Establishment of a Rapid PCR Detection Method for Antrodia salmonea and A. cinnamomea
Meng-Ling Wu,1) Tang-Long Shen,2) Tun-Tschu Chang,1) Chao-Han Chen,1) Ting-Hsuan Hung,2,3) Chih-Yun Lee,1) Chun-Yi Lin2)
【Summary】
Both Antrodia cinnamomea and A. salmonea are fungi of the Polyporaceae. Antrodia cinna- momea, which only grows on the inner side of the trunk of Cinnamomum kanehirai Hayata (Lau- raceae), is a precious medicinal fungus; A. salmonea causes brown heart rot disease of Cunningha- mia konishii Hayata (Cunninghamieae) in Taiwan. It is difficult to distinguish A. cinnamomea from A. salmonea because of their morphological similarities. Antrodia salmonea is commonly used as a counterfeit substitute for A. cinnamomea and sold by dishonest merchants. In 2004, these 2 fungi were first distinguished by the pore surface color of the basidiomata, host preferences, and mat- ing types. However, accurate identification relies on professional training and experience. More- convenient and-persuasive methods are necessary for precisely identifying A. cinnamomea. In this study, we applied 6 fungal primer pairs in polymerase chain reaction (PCR) assays, and analyzed sequences of amplified DNA fragments between A. cinnamomea and A. salmonea. Results showed that one of these primer pairs could amplify a particular DNA fragment from A. salmonea, which was approximately 1.5 kb longer than that from A. cinnamomea. Based on this 1.5-kb difference in sequences, another primer pair named Acl-F/Acl-R was designed for the specific detection of A.
salmonea. A specific fragment, of 219 bp, was yielded only from A. salmonea, whereas no frag- ment was yielded from A. cinnamomea by the PCR assay. In this study, we established a rapid and accurate identification technique for A. salmonea, which will be helpful in rapidly differentiating A.
salmonea from A. cinnamomea. Additionally it also saves significant time as the entire procedure only takes about 1 h and has high sensitivity. This identification method can provide objective evi- dence for industry and public institutions.
Key words: Antrodia cinnamomea, Antrodia salmonea, specific PCR detection.
Wu ML, Shen TL, Chang TT, Chen CH, Hung TH, Lee CY, Lin CY. 2015. Establishment of a rapid PCR detection method for Antrodia salmonea and A. cinnamomea. Taiwan J For Sci 30(1):45-53.
1)Forest Protection Division, Taiwan Forestry Research Institute, 53 Nanhai Rd., Taipei 10066, Taiwan. 林業試驗所森林保護組,10066台北市南海路53號。
2)Department of Plant Pathology and Microbiology, National Taiwan Univ., 1 Roosevelt Rd., Sec. 4, Taipei 10617, Taiwan. 國立台灣大學植物病理與微生物學系,10617台北市羅斯福路四段1號。
3)Corresponding author, e-mail:[email protected] 通訊作者。
Received July 2014, Accepted March 2015. 2014年7月送審 2015年3月通過。
研究報告
應用PCR鑑定技術區分牛樟芝(Antrodia cinnamomea) 與香杉芝(A. salmonea)
吳孟玲1) 沈湯龍2) 張東柱1) 陳昭翰1) 洪挺軒2,3) 李芷芸1) 林雋軼2)
摘 要
牛樟芝(Antrodia cinnamomea)及香杉芝(A. ntrodia salmonea)均屬多孔菌科(Polyporaceae)真菌。
牛樟芝是台灣特有的珍貴真菌,只生長在台灣特有的牛樟樹,貼生於樹幹中空之內面。香杉芝生長在 已腐朽的香杉樹幹,以前被誤認為牛樟芝,於2004年才被認定為Antrodia屬之新種。這兩種真菌由於 形態相似,通常以孢子形態、子實體、寄主偏好和交配型作為區隔。香杉芝常被不肖業者用作牛樟芝 的替代物,但傳統鑑定方法上,此兩種菌種的鑑定需要專業知識及經驗,因此需要真菌鑑定專業人員 參與,鑑定作業實屬不易。在這項研究中,我們測試了六組真菌引子對,並且解析了這兩真菌之間的 序列,我們發現其中一引子對進行PCR增幅時,香杉芝比牛樟芝多出一段1.5 kb之核酸片段,我們並根 據此片段另外設計了一組引子對Acl-F/Acl-R,增幅片段為219 bp,可針對香杉芝進行快速鑑定。本研 究建立快速而準確的牛樟芝與香杉芝菌種鑑定方法,並建立快速鑑定流程,檢測靈敏度高,且鑑定時 間僅需5小時,可提供牛樟芝與香杉芝之鑑定辨識,以及提供產業界及公務部門鑑定之依據。
關鍵詞:牛樟芝、香杉芝、聚合酵素連鎖反應專一性鑑定。
吳孟玲、沈湯龍、張東柱、陳昭翰、洪挺軒、李芷芸、林雋軼。2015。應用PCR鑑定技術區分牛樟芝 (Antrodia cinnamomea)與香杉芝(A. salmonea)。台灣林業科學30(1):45-53。
INTRODUCTION
Cunninghamia konishii Hayata (Cun- ninghamieae) is a coniferous tree that is endemic to Taiwan and grows in broadleaf and coniferous forests at elevations of 1300~
