國立臺灣大學生物資源暨農學院昆蟲學系 碩士論文
Department of Entomology
College of Bioresources and Agriculture National Taiwan University
Master Thesis
載卵壓力和聚集效應對於大頭金蠅產卵行為的影響 Egg load and gregarious effects on Chrysomya megacephala (Diptera: Calliphoridae) oviposition
郭允 Guo, Yun
指導教授:蕭旭峰博士 Advisor: Shiuh-Feng Shiao, Ph.D.
中華民國 107 年 6 月
June 2018
中文摘要
法醫昆蟲學上常利用屍體上出現的麗蠅幼蟲來推估人類或者動物的死後間 隔時間 (post-mortem interval, PMI)。產卵行為會直接影響了幼蟲接下來的競爭、
捕食、生長等,進而影響到死後間隔時間的推算。而在植食性和寄生性的昆蟲當 中,個體需要去平衡壽命上的限制和卵的時效,讓整個的生殖行為能夠有最大化 的效益。牠們通常會調整產卵行為和策略來因應本身數目有限的卵。在高載卵壓 力 (egg load) 的情況之下,可能會促使雌蟲增加搜尋的頻率,來尋找更多的寄主 或者產卵基質,或者讓牠們接受較低品質的寄主。而在麗蠅中,他們需要特定的 物質像是屍體來產下後代,所以載卵壓力可能會影響麗蠅的產卵行為。然而在進 行實驗設計之前,另一項的有關因子聚集效應 (gregarious effect) 也要納入考慮。
聚集效應是麗蠅的產卵行為可以藉由其他同類的存在而被觸發。本研究以大頭金 蠅 (Chrysomya megacephala (Fabricius)) 為材料,主要在探討載卵壓力和聚集效應 對於大頭金蠅的產卵行為的影響,實驗分成四個部分。首先從解剖得知正常麗蠅 的攜卵量,再針對不同共域雌蟲的數量測試是否聚集效應會影響平均產卵量,之 後給予連續產卵刺激以進行生殖週期測試。之後在去除聚集效應的影響下,將單 隻交配過的雌蟲給予連續性的產卵刺激,記錄日齡、產卵量,目的是以日齡作為 載卵壓力的量化指標,隨著時間的增加,載卵壓力隨著增大,來測試雌蟲行為反 應。而在生理攜卵量部分,得到平均每隻雌蟲的攜卵量為 263.4 ± 36.4 個。共域 族群數量的測試結果為平均產卵量會隨著數量增加而提高,在數量到達 5 隻以上 則沒有顯著差異。另外在連續產卵刺激的生殖週期測試,其結果存在著三個週期,
第一個週期平均單隻雌蟲產出 278.9 ± 48.1 個;第二個週期平均單隻雌蟲產出 258.9 ± 36.7 顆;第三個週期平均單隻雌蟲產出 92.7 ± 53.0 顆,產卵間隔平均 9.82 ± 1.03 天。單隻雌蟲產卵測試的結果則是一半以上的組別,會在持續的刺激 下超過 20 天以上才有產卵行為,後續進行檢查體內剩餘的攜卵量的結果顯示,不 論是否出現產卵行為的單隻雌蟲,留在體內的殘存卵量占了絕大部分。總結來說,
共域雌蟲的數量會影響產卵表現,而單隻雌蟲的結果有可能為載卵壓力的影響不 及聚集效應來的顯著,導致未產出的卵佔大多數。
關鍵詞:大頭金蠅、生殖週期、載卵壓力、法醫昆蟲學、產卵行為。
英文摘要
In forensic entomology, the age of necrophagous blow fly maggots that appear on corpse are used to estimate the post-mortem interval (PMI). The oviposition behavior will directly influence competition, predation, growth, etc., which affect PMI calculation. Insect parasitoids and herbivores need to balance the risk of egg and time limitation in order to maximize reproductive success. They adjust their oviposition behavior in response to their limited eggs. High egg load pressure will trigger female to search more intensively, and therefore make them encounter more hosts or medium;
or accept oviposition hosts of lower preference. In blow flies (Diptera: Calliphoridae), they need to find specific materials such as corpse to produce offspring, so the egg load will affect their oviposition behavior. However, another factor must to be taken into consideration, the gregarious effects. Gregarious effects, which means the blow fly oviposition can also be triggered by other companion individuals. In this study, we used Chrysomya megacephala (Fabricius) to measure egg load and gregarious effects.
This study was divided into four stages. The first stage was the dissecting of female flies to obtain the egg amount, the second stage involved determing if female population size will affect the average oviposition quantity, then third gave the flies continuous oviposition stimulus to find the reproductive cycle. On the last stage, after seaparating females into single units, continunous oviposition stimulus was also applied to measure the behavior of single females. Fly age and oviposition quantity were recorded to quantify egg load. Results obtained by dissection indicated the average egg amount in a single female was 263.4 ± 36.4. Results from second stage revealed that as the amount of female increases, the average numbers of eggs laid also increased, but there was no significant difference when the female number increases past five. Continuous oviposition stimulus revealed three individual reproductive
cycles. The first cycle produced 278.9 ± 48.1 eggs, the second cycle produced 258.9 ± 36.7 eggs, and the third cycle produced 92.7 ± 53.0 eggs, with a gap of 9.82 ± 1.03 days in between. In the final stage of the experiment, up to more than half of the groups would oviposit after 20 days of stimulus. After the experiement was over, dissection of the flies from this stage revealed that the amount of eggs laid is significantly lower than the residual eggs in the ovary. The conclusion from these results indicates number of female in a population affects oviposition behavior, and single female behavior is more heavily affected by gregarious effect than egg load, causing lower oviposition.
