國立臺灣大學生物資源暨農學院園藝暨景觀學系 碩士論文
Department of Horticulture and Landscape Architecture College of Bioresources and Agriculture
National Taiwan University Master Thesis
以高通量定序方式分析蝴蝶蘭感染齒舌蘭輪斑病毒及 蕙蘭嵌紋病毒後小型核醣核酸組成之變化
Genome Wide Analysis of Small RNAs from
Odontoglossum ringspot virus and
Cymbidium mosaic virus Infected Phalaenopsis by Deep Sequencing
白 暄 Hsuan Pai
指導教授:張耀乾 博士、林納生 博士
Advisor: Dr. Yao-Chien Alex Chang and Dr. Na-Sheng Lin 中華民國 102 年 1 月
January 2013
I
Contents
Contents ... I Contents of Tables ... V Contents of Figures ... VII
Abstract ... 1
Introduction ... 3
Literature Review ... 6
1. The incidence of virus diseases in orchids ... 6
1.1. Cymbidium mosaic virus (CymMV) ... 6
1.2. Odontoglossum ringspot virus (ORSV) ... 7
1.3. Symptoms and pathogenesis of CymMV and ORSV in orchids ... 8
1.4. Molecular mechanisms of the synergism between CymMV and ORSV ... 10
2. The role of RNA silencing in plant-virus interactions ... 12
2.1. RNA silencing pathways in plants ... 12
2.2. Profiling characteristics of virus-derived small interfering RNAs (vsRNAs) ... 14
2.2.1. Functions of DCL proteins and size distribution of vsRNAs ... 15
2.2.2. Preference of 5’ terminal nucleotide of vsRNAs ... 16
2.2.3. Strand polarity and hotspot distribution of vsRNAs along the virus genome ... 17
2.3. Interactions between virus-modulated RNA silencing and the pathogenesis of viruses in plants ... 18
2.3.1. Roles of viral suppressors of RNA silencing (VSRs) in symptom induction ... 18
2.3.2. Symptom induction through vsRNA-mediated host gene regulation ... 19
2.3.3. Infection induced dysregulation of miRNA expression and its association in plant-virus interactions ... 20
Materials and Methods ... 22
1. Maintenance of CymMV- and ORSV-free plants ... 22
1.1. Plant materials and growing conditions ... 22
1.2. Detecting virus infection in newly purchased plants ... 22
1.2.1. Extracting total RNA from leaf tissues ... 22
II
1.2.2. Reverse transcription-polymerase chain reaction (RT-PCR)... 23
2. Detection of CymMV and ORSV spread in P. amabilis ... 24
2.1. Experimental locations ... 24
2.2. Virus source ... 24
2.3. Virus inoculation and sequential sampling ... 25
2.4. Tissue blotting and hybridization assay for detecting virus ... 26
2.4.1. Tissue blotting ... 26
2.4.2. Preparation of digoxigenin (DIG)-labeled probes ... 27
2.4.3. Hybridization and chemiluminescent detection ... 28
3. Genome wide analysis of small RNAs from ORSV-infected P. amabilis by deep sequencing ... 29
3.1. ORSV inoculation assay and RNA sample preparation ... 29
3.2. Analyzing viral RNA and siRNA accumulation ... 29
3.2.1. Analyzing viral RNA accumulation by Northern blot ... 29
3.2.2. Analyzing viral siRNA accumulation by small RNA Northern blot ... 30
3.3. Small RNA deep-sequencing and annotation ... 31
3.3.1. cDNA library construction and sequencing ... 31
3.3.2. Sequencing data trimming and annotation ... 32
3.3.3. Target prediction for vsRNAs and miRNAs ... 33
3.3.3.1. Analyzing differential expressions of miRNAs ... 33
3.3.3.2. Prediction of putative miRNA target genes ... 33
3.3.3.3. Prediction of putative vsRNA target genes ... 34
3.4. Experimental validation of small RNAs ... 35
3.4.1. Validating miRNA expression by stem-loop quantitative RT-PCR (qRT-PCR) ... 35
3.4.2. VsRNA co-transfection assay in Oncidium protoplasts ... 36
3.4.2.1. Protoplast isolation ... 36
3.4.2.2. Co-transfection by electroporation method ... 37
3.4.2.3. RNA purification and Northern blot analysis ... 37
4. Genome wide analysis of small RNAs from CymMV and double infected P. amabilis by deep sequencing ... 38
4.1. CymMV and mixed inoculation assays ... 38
III
4.2. Analyzing viral RNA and siRNA accumulation ... 38
4.3. Construction of small RNA libraries from CymMV and double infected P. amabilis and bioinformatic analysis ... 39
Results ... 40
1. Preliminary time course assay of CymMV and ORSV infection in P. amabilis ... 40
1.1. ORSV infection in inoculated leaves 2 to 11 dpi ... 40
1.2. ORSV infection spreaded to non-inoculated tissues after 10 dpi... 40
1.3. Accelerated spreading of CymMV infection in mixed inoculated leaves ... 41