2800 m in the central and northern parts of the island. Brown heart rot associated with resupinate, salmon-pink basidiomata in empty rotten trunks of Cun. konishii has vernacu- larly been called Antrodia salmonea. This fungus is similar to A. cinnamomea (Chang and Chou 1995), which bears the vernacular name A. cinnamomea and has only been col- lected from the endemic aromatic tree Cinna- momum. kanehirai Hayata (Lauraceae) in Tai- wan, but the pore surface of its basidiomata
is a different color (Fig. 1). The basidiomata of A. cinnamomea have medicinally been used for treatment of food and drug intoxica- tion, diarrhea, abdominal pain, hypertension, skin itching, and cancer. Both species have a strong bitter taste, believed to indicate the presence of effective medicinal ingredients.
Therefore, it is said that A. salmonea can be substituted for A. cinnamomea. Antrodia cin- namomea and A. salmonea were first thought to be the same species but different varieties which grow on different hosts that exhibit distinct basidiomata. However distinct colo- nies, host preferences, and mating systems showed that they are quite different from each
other; thus, A. salmonea was separated from A. cinnamomea in 2004 (Chang and Chou 2004). Although both A. cinnamomea and A.
salmonea contain some medicinal proper- ties, their economic values enormously differ.
Their high morphological similarities make it difficult to distinguish these 2 fungi solely by visual observation, so there are many prod- ucts on the market purported to contain A.
cinnamomea which are actually made with A.
salmonea. This phenomenon makes accurate identification of A. salmonea and A. cinna- momea important and necessary. Prior to this study, accurate identification relied on profes- sional training and experience. To establish a less time-consuming molecular method for accurate identification, we tested 3 nuclear primer pairs and 3 mitochondrial primer pairs for a polyermase chain reaction (PCR) ex- amination and found sequence differences be- tween these 2 fungi. Based on the difference sequences, another primer pair named Acl-F/
Acl-R was designed for the specific detection of A. salmonea. A rapid and convenient PCR system was established for examination.
MATERIALS AND METHODS Isolates examined
Antrodia cinnamomea isolates included B71, B85, B86, B573, B574, B985, and TF986; A. salmonea isolates included B147, B492, B495, TF971, TF1004, and TF1012.
Isolate culture
Pure cultures were obtained from basidi- omata and grown at 25℃ on malt extract agar (MEA: 2% malt extract, 2% glucose, and 2%
Bacto agar) and potato dextrose agar (PDA).
Petri dishes with an 85-mm internal diameter and which contained 20~25 ml of agar media were inoculated with a piece of mycelium at the center, kept at 25℃, and macroscopically examined after 2 and 6 wk of incubation.
Fungal DNA extraction
The DNA extraction method was modi- fied from Hung (1999). A 0.4×0.4-cm agar plate was scraped with cultured A. cinna- momea and A. salmonea hyphae and placed into an Eppendorf tube; then, the hyphae Fig. 1. Basidiomata of Antrodia cinnamomea (A) and A. salmonea (B). Colony morphology of A. cinnamomea (C) and A. salmonea (D) after growing on MEA plates for 3 wk at 25℃.
(A) (B) (C) (D)
were homogenized with a plastic pestle. DNA extraction buffer (0.9 ml; pH 8.0: 100 mM Tris-HC1, 100 M EDTA, 250 mM NaC1) and 0.1 ml Sarkosyl (20%) were added. The suspension was vortexed and incubated at 55℃ for 1h. The tube was centrifuged at 6000 rpm for 5 min. The supernatant (0.8 ml) was saved, 100 μl of 5 M NaC1 and 100 μl of CTAB/NaC1 (10% CTAB in 0.7 M NaC1) were added and incubated at 65℃ for 10 min.
