胞週期的進程內,但 仍具有分裂的潛力,例如:肝細胞。G0 期也常發生在缺少養份而導致生長 抑制的狀況,此時細胞週期也會停滯(Schafer, 1998)。 細胞週期所需要的時間反映了細胞的狀態,癌變的細胞常常有異常的 分裂週期,導致細胞生長較正常細胞快速,因而增加了癌症惡化的情形, 因此抑制癌細胞的細胞週期也為一種抗腫瘤的方法。. 35.
(53) 圖 2-11. 細胞週期分佈圖. (資料來源:The Science Creative Quarterly). 二、細胞週期的調控(Cell Cycle Regulation) 細胞週期可由兩種重要蛋白所調控,Cyclin 家族與 Cyclin-dependent kinases(CDK)家族。CDK 為絲胺酸/酥胺酸蛋白激酶家族,家族中各成員會 在不同的細胞週期被特異性的 cyclin 蛋白質所調控,而調控細胞週期進行 會由 CDK 的活化與否來確定(van den Heuvel and Harlow, 1993)。目前在哺 乳動物中至少有七種 CDKs(Pines, 1995),分別為:CDK4、CDK6、CDK2(G1 期活化),CDK2(S 期活化),CDK2/CDK1(G2 期與 M 期活化),CDK7(各 細胞週期)。 CDK 的活性可由 CDK inhibitor 所抑制,而 CDK inhibitor 又可分為兩個 家族, INK4 家族以及 KIP/CIP 家族(Sherr and Roberts, 1995)(表 2-5)。INK4 家族包括 p15 (INK4B)、p16 (INK4A)、p18(INK4C)以及 p19(INK4D),作用. 36.
(54) 為使 G1 期的 CDK 失活,例如:CDK4、CDK6。這些 CDKI 功能為在 CDK 與 cyclin 結合前,和 CDK 組成一個穩定複合體,以防止 cyclin D 的作用 (Carnero and Hannon, 1998)。 KIP/CIP 家族包括了 p21(CIP1/WAF1),p27(CIP2)以及 p57(KIP2),作用 為使 CDK/cyclin 複合體失去活性(Lee et al., 1995),也會使 cyclin B-CDK1 的複合體失活(Hengst and Reed, 1998)。 Cyclins 蛋白主要在細胞週期中被合成,目前已被探討研究的有 8 種, 分別為 Cyclin A、Cyclin B1、Cyclin B2、Cyclin B3、Cyclin C、Cyclin D1、 Cyclin D2、Cyclin D3、Cyclin E、Cyclin F、Cyclin G 以及 Cyclin H,這些 Cyclins 在 N-端與 CDKs 鍵結處皆具有約 150 個胺基酸的相同區域,稱之 為 cyclin-box,在此可進行 Cyclins 與 CDKs 的結合(Evans et al., 1983),而 使細胞週期持續進行。. 表2-1. CDK inhibitor抑制CDK活性(資料來源:(Sherr and Roberts, 1995). 37.
(55) 在cyclins中,cyclin D為一生長的指標,也為有絲分裂及細胞週期提供 一個連結。在G1的早期,cyclin D1會CDK4或CDK6形成複合物,而導致細 胞週期的持續進行;異常的cyclin D1表現會在許多癌症中表現。cyclin D2 與cyclin D3則具有組織專一性(tissue specificity),較常在T 細胞型態的白血 球出現,其他組織例如乳癌組織則較少或沒有表現。而在不同之細胞週期, cyclin與CDK組成的複合體也不盡相同,以下將以週期為單位分別闡述(圖 2-10.): I. G1/S phase: 細胞在G1/S時期,主要表現的為Cyclin D與CDK6、Cyclin D與 CDK4以及CyclinE與CDK2的結合。此結合主要存在於G1 phase,並在 G1/S的過渡期間被分解。. II.G2/M phase: 在進入G2/M期之前,Cyclin A在S期中與CDK2形成複合物 (Girard et al., 1991; Walker and Maller, 1991),功能是促進DNA複 製。而G2/M期主要表現的複合體為Cyclin A與CDK1、Cyclin B與 CDK1 (cdc2) 的結合。. 38.
(56) 圖2-12. 細胞週期各調控蛋白分佈示意圖 (資料來源:www.bioon.com). 三、細胞週期檢查點(Cell cycle checkpoint) 檢查點的功能異常在許多癌症中會被發現(Vermeulen et al., 2003),因此 檢查點異常可能為造成細胞週期異常的原因之一。細胞週期檢查點對細胞 週期是一個重要的調控機制,正常之細胞具有兩個檢查點,其一為 G1/S 期檢查點:當生長因子作用在 G1 期,需要 Cyclin protein 及 CDK 同時結合 形成複合體,才能通過 G1 Checkpoint 進入到下一期(Long, 2005),而非繼 續停留於 G0 期。G1 期停滯為 p53 依賴型(Little et al., 1995),而在不同種 類的細胞株可發現抑制 p53 會增加化學療法及放射線療法的抵抗性(Russell. 39.
