Safrole Suppresses Murine Leukemia WEHI-3 Cells In Vivo, and Stimulates Macrophages Phagocytosis and Natural Killer Cell Cytotoxic Activity in Leukemia Mice
Fu-Shun Yu1, Jai-Sing Yang2, Chun-Shu Yu3, Chi-Cheng Lu4, Jo-Hua Chiang4, Hsiung-Kwang Chung5,6, Chien-Chih Yu3, Chih-Chung Wu7, and Jing-Gung Chung8,9,*
1Department of Dental Hygiene, China Medical University, Taichung 404, Taiwan
2Department of Pharmacology, China Medical University, Taichung 404, Taiwan
3School of Pharmacy, China Medical University, Taichung 404, Taiwan
4Department of Life Sciences, National Chung Hsing University, Taichung 402, Taiwan
5Graduate Institute of Clinical Medical Science, China Medical University, Taichung 404, Taiwan
6Department of Otorhinolaryngology-Head & Neck Surgery, China Medical University Hospital, Taichung 404, Taiwan
7Department of Nutrition and Health Sciences, Chang Jung Christian University, Tainan 711, Taiwan
8Department of Biological Science and Technology, China Medical University, Taichung 404, Taiwan
9Department of Biotechnology, Asia University, Wufeng, Taichung 413, Taiwan
Running title: Safrole inhibits murine leukemia WEHI-3 cells in vivo.
Correspondence to: Jing-Gung Chung, Department of Biological Science and Technology, China Medical University, No 91, Hsueh-Shih Road, Taichung 404,
Taiwan. Tel: +886 4 2205 3366 ext 2161, Fax: +886 4 2205 3764, e-mail:
ABSTRACT. Many anticancer drugs (phytochemicals) are obtained from natural products. However, some of phytochemicals have mutagenic effects. Our earlier studies have been shown that safrole is a component of Piper betle inflorescence, have been reports it is a carcinogen but we did found that safrole induced apoptosis of human oral cancer in vitro and inhibited this cancer cells in xenograft tumor cells in vivo. However, there is no information to show safrole could promote immune
responses. Thus, we hypothesized that safrole would have immune responses on leukemia mice in vivo. We used leukemia cells (WEHI-3) to generate the leukemia mice then oral treatment with or without safrole, animal were sacrified for examining the cell marker population, the weights of spleen and liver, phagocytosis of macrophage and the activities of natural killer cells. Results indicated that safrole increased the body weight, decreased the weights of spleen and liver of leukemia mice. Furthermore, safrole promoted the activities of macrophages phagocytosis and NK cells from leukemia mice compared with the untreated leukemia mice. After examining the cell marker population, we found that safrole promoted the levels of CD3 (T cells) and CD19 (B cells) and Mac-3 (macrophage) but did not affect CD11b (monocytes) in leukemia mice.
Keywords: Safrole; murine leukemia WEHI-3 cells; macrophages phagocytosis; NK cells cytotoxic activity; in vivo
INTRODUCTION
It is well documented that diet can play a vital role in cancer prevention.
Nowadays numerous scientists are looking for food materials from natural plant which can potentially prevent the cancer occurrence. Increase consumption of a plant-based diet can reduce the risk of cancer development from epidemiological and animal studies (Mahmoud et al. 2000; Mutoh et al. 2000; Wenzel et al. 2000). Cancer is the major cause death in human population and leukemia is one of the cancers for causing dearth in humans worldwide. About 3.7 individuals per 100,000 die each year from leukemia in U.S (Jensen et al. 2004). About 2.1 individuals per 100,000 thousand die per year of leukemia in Taiwan that based on 2008 reports of the Department of Health, Executive Yuan, R.O.C. (Taiwan). Conventional treatments for patients with leukemia are radiotherapy, chemotherapy, or a combination of both but have proven unsatisfactory.
In Taiwan, it was reported that people chewing betel quid containing piper betle inflorescence can generate a high concentration of safrole (420 μM) in the saliva (Wang and Hwang 1993). Although safrole have been recognized to be as a group 2B carcinogen (safrole is a documented rodent carcinogen) but no adequate evidences elucidating the relationship between exposure to safrole and human cancers (1976).