An equal volume of chloroform/isoamyl al- cohol (24: 1) was added, mixed thoroughly, and spun at 11,000 rpmfor 5 min. The aque- ous suspension was saved. Equal volumes of phenol/chloroform/isoamyl alcohol (25: 24:
1) were added, mixed thoroughly, and spun at 11,000 rpm for 5 min. The aqueous suspen- sion was saved. Isopropanol 0.6 volume was added to precipitate the nucleic acids, and the mixture was spun at 12,000 rpm for 30 min.
The pellet was washed with 70% ethanol to remove the residual CTAB. Briefly, the pel- let was dried and resuspended in 100 µl of TE buffer (pH 8.0: 10 mM Tris and 1 mM EDTA), and stored at -20℃.
PCR conditions
A PCR was conducted in 25-µl reaction mixtures (10 mM Tris-HCl at pH 8.7, 50 mM KCl, 2 mM MgCl2, 0.25 mM of each dNTP, 1.25 units of Taq DNA polymerase, 0.2 µM each primer, and 2 µl of extracted DNA).
All PCR reagents were procured from super- therm taq (Invitrogen, MA, USA). The PCR was carried out with the following setup: ini- tial incubation at 94℃ for 2 min, followed by 30 cycles of a denaturing step at 94℃ for 30 s, an annealing step at 58℃ for the ML3/ML4 primers and 56℃ for the Acl-F/Acl-R prim- ers for 30 s, extension at 72℃ for 1.5 min for the ML3/ML4 primers and 20 s for the Acl-F/
Acl-R primers, and a final extension of 5 min at 72℃. The amplified DNA was analysed
on 1.5% agarose gels stained with ethidium bromide. Fluorescence of the DNA bands was visualized with ultraviolet light (302 nm) with the AlphaImage 2200 system (AlphaInnotech, CA, USA).
Primers for the PCR examination
Six pairs of common primers were tested: including 3 pairs of nuclear primers (ITS1F/ITS4, NS3/NS4, and NS5/NS6) and 3 pair of mitochondrial primers (MS1/MS2, ML1/ML2, and ML3/ML4) (Gardes and Bruns 1993, White et al. 1990). According to the sequencing results, a specific primer pair was designed from the 2.4-kb fragment de- rived from A. salmonea named Acl-F/Acl-R.
All primers sequences are shown in Table 1.
The 900- and 2384-bp fragments yielded by ML3/ML4 of several individuals of A.
cinnamomea and A. salmonea were also se- quenced and analyzed. The genetic affiliation was analyzed by DNAStar (Madison, WI, USA).
RESULTS PCR examination
For the first step, 6 pairs of common primers were used. PCR results for 5 pairs of primers showed no difference in product sizes between A. cinnamomea and A. sal- monea (Fig. 2), although the sequences con- tained certain variances. Only the ML3 and ML4 primers gave quite different results; an 871-bp fragment was amplified from A. cin- namomea, whereas a 2.4-kb fragment was amplified from A. salmonea (Fig. 3). Further PCR examination of the ML3/ML4 primer pair for other common pathogen fungi was also carried out, and different fragment sizes were found among them (Fig. 4). The primer pair, Acl-F/Acl-R, was designed based on the ML3/ML4 PCR product of A. salmonea
Table 1. Classification and sequences of primers used in this study Primer Sequence
Nuclear primers ITS1F 5’-CTTGGTCATTTAGAGGAAGTAA-3’
ITS4 5’-TCCTCCGCTTATTGATATGC-3
NS3 5’-GCAAGTCTGGTGCCAGCAGCC-3’
NS4 5’-CTTCCGTCAATTCCTTTAAG-3’
NS5 5’-AACTTAAAGGAATTGACGGAAG-3’
NS6 5’-GCATCACAGACCTGTTATTGCCTC-3’
Mitochondrial primers MS1 5’-CAGCAGTCAAGAATATTAGTCAATG-3’
MS2 5’-GCGGATTATCGAATTAAATAAC-3’
ML1 5’-GTACTTTTGCATAATGGGTCAGC-3’
ML2 5’-TATGTTTCGTAGAAAACCAGC-3’
ML3 5’-GCTGGTTTTCTACGAAACATATTTAAG-3’
ML4 5’-GAGGATAATTTGCCGAGTTCC-3’
Specific primers Acl-F 5’-AGAACACGAATACAAGGTC-3’
Acl-R 5’-GCAAGTTACCCATTTTATCAA-3’
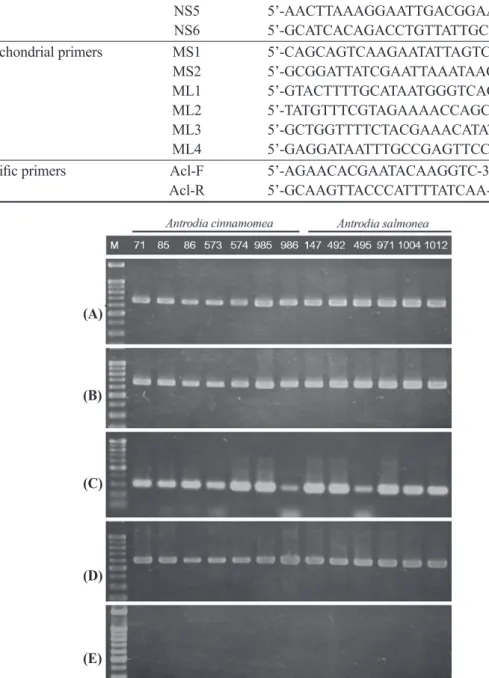
Fig. 2. Results of PCR amplification of 5 primer pairs, (A) ITS1F/ITS4, (B) NS3/NS4, (C) NS5/NS6, (D) MS1/MS2, and (E) ML1/ML2. The PCR test showed no differences between A.