Key words: Chrysomya megacephala, reproductive cycle, egg load, forensic entomology, oviposition behavior.
目錄
中文摘要 ...i
英文摘要 ... iii
表目錄 ... vii
圖目錄 ... viii
壹、前言 ... 1
貳、往昔研究 ... 2
一、法醫昆蟲學 ... 2
二、麗蠅在法醫昆蟲學的重要性 ... 5
三、大頭金蠅生物學 ... 7
四、影響大頭金蠅的產卵因子 ... 7
五、試驗目的 ... 10
參、材料與方法 ... 11
一、受測麗蠅的飼養 ... 11
二、成熟雌蟲卵量測定 ... 11
三、試驗設置 ... 12
(一) 共域雌蟲數量對於產卵表現測試 ... 12
(二) 大頭金蠅生殖週期測試 ... 13
(三) 單隻雌蟲載卵壓力測試 ... 13
四、統計分析 ... 13
肆、結果 ... 15
一、成熟雌蟲卵量測定 ... 15
二、共域雌蟲數量對於產卵表現量測試 ... 15
三、大頭金蠅雌蟲的生殖週期測試 ... 16
四、單隻雌蠅的載卵壓力測試 ... 17
伍、討論 ... 18
一、成熟雌蟲卵量測定 ... 18
二、共域雌蟲數量對於產卵表現量測試 ... 18
三、大頭金蠅的生殖週期測試 ... 20
四、單隻雌蟲的載卵壓力測試 ... 20
陸、結論 ... 23
參考文獻 ... 24
表目錄
表一、大頭金蠅 (Chrysomya megacephala) 生殖週期產卵量之常態分布測定結 果 ... 28 表二、大頭金蠅 (Chrysomya megacephala) 不同共域雌蟲數量組別間產卵量差 異的統計分析結果 ... 29 表三、大頭金蠅 (Chrysomya megacephala) 單隻雌蟲載卵壓力測試結果 ... 30
圖目錄
圖一、大頭金蠅 (Chrysomya megacephala) 雌蟲腹部及卵巢圖 ... 31 圖二、大頭金蠅 (Chrysomya megacephala) 不同共域雌蟲數量的平均產卵量比 較 ... 32 圖三、大頭金蠅 (Chrysoma megacephala) 雌成蟲在 5 隻共域時之生殖週期產卵 組數累計圖 ... 33 圖四、大頭金蠅 (Chrysoma megacephala) 雌成蟲在 5 隻共域時之生殖週期產卵 量分布圖 ... 34 圖五、大頭金蠅 (Chrysomya megacephala) 生殖週期實驗中三個週期平均單隻 雌蟲產卵量之比較結果 ... 35 圖六、大頭金蠅 (Chrysomya megacephala) 生殖週期測試存活數量與日齡的對 照圖 ... 36 圖七、大頭金蠅 (Chrysoma megacephala) 雌成蟲在單雌載卵壓力測試出現產卵 行為組數總計圖 ... 37 圖八、大頭金蠅 (Chrysomya megacephala) 單隻雌蟲載卵壓力測試產卵量分布 圖 ... 38
壹、前言
法醫昆蟲學應用在各種不同層面上的問題,從比較嚴重的刑事案件到簡單的 某些倉儲中所出現的所引起的倉儲問題 (Hall, 2001)。而在以昆蟲或其他節肢動物 作為證據來處理刑事案件的狹義法醫昆蟲學中,科學家常利用食腐性的雙翅目蠅 類,特別是許多食腐性麗蠅 (blow fly),來估算人類或者其他動物的死後間隔時 間 (post-mortem interval, PMI),也就是自死亡發生到屍體被發現的這段時間 (Catts and Goff, 1992; Amendt et al., 2004)。目前科學家普遍相信麗蠅雌蟲會在死亡 發生後數分鐘之內抵達屍體並產卵於上面 (Amendt et al., 2004)。因此若能推估屍 體上的麗蠅幼蟲之發育日齡,則可以得知產卵的起始時間,而這樣的發育時間會 與死亡時間十分的接近,只有極短的差距。
然而,動物會因應環境資源的分布不同而有產卵偏好 (Ellis, 2008),在許多 的實際案例中,如溫度、族群數量、屍體有無被隱蔽等影響之下,會造成產卵時 間與死亡時間的不相近,若直接以幼蟲的發育時間去估算勢必會造成誤差。因此 探討影響麗蠅的產卵因子,找出可能會影響產卵的因子,並提出可能的校正資料,
是有其必要的。
在北台灣的中低海拔,人口眾多的地區所進行的調查結果顯示,麗蠅科 (Calliphoridae),金蠅屬 (Chrysomya) 的大頭金蠅 (C. megacephala (Fabricius)) 以 及同屬的紅顏金蠅 (C. rufifacies (Macquart)) 為食腐性麗蠅中最常見的種類 (Chang, 2003)。而同樣在南台灣的調查中,這兩種的麗蠅在當地也是最為優勢的 食腐性麗蠅種類 (Jian, 2004)。因此這兩種為值得研究的對象。而在大頭金蠅中,
對於影響產卵因子的研究中,除了低溫、濕度、或是屍體在被隱蔽的狀態下會有 所影響 (Amendt et al., 2004),另外在產卵基質、光線以及產卵基質的腐敗程度上 有一定的探討 (Singh and Bharti, 2001; Yang and Shiao, 2012; Liao, 2016)。
而除了上述提及的產卵因子以外,還有許多因子未被探討。在現有的結果之 下將可能的產卵因子加入研究及考慮,可以對於大頭金蠅本身的產卵偏好以及估
算資料的校正上有著更多的修正。
貳、往昔研究 一、法醫昆蟲學
法醫昆蟲學 (forensic entomology) 在昆蟲學中為相當獨特的領域。Keh (1985) 將法醫昆蟲學定義為以昆蟲及其他節肢動物之相關研究應用於法律相關 議題的學門。在這個定義之下,Lord 和 Stevenson (1986) 將法醫昆蟲學分成三個 部分,都市昆蟲學 (urban entomology)、儲物昆蟲學 (stored-product entomology)、
以及狹義的法醫昆蟲學 (medicocriminal entomology)。都市昆蟲學主要研究的議題 是在人類生活環境中的昆蟲問題,例如白蟻造成的木造建築或者家具的破壞、蠅 類造成的環境衛生問題等等。而儲物昆蟲學則是處理貨物或者倉儲食物中出現的 昆蟲、節肢動物或者是部分的殘肢所引起的法律問題。例如穀片中出現的蟲體碎 片、家用面紙上的蜘蛛等例子。
狹義的法醫昆蟲學,強調在於利用昆蟲或者其他節肢動物,以解決法律犯罪 的事件,通常為刑事之暴力犯罪案件,如凶殺案等 (Hall, 1990)。在犯罪調查中利 用食腐性昆蟲有以下的特點 (Catts and Goff, 1992):
(1) 在屍體暴露出來後,昆蟲通常能夠發現屍體並快速地抵達。
(2) 調查人員能夠透過檢查屍體上的昆蟲的生長狀態跟存活狀況,以及不同 的組成來了解屍體本身的狀態。例如麗蠅會尋找在屍體上的自然開口或傷口並產 卵在這樣開口上 (Bourel et al., 2003)。如果有些昆蟲出現在屍體上但是這些昆蟲 並不自然存在於該棲地,則可以推測這個屍體本身可能不是第一現場,可能是被 移動到此處來的 (Keh, 1985)。
(3) 因為屍體的出現代表的是一種稀少的資源,有著時間和質量上的限制 (Beaver, 1977),許多的昆蟲會想要爭奪此有限資源,作為食物或者是產卵的資源 (Thompson et al., 2013)。隨著時間的過去,屍體會因為腐敗而經歷過好幾個時期,
而在每個時期會藉由腐敗分解產生許多的化學物質,這些化學物質會吸引特定昆 蟲前來,所以屍體周圍的動物相是可以受到預測的。通常這樣的動物相會提供一 些視覺上的資訊,但是這些資訊並不一定會受到重視。
最早的法醫昆蟲學可以追朔到西元十三世紀中國南宋時期理宗和端宗年間,
宋慈所撰寫的洗冤錄中所記錄到利用蠅類會被血腥味吸引,進而偵破兇殺案件 (Catts and Goff, 1992)。到了十九世紀初,麗蠅會在屍體分解的早期出現並產卵的 現象有被科學家所記錄,但是並沒有將死亡時間跟麗蠅的出現加以連結。一直到 十九世紀後半葉,Yovanovich (1888) 與 Mégnin (1894) 才正式將屍體上的昆蟲相 的演替跟屍體腐敗時間的估算加以連結。
在狹義的法醫昆蟲學當中,最主要的討論範圍是利用昆蟲資料來推算死後估 計時間 (PMI),也就是從人或者動物死亡之後開始計算,到屍體被發現的時間 (Amendt et al., 2004)。估計死亡時間在法律案件上的調查上有著十分重要的意義,