2. Analysis of small RNAs from ORSV-infected P. amabilis by deep sequencing ... 42
2.1. Analysis of ORSV infection ... 42
2.2. Deep sequencing of small RNAs from ORSV-infected P. amabilis ... 43
2.3. Characteristics of ORSV viral siRNAs (vsRNAs) ... 45
2.3.1. ORSV vsRNA populations in inoculated (Oi) and non-inoculated (Oc) tissues ... 45
2.3.2. Size distribution, strand polarity, and 5’-end nucleotide preference of ORSV vsRNAs ... 45
2.3.3. Genome mapping, coverage and hotspots of ORSV vsRNAs ... 46
2.3.4. Specific vsRNA hotspot in ORSV 3’-UTR ... 47
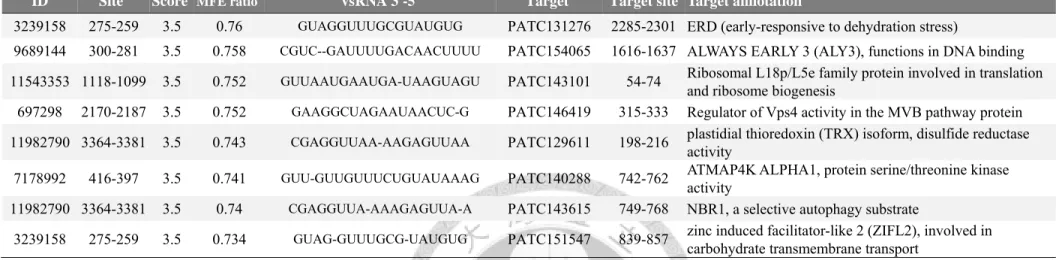
2.3.5. Functional analysis of ORSV 3’-UTR hotspot vsRNA ... 48
2.4. Identification and analysis of conserved miRNAs ... 49
2.4.1. miRNA populations ... 49
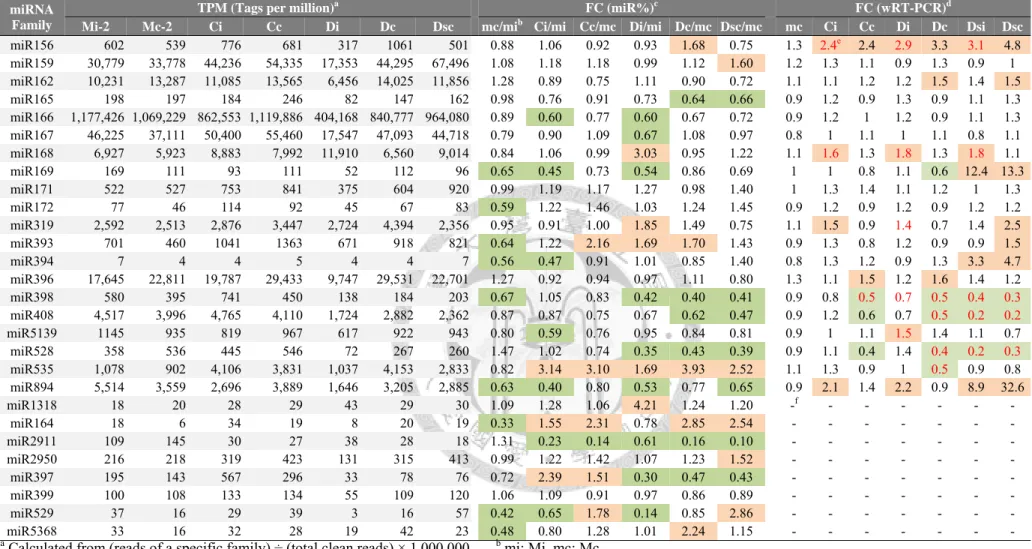
2.4.2. Differential expressions of miRNAs in response to ORSV infection ... 50
2.4.3. Target prediction and possible roles of ORSV-infection responsive miRNAs ... 51
3. Analyses of small RNAs from CymMV and mixed infected P. amabilis by deep sequencing ... 53
3.1. Analysis of CymMV and mixed infection... 53
3.2. Deep sequencing of small RNAs from CymMV and mixed infected P. amabilis ... 55
3.3. Characteristics of vsRNAs in CymMV and doubly infected tissues ... 56
3.3.1 VsRNA populations in singly and doubly infected tissues ... 56
3.3.2. Characteristics of vsRNAs in CymMV and double infected tissues ... 57
IV
3.3.3. Genome mapping and coverage of CymMV vsRNAs ... 58 3.3.4. Specific ORSV vsRNA hotspots occurred in mixed infected tissues ... 59 3.3.5. Prediction of potential P. amabilis target transcripts of vsRNAs ... 59 3.4. Identification and analysis of conserved miRNAs in CymMV and mixed
infected Phalaenopsis amabilis ... 61 3.4.1. miRNA populations ... 61 3.4.2. Differential expressions and predicted targets of CymMV and double
infection responsive miRNAs ... 62 Discussion ... 64
1. Synergistic enhancement of CymMV infection by ORSV co-inoculation in
Phalaenopsis amabilis ... 64 2. The leading roles of DCL4 and DCL2 in P. amabilis ... 66 3. Asymmetrical strand polarity and 5’-end nucleotide identity of CymMV and
ORSV vsRNAs ... 66 4. Differential distribution of CymMV and ORSV vsRNA along viral genomes ... 68 5. VsRNA-mediated host gene silencing may underly the mechanism of symptom
formation ... 70 6. Roles of infection responsive miRNAs in Phalaenopsis-virus interactions ... 71 7. The involvement of novel miRNAs and other small RNAs in response to viral
stresses ... 74 Conclusion and Future Directions ... 76 References ... 78
V
Contents of Tables
Table 1. Summary of total reads in small RNA libraries constructed from mock- and
ORSV-inoculated tissues. ... 98
Table 2. Summary of unique reads in small RNA libraries constructed from mock- and ORSV-inoculated tissues. ... 99
Table 3. The major sequences and total reads of abundant miRNAs in small RNA libraries constructed from mock- and ORSV-inoculated tissues. ... 100
Table 4. Expression fold-changes of some abundant miRNAs in small RNA libraries constructed from mock- and ORSV-inoculated tissues. ... 105
Table 5. Putative target genes of ORSV infection responsive miRNAs. ... 106
Table 6. Summary of total reads in small RNA libraries constructed from mock-, CymMV-, and CymMV and ORSV mixedly-inoculated tissues. ... 108
Table 7. Summary of total unique reads in small RNA libraries constructed from mock-, CymMV-, and CymMV and ORSV mixedly-inoculated tissues. ... 109
Table 8. Potential ORSV vsRNA targeting Phalaenopsis genes. ... 110
Table 9. Potential Di library-specific ORSV vsRNA targeting Phalaenopsis genes. ... 114
Table 10. Potential CymMV vsRNA targeting Phalaenopsis genes. ... 116
Table 11. Potential CymMV vsRNA targeting Phalaenopsis genes... 117
Table 12. Potential Di library-specific CymMV vsRNA targeting Phalaenopsis genes. (continued) ... 121
Table 13. The major sequences and total reads of abundant miRNAs in small RNA libraries constructed from mock-, CymMV-, and CymMV and ORSV mixedly-inoculated tissues. ... 128
Table 14. Expression fold-changes of some abundant miRNAs in mock-, CymMV-, or mixedly-infected tissues. ... 137