cinnamomea and A. salmonea. M: 100-bp ladder marker.
(A)
(B)
(C)
(D)
(E)
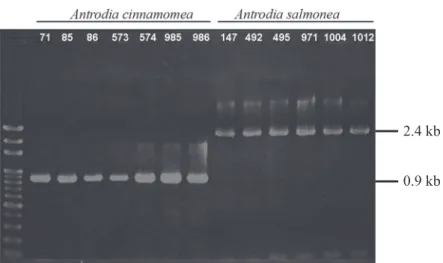
Fig. 3. PCR results of the ML3/ML4 primer pair, showing a significant difference of Antrodia cinnamomea and A. salmonea, which yielded 871- and 2384-bp fragments, respectively. M: 100-bp ladder marker.
Fig. 4. Amplification of the ML3/ML4 primer pair for common fungal pathogens in Taiwan.
M: 100-bp ladder marker; lane 1: Antrodia cinnamomea; lane 2: A. salmonea. Lanes 3 to 13 are: Gloeophyllum trabeum, G. sepiarium, G. abietinum, Trametes versicolor, Lentinus lepideus, Asp. niger, Asp. restrictus, Chaetomium globosum, Cladosporium cladosporioides, Cla. herbarum, and Phellinus noxius, respectively.
and then tested on A. cinnamomea and A.
salmonea. Antrodia salmonea yielded a 219- bp fragment, but none was found in A. cinna- momea (Fig. 5).
Sequence alignment and phylogenetic analysis
The 2384-bp sequences of the 2 frag- ments were quite similar; the 871-bp fragment amplified from A. cinnamomea matched the
front and rear ends of the 2384-bp fragment of A. salmonea (Fig. 6). Seven sequences acquired from A. cinnamomea were submit- ted to GenBank, KP338795, KP338796, KP338797, KP338798, KP338799, KP338800, and KP338801; 6 from A. salmonea were also submitted as KP338802, KP338803, KP338804, KP338805, KP338806, and KP338807 to GenBank. The genetic affilia- tion was analyzed by DNAStar (Fig. 7).
2.4 kb 0.9 kb
M 1 2 3 4 5 6 7 8 9 10 11 12 13
DISCUSSION AND CONCLUSIONS Out of these 6 primer pairs, only ML3/
ML4, which are mitochondrial primers, yield- ed distinct PCR fragments (Figs. 2, 3). We
also tested ML3/ML4 on other common fun- gal pathogens in Taiwan (Fig. 4), and found that ML3/ML4 could be a universal primer pair for the rough rapid pathogen detection of these Taiwanese fungi. For more-precise Fig. 5. Amplification of the specific primer pair, Acl-F/Acl-R, on Antrodia cinnamomea and A.
salmonea. A 219-bp fragment was only found in A. salmonea. M: 100-bp ladder marker.
0.2 kb
Fig. 6. Relative sketch of the PCR products of Antrodia cinnamomea and A. salmonea amplified by the ML3 and ML4 primers. The front and rear ends of the 871-bp fragment matched to the 5’ and 3’ ends of the 2384-bp fragment, respectively.