正確的估計死亡時間可以與嫌犯的不在場證明相互對照,藉此來支持或者否定嫌 犯的不在場證明。昆蟲的種類多、棲地很廣,在死亡事件發生之後,往往最先抵 達的是昆蟲。加上前面所提的演替相及動物相可以隨著腐敗分解的情況可以預測,
只要能掌握昆蟲的生活史及相關的基本資料,就能夠利用這些資料進行死後時間 估計,而且能夠相當的精確 (Gennard, 2007)。
以昆蟲資料來推算 PMI 的方式主要可以分為兩種 (Catts and Goff, 1992)。在 屍體分解腐敗的早期,利用屍體上發育成長為成蟲的昆蟲物種的日齡來推算 PMI,
最常見的就是利用麗蠅或者肉蠅等雙翅目食腐性種類來進行短期的 PMI 估算。另 一種則是在屍體分解的中長期,利用昆蟲相的演替來進行比較長期的 PMI 估算。
在屍體分解的過程當中,生物、化學會隨著時間會有著很大的變化,根據不同的 學者對於腐敗時期的分法也不盡相同,從早期提出的八個時期到近代大部分的學 者都採用四到五個時期。這裡引用分為四個時期的腐敗過程 (Catts and Goff, 1992)。從一開始的新鮮期 (fresh stage) 開始,這段期間會經過許多的化學及生理
上的變化,但並不會有明顯的味道以及屍體本身並不會有明顯的腫脹發生,出現 在附近或屍體上的昆蟲通常只有少數的螞蟻和麗蠅。等到經過幾個小時之後,會 根據溫度以及環境條件的不同,而進入到膨脹期 (bloated stage),這時段屍體本身 會因為微生物的分解而開始產生氣味,通常會伴隨著氣體的產生而開始造成內部 的膨脹,加上許多的蠅類開始出現在旁邊。接下來進入腐敗期 (decayed stage),
會有著大量的腐生蠅類的幼蟲,這些幼蟲會從屍體內部鑽出,使膨脹的部分塌陷,
同時也會有部分的鞘翅目物種會作為捕食者來這裡捕食。等到進入最後一個時期 乾燥期 (dried stage),大部分的蠅類幼蟲會離開屍體而進入蛹期,只有少部分的 可能會留在上面,同時因為整個屍體的軟組織幾乎都被昆蟲及微生物消耗殆盡,
剩下皮毛、骨骼等的硬組織,所以會有些專門取食這些硬組織的鞘翅目昆蟲前來 取食,最後的階段通常會維持最久的時間,可以長達幾年。
而在此屍體分解過程中,會出現在屍體上及其周邊的昆蟲大致上可以分成四 類 (Catts and Goff, 1992; Hall and Haskell, 1995):
(1) 食腐性種類:許多的雙翅目昆蟲,特別是麗蠅科和肉蠅科 (Sarcophagidae) 中的種類為食腐類,另外鞘翅目中的埋葬蟲科 (Silphidae)、鰹節蟲科 (Dermestidae) 亦有某些種類為食腐性。這些種類的成蟲會取食屍體並在上面進行產卵,卵孵化 之後的幼蟲的生長會在屍體上,這類的昆蟲是用來估算 PMI 的主要依據。
(2) 捕食或者寄生種類:在屍體及周邊捕食或者寄生在上述的食腐性物種,
例如某些隱翅蟲科 (Staphylinidae),或者是某些閻魔蟲科 (Histerdidae) 的種類,
或者是膜翅目的寄生性蜂類等,此類的重要性是僅次於食腐性的物種。
(3) 雜食性種類:例如螞蟻、部分蜂類可能會同時取食食腐性的昆蟲,從卵 到成蟲都有可能是捕食的範圍,同時也會以屍體組織作為食物來源。如果這類的 昆蟲數量太過於龐大的時候,有可能會影響屍體的分解速率。
(4) 偶發性種類:此類的昆蟲或者是節肢動物,僅只把屍體作為生活棲地的 一部分來看待,如某些的蟎類。也有可能只是正好經過屍體附近的昆蟲,會出現
僅是偶然。
上述四種類型的昆蟲在不同時期造成屍體上的昆蟲相的演替可以做為長期 PMI 估算的基礎以及重要的參考資料,但是有關於短期的估算則是要從特定的物 種來進行。
二、麗蠅在法醫昆蟲學的重要性
如前面所提到的,短期的 PMI 估算中,雙翅目的蠅類扮演的角色十分的重 要,而在多數的法醫昆蟲學估算 PMI 的研究當中,食腐性的麗蠅為最重要的類群 (Byrd and Castner, 2001; Campobasso et al., 2001)。麗蠅在雙翅目中屬於短角亞目 (Brachycera),有縫組 (Schizophora),有瓣類 (Calyptratae),狂蠅總科 (Oestroidea) 的麗蠅科 (Calliphoridae) (Kano and Shinonaga, 1968; Marinho et al., 2012)。大部分 的麗蠅外觀具有金屬光澤。多數種類的雄蟲前額窄,而雌蟲則是較寬,從外觀來 看雄蟲的兩個複眼會緊貼在一起,為合眼式,雌蟲則是眼距較寬,為離眼式。幼 蟲的體色通常為白色或者黃色;終齡的幼蟲末節邊緣具有六個或者更多的錐狀突 起;腹部末端具後氣孔 (Kano and Shinonaga, 1968)。許多的麗蠅會受到屍體、排 泄物、組織傷口的引誘,除了在法醫昆蟲學上重要之外,亦是相當重要的分解者 和清除者。雙翅目除了麗蠅之外還有其他可能具有法醫昆蟲重要性的類群包括:
肉蠅科 (Sarcophagidae)、家蠅科 (Muscidae)、水虻科 (Stratiomyidae)、蚤蠅科 (Phoridae) (Byrd and Castner, 2001; Marchenko, 2001)。
由於麗蠅在通常在死亡事件發生之後極短的時間內就能夠到達並進行產卵 行 為 (Bourel et al., 2003),此間隔時間可能少於幾小時內甚至是幾分鐘內 (Amendt et al., 2004)。而在一定的溫度之下可以利用有效積溫法則 (law of effective accumulated temperature,可以用公式 DT = K 來表示,其中 D = 完整生 長發育所需時間;T = 完成該發育階段的平均溫度;K = 常數。但一般昆蟲發育 起始溫度皆在 0℃以上,在發育溫度起點以上的溫度才是對生長發育有效的溫度。
昆蟲有效發育的過程中所接受的總熱量應該為有效溫度的總和,因此將公式修正
為 D (T-C) = K;C = 發育起點溫度;T-C = 發育平均有效溫度),而每一種類 之麗蠅的發育所需時數為固定。例如常見的大頭金蠅在 26℃ 條件下,從卵、幼 蟲到化蛹、羽化為成蟲的時間約為 12 天 (Gabre et al., 2005)。在已建立發育時間 和麗蠅生活史基礎資料的前提之下,藉由屍體上採集到的幼蟲日齡,可以推估該 屍體的死亡時間。
基於上面的理由,死亡時間和麗蠅的產卵時間的間隔成為很重要的議題。通 常研究者認為麗蠅不會進行夜間產卵 (Amendt et al., 2008),所以在一般計算 PMI 的時候,必須考慮產卵的時間會不會與夜晚的時間重疊,若是有,則通常會認定 產卵時間是夜晚來臨前的傍晚。麗蠅在尋找產卵基質時,是以飛行的方式找尋產 卵基質,有研究顯示麗蠅在夜晚無法飛行,而主要的原因推測可能是氣溫 (Wooldridge et al., 2007)。而在歐洲的研究顯示,在正常的狀況之下,麗蠅不會在 夜間進行產卵,但是可能會在特殊的狀況之下,如較高溫的夜間等,誘發麗蠅的 產卵行為 (Amendt et al., 2008)。
以麗蠅幼蟲日齡進行 PMI 估算的問題,在於如何準確的估算幼蟲的日齡。
藉由實驗室內的飼養特定的種類,記錄不同溫度之下的生長曲線,此資料能夠用 來跟實際案例中採集的幼蟲做比較,求得 PMI 數值 (Anderson, 2000)。除了用幼 蟲的生長曲線來估算之外,也可以利用蠅卵來估算 PMI (Anderson, 2004)。甚至能 夠以羽化成蟲所遺留的蛹殼的碳水化物含量的變化量,會隨著時間改變,利用這 點來估算 PMI (Zhu et al., 2007)。
除了幼蟲本身的研究之外,環境因子也會造成 PMI 的誤差 (Benecke, 2001)。
例如,Gennard (2007) 研究美國佛羅里達州地區的屍體演替情況,發現即便相同 的地區,在不同的季節、甚至不同日期的麗蠅種類豐度、消長情況皆有差異,此 差異會造成屍體分解速度的不同,因此在估算 PMI 時除了地區之外,還要考慮 季節等因素。Joy et al. (2006) 於美國西維吉尼亞州之調查發現,屍體上的蛆團溫 度與周遭的氣溫有高度的相關,在陽光直射的情況之下,蛆團的溫度會顯著高於
在有遮陰者。光照、蛆團溫度等因素皆會影響屍體的分解速率,進而影響 PMI 的 估算。
三、大頭金蠅生物學
大頭金蠅 (Chrysoma megacephala (Fabricius)),屬於雙翅目,麗蠅科,金蠅 亞科 (Chrysomyinae),金蠅屬 (Chrysoma) (Wells et al., 1999)。成蟲體長 8.0~10.0 mm,身體呈藍色或者綠色金屬光澤,複眼大而呈現紅色,頰區有橘黃色的毛 (Kano and Shinonaga, 1968; Byrd and Castner, 2001)。原產於東洋區及澳洲區,現 在則是普遍分布在非洲、美洲、亞洲,為世界性廣泛分布之具法醫重要性麗蠅 (Sukontason et al., 2008)。大頭金蠅在野外的成蟲壽命約一個月,但是實驗室飼養 的成蟲往往可以存活到兩個月,甚至更久。幼蟲會生長經過三個齡期之後進入後 食期,在後食期的階段,幼蟲會離開原本取食的區域,找尋適合的地方化蛹。當 卵被產下之後,需要經過約一天的時間來孵化,這時間有可能從幾小時到一天但 不會超過一天。在溫度 27℃、相對溼度 60~70% 的條件下,從卵孵化算起,完整 的幼蟲期需要 5.4 天,蛹期則是 5.3 天。羽化之後的成蟲需要 6.6 天來達到性成 熟,並且有能力進行產卵,而大頭金蠅雌蟲一次可以產出 223.7 顆卵 (Gabre et al., 2005)。