Safrole have been reported to cause marked [Ca2+]i elevation and decreased cell viability in human osteosarcoma cells (Lin et al. 2006) and to bind to DNA then to form safrole-DNA adduct formation (Daimon et al. 1997; Daimon et al. 1998; Lee et al. 2005).
It was reported that safrole reduced the bactericidal activity of polymorphonuclear leukocytes (PMNs) against A. actinomycetemcomitans (American Type Culture Collection [ATCC] 33384, serotype c) and safroel also
reduced the release of superoxide anions from PMNs (10). Our previous studies have been shown that safrole induced apoptosis in human oral cancer HSC-3 cells in vitro and in xenograft mice model in vivo (Yu et al. ). Currently there are no reports to show the effects of safrole on the immune system of leukemia mice in vivo. In the present, we demonstrated that safrole can promoted phagocytosis of macrophages and increase activities of natural killer cells from leukemia BLAB/c mice in vivo.
MATERIALS AND METHODS
Materials and reagents
Safrole and dimethyl sulfoxide (DMSO) were obtained from Sigma-Aldrich Corp.
(St. Louis, MO, USA). RPMI-1640 medium, fetal bovine serum (FBS), penicillin- streptomycin and L-glutamine were obtained from Invitrogen Life Technologies (Carlsbad, CA, USA).
Murine WEHI-3 leukemia cell line
The WEHI-3 murine myelomonocytic leukemia cell line was obtained from the Food Industry Research and Development Institute (Hsinchu, Taiwan). Cells were cultured in culture plastic flasks (75-cm2) at 37°C under a humidified 5% CO2
atmosphere in RPMI-1640 medium supplemented with 10% FBS, 2 mM L-glutamine, 100 units/ml penicillin and 100 μg/ml streptomycin. The cells were cultivated for two complete cycles in an incubator.
Male BALB/c Mice
Fifty male BALB/c mice 8 weeks of age and approximately 22-28 g in weight were obtained from the Laboratory Animal Center, College of Medicine, National
Taiwan University (Taipei, Taiwan).
Establish leukemia mice and safrole treatment
Fifty BALB/c mice were randomly divided into 5 groups to receive different treatments. Then thirty mice from randomly 3 groups were intraperitoneally (i.p.) injected with 1×105 WEHI-3 cells for 2 weeks and then were randomly separated into 3 groups as a model of leukemia.
Group I served as a control (10 animals). Group II mice were treated with oliver oil as vehicle (10 animals). Group III animals were treated with oliver oil (10 animals) after intraperitoneal injection WEHI-3 cells. Group IV mice were treated with safrole (e mg/kg) in oliver oil (10 animals) after intraperitoneal injection of WEHI-3 cells. Group V animals were treated with safrole (16 mg/kg) in oliver oil (10 animals) after intraperitoneal injection of WEHI-3 cells. Safrole was administered by oral gavage to the treatment groups the above dose daily for up to 3 weeks before being weighed (Kwon et al. 2009).
Body, spleen and liver tissues weights
All animals were weighed and blood withdrawn. Spleen and liver samples were isolated and weighed individually.
Quantification of phagocytic activity of macrophages
PHAGOTEST kit (Glycotope Biotechnology GmbH, Heidelberg, Germany) was used to measure the phagocytosis as previously described (Kwon et al. 2009).
Approximately 1×105 leukocytes in 100 μl whole blood from each treatment of each group was incubated for 1 h at 37°C with fluoresce in isothiocyanate-labelled E. coli (20 μl). The quenching solution (100 μl) was added to the reaction according to the
manufacturer’s instruction. After the completion of phagocytosis by monocytes/macrophages, DNA was stained according to the manufacturer’s protocol.
Cells from each sample were analyzed by flow cytometery as previously described (Kwon et al. 2009). Fluorescence data were collected on 10,000 cells and analyzed using the CELLQUEST software.