Table 15. Putative target genes of virus infection-responsive miRNAs. ... 138
Table S1. Primers used in RT-PCR and stem-loop qRT-PCR. ... 140
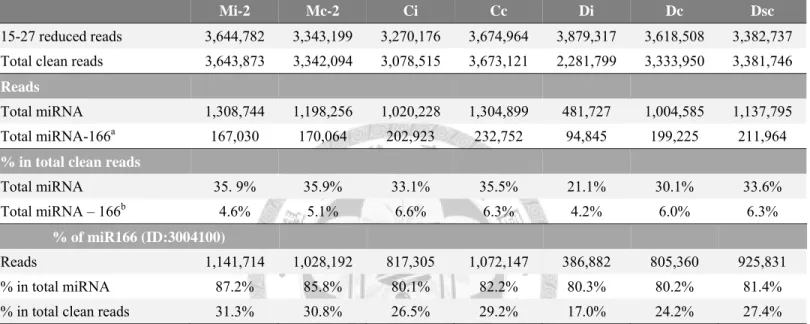
Table S2. Information about the super-abundant miR166 (ID:3004100) tag small RNA libraries constructed from mock- and ORSV-infected tissues. ... 143 Table S3. The non-redundant list of the top 50 abundant CymMV vsRNAs in Ci, Di and
VI
Dc libraries. ... 144 Table S4. The non-redundant list of the top 50 abundant ORSV vsRNAs in Oi and Di
libraries. ... 146 Table S5. List of ORSV vsRNAs specifically sequenced in Di library. ... 149 Table S6. List of CymMV vsRNAs specifically sequenced in Di library. ... 150 Table S7. The super-abundant miR166 (ID:3004100) tag in small RNA libraries of
mock-, CymMV-, and mixedly-inoculated tissues. ... 151
VII
Contents of Figures
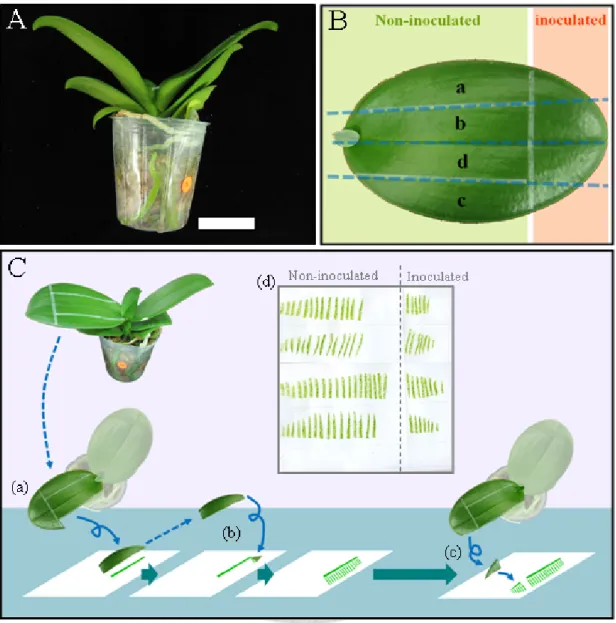
Fig. 1. Diagrams of leaf tip-inoculation and tissue-blotting methods used in the infection
time-course assays. ... 152
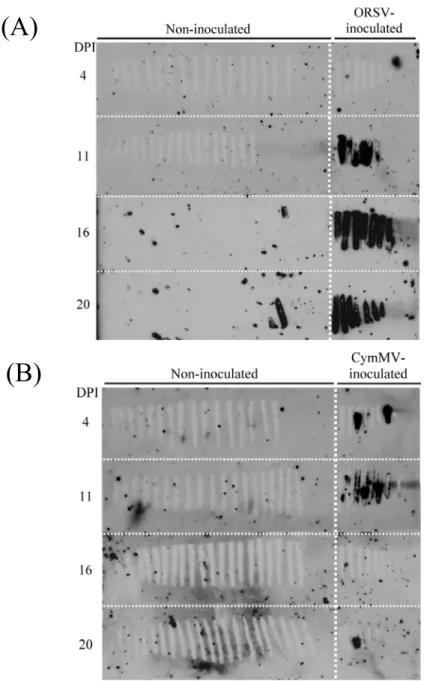
Fig. 2. Time-course detection of ORSV infection in ORSV-inoculated Phalaenopsis. ... 153
Fig. 3. Time-course detection of ORSV infection in one month period in ORSV-inoculated Phalaenopsis. ... 154
Fig. 4. Time-course detection of virus infection in ORSV or CymMV singly inoculatedPhalaenopsis. ... 155
Fig. 5. Time-course detection of virus infection in mixedly-inoculated Phalaenopsis. ... 156
Fig. 6. Diagrams of leaf tip-inoculation and tissue-blotting methods used in the inoculation assays. ... 157
Fig. 7. Virus infection of ORSV-inoculated Phalaenopsis at 10 days post inoculation. ... 158
Fig. 8. Accumulation of ORSV viral RNA and virus-specific siRNA in Phalaenopsis. ... 159
Fig. 9. Size distribution of small RNAs in mock- or ORSV-inoculated Phalaenopsis. ... 160
Fig. 10. Annotation procedures of deep sequencing data. ... 161
Fig. 11. Abundance and size distribution of ORSV vsRNAs in ORSV-infected Phalaenopsis. ... 162
Fig. 12. Nucleotide composition of ORSV vsRNAs in ORSV-infected Phalaenopsis. ... 163
Fig. 13. Distribution of vsRNAs along ORSV genome corresponding to reads from Oi and Oc libraries. ... 164
Fig. 14. Alignment of tobamovirus 3’-untranslated region (3’-UTR) conserved sequence. Identical bases among 22 viruses are highlighted in grey. ... 165
Fig. 15. Location and frequency of abundant 3’-UTR-originating ORSV positive stranded vsRNAs. ... 166
Fig. 16. ORSV viral RNA accumulation in ORSV and synthetic siRNAs co-transfected protoplasts. ... 167
Fig. 17. The abundance of miRNA families in Mi, Mc, Oi, and Oc libraries. The percentage was calculated from sum of Mi, Mc, Oi, and Oc libraries. ... 168
Fig. 18. Expression levels of some ORSV-infection responsive miRNAs validated by stem-loop qRT-PCR. ... 169
Fig. 19. Expression of some miRNAs in ORSV- or mock-inoculated tissues validated by stem-loop qRT-PCR. ... 170
VIII
Fig. 20. Virus infection of CymMV- and doubly-inoculated Phalaenopsis at 10 days
post inoculation (DPI). ... 172 Fig. 21. Accumulation of CymMV and ORSV viral RNA and virus-specific siRNA in
Phalaenopsis. ... 173 Fig. 22. Size distribution of small RNAs in mock-, CymMV-, or CymMV and ORSV
mixedly-inoculated Phalaenopsis. ... 174 Fig. 23. Abundance and size distribution of CymMV vsRNAs in CymMV, or CymMV
and ORSV mixedly-inoculated Phalaenopsis. ... 175 Fig. 24. Abundance and size distribution of ORSV vsRNAs in CymMV and ORSV
mixedly-inoculated Phalaenopsis. ... 176 Fig. 25. Nucleotide composition of 5’-end of CymMV vsRNAs in CymMV, or CymMV
and ORSV mixedly-inoculated Phalaenopsis. ... 177 Fig. 26. Nucleotide composition of 5’-end of ORSV vsRNAs in CymMV and ORSV
mixedly-inoculated Phalaenopsis. ... 178 Fig. 27. Distribution of vsRNAs along CymMV genome corresponding to reads from Ci,
Cc, Di, Dc and Dsc libraries. ... 179 Fig. 28. Distribution of vsRNAs along ORSV genome corresponding to reads from Di,
Dc, and Dsc libraries. ... 180 Fig. 29. The abundance of miRNA families in Mi-2, Mc-2, Ci, Cc, Di, Dc, and Dsc