大頭金蠅在台灣是優勢食腐性物種,在有屍體暴露的情況之下,很容易吸引 牠的前來。麗蠅利用嗅覺的感受器來進行長距離的尋找,當到達近距離的時候,
便會切換視覺來確認屍體或者產卵基質的狀態 (Gomes et al., 2007)。自然的開口 如口、鼻、耳,或者傷口都會成為麗蠅產卵的首要目標。在正常狀況下,自然開 口通常是唯一可以讓剛孵化的一齡幼蟲進入內部軟組織的地方。當雌蟲找尋到適 當的自然開口,他們通常會進入口內或者在口的附近產下卵,之後便等待卵的孵 化,開始整個分解的消長。
四、影響大頭金蠅的產卵因子
許多的因子會影響大頭金蠅的產卵,除了前面提及的環境因子會影響屍體的 分解速率外,其實也會直接影響大頭金蠅的產卵行為 (Amendt et al., 2004)。除了 環境因子之外,產卵基質本身也佔有很大的比重。Yang and Shiao (2012) 利用不 同條件的豬肝進行實驗,來對於大頭金蠅和紅顏金蠅進行產卵偏好性的實驗,而 實驗的結果指出產卵行為會受到基質本身的大小,以及同種或者異種間幼蟲的存 在會影響產卵的偏好。同時這次的結果也指出視覺在找尋產卵基質的過程當中,
扮演相當重要的角色。
另外的研究則注重在基質本身的狀態是否會對於麗蠅產生影響。大頭金蠅會 容易被中大型的屍體所吸引,同時如果基質本身是處於低水分的狀態,則會阻止 大頭金蠅的產卵行為 (Erzinçlioğlu, 1996)。濕度的條件本身也會是一個相當重要 的影響要素,要是麗蠅產卵在一個乾燥的基質上,卵便會跟著容易一起脫水,導 致卵的損害,最終難以孵化 (Davies, 1950)。然而大部分的麗蠅通常能夠辨認基質 的乾燥程度來決定產卵與否。
基質的腐敗程度同時也會受到影響,之前的研究顯示超過 8 天以上的產卵 基質與新鮮的產卵基質,麗蠅幾乎不會選擇腐敗程度較高的產卵基質 (George et al., 2012)。而在測試腐敗 4 天與新鮮的產卵基質的偏好性測試,結果顯示兩者之
間並沒有顯著的差異,可以認定在腐敗程度不超過 4 天的情況下,不會對於麗蠅 的偏好性有顯著的影響 (Liao, 2016)。
另外在眾多野外研究當中,會去探討所謂的夜間產卵問題。在眾多的野外調 查當中,會不會有夜間產卵的問題一直是沒有一個肯定的答案,有些物種的麗蠅 可能會在夜間丟出的屍體上進行產卵,然而有些物種則是沒有這樣的行為(Singh and Bharti, 2001; Baldridge et al., 2006; Barnes et al., 2015; Williams et al., 2017),而 這樣的結果只能推測可能會跟當時實驗進行的野外環境的溫度及附近的光源是否 存在有關。
然而以麗蠅的產卵情況及之前的研究紀錄中,皆顯示牠們一旦找到產卵基質
之後,能夠在短時間內產下卵。以食腐性的昆蟲而言,屍體有如突然出現的資源,
而且皆未被占用 (Beaver, 1977),雌蠅必須要能夠快速產卵的情況之下,在抵達之 前便是攜帶的成熟的卵,如果攜帶的卵未能夠找到適當的產卵基質給產下,對個 體而言會產生生殖上的壓力,稱為載卵壓力 (egg load)。載卵壓力一詞主要是在 探討關於植食性和寄生性昆蟲的生殖策略和選擇上, 在前人研究所提到的 整理中,這樣需要專一性產卵物質的昆蟲中,如果 在找不到適合產卵的情 況之下,成熟的卵會給予個體的壓力稱為載卵壓力。這樣的壓力會在卵熟 成之後隨著時間的增加而逐漸的提高,而這樣隨著時間的提高會導致一些 行為上的改變,主要可以分為幾種可能,第一種是會增加雌蟲去尋找寄 主 或者是產卵基質的 反 應跟行動能力,藉由主動搜尋來提高接觸的機率,以 便能夠把卵給產出。第二種是會接受跟一般的寄主或產卵基質相比較,品 質較低的寄主或基質。這樣是增加接受度來達到把卵產出的目的。第三種 則是在接觸到合適的產卵基質跟寄主之後,會提升產卵的速度以及產卵的 總量 (Minkenberg et al., 1992; Ellers et al., 2000)。雖然這是在植食性跟寄 生性的昆蟲上所得到的結果,但是麗蠅在需要特定的產卵基質的這個條件 上來看,可以視為比較接近寄生性的性質,在無法找到適合的產卵基質的 情況之下, 麗蠅可能有著不小的載卵壓力。 另外藉由前面對於載卵壓力研 究,都是以產卵表現作為載卵壓力的大小,在不同的日齡會有不同的 產卵 表現量 (Dieckhoff et al., 2014)。因此,如果要測定載卵壓力的存在與否,
應該能夠藉由產卵表現以及產卵量來判斷。而在麗蠅的產卵行為的研究中,
通 常 麗 蠅 雌 蟲 在 找 到 合 適 的 基 質 之 後 , 一 次 將 體 內 全 部 的 卵 進 行 產 出 (Browne, 1993),所以綜合以上的結果,可以藉由量測產卵量來決定
在之前實驗中發現到, 麗蠅的產卵會因為複數個體的存在而被觸發,
這樣的效性稱為聚集產卵或者聚集效應 (gregarious effect),同時單隻雌蟲 在提供產卵基質的情況之下,即便是新鮮的產卵基質依舊不會有產卵行為
(Yang, 2014)。然而這樣的結果是否只有在單雌的 成蟲上會出現,在從單隻 到十幾隻的連續數量中有沒有出現產卵行為的表現和產卵量的差 異在當時 的結果中並沒有提及。
五、試驗目的
結合了上述的因子,為了瞭解載卵壓力是否會影響麗蠅的產卵行為,同時比 較在產卵基質的存在的條件之下,麗蠅的族群數量、載卵壓力在產卵行為上哪個 佔有比較重要的影響力,以及建立大頭金蠅的生殖週期資料。本研究以大頭金蠅 (Chrysomya megacephala) 為材料,首先收集大頭金蠅的本身生理狀態的攜卵量資 料,將發育完整且經過交尾的雌蟲進行卵巢解剖與計數卵數,希望能夠了解雌蟲 本身的生理條件與正常的攜卵量。之後再做不同共域數量的產卵表現量,重複之 前的試驗方式但是將共域數量連續,藉此能找出共域數量不同是否能夠刺激產卵 表現量。再來給予連續產卵刺激來收集麗蠅的生殖週期資料,將大頭金蠅多久能 夠進行一次產卵以及產卵量給予紀錄,之後以此作為對照。藉此來比較單隻雌蟲 在面對逐漸提高的日齡但是產卵基質依舊存在的情況之下,會如何選擇。將希望 能夠透過這類資料的收集,提出一個大頭金蠅在面對族群數量影響跟載卵壓力存 在的狀況時,這兩個因子哪個是比較重要的影響力,同時提供一些有關於 PMI 估 算的修正。
參、材料與方法 一、受測麗蠅的飼養
材料為從台灣大學農業昆蟲館附近經由陷阱捕捉且在實驗室內已有長時間 累代飼養的大頭金蠅 (Chrysoma megacephala),在直立式恆溫電氣生長箱下,控 制光週期為 L : D = 12 : 12,溫度 27 ± 0.5℃ 下進行累代飼養,以進行試驗。成 蟲飼養在 30 cm × 30 cm × 30 cm 四面具有尼龍紗網的塑膠飼養箱內,內部放置 清水和由紅糖和奶粉以 1:1 混合而成的飼料供麗蠅自由取食。由於麗蠅的雌蟲在 羽化之後需要補充額外的蛋白質等營養才能達到性成熟並且可以交尾,交尾行為 會出現在性成熟之後 (Norris, 1965)。在羽化後的一天到兩天之內給予新鮮的豬肝 作為成蟲的蛋白質補充,羽化過後的麗蠅約 7 天達到性成熟的階段,可以進行產 卵。給予性成熟的麗蠅新鮮的豬肝作為產卵基質,會選擇豬肝是因為該物質是對 麗蠅最具吸引力的物質 (Bunchu et al., 2008)。待雌蟲產卵於豬肝之後,將卵移到 250 cc 容器中,利用 Hong (1995) 開發的大量飼養麗蠅的方法,以魚粉:酵母粉:
洋菜粉:水 = 25 : 10 : 1 : 150 比例混合煮沸凝固的培養基中飼養。幼蟲會放在人 工培養基持續飼養直到幼蟲進入後食期 (post-feeding),套入有鋪木屑的套袋以利 幼蟲化蛹。將蛹收入飼養箱內,等待羽化。重複此步驟,以維持族群。受測的麗 蠅在羽化後 0~2 天為剛羽化時期並給予蛋白質補充,2~6 天為等待完整卵巢發育 和性成熟,實驗的麗蠅皆在羽化 7 天放入實驗裝置,等待靜置一天之後才會進行 實驗。
二、成熟雌蟲卵量測定
為了確定單一隻雌蟲在一次卵成熟之後能夠產生多少的卵,在累代飼養的族 群當中,隨機挑選羽化後 7 天至 10 天內,挑選性成熟、有交配並懷孕的雌蟲進行 解剖並計算成熟的卵量。麗蠅捕捉後將其放入 - 20℃ 的冰箱中放置 5 分鐘。當 確定雌蟲失去活動能力之後,使用解剖工具將腹部完整的剪下,放入滾水煮沸,