Whole blood samples and immunofluorescence staining for surface markers from leukemia mice after exposed to safrole
At the end of treatment, blood samples of 1 ml from all experimental mice were collected before mice were sacrificed. Collected blood sample were individually exposed to 1× Pharm LyseTM lysing buffer (BD Biosciences, San Jose, CA, USA) for lysing of the red blood cells and then all samples were centrifuged for 15 min at 1500 rpm at 4°C. The isolated white blood cells from each treatment were stained by the FITC anti-mouse CD3, PE anti-mouse CD19, PE anti-mouse Mac-3 and FITC anti- mouse CD11b antibodies (BD Pharmingen Inc, San Diego, CA, USA) before being analyzed to determine the levels of cell marker by flow cytometry as previously described (Kwon et al. 2009; Lin et al. 2009)
Quantification of NK cell cytotoxic activity
At the end of treatment, about 1×105 leukocytes from the spleens of all treatments in 1 ml of RPMI-1640 medium were cultured in each well of 24-well culture plates for 24 h. About 2.5×107 of YAC-1 cells were cultured in 15-mL tubes with serum-free RPMI-1640 medium and then PKH-67/Dilunt C buffer (Sigma- Aldrich, St Louis, MO, USA) was added to the cells, mixed thoroughly for 2 min at 25˚C then 2 ml PBS was added for 1 min. RPMI-1640 medium at 4 ml was added for a 10 min-incubation then were followed by centrifugation at 1200 rpm of 25˚C.
YAC-1 cells in 100 μl were placed on 96-well plates before the addition of the leukocytes from each treatment to the wells for 12 h and determination of NK cell activity by flow cytometry (FACS CaliburTM, Becton Dickinson, NJ, USA) as previously described (Tang et al. 2009; Yu et al. 2006).
Statistics analysis
The results were expressed as mean ± S.D. and the difference between control and safrole treated groups was analyzed by one-way ANOVA followed by Dunnett's multiple comparison test and Student’s t test. p values of less or equal to 0.05 were taken as significant.
Results
Safrole affected the body, spleen and liver weight in WEHI-3 leukemia BLAB/c mice
At the end of safrole treatment, body weights of each animal were weighed then spleen or liver tissues were isolated and were weighed and results were showed in Figure 1A, B, C and D. Results indicated that safrole increased the body weights of each treatment after compared to the leukemia control (Fig. 1A) but significantly decreased the spleen (Fig. 1B and C) and liver (Fig. 1D) weights.
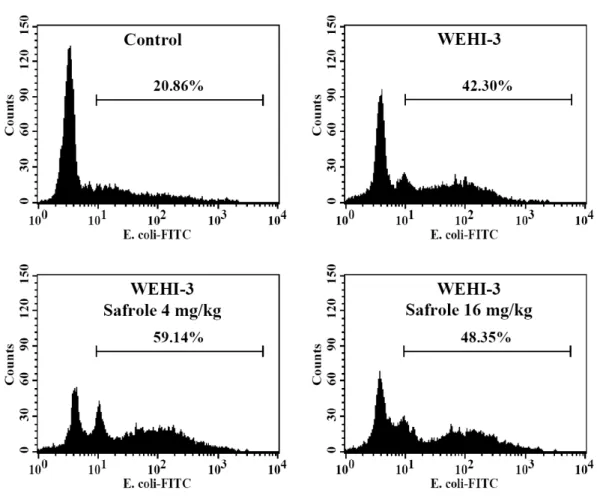
Safrole promoted the phagocytosis by macrophage from peripheral blood mononuclear cell (PBMC) and peritoneal cavity in WEHI-3 leukemia BALB/c mice
In order to investigate whether or not safrole affected phagocytosis, the leukocytes from safrole-treated or untreated groups were isolated and phagocytic
activity was measured. The data from Figure 2 demonstrated that safrole (4 and 16 mg/kg/day) promoted the activity of phagocytosis from PMBC (4 mg/kg/day: 22.9%;
16 mg/kg/day: 10.4%) (Fig. 2A and B) and peritoneal cavity (4 mg/kg/day: 62.5%; 16 mg/kg/day: 112.5%) (Fig. 2C and D) from leukemia mice.