libraries. ... 181 Fig. 30. Fold changes of expression levels of some virus infection-responsive miRNAs
in mock-, CymMV-, or CymMV and ORSV mixedly-inoculated Phalaenopsis
validated by stem-loop qRT-PCR. ... 182 Fig. 31. Fold changes of expression levels of some miRNAs in mock-, CymMV-, or
CymMV and ORSV mixedly-inoculated Phalaenopsis validated by stem-loop
qRT-PCR. ... 183
Fig. S1. Plasmids used for generating ORSV- or CymMV-specific RNA probes for
detecting virus infection. ... 184 Fig. S2. Plasmids used for generating ORSV- or CymMV-specific RNA probes for
detecting vsRNA accumulation. ... 185 Fig. S3. Percentages of CymMV and ORSV vsRNAs of total clean reads in small RNA
IX
libraries. ... 186 Fig. S4. Distribution of 21- and 22-nt viral siRNAs along ORSV genome corresponding
to reads from Oi library. ... 187 Fig. S5. Validation of miR166 abundance. ... 188 Fig. S6. Size distribution of small RNAs modified with the removal of the
super-abundant miR166 tag. ... 189 Fig. S7. Modified relative abundance of miRNA families after removal of the
super-abundant miR166 tag. ... 190 Fig. S8. The redundancy of top 50 abundant vsRNA tags in Oi, Ci, Di, and Dc libraries. .... 191
1
Abstract
Cymbidium mosaic virus (CymMV) and Odontoglossum ringspot virus (ORSV) are the most prevalent viruses infecting orchids and causing economic losses worldwide.
Upon virus infection, small RNA mediated antiviral RNA silencing response is activated. Such biotic stress may affect virus-specific interfering RNAs (vsRNAs) or microRNA (miRNA) regulated host gene expression. To advance our understanding of the offense-defense interactions between CymMV, ORSV and orchids, this study employed deep sequencing to analyze small RNAs from virus infected Phalaenopsis.
The leaf tip-inoculation method first distinguished early and late stages of infection in non-inoculated and inoculated tissues at ten days post inoculation (dpi). Small RNA Solexa sequencing generated 11 libraries with more than five million reads each from CymMV and ORSV singly and doubly inoculated leaves and from mock-inoculated leaves. Generally, CymMV and ORSV vsRNAs were predominantly 21 and 22 nucleotides (nt), with excess positive polarity accumulating in single inoculations.
While most CymMV vsRNAs were derived from RNA-dependent RNA polymerase (RdRp) coding regions, ORSV vsRNAs encompassed the coat protein coding gene and 3’-untranslated region, with a specific hotspot residing in the pseudoknot upstream to the 3’-terminal tRNA-like structure. These results suggest Dicer-like (DCL) 4 and DCL2 homologs play a leading role in mediating antivirus RNA silencing in P.
amabilis using single-stranded RNA derived secondary structure as templates. Biased distribution of 5’ terminal adenosine (A), uridine (U), cytosine (C) and underrepresented 5’-guanine (G) indicate vsRNAs could be recruited into multiple Argonaute (AGO) complexes. Under mixed infection, chlorotic necrosis symptoms appeared specifically in inoculated tissues at 10 dpi, and accelerated spreading and
2
enhanced viral titer of CymMV also occurred. The proportion of CymMV vsRNAs in total small RNAs ranged from 5.83% in singly infected tissues to 27.9% in doubly infected tissues, providing evidence of the enhancement of CymMV titer. While most vsRNA features remained unchanged in double inoculations, three additional prominent ORSV vsRNA hotspot peaks were observed. In silico prediction revealed Phalaenopsis transcript hotspots that are potential targets for vsRNA are also likely to be involved in symptom formation. The virus infection also modulated miRNA expression—for example, miR156, miR168, miR894 were up regulated and miR398, miR408, miR528 were down regulated after CymMV or ORSV infection. These infection responsive miRNAs participate in a broad spectrum of cellular processes like hormone and metabolite assimilation, signal transduction, and oxidative stress calibration. Taken together, the deep sequencing provided a global profile of vsRNAs and miRNAs in Phalaenopsis under CymMV and ORSV infection. Further research should provide valuable insights into small RNA-mediated virus-plant interactions.