目的是為了讓整個卵巢被煮熟固定,並讓每一顆卵能夠輕易地辨別。之後放在解 剖顯微鏡底下,紀錄完整的樣子之後進行攜卵量的計算。如果在計算的過程中有 發現煮熟的狀況並不完全,會將已取出的卵巢再進行第二次的煮滾,讓卵的顏色 呈現白色表示蛋白已經被煮熟而且是處於固定的狀態。將煮熟過後的卵巢先進行 照相及完整大小的測量,之後再利用鑷子將整個的卵巢逐步的分離開來讓卵能夠 盡量全部分開,卵攤平在整個平面上,再進行卵量的計算,計算一隻雌蟲體內能 夠攜帶多少的卵量,以及正常發育的卵大小。將此生理攜卵量的結果和後面的實 驗結果作為對照用之外,同時了解麗蠅雌蟲一次卵巢的最大發育量。此測定會重 複 15 次,共使用 15 隻雌蟲。
三、試驗設置
產卵行為的試驗設置在 30 cm × 30 cm × 30 cm 四個面具有尼龍紗網的塑 膠飼養箱內,將飼養的麗蠅隨機挑選性成熟且尚未產過的卵的雌蠅進行以下的試 驗。飼養箱內部放置清水和紅糖供麗蠅自由取食,水放在 250 cc 塑膠杯並利用 吸水紙來讓麗蠅取用。生長箱內的光週期為 L : D = 12 : 12,溫度 27 ± 0.5℃。提 供新鮮豬肝作為產卵基質,產卵基質固定放在飼養箱的正中央。豬肝為當天由傳 統市場所購買的新鮮豬肝,豬肝裝在 250 cc 塑膠杯並且在底部會先放半張摺疊好 的吸水紙,再將產卵基質放入杯子內,避免基質的水分過多而導致雌蟲淹死的情 況,紙張會被產卵基質完全的覆蓋住,並不會讓雌蟲接觸到紙張。塑膠杯為避免 上面有氣味的殘留影響雌蠅的選擇,故不會重複使用。
(一) 共域雌蟲數量對於產卵表現測試
日齡 7 天的雌蟲將分別各以 1~10 隻為一組放入試驗設置之中,提供產卵基 質豬 100 g。試驗總共進行 48 小時,每 24 小時記錄一次上面的產卵量,並將產卵 基質做更換,將產卵基質上的卵塊以軟鑷子和刷毛取下,並將卵塊分離。計算不 同雌蟲數量組別在產卵基質上面的產卵數,之後經由統計方法推論共域雌蟲數量 對於產卵表現的影響。2~10 隻為一組的雌蟲各進行 10 組重複,單隻雌蟲進行 20
次重複,共使用 540 隻雌蠅。
(二) 大頭金蠅生殖週期測試
將日齡 7 天的雌蟲 5 隻為一組放入試驗設置之中,從日齡 8 天起,每天給予 產卵基質 100 g,每 24 小時記錄一次上面的產卵量,並同時記錄成蟲的存活數量,
為了避免卵或者是產卵基質的腐敗會影響雌蟲的選擇,產卵基質會每天進行更換。
因為在前測以及觀察中,發現壽命約略 40 天左右,因此決定紀錄直到日齡 42 天 結束,之後進行總產卵量、平均產卵量、產卵間隔的天數的計算和統計分析。實 驗進行 15 組重複,共使用 75 隻雌蠅。
(三) 單隻雌蟲載卵壓力測試
將實驗室飼養的麗蠅隨機單隻雌蟲當作一組放入試驗設置,從日齡 8 天開始,
每天給予產卵基質 100 g,每 24 小時記錄一次上面的產卵量,並同時記錄單隻雌 蟲的存活狀況。同樣為避免之前的產卵或者產卵基質的腐敗會影響雌蟲的選擇,
基質會在有產卵時進行更換,若沒有則不會讓產卵基質超過 3 天進行更換。紀錄 從日齡第 8 天一直持續到 42 天結束。在實驗結束時會將雌蟲本身保存下來,進行 解剖並計算有多少的卵在雌蟲體內,抑或是卵被雌蟲給吸收走等可能的狀況進行 紀錄。之後進行卵量、產卵間隔天數、第一次產出的日齡計算和分析。實驗進行 20 組重複,共使用 20 隻雌蠅。
四、統計分析
統計分析是採用軟體 PAST Version 3.0 (Hammer et al., 2001)。在生理攜卵量 的測定中,計算的卵數目會用常態分布來檢定,並計算平均數和標準差。共域雌 蟲數量對於產卵表現測試實驗中,利用 Kruskal-Wallis test 和 Dunn's post hoc 來 分析組間表現量的差異。而在生殖週期的結果當中,每日收集的卵量會先進行平 均,之後找出產卵間隔,有的話會計算日期的平均及標準差,再來是有無規律性 的分布,如果有則會按照產卵平均跟日期來區分週期,計算一個週期的平均產卵 量,之後進行週期之間的比較。實驗計算的卵量會經過平均然後先進行常態分佈
檢定,如果全部的組別結果都通過常態分佈檢定,則會使用 ANOVA 進行組間的 差異比較分析。反之,如果有任何一組的卵量資料無法通過常態分布檢定,則會 繼續使用 Kruskal-Wallis test 和 Dunn's post hoc 來分析。
肆、結果 一、成熟雌蟲卵量測定
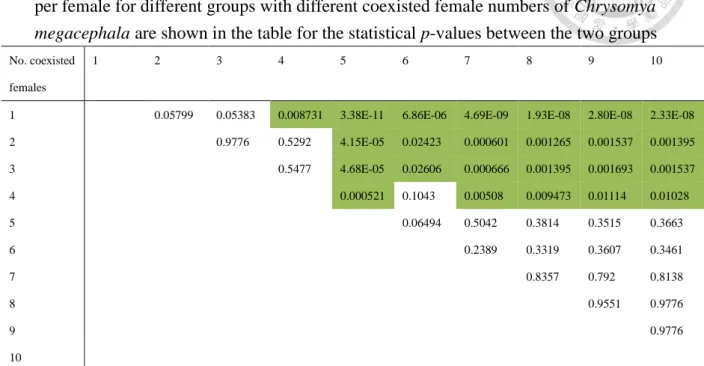
在此測定中,將解剖過後的卵巢進行影像紀錄,接下來再進行卵數計量。在 對照煮過與未煮過的樣本,發現未煮過的不僅在將卵進行單顆分離的過程中不易 分離,容易造成卵本身的破裂。相較於煮過在肉眼辨識下就可以清楚的觀察到卵 巢以及單顆卵之間的界限,雌蟲的腹部長度約 5 mm,而完整發育的卵巢是兩側 對稱且呈現圓球形。完整發育的卵巢可佔整體腹部的一半以上的體積 (圖一 A)。
15 隻雌蠅的總攜卵數為 3951 顆,平均每隻雌蟲的攜卵量為 263.4 ± 36.4 顆,經過 常態分布檢定後不是常態分佈 (Shapiro test : W = 0.1612 , p > 0.05),之後進行與生 殖週期與單雌的統計分析採用無母數的 Kruskal-Wallis test。而這樣的結果可以提 供後面實驗上的平均產卵量以及最大產卵上限來作為對照。
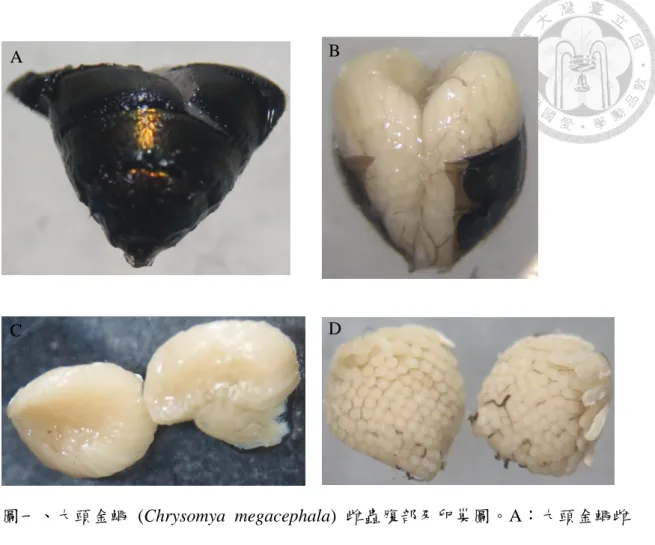
二、共域雌蟲數量對於產卵表現量測試
在族群數量對於產卵表現量的實驗中,單隻雌蟲的部分是沒有收集到任何的 卵 (N = 20)。在 2~10 隻的產卵量結果分別為 2 隻雌蟲組別平均產下 52.9 ± 21.2 顆;
3 隻雌蟲組別平均產下 50.9 ± 27.4 顆;4 隻雌蟲組別平均產下 70.5 ± 20.1 顆;5 隻 雌蟲組別平均產下 111.4 ± 11.7 顆;6 隻雌蟲組別平均產下 97.5 ± 6.2 顆;7 隻雌蟲 組別平均產下 103.2 ± 19.2 顆;8 隻雌蟲組別平均產下 103.2 ± 4.7 顆;9 隻雌蟲組 別平均產下 103.1 ± 8.1 顆;10 隻雌蟲組別平均產下 104 ± 14.1 顆 (圖二)。經統 計分析之後,平均產卵量在 5~10 隻的組別沒有顯著的差異,表示在雌蟲數量來 到 5 隻以上時,增加雌蟲數量並不會讓平均產卵量再往上提高,組跟組之間沒有 顯著差異。而在 2~4 隻組別的結果則是有產出但是產卵量與 5 隻以上有差別,表 示雖然有產卵行為但是在這樣的雌蟲數量條件下平均產卵量不足。而單隻的產卵 量的結果雖然是沒有產出,但在統計分析上顯著於 4~10 隻的組別但是卻與 2~3 隻的組別沒有顯著差異,表示這樣的 2~3 隻平均產卵量在結果上來看有產出但是
產卵量不足。就整體結果而言,平均產卵量的表現會隨著雌蟲數量的增加而有望 上增加的趨勢,來到 5 隻為一組之後會達到平穩而沒有再往上增加 (表二)。
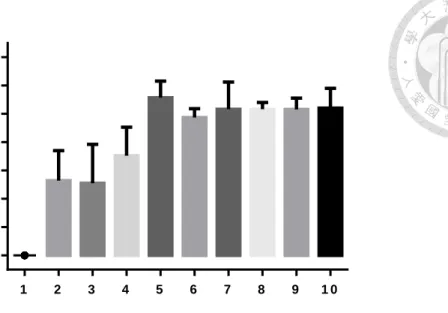
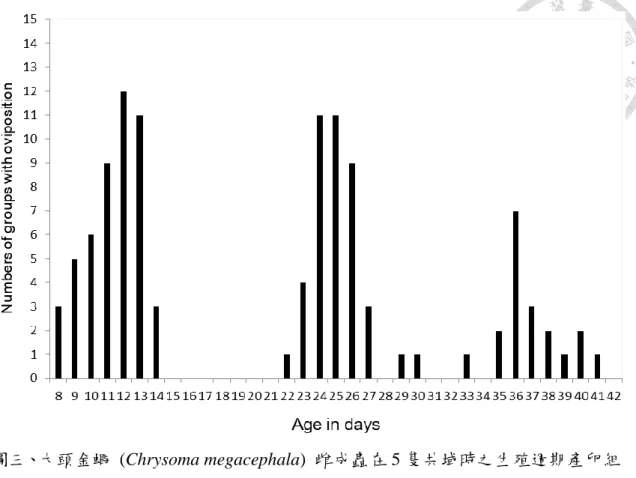
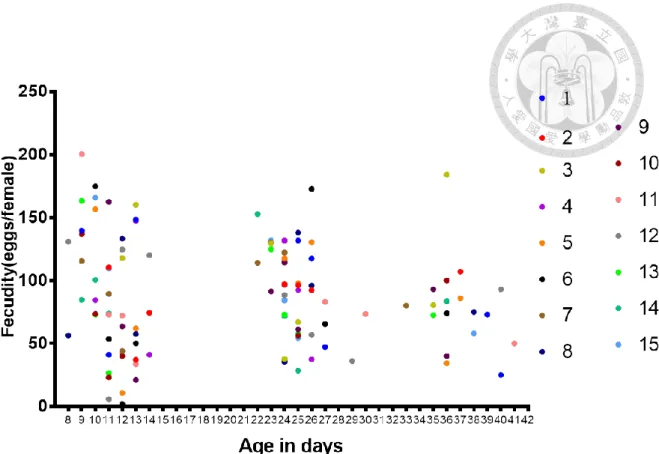
三、大頭金蠅雌蟲的生殖週期測試
在大頭金蠅的生殖週期實驗中,先將產卵高峰找出來確定週期的出現日齡 (圖三、圖四),在 15 組的累積中,有產卵行為出現的日齡有呈現集中在特定一個 區段的日齡,並且區段跟區段之間有明顯的間隔出現。將產卵行為密集出現的區 段視為一個週期的話,可以發現出現三次的產卵週期,第一個產卵週期出現在日 齡 8~14 天,第二個產卵週期出現在日齡 21~27 天,第三個產卵週期出現在日齡 35~41 天。在最後一次產卵的日齡到下次亦有產卵行為出現的日齡視為最小間隔,
再去計算最小間隔,最小間隔為 9.82 ± 1.03 天。代表著這些麗蠅能夠產出一代跟 一代的間隔時間大約 10 天,而這段時間即便有產卵基質的存在,但雌蟲並沒有產 卵行為的表現。
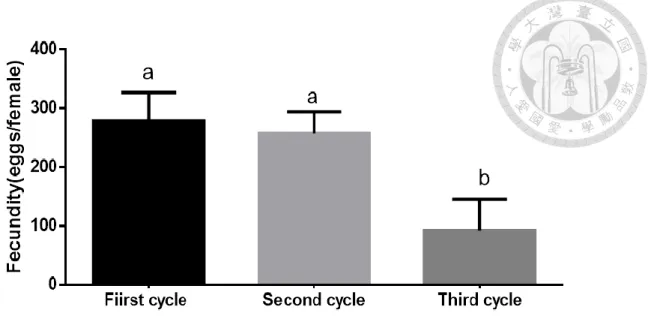
而將每一組的三個週期所產出的卵量進行平均,分別為第一個週期每組平均 產出 278.9 ± 48.1 顆;第二個週期每組平均產出 258.9 ± 36.7 顆;第三個週期每 組平均產出 92.7 ± 53.0 顆。先經過常態分佈檢定,第一週期的平均產卵量 (Shapiro test : W = 0.009636, p < 0.05) 和第三週期的平均產卵量 (Shapiro test : W = 0.01648, p < 0.05) 是常態分佈而第二週期不是 (Shapiro test : W = 0.5199, p > 0.05) (表一),所以在後面比較三個週期之間有差異時,使用 Kruskal-Wallis test 和
D
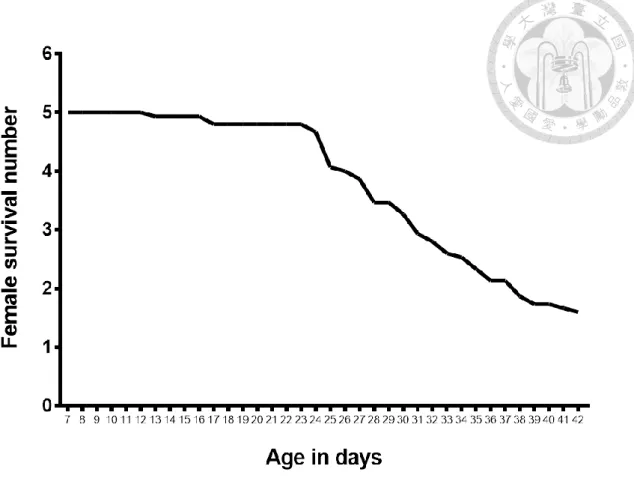