(57) et al., 1995)。 其二檢查點為 G2/M 期檢查點,當 DNA 受損會使細胞週期停滯在 G1 期或是 G2 期。停滯於 G1 期會使得 DNA 在複製之前先進行修補作用,而 停滯於 G2 期則會在有絲分裂中的染色體分裂前先進行修補的動作(Maity et al., 1994)。. 四、Wnt/-catenin pathway 參與細胞週期調控(Cell cycle progression) Wnt/-catenin pathway 可藉由胚胎發育(embryonic development)的 形態來調控細胞增生,這狀態廣泛的在人類疾病中被探討,特別是癌 症(Clevers, 2006; Logan and Nusse, 2004; Polakis, 2000 ; Reya and Clevers, 2005)。已從文獻得知,Wnt/-catenin pathway 可藉由增加 c-myc 以及 cyclin D1 的表現來提升 G1 期的進程(Tetsu and McCormick, 1999)。再者,Wnt 的標的基因 Axin2 的蛋白表現同樣也在有絲分裂期 (Mitosis)觀察到其表現量的高峰期(Hadjihannas and Behrens, 2006);不 僅如此,GSK3、APC、Axin2 皆為-catenin 裂解複合體的成員,也發 現三者對於有絲分裂(Mitosis)皆有直接的調控功能(Aoki and Taketo, 2007; Hadjihannas and Behrens, 2006; Kockeritz et al., 2006)。近期文獻指 出,細胞在有絲分裂 Mitosis 狀況下,也可調控細胞週期。LRP6 活化 為依賴細胞週期作用,藉由 cyclin Y/CDK14 在 G2/M 期時磷酸化 LRP6,使其活化,並且增加接受器的感受性來增強對 Wnt/-catenin pathway 的調控(Davidson and Niehrs 2010) (圖 2-11.)。Wnt/-catenin. 40.
(58) pathway 在細胞週期 G2/M 期中會表現出高峰點(Orford et al., 1999),因 -catenin 為有絲分裂期的重要調控因子(Huang et al., 2007a)。因此,不 僅 Wnt/-catenin pathway 的標的基因會調控細胞增生能力,細胞週期 也可藉由調控 Wnt/-catenin pathway,來影響細胞增生的進展。. 圖 2-13. Wnt/-catenin pathway 可被活化之細胞週期調控示意圖 (資料來源:The Science Creative Quarterly). 41.
(59) 第六節. Wnt/-catenin pathway 與細胞轉移之相關性. 一、 癌細胞的轉移(cell migration): 當原位惡性腫瘤(primary tumor)的病灶透過血液或是淋巴至其他器 官,這段過程即稱為轉移(metastasis) (Garbisa et al., 1986)。 轉移的過程包含五個步驟(圖 2-12.):. 圖 2-14. 為癌細胞轉移示意圖 (資料來源:Nature Reviews). 42.
數據


+7



相關文件
Center for Faculty Development, Department of medical education, Taichung Veterans General hospital 1 School of Medicine, National Yang-Ming University, Taipei, Taiwan 2.
臺大機構典藏NTUR (National Taiwan University 二 Repository, http://ntur.lib.ntu.edu.tw) 經驗與協助推 動臺灣學術機構典藏TAIR (Taiwan Academic Institutional Repository,
Students are asked to collect information (including materials from books, pamphlet from Environmental Protection Department...etc.) of the possible effects of pollution on our
The accuracy of a linear relationship is also explored, and the results in this article examine the effect of test characteristics (e.g., item locations and discrimination) and
北京中醫藥大學 天津中醫藥大學 上海中醫藥大學 遼寧中醫藥大學 南京中醫藥大學 浙江中醫藥大學 廈門大學 福建中醫藥大學 江西中醫藥大學 暨南大學
Center for Faculty Development, Department of Medical Education, Taichung Veterans General Hospital 1 Department of Nursing, Hungkuang University, Taichung, Taiwan 2. Section
Center for Faculty Development, Department of Medical Education, Taichung Veterans General Hospital 2 School of Medicine, National Yang-Ming University, Taipei, Taiwan 3.
The e xfoliated oral buccal cells and blood samples were collected for the assay of micronucleus frequency (MNF) and comet assay.. We find that there are higher MNF