3
Introduction
The Orchidaceae is an immense family composed of more than 800 genera, about 25,000 native species and over 110,000 cultivars (Sheehan, 2003). Ornamental orchids commonly seen in the market include Cattleya, Cymbidium, Dendrobium, Oncidium, Paphiopedilum, and especially Phalaenopsis (the moth orchid). People have long appreciated the fascinating array of colors, elegant flower shapes, ease of growing moth orchids indoors, as well as the long duration of their inflorescence. Phalaenopsis are native throughout Sri Lanka, southern India, New Guinea, northern Australia, China, Vietnam, the Philippines and other tropical and subtropical mountain areas. Taiwan is the habitat of two native Phalaenopsis species, including P. aphrodite and P. equestris (李,2005). Taking advantage of the optimal climate for growing Phalaenopsis, and aided by fine cultivation protocols, Taiwan is well situated as a major exporter. The Council of Agriculture has chosen the Phalaenopsis as one of the top four important export commodities in 2004, and which has become third in wholesale value of exported agricultural products since 2008 (楊, 2010). However, Taiwan Phalaenopsis exports are now facing competition with other countries such as China, America, and especially the Netherlands. A major challenge is virus infection, since virus indexing techniques have not been routinely applied at the early stage of orchid industry development in Taiwan (張, 2006).
Of numerous orchid pests and diseases, viruses are the most troublesome and no efficient measures can be used to control virus diseases. Virus infections can result in retarded and weakened growth of orchid plants, even though the plants appear symptomless. In addition, the hidden risk of latent infection after shipment also poses a
4
thorny problem (張, 2007). Since large-scale cultivation via tissue culture has become the trend in orchid production, virus infections through mechanical transmission have become a major threat (鄭等, 2008). Of more than 50 orchid-infecting viruses, Cymbidium mosaic virus (CymMV) and Odontoglossum ringspot virus (ORSV) are the most prevalent worldwide. Isolation of CymMV and ORSV was first reported in the 1950s (Jensen, 1950;Jensen and Gold, 1951). Co-infection with these two viruses is commonly observed in orchid nurseries, and synergistic effects between CymMV and ORSV have long been reported (Ajjikuttira et al., 2005). Several studies have recorded the characteristics of virus particles and their induced symptoms, and also decoded the complete genome sequences (Ryu and Park, 1995;Wong et al., 1997). However, most studies aimed to establish rapid virus diagnosis techniques (Chen et al., 2010; Chia et al.,1992; Eun and Wong, 1999; Hu and Wong, 1998; Lee et al., 2011; Lee and Chang, 2010; Ryu et al., 1995; Seoh et al., 1998; Tanaka et al., 1997), few have probed the molecular determinants of CymMV and ORSV pathogenesis (Ajjikuttira et al., 2005;
Hu et al., 1998;Lu et al., 2009) and the mechanism of infection remains largely unknown.
Recently, pathogen derived resistance is one of the interesting areas in biotechnological research on Phalaenopsis. Investigating the interaction between CymMV or ORSV and host plants will provide useful knowledge for establishing a foundation of virus-proofing techniques and generating virus-resistant plants.
Sequence-specific RNA silencing has been revealed as one of a number of general antiviral mechanisms in plants (Angell and Baulcombe, 1997; Dougherty, 1994; Ruiz et al., 1998; and reviewd in Llave, 2010). RNA silencing is triggered by double-stranded
5
or self-folding RNAs that are processed into small and functioning fragments by RNase III-type DICER enzymes. The small RNAs are mainly categorized in plants into small interfering RNAs (siRNAs) or microRNAs (miRNAs) according to their origin and biogenesis pathways (reviewd in Naqvi et al. 2009 and Vazquez et al., 2010).
Virus-derived small interfering RNAs (vsRNAs) usually accumulate at high levels in infected plant tissues. VsRNAs are further recruited into RNA-induced silencing complex (RISC) and guiding sequence-specific viral RNA cleavage. Furthermore, recent studies have provided evidence that RNA silencing may in turn play an important role in disease induction, since plant endogenous miRNAs may be dysregulated upon virus infection, and vsRNAs are also potentially targeted to plant mRNAs rather than viral RNAs (Moissiard and Voinnet, 2006; Qi et al., 2009; and reviewd in Wang et al., 2012).
This study investigated the profile of small RNAs in P. amabilis plants with ORSV and CymMV single- or mixed-infection and focused on 1) analyzing the characteristics of CymMV and ORSV vsRNAs, such as abundance, size classes, strand polarities, and hotspot distribution along the viral genome, and 2) the expression levels of several conserved miRNAs which respond to virus infection. Possible roles of these small RNAs in defense and counter-defense interactions are also explored by predicting potential Phalaenopsis mRNA targets of miRNAs and vsRNAs. Furthermore, the study also addresses the synergistic effect between CymMV and ORSV.
6
Literature Review 1. The incidence of virus diseases in orchids
Virus diseases have long been a serious problem in orchid cultivation. Plant viruses are infectious obligate parasitic nanoparticles constituted by an RNA or DNA genome encapsidated inside its coat protein (CP) shells. More than 50 viruses are reported to infect orchids and about 30 of them have been described in detail (Ajjikuttira and Wong, 2009). Six of those 30 viruses have been observed in Taiwan, including Capsicum chlorosis virus (CaCV), Carnation mottle virus (CarMV), CymMV, Cucumber mosaic virus (CMV), ORSV, and Phalaenopsis chlorotic spot virus (PhCSV). Of these, CymMV and ORSV are the most prevalent and cause serious economic loss (鄭等, 2008; Zettler et al., 1990).
1.1. Cymbidium mosaic virus (CymMV)
CymMV belongs to the family Alphaflexiviridae, genus Potexvirus. It has a positive-sense single-stranded RNA [(+)-ssRNA] monopartite genome with five open reading frames (ORFs) encoding a 160-kilo Dalton (kDa) RNA-dependent RNA polymerase (RdRp), three triple-gene-block proteins (TGBps), and a 24-kDa CP. The TGBps have been recognized as movement proteins (MP). The genomic RNA is 5’-capped and about 6200 nucleotides (nt) in length (e.g. GenBank accession no.