unn's post hoc 進行統計分析。而結果顯示,第一和第二週期之間平均產卵量沒有顯著 差異,第三週期和第一以及第二週期有顯著差異,代表在連續給予產卵基質的情 況之下,在第一次及第二次的產卵是能夠表現正常的產卵量,而到了第三個週期 時,即便有產卵行為的產生,但是產卵量卻不足 (圖五)。而整個實驗下來平均每 組的總產卵量為 630.9 ± 65.0 顆 (Shapiro test : W = 0.2383, p > 0.05),可以視為雌 蟲在整個實驗中的總共生殖能力。至於族群存活天數的結果,存活的天數從羽化 後作為日齡開始起點算起,整個平均存活天數為 33.2 ± 8.3 天,而整體存活數量與日齡的變化則是在日齡 26 天的時候平均會有 4 隻還存活的狀態,而在日齡 31 天的時候平均會有低於 3 隻還存活的狀態,最後在日齡 37 天的時候存活平均數量 會低於 2 隻 (圖六)。這樣存活的結果可以代表在進到第三個週期的時候,就已經 會有不少的雌蟲死亡,而且會在第三個週期的當中,雌蟲的數量會持續降低。
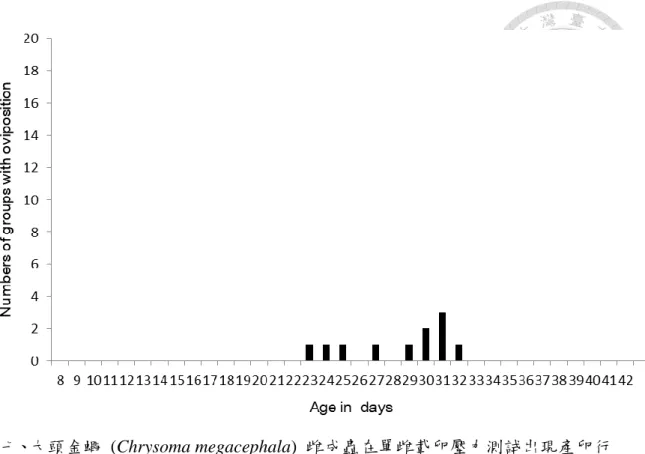
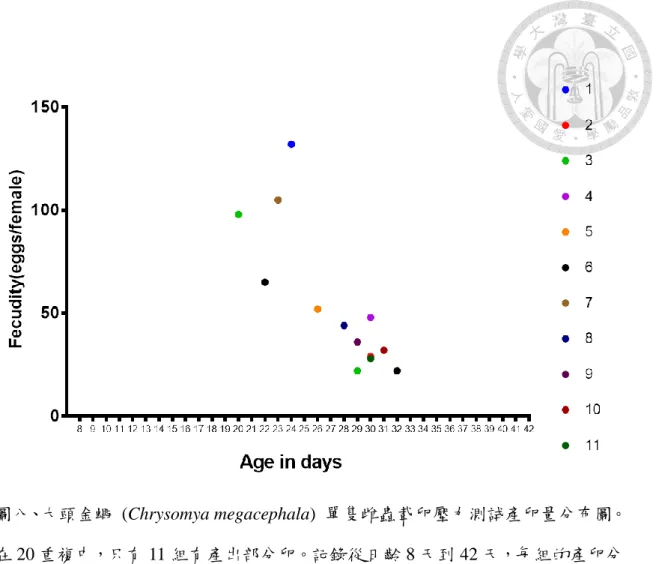
四、單隻雌蠅的載卵壓力測試
在單隻載卵壓力測試的結果當中,在 20 組當中 11 組的單隻雌蟲在隨著日齡 的增加之下有產卵行為的產生,另外 9 組雌蟲,在到實驗結束的時間點 (日齡 42 天) 時並沒有產卵行為的產生 (表三)。將有產卵組別進行初始產卵的日齡紀錄並 平均,11 組當中有初始產卵的平均日齡是 27.5 ± 3.0 天 (圖七),而在正常的生殖 週期中正常的第一次產卵天數為 8~14 天。有產出 11 組的平均產卵量為 67.4 ± 38.8 顆 ( 圖 八 ) 與 生 殖 週 期 中 的 第 一 週 期 進 行 統 計 分 析 , 發 現 有 顯 著 差 異 (Kruskal-Wallis test: p = 1.854E-05, p < 0.05),代表在即便單雌有產出卵的情況之下,
產卵量與正常生殖週期的相比是較少的。而將 11 組有產卵的組別的平均產卵量與 9 組未產卵的組別進行產卵量的統計分析,發現是有顯著差異的 (Kruskal-Wallis test: 0.0001601, p < 0.05),表示在單雌產出與不產出的是有差異的。
但是將 20 組全部的雌蟲全部進行檢查體內剩餘的攜卵量,將整個 20 組整體 每一組的產出與未產出的產卵量進行統計分析,結果產出的產卵量與未產出的存 留量結果呈現顯著差異 (Kruskal-Wallis test: 0.0000001214, p < 0.05),這表示在單 雌且產卵基質持續存在的情況之下,有產出的組別即便有產出部分的卵,但是大 部分的卵還是選擇留存在體內。而沒有產出的組別中,大部分的卵並沒有產出,
而且這些卵也在雌蟲體內留存到了實驗結束的日齡 42 天。
伍、討論 一、成熟雌蟲卵量測定
在此測定的結果中,利用 Harterreiten-Souza and Pujol-Luz (2012) 中提出了 有關於麗蠅雌蟲的卵巢到的結構的結果中,成功的將卵給取出並計算數量。在 Chaiwong et al. (2012) 的研究結果當中,認為雌蟲的卵巢發育需要約 10 天,但在 Gabre et al. (2005) 提出的結果中提到最短的發育天數是 6.6 天。儘管兩者之間的 發育所需天數有差異,但是測定中解剖的雌蟲結果來看,並沒有出現卵巢發育不 足的情況。另外在 Barton (2001) 提及卵巢發育的狀況和蛋白質以及含氮物質的攝 取有著正相關。Browne (1993) 提及麗蠅的卵巢是屬於同時間一起成熟的類型,
而這樣一起成熟卵巢期產卵行為也會傾向於一次性的將卵全部產出,而在沒有產 出或者產完之前不會進行再次發育。結合上述的結果來推論,麗蠅雌蟲在正常況 狀下能夠產出的卵量上限應為生理一次發育的上限,也就是在整批卵在雌蟲沒有 全部產出或者吸收完畢之前,不會進行下一次的發育。這樣的結果可以用在後面 的實驗結果中,作為這樣的產卵表現量是否就是就是全部生理所有的攜卵量,可 以判斷雌蟲的產卵是否為全部產出。
二、共域雌蟲數量對於產卵表現量測試
在族群數量對於產卵表現的結果當中,顯示在單雌的時候沒有產卵的結果顯 著的高於 4~10 隻為一組的組別,但在與 2~3 隻為一組的結果即便在單隻雌蟲沒 有產下任何卵的情況之下,在統計上沒有顯著差異。而 2~3 隻為一組的表現量與 5~10 隻為一組的表現量有著顯著的差異。而在 4 隻為一組的結果雖然和 5、7、8、
9、10 隻為一組的結果有顯著差異,但是與 6 隻為一組卻沒有顯著差異。綜合以 上的相互比較結果,可以得出在單隻雌蟲的情況之下,在日齡 8~12 天的雌蟲儘 管給予了合適的產卵基質,但 48 小時之內都沒有進行產卵。而隨著雌蟲數量的增 加,會增加雌蟲產卵的意願,單以產卵量來看,只要有超過一隻以上的同伴,就
能夠刺激雌蟲在有正常產卵基質的情況之下產卵,可是產卵量的多少則是會在特 定往上增加。以結果來說,存在著 5 隻以上的雌蟲的時候,給予產卵基質會讓平 均產卵表現量沒有顯著差異,即能夠在固定的產卵基質下,有相同的表現量。這 結果與 Yang (2014) 在聚集產卵實驗中的結果相似。當時的研究指出在群聚產卵 的實驗當中,雌蟲的數目會影響麗蠅的產卵意願,甚至會更進一步影響產卵的平 均量,稱為聚集效應。而當時所提出的推論指出由於這樣的效應會影響到子代的 數量及存活生長,進而影響到種內與種間的競爭。另外從麗蠅如何尋找產卵基 質來看,在 以往的研究來說,認為是利用嗅覺尋找 產卵基質,在近距離的 時候視覺可能也有影響 (Wall and Fisher, 2001)。在之後的研究中,麗蠅尋 找適當的產卵基質可能是依賴嗅覺進行,但是當要決定降落位置時,視覺 才是最重要的因子 (Gomes et al., 2007)。麗蠅會利用視覺來尋找合適的產 卵基質,並且會確認產卵基質上有沒有其他種類或者是同種類的幼蟲已存 在在上面,進而對於不同處理的產卵基質 做出不同的產卵偏好行為 (Yang, 2014)。結合上述前人 的研究結果,麗蠅對於產卵基質的存在之外, 會考量 整體環境的狀況。
而在另外一篇研究提出的推論是大頭金蠅的幼蟲本身需要藉由一定密度的 存在來產生蛆團熱,藉此增加生長效率以及取食效率 (Beaver, 1977)。因為麗蠅幼 蟲本身的活動範圍除了在終齡停止進食到化蛹前會增加其活動範圍之外,從卵孵 化開始的時期就幾乎只會在取食範圍附近活動,所以麗蠅雌蟲的選擇,會直接影 響幼蟲的數量及生長。所以對於雌蟲來說,聚集在一起產卵,除了可以增加幼蟲 數量進而增加取食效率之外,數量多的幼蟲可以藉由蛆團熱來加速幼蟲的生長,
對於種間的競爭更加有利。然而藉由前面生理攜卵實驗與數量的表現量相互比較,
會發現表現量跟生理的攜卵量有一段不小的差距,換句話說,雌蟲有可能沒有把 全部的卵生完,對此 Shiao and Yeh (2008) 的研究中指出,幼蟲的密度太高,
反而會讓幼蟲的存活率下降,這樣或許可以解釋即便數量不斷提高,但是
平均產卵量卻沒有達到生理攜卵量的上限。
三、大頭金蠅的生殖週期測試
在生殖週期的結果中,發現麗蠅在族群數量充足的情況之下,連續提供適合 的產卵基質,會讓產卵行為呈現出週期性。而週期與週期之間的間隔符合先前的 研究,正常的雌蟲所需要的卵巢發育時間約為 8 天 (Chaiwong et al., 2012)。然而 這當中的週期並不是僅有一天進行產卵行為,在結果中,產卵行為在一個週期中 會持續 3~5 天,呈現連續性的將卵釋放出來。針對這點,可能是如果前面實驗設 置的問題,產卵基質的大小不足以讓雌蟲把全部的卵給產出,但是由於是連續性 的給予並更換產卵基質,在族群數量與產卵基質的條件都良好的情況之下,這樣 會迫使雌蟲將體內全部的卵給產出,也因此在第一和第二個週期當中,整個週期 的平均產卵量是可以接近生理攜卵量測定結果的上限。
在之前研究中所做出的大頭金蠅生命表中,認為在羽化後 10 天有著最高的 生殖能力。如果與結果對照來看,大概是第一週期出現的時間點。另外該研究中 的大頭金蠅的存活率會在羽化後日齡 25 天後開始下降,這個研究結果也與生殖週 期中的存活符合這樣的趨勢 (Gabre et al., 2005)。也就是第一週期和第二週期的平 均產卵量顯著高於第三週期,這樣的結果可能代表著隨著日齡的增加和生產次數 的增加,雌蟲本身的生殖和發育能力可能會有所下降,即便有適合的產卵基質,
雌蟲也因為已經不是有著最高生殖能力的時間點而產出的卵量不夠。而另外一個 可能的原因在於本身的壽命上,在實驗當中,當第二週期結束之後,會有部分雌 蟲自然死亡,而當初實驗設置中的雌蟲數目為 5 隻一組,然而當有部分因為自然 死亡之後,數量就會來到 5 隻以下的情況,根據之前的族群數量的實驗結果,這 樣有可能會造成產卵表現量的降低,而導致第三週期的平均產卵量的下降。
四、單隻雌蟲的載卵壓力測試
在單隻雌蟲的結果當中,初始的產卵日齡超過 20 天以上,在 Minkenberg et al.