Fig. 7. Phylogenetic inference based on the ML3/ML4 amplified fragments showing monophyletic cladogenesis of Antrodia cinnamomea and A. salmonea.
identification and sequencing, fragments of ML3/ML4 yielded from A. cinnamomea (0.9 kb) and A. salmonea (2.4 kb) were analyzed and showed that there was a difference be- tween these two with an extra insert in A.
salmonea (Figs. 6, 7). Thus a more-specific primer pair, Acl-F/Acl-R, of A. salmonea was then designed in this study. The PCR exami- nation showed that only A. salmonea yielded a specific 219-bp fragment (Fig. 5). A protein analysis was also carried out, and 2 open read- ing frames, encoding 61 and 196 amino acids, were found in the 1.5-kb fragment yielded from A. salmonea. These 2 proteins are likely homing endonucleases due to the LAGLI- DADG motif which has been related to intron movement. Similar structures were also report- ed in other fungi, such as Avarinvillea erecta, Trichoplax adhaerens, and Saccharomyces sp., and Scleractinian corals. Further research of these 2 possible genes and protein activi- ties may be probed in evolution study in the future. Obviously, ML3/ML4 can distinguish these 2 similar fungi, i.e., the Acl-F/Acl-R primer pair yielded a shorter fragment and took only 1 h for the PCR.Therefore, it is pos- sible to avoid professional identification tech- niques for A. cinnamomea and A. salmonea (Chang and Wang 2005). Moreover, it would be hard to culture and carry out morphological recognition, since most commercial products made of A. cinnamomea or A. salmonea are powders and capsules. In this study, we estab- lished a rapid and accurate identification tech- nique for A. cinnamomea and A. salmonea, which can be helpful when directly examining commercial products. For practical examina- tions, however, morphological observations of these fungi are still recommended as primer pairs of ML3/ML4 were used to distinguish A.
salmonea from A. cinnamomea. At the same time, the advanced specific PCR with Acl-F/
Acl-R yielded a 219-bp fragment which can
be used to determine whether A. salmonea has been added to commercial products.
LITERATURE CITED
Chang TT, Chou WN. 1995. Antrodia cin- nemomea sp. nov. on Cinnamomum kanehirai in Taiwan. Mycol Res 99:756-8.
Chang TT, Chou WN. 2004. Antrodia cinna- momea reconsidered and A. salmonea sp. nov.
on Cunninghamia konishii in Taiwan. Bot Bull Acad Sin 45:347-52.
Chang TT, Wang WR. 2005. Basidiomatal formation of Antrodia cinnamomea on nutrient agar media. Bot Bull Acad Sin 46:151-4.
Dujon B, Colleaux L, Jacquier A, Michel F, Monteilhet C. 1986. Mitochondrial introns as mobile genetic elements: the role of intron- encoded proteins. Basic Life Sci 40:5-27.
Gardes M, Bruns T. 1993. ITS primers with enhanced specificity for asidiomycetes--appli- cation to the identification of mycorrhizae and roots. Mol Evol 2:113-8.
Guha TK, Hausner G. 2014. A homing en- donuclease with a switch: characterization of a twintron encoded homing endonuclease. Fun- gal Genet Biol 65:57-68.
Hung TH, Wu ML, Su HJ. 1999. Develop- ment of a rapid method for the diagnosis of citrus greening disease using the polymerase chain reaction. J Phytopathol 147:599-604.
Kurokawa S, Yamasaki T, Komatsu T, Wa- tanabe KI, Ohama T. 2006. Degenerated rec- ognition property of a mitochondrial homing enzyme in the unicellular green alga Chlamy- domonas smithii. Plant Mol Biol 62:141-50.
Mullineux ST, Willows K, Hausner G. 2011.
Evolutionary dynamics of the mS952 intron: a novel mitochondrial group II intron encoding a LAGLIDADG homing endonuclease gene. J Mol Evol 72:433-49.
Sethuraman J, Okoli CV, Majer A, Corkery TL, Hausner G. 2008. The sporadic occur-
rence of a group I intron-like element in the mtDNA rnl gene of Ophiostoma novo-ulmi subsp. americana. Mycol Res 112:564-82.
Szczepanek T, Gora M, Monteilhet C, Wy- socka M, Lazowska J, Golik P. 2006. In vivo analysis of the relationships between the splic- ing and homing activities of a group I intron- encoded I-ScaI/bi2-maturase of Saccharomy- ces capensis produced in the yeast cytoplasm.
FEMS Yeast Res 6:823-35.
Szczepanek T, Jamoussi K, Lazowska J.
2000. Critical base substitutions that affect the splicing and/or homing activities of the group
I intron bi2 of yeast mitochondria. Mol Gen Genet 264:137-44.
White T, Bruns T, Lee S, Taylor J. 1990.
Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In:
PCR-protocols a guide to methods and ap- plications. Innis MA, Gelfand DH, Sninski JJ, White TJ, editors. San Diego, CA: Academic Press. p 315-22.
Yang J, Zimmerly S, Perlman PS, Lambow- itz AM. 1996. Efficient integration of an intron RNA into double-stranded DNA by reverse splicing. Nature 381:332-5.