Safrole affected on surface markers of whole blood cells from WEHI-3 leukemia BALB/c mice
In order to investigate whether safrole affected the levels of cell surface marker, leukocytes from safrole-treated or untreated groups were isolated and levels of CD3, CD19, Mac-3 and CD11b were measured. The data for cell markers are presented in Figure 3A, B, C, D, E, F and G. The results indicated that safrole increased the levels of CD3 (Fig. 3A and B) (4 mg/kg/day:22.3%), CD19 (Fig. 3C) (4 mg/kg/day: 38.4%) and Mac-3 (Fig. 3F and G) (4 mg/kg/day: 61.1%; 16 mg/kg/day: 66.6%) but did not affected CD11b (Fig. 3D and E) when compared to the WEHI-3 only treated groups.
Safrole affected NK cell activity of splenocytes in WEHI-3 leukemia BALB/c mice
In order to investigate whether safrole is able to act on NK cell activity, leukocytes from safrole-treated or untreated groups were isolated and NK cell activity was determined. The results presented in Figure 4, which indicated that the YAC-1 target cells were killed by NK cells from safrole treated group when compared to the untreated groups at 16 mg/kg/day at target cells ratio of 25:1 and 50:1. This dose was effective at both target ratios and increased the activity of NK cells (16 mg/kg/day, 100.5% at target cells ratio of 25:1; 66.4% at target cells ratio of 50:1).
DISCUSSION
Several reports have been shown that safrole act as a carcinogen and it can binding to DNA of cells (Daimon et al. 1997; Daimon et al. 1998; Lin et al. 2006). It was also reported that safrole decreased cell viability and cause marked [Ca2+]i elevation in human osteosarcoma cells (Lin et al. 2006). Recently our primary reports also demonstrated that safrole induced cytotoxic effects and induction of apoptosis in human oral cancer cells in vitro and xenograft animal model in vivo (Yu et al. ).
However, there is no available information to address the effect of safrole on the immune responses in leukemia mice in vivo. Herein, we established leukemia BLAB/c mice through the injection of WEHI-3 cells, and then chronically treated mice with safrole. Our findings indicated that safrole decrease the weights of animal body, the liver and spleen in leukemia mice after compared to the leukemia mice without treatment with safrole (control) (Fig. 1).
Furthermore, we also found out that safrole promoted the phagocytosis of macrophage (Fig. 2) and elevated the activities of NK cells (Fig. 4). Thus, safrole could not only increase the humoral immune response (B cells and promoted macrophage activities), but also the cellular immune response (T cells) when the leukemia mice were treated with safrole. The safrole significantly enhanced NK cell activities (Fig. 4) from leukemia mice. It is well known that macrophage phagocytosis and NK cell activities both are played major roles for immune responses after animals were exposed to antigen Human T cell and monocyte modulating activity of Rhizoma typhonii in vitro (Mulligan et al. 2008; Shan et al. 2001; Thomas et al. 1985). These findings suggested that safrole increased the immune response and might increase the activities of macrophage and NK cells against leukemia in general. These observations are in agreement with an earlier study in normal mice showing that oral treatment of safrole also promoted immune responses (data not shown).
We also examined the cell markers from leukemia mice after oral treatment of safrole and results indicated that the percentages of CD3 (T cells), CD19 (B cells) and Mac-3 (macrophage) were significantly increased in safrole-treated leukemia mice, but that the population of CD11b (monocytes) were not significantly effected in safrole treated leukemia mice. It was reported that CD19 antigen exists on cell surface membranes of non-activated B lymphocytes (Krop et al. 1996). CD3 and CD19 positive cells represented the percentages of T and B cells increase from safrole- treated leukemia mice and this is in comprisable to Figure 3. It was reported that the B-cell differentiation requires the interaction of various cytokines, which are secreted from macrophages or T cells (Mitsuzumi et al. 1998). Our results showed that safrole could increase B cells (elevated the population of CD19), the augmenting effect of safrole may through the antibody responses resulted from expansion and differentiation of mainly monocytes and macrophages. The results suggested that safrole inhibited leukemia-related spleen growth.