AB197937, AF016914, AM055720, AY571289, EF125178, EF125179, EF125180, EU314803, HQ681906, NC_001812), excluding the 3’-poly(A) tail (Ajjikuttira and Wong, 2009; Wong et al., 1997). The non-enveloped, flexuous and filamentous virions
7
with a length of ~480 nm and a width of ~13 to 18 nm are constituted by 5.6% RNA and 94% protein. The axial canal is indistinct. The thermo inactivation point (TIP) is about 60 to 70o C and the longevity in vitro (LIV) is reported to be around 25 days. It is easily transmitted through mechanical wounds, since the virions are quite stable, but no known vectors have been reported to date (Ajjikuttira and Wong, 2009; Frowd and Tremaine, 1977; Jenson and Gold, 1955; Murakishi, 1958). CymMV was first found in Cymbidium orchids (Jenson, 1950), and its natural hosts include several orchid families such as Phalaenopsis, Cymbidium, Cattleya, Epidendrum, Laelia, Laeliocattleya, Oncidium, Vanda, Vanilla and Zygopetalum. Local lesions or patches are formed in CymMV-inoculated leaves of Cassia occidentalis and Chenopodium quinoa, thus these plants are commonly used as indicators. In addition, Nicotiana benthamiana are systemic hosts of CymMV and are often used as experimental hosts for inoculation assays and virion propagation (Ajjikuttira and Wong, 2009; Faccioli and Marani, 1979;
Hiruki et al., 1980; Jenson and Gold, 1955; Kado and Jenson, 1964; Murashiki, 1958).
1.2. Odontoglossum ringspot virus (ORSV)
ORSV belongs to the Tobamovirus group and had previously been regarded as Tobacco mosaic virus orchid strain (TMV-O). ORSV was reclassified as a separate species after nucleotide sequence analysis and other phylogenic evidence accumulated (Edwardson and Zettler, 1988). It has a (+)-ssRNA monopartite genome with three ORFs. ORF1 encodes 126- and 183-kDa RdRp proteins, the latter expressed by read-through strategy. Other ORFs encode 34-kDa MP and 18-kDa CP (Ajjikuttira and Wong, 2009; Ryu and Park, 1995). The genomic RNA of ORSV is about 6600 nt in length (e.g. GenBank accession no. AY571290, DQ139262, NC_001728, ORU3458),
8
with a 5’-capped structure but lacking the 3’-poly(A) tail. Instead, its 3’-untranslated region (3’-UTR) is comprised of a tRNA-like structure (TLS) in the terminus and three upstream consecutive homologous regions, forming a pseudoknot (PK) chain (Chng et al., 1996). The virus particles are rigidly rod-shaped, with a length of ~300 nm and a width of ~18 nm and comprised of ~5% RNA. ORSV is inactivated at about 90oC and is mechanically transmitted, without recognized vectors (Ajjikuttira and Wong, 2009;
Corbett, 1967; Edwardson and Zettler, 1986; Jenson and Gold, 1951; Kado et al., 1986;
Paul et al., 1965). Previous study has shown ORSV virions can retain infectivity for more than ten years (Inouye, 1983). ORSV was first reported infecting Odontoglossum grande (Jensen and Gold, 1951). Other natural hosts include Odontoglossum, Cymbidium, Cattleya, Dendrobium, Epidendrum, and Zygopetalum. For biological diagnosis of ORSV infection, Gomphrena globosa and C. quinoa can be used as indicators by forming local lesions on the inoculated leaves. N. benthamiana is one of the systemic hosts of ORSV, showing mild mosaic and distorted emerging leaves after infection and is commonly used as an experimental host (Ajjikuttira and Wong, 2009;
Navalinskienë et al., 2005; Wisler et al., 1979).
1.3. Symptoms and pathogenesis of CymMV and ORSV in orchids
Variable symptoms of CymMV and ORSV have been observed in orchids and are variable by cultivars, growing stages, and environmental factors, such as seasonal changes. Despite this variation, classical symptoms shoed chlorotic spots formed on leaves of CymMV singly infected Phalaenopsis; the spots were 0.2-0.5 cm in diameter at first, then gradually enlarged or became necrotic at later stages (陳等, 2006).
Common symptoms of CymMV in orchids include chlorosis streak or stripe, necrosis
9
spots or line patterns on leaves and stems. In addition, the necrotic type symptoms in florescence are specific to CymMV (張, 2006; 陳等, 2006). The most prevalent symptoms caused by ORSV infection include irregular chlorotic patches, ringspots, mosaic, mottle, streak, color breaking, and necrosis in orchids, with mosaic and streak.
However, the incidence of symptom formation with single infection by ORSV is much lower than when infected with CymMV alone. In particular, ORSV symptoms were minimal in juvenile Phalaenopsis and Oncidium before the plants grew up and flowered (張, 2006).
Co-infection with both CymMV and ORSV has been frequently observed in orchid nurseries, and synergism exists between the two viruses (張, 2006; Hadley et al., 1987).
Latent virus infection could result in retarded growth and reduced flower quality, even though infected plants are sometimes symptomless. An increase of 65% and 21% in inflorescence size and photosynthetic capacity, respectively, were obtained in virus-eradicated Oncidium Gower Ramsey, demonstrating the impact of virus infection on plant growth (Chia and He, 1999).
Infection with CymMV was detected in rub-inoculated leaves of Dendrobium × Jaquelyn Thomas 'Uniwai Supreme' and 'Uniwai Prince' nine days post inoculation (dpi) on average. Systemically movement of CymMV was first detected in the root (about 17 dpi) and then in leaves (about 20 dpi). Slash (13/15) and cut (13/22) inoculation was less efficient than rub inoculation (41/41) for CymMV (Hu et al., 1994). ORSV was inoculated only through rub-inoculation method and systemic movement was not detected in the dendrobiums, indicating CymMV is more easily spread through cultivation practices than ORSV (Hu et al., 1994). Wong et al. (1994) conducted a
10
survey of CymMV and ORSV incidence in Singapore and found 54.6% plants in orchid farms infected with CymMV, 4.0% with ORSV and 14.2% with both viruses. On the other hand, in the Singapore Botanical Garden, 34.5% of orchids were detected to have CymMV, 0.3% with ORSV and 8.3% had mixed infection. These studies indicate higher incidence of CymMV which might suggest a greater susceptibility to CymMV than to ORSV in orchids. Furthermore, frequent flower cutting in farms increased the risk of mechanical transmission (Wong et al., 1994).