(1992) 提及載卵壓力的結果中,日齡的提高的確對於個體本身提高載卵壓力,而 會讓原先不想產卵的雌蟲,被迫要選擇產出部分的卵來降低這個壓力,或者是採 取另外的策略來應對這逐漸提高的載卵壓力。而單雌結果與當初族群數量結果中 的單隻的結果有所差異。在原先低日齡且產卵基質的情況相同之下,單隻雌蟲是 選擇不產卵,但是當日齡的提高之下,單隻雌蟲被迫釋放出部分的卵。但是還是 有約一半的雌蟲組別依然選擇沒有任何卵的產出來應對逐漸提高的日齡。這樣或 許在載卵壓力這個部分,提高日齡對於大頭金蠅的雌蟲的影響可能不大,儘管有 部分的個體有產卵反應,但是這樣的產卵量是低於整個生殖週期中完整的產卵總 量。
而為了更進一步的確定這些沒有產卵的組別究竟是否有選擇其他的策略來 對應提高的載卵壓力,像是將卵給吸收掉來做為營養,決定將全部的組別進行體 內剩餘卵的檢查以及將產出和體內剩餘卵量的計算。而在經過統計分析之後,結 果顯示體內剩餘卵量顯著高於產出的卵量。這表示在單隻雌蟲的情況之下,雌蟲 就算在有適合產卵基質的情況下,選擇不產卵的為大多數,而且不僅是不選擇產 卵,在提高日齡之後,大部分的雌蟲依舊選擇將卵保留在體內,而沒有進行更進 一步的反應。在有關於載卵壓力的研究當中,多半認為生物本身需要去平衡自身 的生殖資源的投資和自身生存的效率兩者之間來做平衡,而在這當中提到如果對 於載卵壓力提高沒有產生產卵行為的反應,可能是自身有很好的方式去調節,或 者是本身的尋找基質的能力很好而不表現產卵行為 (Dieckhoff et al., 2014)。
而就算有產卵行為的出現,在初始的產卵日期,與生殖週期的初始產卵日相 比,落後了不少的天數。而在平均產卵量的部分,遠低於正常的生殖週期結果的 平均卵量。對於這樣的結果,Goodbrod and Goff (1990) 中的結果提出在幼蟲中,
個體數量越高時會有協力取食的現象。雌蟲可能認為,在沒有同伴的情況之下,
單一雌蟲產出的幼蟲數量不足以提高取食效率。而在沒有聚集效應來影響雌蟲產 卵的情況之下,雌蟲對於提高日齡作為載卵壓力反應是認為本身不受到影響才會
不將卵產出。
而此結果另外的可能,則是在於刺激產卵因子方面的重要性,什麼能夠有效 的刺激大頭金蠅雌蟲產卵一直都是複雜且牽涉許多的因子。在 Yang and Shiao (2012); Yang (2014)及 Yang (2014) 的結果當中,提及了視覺可能是主要因子的結 論。而在單雌產卵結果中,在沒有了聚集效應的情況之下,單雌對於載卵壓力的 影響可以認為是沒有,而在族群對於數量的結果中,雌蟲數量的差距則是會影響 著產卵平均量的表現。綜合以上這兩者的結果而言,大頭金蠅的雌蟲的產卵可能 是首先要尋找產卵基質,接著利用視覺判斷是否要為合適的產卵基質,這包括了 同種或者其他種類的卵塊或者幼蟲的存在,和產卵基質附近同種成蟲的存在。而 對於載卵壓力的影響,在這些結果中並沒有聚集效應來的重要。
陸、結論
麗蠅的產卵行為會影響其產卵量,進而直接影響幼蟲的數量,而幼蟲的數量 多寡會造成接下來的捕食、競爭等交互作用。最終造成族群的存活率與生長速率 的改變,進而去影響到 PMI 估算時候的誤差與判斷。本研究以大頭金蠅為材料,
探討聚集效應與載卵壓力對於麗蠅產卵的影響,以及收集麗蠅生殖週期的基礎資 料。
在此研究中的結果顯示,在麗蠅的生殖週期結果中取得了產卵量、產卵週期 出現的日齡,以及整體族群的存活狀況。其中前兩個週期的產卵量顯著高於第三 個週期。而在共域蟲數對於雌蟲的產卵表現量的結果,則是與前人的結果相同,
在五隻以上的共域數量下,大頭金蠅會表現正常的產卵量,而低數量共域的雌蟲 即便在給予產卵基質的情況下,表現量則是顯著低於高數量共域。推測大頭金蠅 除了需要產卵基質之外,族群數量可能是一個重要的刺激因子。
單隻雌蠅的載卵壓力測試結果顯示即便提高日齡來藉此增加載卵壓力,但是 大頭金蠅給予的產卵反應卻沒有顯著,在計算完體內的殘留卵量以及產出的卵量 作分析之後,體內留存卵量顯著高於產出卵量。這樣的結果可能對於在大頭金蠅 面對載卵壓力及族群數量兩個因子時,且產卵基質存在時,族群數量可能有比較 大的影響來刺激產卵,而載卵壓力可能對於大頭金蠅雌蟲產卵行為的影響並不 高。
以此結果來看,若在往後的進行相關產卵行為的研究的時候,在數量高於五 隻以上時,才會有正常的產卵表現量,這個結果不論是在飼養或者實驗設置中的 共域雌蟲數量是需要列入考量的。而在往後產卵因子的探討當中,可能也需要將 雌蟲數量有比較優先的順位來做考量。另外在單隻雌蟲對於產卵基質並沒有反應 的部分,可能可以應用於極端的狀況下來推算對於 PMI 估算誤差有所幫助,也 可能代表某些刺激產卵的因子要一併存在的情況之下,才能刺激麗蠅的產卵。
參考文獻
Amendt J, Krettek R, Zehner R. 2004. Forensic entomology. Naturwissenschaften 91:
51-65.
Amendt J, Zehner R, Reckel F. 2008. The nocturnal oviposition behaviour of blowflies (Diptera: Calliphoridae) in Central Europe and its forensic implications.
Forensic Sci Int 175: 61-64.
Anderson GS. 2000. Minimum and maximum development rates of some forensically important Calliphoridae (Diptera). J Forensic Sci 45: 824-832.
Anderson GS. 2004. Determining time of death using blow fly eggs in the early postmortem interval. Int J Legal Med 118: 240-241.
Baldridge RS, Wallace SG, Kirkpatrick R. 2006. Investigation of nocturnal oviposition by necrophilous flies in central Texas. J Forensic Sci 51: 125-126.
Barnes KM, Grace KA, Bulling MT. 2015. Nocturnal oviposition behavior of forensically important Diptera in Central England. J Forensic Sci 60:
1601-1604.
Barton BL. 2001. Quantitative aspects of the regulation of ovarian development in selected anautogenous Diptera: integration of endocrinology and nutrition.
Entomol Exp Appl 100: 137-149.
Beaver RA. 1977. Non-equilibrium `island' communities: Diptera breeding in dead snails. J Anim Ecol 46: 783-798.
Benecke M. 2001. A brief history of forensic entomology. Forensic Sci Int 120: 2-14.
Bourel B, Callet Bt, Hédouin V, Gosset D. 2003. Flies eggs: a new method for the estimation of short-term post-mortem interval? Forensic Sci Int 135: 27-34.
Browne LB. 1993. Physiologically Induced changes in resource-oriented behavior.
Annu Rev Entomol 38: 1-23.
Bunchu N, Sukontason KL, Olson JK, Kurahashi H, Sukontason K. 2008. Behavioral responses of Chrysomya megacephala to natural products. Parasitol Res 102:
419-429.
Byrd JH, Castner JL. 2001. Insects of forensic importance. pp. 43-80. In: Byrd JH, Castner JL, (eds). Forensic Entomology: The Utility of Arthropods in Legal Investigations. CRC Press, Boca Raton.
Campobasso CP, Di Vella G, Introna F. 2001. Factors affecting decomposition and Diptera colonization. Forensic Sci Int 120: 18-27.
Catts EP, Goff ML. 1992. Forensic entomology in criminal investigations. Annu Rev Entomol 37: 253-272.
Chaiwong T, Sukontason K, Chaisri U, Kuntalue B, Vogtsberger RC, Sukontason KL.
megacephala (Diptera: Calliphoridae). Int J Parasitol Res 4: 65-70.
Chang CY. 2003. Taxonomic studies of forensically important blow flies from northern Taiwan (Diptera: Calliphoridae) [M.S. thesis]. Taiwan: National Taiwan University. 110 pp.
Davies L. 1950. The hatching mechanism of muscid eggs (Diptera). J Exp Biol 27:
437-445.
Dieckhoff C, Theobald JC, Wäckers FL, Heimpel GE. 2014. Egg load dynamics and the risk of egg and time limitation experienced by an aphid parasitoid in the field. Ecol Evol 4: 1739-1750.
Ellers J, Sevenster JG, Driessen G. 2000. Egg load evolution in parasitoids. Am Nat 156: 650-665.
Ellis AM. 2008. Incorporating density dependence into the oviposition preference–
offspring performance hypothesis. J Anim Ecol 77: 247-256.
Erzinçlioğlu Z. 1996. Blowflies. UK: Richmond Publishing Company.