In the present study, we also found that safrole could promote immune responses in BALB/c leukemia mice in vivo. The safrole can act as a potent immunological adjuvant in vivo and its application provides an effective strategy to improve the efficacy of immune responses.
Declaration of Conflicting Interests
The authors have no conflicts of interests.
ACKNOWLEDGEMENTS
This study was supported by Grant DOH100-TD-C-111-005 from Department of Health, Executive Yuan, R.O.C (Taiwan).
REFERENCES
1976. IARC monographs on the evaluation of the carcinogenic risk of chemicals to man: some naturally occurring substances. IARC Monogr Eval Carcinog Risk Chem Man 10:1-342.
Daimon H, Sawada S, Asakura S, Sagami F. 1997. Analysis of cytogenetic effects and DNA adduct formation induced by safrole in Chinese hamster lung cells.
Teratog Carcinog Mutagen 17(1):7-18.
Daimon H, Sawada S, Asakura S, Sagami F. 1998. In vivo genotoxicity and DNA adduct levels in the liver of rats treated with safrole. Carcinogenesis 19(1):141-6.
Jensen CD, Block G, Buffler P, Ma X, Selvin S, Month S. 2004. Maternal dietary risk factors in childhood acute lymphoblastic leukemia (United States). Cancer Causes Control 15(6):559-70.
Krop I, Shaffer AL, Fearon DT, Schlissel MS. 1996. The signaling activity of murine CD19 is regulated during cell development. J Immunol 157(1):48-56.
Kwon SH, Nam JI, Kim SH, Kim JH, Yoon JH, Kim KS. 2009. Kaempferol and quercetin, essential ingredients in Ginkgo biloba extract, inhibit interleukin- 1beta-induced MUC5AC gene expression in human airway epithelial cells.
Phytother Res 23(12):1708-12.
Lee JM, Liu TY, Wu DC, Tang HC, Leh J, Wu MT, Hsu HH, Huang PM, Chen JS, Lee CJ and others. 2005. Safrole-DNA adducts in tissues from esophageal cancer patients: clues to areca-related esophageal carcinogenesis. Mutat Res 565(2):121-8.
Lin HC, Cheng HH, Huang CJ, Chen WC, Chen IS, Liu SI, Hsu SS, Chang HT, Huang JK, Chen JS and others. 2006. Safrole-induced cellular Ca2+ increases and death in human osteosarcoma cells. Pharmacol Res 54(2):103-10.
Lin JP, Yang JS, Lu CC, Chiang JH, Wu CL, Lin JJ, Lin HL, Yang MD, Liu KC, Chiu TH and others. 2009. Rutin inhibits the proliferation of murine leukemia WEHI-3 cells in vivo and promotes immune response in vivo. Leuk Res 33(6):823-8.
Mahmoud NN, Carothers AM, Grunberger D, Bilinski RT, Churchill MR, Martucci C, Newmark HL, Bertagnolli MM. 2000. Plant phenolics decrease intestinal tumors in an animal model of familial adenomatous polyposis. Carcinogenesis 21(5):921-7.
Mitsuzumi H, Kusamiya M, Kurimoto T, Yamamoto I. 1998. Requirement of cytokines for augmentation of the antigen-specific antibody responses by ascorbate in cultured murine T-cell-depleted splenocytes. Jpn J Pharmacol 78(2):169-79.
Mulligan JK, Lathers DM, Young MR. 2008. Tumors skew endothelial cells to disrupt NK cell, T-cell and macrophage functions. Cancer Immunol Immunother 57(7):951-61.
Mutoh M, Takahashi M, Fukuda K, Komatsu H, Enya T, Matsushima-Hibiya Y, Mutoh H, Sugimura T, Wakabayashi K. 2000. Suppression by flavonoids of cyclooxygenase-2 promoter-dependent transcriptional activity in colon cancer cells: structure-activity relationship. Jpn J Cancer Res 91(7):686-91.