1.4. Molecular mechanisms of the synergism between CymMV and ORSV
Infection with multiple viruses from different groups often shows syngeristic interaction, and results in intensified symptoms and higher amounts of virus accumulation. Synergism has also been noticed in double infections of potexvirus and tobamovirus (Goodman and Ross, 1974; Taliansky et al., 1982). Severe necrosis and sunken patches more frequently occur in CymMV and ORSV mixedly infected orchids than singly infected ones (鄭等, 2008). The synergism between CymMV and ORSV has also been observed in transfected protoplasts (Hu et al., 1998). In protoplasts isolated from Dendrobium ‘Sonia’ petals, CymMV genomic viral RNA was detectable six hours after electroporation and reached a maximum accumulation at 18 hours. ORSV genomic RNA was also detected beginning six hours after electroporation but reached a maximum accumulation later at 24 hours. Accelerated ORSV viral RNA accumulation in co-infected protoplasts was observed, taking only half the time (12 h) to reach the maximum accumulation. The accumulation of both sense and antisense strands of CymMV and ORSV viral RNA was increased in mixed infections. The positive and negative strands of ORSV showed approximately 6- and 12-fold accumulation
11
compared to single infected protoplasts, respectively. The negative strands of CymMV also increased 5-fold in co-infected protoplasts compared to singly infected ones, indicating increased replication of both CymMV and ORSV during co-infection (Hu et al., 1998).
Viral MP and CP are often required for the virus movement through plants. Several studies demonstrate taxonomically distinct viruses display reciprocal transport functions.
For example, TMV MP can complement the defective cell-to-cell movement of CMV, Barley stripe mosaic virus (BSMV), Potato virus X (PVX) and other unrelated viruses (Cooper et al., 1996; Morozov et al., 1997; Solovyev et al., 1996). The cell-to-cell movement of an MP-deficient CymMV was restored in ORSV MP-expressing transgenic N. benthamiana. Similarly, cell-to-cell movement of a MP-deficient ORSV was rescued in CymMV TGBp1-expressing plants, suggesting reciprocal functions between CymMV TGBps and ORSV MP. On the other hand, systemic movement of CP-deficient CymMV was supported by ORSV CP. However, CymMV CP did not support the long-distance movement of CP-deficient ORSV (Ajjikuttira et al., 2005).
In summary, more pronounced disease symptoms are often observed in double infections and may be related to increased accumulation of either or both viruses. (Hu et al., 1998; Ajjikuttira et al., 2005). Functional complementation of MPs and CPs of CymMV and ORSV facilitates virus movement and may enhance the susceptibility to the other virus after infection with either one (Ajjikuttira et al., 2005). However, though it is generally agreed that intensified symptom formation occurs during co-infection as a consequence of the synergism between CymMV and ORSV, the molecular details of virus accumulation and symptom formation in double infections have not been determined.
12
2. The role of RNA silencing in plant-virus interactions 2.1. RNA silencing pathways in plants
The central dogma of molecular biology describes the cascade of transferring sequential genetic information through transcription of DNA to RNA, and translation of RNA to protein. However, researchers have identified several regulatory non-coding RNAs which are not translated into proteins. Non-coding RNAs, such as transfer RNA (tRNA) and ribosomal RNA (rRNA), were first characterized in the 1960s and 1980s, respectively, whereas small RNAs were not noticed and studied intensively until the 1990s (Fire et al., 1998; Napoli et al., 1990; Romano and Macino, 1992; van der Krol et al., 1990a and 1990b). Small RNAs are 20 to 40 nt-long molecules present in most eukaryotes that regulate gene expression in a sequence-specific matter. Various small RNA classes, such as micro RNA (miRNA), small-interfering RNA (siRNA), and piwi-interacting RNA (piRNA) have been identified and categorized on the basis of their biogenesis and precursor structure. In plants, the best described classes are siRNA and miRNA (e.g. Bazzini et al., 2007; for recent reviews, see Naqvi et al., 2009).
Biogenesis of siRNAs and miRNAs relies on the overlapping requirement of dsRNA, as precursor and conserved protein families are involved. The major difference is that miRNAs are derived from imperfectly base-paired hairpin-loop transcripts of miRNA genes generated by RNA polymerase II, whereas siRNAs are generated from longer double-stranded RNA precursors, both endogenously or exogenously (Elbashir et al., 2001; Vazquez et al., 2010). After the short duplexes are processed from precursors by RNase-III DICER-LIKE (DCL) enzymes, and methylated by HEN1 methyltransferase, they are unwound by ARGONAUTE (AGO) effectors, then
13
assembled into RNA-induced silencing complex (RISC) to provide sequence-specific targeting (Schauer et al., 2002; Papp et al., 2003; Tang et al., 2003; Bernstein et al., 2001; Hamilton and Baulcombe, 1999; Hammond et al., 2000). They then mediate either transcriptional regulation such as DNA methylation and histone modification, or post-transcriptional regulation through mRNA cleavage or translational inhibition (Aukerman and Sakai,2003; Chen, 2004; Doench et al., 2003; Hammond et al., 2001;
Hutvagner and Zamore, 2002). The consequently repressed expressions of miRNA/siRNA-targeted genes are thus subject to the name “gene silencing,” “RNA silencing” or “RNA interference (RNAi)” (Jones-Rhoades et al., 2006; Mallory and Vaucheret, 2010; Molnar et al., 2011; Naqvi et al., 2009; Voinnet, 2009; Zamore et al., 2000).
SiRNA-mediated antiviral RNA silencing plays a critical role in plant-virus interactions. Double-stranded viral RNAs, such as replication intermediates or self-annealing hairpin structures, can serve as templates for DCL processing. The virus-derived siRNAs (vsRNAs) are recruited by RISC and guide RISC to cleave single-stranded viral RNAs. In addition, plant endogenous RNA-dependent RNA polymerases (RDRs) are also involved in synthesizing dsRNAs from single-stranded viral RNAs. The dsRNAs again serve as substrates for the DCL-dependent formation of secondary vsRNAs, and thus amplify the antiviral cascade (Angell and Baulcombe, 1997; Dougherty, 1994; Ruiz et al., 1998; Blevins et al., 2006; Llave, 2010).