Gabre RM, Adham FK, Chi H. 2005. Life table of Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae). Acta Oecol 27: 179-183.
Gennard DE. 2007. The breadth of forensic entomology. pp. 1-18. Forensic entomology: an introduction. John Wiley & Sons Ltd, England.
George KA, Archer MS, Toop T. 2012. Effects of bait age, larval chemical cues and nutrient depletion on colonization by forensically important calliphorid and sarcophagid flies. Med Vet Entomol 26: 188-193.
Gomes L, Gomes G, Casarin FE, Silva IMd, Sanches MR, Von Zuben CJ, Fowler HG.
2007. Visual and olfactory factors interaction in resource-location by the blowfly, Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae), in natural conditions. Neotrop Entomol 36: 633-639.
Goodbrod JR, Goff ML. 1990. Effects of larval population density on rates of development and interactions between two species of Chrysomya (Diptera:
Calliphoridae) in laboratory culture. J Med Entomol 27: 338-343.
Hall RD. 1990. Medicocriminal entomology. pp. 1-8. In: Elmer Paul Catts, Haskell NH, (eds). Entomology & Death: A Procedural Guide. Joyce's Print Shop, Clemson.
Hall RD. 2001. Introduction: perceptions and status of forensic entomology. pp. 1-15.
In: Jason H. Byrd, James L. Castner, (eds). Forensic Entomology: The Utility of Arthropods in Legal Investigations. CRC Press, Boca Raton.
Hall RD, Haskell NH. 1995. Forensic entomology: applications in medicolegal investigation. In: C W, (ed). Forensic Sciences. Matthew Bender, New York.
Hammer O, Harper D, Ryan P. 2001. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol Electron 4: 1-9.
Harterreiten-Souza ES, Pujol-Luz JR. 2012. Comparative morphology of the
spermathecae of some species of Chrysomya Robineau-Desvoidy and Cochliomyia Townsend (Diptera, Calliphoridae). Rev Bras Entomol 56: 54-58.
Hong ZZ. 1995. The life table and mass rearing of Chrysomya megacephala (Fabricius) [M.S. thesis]. Taiwan: National Taiwan University. 77 pp.
Jian MZ. 2004. The insect fauna on disposed carrions in Taiwan area [M.S. thesis].
Taiwan: Central Police University. 132 pp.
Joy JE, Liette NL, Harrah HL. 2006. Carrion fly (Diptera: Calliphoridae) larval colonization of sunlit and shaded pig carcasses in West Virginia, USA. Forensic Sci Int 164: 183-192.
Kano R, Shinonaga S. 1968. Calliphoridae (Insecta: Diptera). Fauna Japonica. Tokyo:
Tokyo Biogeographical Society of Japan. 181 pp.
Keh B. 1985. Scope and applications of forensic entomology. Annu Rev Entomol 30:
137-154.
Liao CS. 2016. Effects of early decomposition time on blow fly (Diptera:
Calliphoridae) oviposition [M.S. thesis]. Taiwan: National Taiwan University.
55 pp.
Lord WD, Stevenson JR. 1986. Directory of Forensic Entomologists. Washington, D.C.: Defense Pest Information Analysis Center; Walter Reed Army Medical Center. 42 pp.
Mégnin J. 1894. La faune des cadavres: application de l’entomologie a la médecine légale. Paris: Masson et Gauthiers-Villars.
Marchenko MI. 2001. Medicolegal relevance of cadaver entomofauna for the determination of the time of death. Forensic Sci Int 120: 89-109.
Marinho MAT, Junqueira ACM, Paulo DF, Esposito MC, Villet MH, Azeredo-Espin AML. 2012. Molecular phylogenetics of Oestroidea (Diptera: Calyptratae) with emphasis on Calliphoridae: Insights into the inter-familial relationships and additional evidence for paraphyly among blowflies. Mol Phylogen Evol 65:
840-854.
Minkenberg OPJM, Tatar M, Rosenheim JA. 1992. Egg Load as a Major Source of Variability in Insect Foraging and Oviposition Behavior. Oikos 65: 134-142.
Norris KR. 1965. The bionomics of blow flies. Annu Rev Entomol 10: 47-68.
Shiao SF, Yeh TC. 2008. Larval competition of Chrysomya megacephala and Chrysomya rufifacies (Diptera: Calliphoridae): behavior and ecological studies of two blow fly species of forensic significance. J Med Entomol 45: 785-799.
Singh D, Bharti M. 2001. Further observations on the nocturnal oviposition behaviour of blow flies (Diptera: Calliphoridae). Forensic Sci Int 120: 124-126.
Sukontason K, Piangjai S, Siriwattanarungsee S, Sukontason KL. 2008. Morphology
rufifacies in Thailand: application in forensic entomology. Parasitol Res 102:
1207-1216.
Thompson CR, Brogan RS, Scheifele LZ, Rivers DB. 2013. Bacterial interactions with necrophagous flies. Ann Entomol Soc Am 106: 799-809.
Wall R, Fisher P. 2001. Visual and olfactory cue interaction in resource‐location by the blowfly, Lucilia sericata. Physiol Entomol 26: 212-218.
Wells JD, Byrd JH, Tantawi TI. 1999. Key to third-instar chrysomyinae (Diptera:
Calliphoridae) from carrion in the continental United States. J Med Entomol 36:
638-641.
Williams KA, Wallman JF, Lessard BD, Kavazos CRJ, Mazungula DN, Villet MH.
2017. Nocturnal oviposition behavior of blowflies (Diptera: Calliphoridae) in the southern hemisphere (South Africa and Australia) and its forensic implications. Forensic Sci Med Pathol 13: 123-134.
Wooldridge J, Scrase L, Wall R. 2007. Flight activity of the blowflies, Calliphora vomitoria and Lucilia sericata, in the dark. Forensic Sci Int 172: 94-97.
Yang ST. 2014. Taxonomy of blow flies in Taiwan and reproductive behavior of Chrysomya megacephala (Diptera: Calliphoridae) [Ph.D dissertation]. Taiwan:
National Taiwan University. 167 pp.
Yang ST, Shiao SF. 2012. Oviposition preferences of two forensically important blow fly species, Chrysomya megacephala and C. rufifacies (Diptera: Calliphoridae), and implications for postmortem interval estimation. J Med Entomol 49:
424-435.
Yovanovich P. 1888. Entomologie appliquée ã la Médecine légale. Olliver-Henry, Paris.
Zhu GH, Xu XH, Yu XJ, Zhang Y, Wang JF. 2007. Puparial case hydrocarbons of Chrysomya megacephala as an indicator of the postmortem interval. Forensic Sci Int 169: 1-5.
表一、大頭金蠅 (Chrysomya megacephala) 生殖週期產卵量之常態分布測定結果。
下表為生殖週期測定中三個週期的平均產卵量及總產卵量做常態分布測定的結果 Table 1. Normality test results of the reproductive cycle of Chrysomya megacephala.
The following table shows the results of the normal distribution test of the average fecundity for the three reproductive cycles and total egg production
Total egg number (Mean ± SD)
N* P-value
Total egg number per group First cycle
Second cycle Third cycle
630.9 ± 65.0 15 0.2383 278.9 ± 48.1 15 0.009636*
258.9 ± 36.7 15 0.5199 92.7 ± 52.9 15 0.01648*
N* = replication for each experiment, five female flies were used for each experiment.
*: p < 0.05.
表二、大頭金蠅 (Chrysomya megacephala) 不同共域雌蟲數量組別間產卵量差異 的統計分析結果。表格中為各兩組之間產卵量差異統計 p 值
Table 2. The statistical analysis results of the differences between the egg numbers laid per female for different groups with different coexisted female numbers of Chrysomya megacephala are shown in the table for the statistical p-values between the two groups
No. coexisted females
1 2 3 4 5 6 7 8 9 10
1 0.05799 0.05383 0.008731 3.38E-11 6.86E-06 4.69E-09 1.93E-08 2.80E-08 2.33E-08
2 0.9776 0.5292 4.15E-05 0.02423 0.000601 0.001265 0.001537 0.001395
3 0.5477 4.68E-05 0.02606 0.000666 0.001395 0.001693 0.001537
4 0.000521 0.1043 0.00508 0.009473 0.01114 0.01028
5 0.06494 0.5042 0.3814 0.3515 0.3663
6 0.2389 0.3319 0.3607 0.3461
7 0.8357 0.792 0.8138
8 0.9551 0.9776
9 0.9776
10
The Dunn's post hoc analysis was used to show whether there was a significant difference in the average egg production between the two groups. The green block indicates p < 0.05.
表三、大頭金蠅 (Chrysomya megacephala) 單隻雌蟲載卵壓力測試結果。受試單 隻雌蟲每一組的產卵量及產卵日齡,在日齡 42 天結束之後,解剖計算殘存在體內 卵量
Table 3. Egg load test results for a single Chrysomya megacephala female. Numbers of egg deposited and time of oviposition in 20 single female tests. Residual egg numbers were counted after dissection for the 42-day old female
Replicate
Total egg number deposited
Residual egg number in ovary
Oviposition time (days after emergence) and egg number
1 132 121 24(132)
2 29 175 30(29)
3 120 102 29(22), 33(98)
4 48 167 30(48)
5 52 185 26(52)
6 87 158 22(65), 32(22)
7 105 142 23(105)
8 44 185 28(44)
9 36 125 29(36)
10 32 148 31(32)
11 28 169 30(28)
12 0 224 -
13 0 217 -
14 0 202 -
15 0 218 -
16 0 244 -
17 0 251 -
18 0 213 -
19 0 252 -
20 0 234 -
圖一、大頭金蠅 (Chrysomya megacephala) 雌蟲腹部及卵巢圖。A:大頭金蠅雌 蟲腹部。B:剪開腹部前兩節表皮的成熟麗蠅雌蟲,可以看到卵巢本身佔了腹部 很大的一部分。C:未煮過的卵巢,卵跟卵之間的界限無法看清楚。D:煮過的卵 巢,每顆卵可以用肉眼就能辨識,容易分開且不會破裂。
Figure 1. Chrysomya megacephala female abdomen and ovary. A, Abdomen, ventral view; B, Ovaries account for a large part of the abdomen, as shown after the dissection of gravid female; C, Uncooked ovaries, the space between eggs and eggs could not be identified clearly; D, Cooked ovaries, each egg can be identified with the naked eye, easy to separate and not easily broken.
C A
D B