Shan BE, Zhang JY, Li QX. 2001. [Human T cell and monocyte modulating activity of Rhizoma typhonii in vitro]. Zhongguo Zhong Xi Yi Jie He Za Zhi 21(10):768-72.
Tang NY, Yang JS, Lin JP, Hsia TC, Fan MJ, Lin JJ, Weng SW, Ma YS, Lu HF, Shen JJ and others. 2009. Effects of Agaricus blazei Murill extract on immune responses in normal BALB/c mice. In Vivo 23(5):761-6.
Thomas PT, Ratajczak HV, Aranyi C, Gibbons R, Fenters JD. 1985. Evaluation of
host resistance and immune function in cadmium-exposed mice. Toxicol Appl Pharmacol 80(3):446-56.
Wang CK, Hwang LS. 1993. Phenolic compounds of betel quid chewing juice. Food Sci 20(44):458-471.
Wenzel U, Kuntz S, Brendel MD, Daniel H. 2000. Dietary flavone is a potent apoptosis inducer in human colon carcinoma cells. Cancer Res 60(14):3823- 31.
Yu FS, Yang JS, Yu CS, Lu CC, Chiang JH, Lin CW, Chung JG. Safrole Induces Apoptosis in Human Oral Cancer HSC-3 Cells. J Dent Res 90(2):168-74.
Yu FS, Yu CS, Chan JK, Kuo HM, Lin JP, Tang NY, Chang YH, Chung JG. 2006.
The effects of emodin on the expression of cytokines and functions of leukocytes from Sprague-Dawley rats. In Vivo 20(1):147-51.
Figure Legends
Fig. 1. Safrole affected the body, spleen and liver weights of leukemia BALB/c mice.
The mice were intraperitoneally injected with WEHI-3 cells (1×105 cells/100 μl) for 2 weeks and then orally treated with or without safrole (4 and 16 mg/kg/mice) for 3 weeks. The body (A), spleen (B, C) and liver (D) weights were weighted as described in “Materials and methods”. Each point is mean ± S.D (n=10).*p<0.05 was considered significant when compared with the WEHI-3 leukemia mice.
Fig. 2. Safrole stimulated phagocytotic activity of PBMC and peritoneal cavity in leukemia BALB/c mice. Macrophages were isolated from PBMC (A and B) and peritoneal cavity (C and D) of each group from leukemia BALB/c mice after exposure to 4 and 16 mg/kg/day of safrole by oral administration for 21 days. The percentages of phagocytosis with phagocyte green fluorescent particles (FITC-E. coli.) at safrole oral treatment were determined by flow cytometric analysis as described in “Materials and methods”. Each point is mean ± S.D (n=10). *p<0.05 was considered significant when compared with the WEHI-3 leukemia mice.
Fig. 3. Safrole affected the levels of cell markers in white blood cells from leukemia BALB/c mice. The mice were intraperitoneally injected with WEHI-3 cells for 2 weeks and orally treated with or without safrole for 3 weeks. Blood was collected from each group animal and was analyzed for cell markers (A: CD3; B: CD19; C:
Mac-3 and D: CD11b) by flow cytometry as described in “Materials and methods”.
The data is expressed mean ± S.D. of three experiments. Each point is mean ± S.D.
(n=10). *p<0.05 was considered significant when compared with the WEHI-3 leukemia mice.
Fig. 4. Safrole affected activity of natural killer (NK) cells in leukemia BALB/c mice.
The YAC-1 target cells were killed by NK cells of splenocytes from the mice after treatment with safrole by oral administration at 4 and 16 mg/kg/day in target cells ratio of 25:1 and 50:1 Each point is mean ± S.D.*p<0.05 was considered significant when compared with the WEHI-3 leukemia mice (n=10).
Figure 1 A
B
C
D
Figure 2 A PMBC
B PBMC
C Peritoneal cavity
D Peritoneal cavity
Figure 3 A
B
C
D
E
F
G
Figure 5