Furthermore, several studies have revealed siRNAs are mobile molecules and spread throughout the plant via vascular bundles (Palauqui et al., 1997; Jorgensen et al., 1998;
Voinnet et al., 1998; Vance and Vaucheret, 2001), suggesting vsRNAs may act as signal molecules and confer systemic antiviral silencing throughout the plants.
14
In addition, numerous plant viruses have evolved viral suppressors of RNA silencing (VSRs) as a counter-defense strategy, and even vsRNAs are mediators of viral diseases (Burgyán and Havelda, 2011; Lu et al., 2008). Moreover, VSRs may interfere with the host miRNA pathway and dysregulate normal plant development, whereas vsRNAs highly complementary to a host sequence could also induce silencing of the host genes (Dunoyer and Voinnet, 2005; Wang et al., 2004; Wang et al., 2012; Qi et al., 2009). Collectively, the RNA silencing machinery plays an important role in regulating plant growth and also constitutes a complex layer of defense and counter-defense interactions between plants and viral pathogens.
2.2. Profiling characteristics of virus-derived small interfering RNAs (vsRNAs) High accumulation of vsRNAs is frequently observed in virus-infected plants and is recognized as a hallmark of antiviral RNA silencing machinery. Several proteins are involved in the antiviral silencing mechanism. Thoroughly studied in Arabidopsis thaliana, vsRNAs are mainly 21-nt and 22-nt through processing of DCL4 and DCL2, respectively (Blevins et al., 2006). Different AGOs associate with vsRNAs based on the 5’-terminal nucleotide classes and guide different modes of silencing pathways. For example, AGO1- and AGO10-containing RISC complexes often mediate mRNA cleavage or translational inhibition, while AGO4 mainly directs transcriptional silencing through DNA methylation (Hutvagner and Simard, 2008; Mi et al, 2008; Yan et al., 2010). Consequently, characteristics such as size, strand polarity, hotspots of DCLs processing and 5’-nucleotide preference of vsRNAs are associated with these proteins.
High throughput sequencing techniques provide an efficient way to analyze small RNAs and have been used to profile vsRNAs. Some features of vsRNAs produced from
15
diverse groups of viruses in different hosts are described below.
2.2.1. Functions of DCL proteins and size distribution of vsRNAs
Specific size classes of small RNAs are produced via different DCLs. DCL1 and DCL4 generate 21-nt small RNAs. DCL2 and DCL3 process 22-nt and 24-nt small RNAs respectively (Blevins et al., 2006; Vaucheret, 2006). Viral RNAs are differentially processed by multiple DCL proteins. Since DNA viruses tend to multiply in the nucleus but RNA viruses normally replicate in the cytoplasm, the major DCLs mediating vsRNA production from DNA or RNA viruses are different (Blevins et al.,2006; Wang et al., 2012). As shown in A. thaliana mutants, all four types of DCLs target DNA viruses such as Cabbage leaf curl virus (CaLCuV) and Cauliflower mosaic virus (CaMV), whereas DCL4 and DCL2 mainly affect RNA viruses such as CMV, Tobacco rattle virus (TRV), and Oilseed rape mosaic virus (ORMV) (Blevins et al., 2006;
Bouché et al., 2006; Deleris et al., 2006).
High throughput sequencing has revealed similar profiles of vsRNA size class distribution. In most cases, vsRNAs from RNA viruses are mainly 21-nt, followed by 22-nt class. For instance, a number of viruses have been reported as generating more 21-nt vsRNAs, including TMV, TRV and CMV in Arabidopsis, Bamboo mosaic virus (BaMV), Tunip mosaic virus (TuMV), Pepper mild mottle virus (PMMoV) and PVX in N. benthamiana, Watermelon mosaic virus (WMV) and Melon necrotic spot virus (MNSV) in Cucumis melo, Tomato yellow leaf curl virus (TYLCV) in Solanum lycopersicum and several grape viruses such as Grapevine rupestris stem-pitting associated virus (GRSPaV), Grapevine fleck virus (GFkV), Grapevine red globe virus (GRGV), Grapevine asteroid mosaic-associated virus (GAMaV) and Grapevine
16
rupestris vein feathering virus (GRVFV) in Vitis vinifera Pinot Noir (Donaire et al., 2009; Lin et al., 2010; Pantaleo et al., 2010; Qi et al., 2009). Furthermore, the patterns of vsRNA size distribution of BaMV and Rice stripe virus (RSV) did not differ according to host plant species (Lin et al., 2010; Xu et al., 2012; Yan et al., 2010). The only exception was Cymbidium ringspot virus (CymRSV), with more 22-nt vsRNAs over 21-nt found in infected N. bentahmiana, which is possibly related to the activity of silencing suppressor P19 of CymRSV to sequester 21-nt sRNAs (Donaire et al., 2009).
2.2.2. Preference of 5’ terminal nucleotide of vsRNAs
Since the 5’ nucleotide identity dictates the loading of vsRNAs into a particular AGO effector (Czech and Hannon, 2011; Kim, 2008), the base composition of vsRNAs 5’ termini may infer different downstream pathways involved in RISC-mediating silencing mechanisms. Most viruses studied revealed preference for 5’-uridine (U) or 5’-adenosine (A). The sense-stranded vsRNAs with a 5’-U were more abundant for CMV, CymRSV, MNSV, PMMoV, RSV, TRV and TYCV, while 5’-U or A were observed equally for WMV, TMV, TuMV and PVX sense-stranded vsRNAs (Donaire et al., 2009). When it comes to antisense-stranded vsRNAs, 5’-U, A, or 5’-cytosine (C) were similarly represented for most viruses (Donaire et al., 2009; Qi et al., 2009). In particular, grape viruses such as GRSPaV, GFkV, GRGV, GAMaV and GRVFV were observed to favor 5’-C (Pantaleo et al., 2010). Nonetheless, a clear tendency to avoid 5’-guanidine (G) was observed in the cases above. The diverse categories of 5’-nucleotide of vsRNAs also suggest that vsRNAs may potentially be recruited into multiple AGO-containing complexes (Donaire et al., 2009).
17
2.2.3. Strand polarity and hotspot distribution of vsRNAs along the virus genome Double-stranded viral RNA formed by direct hybridization between (+)- and (–)-strand replication intermediates or synthesized by host RDR from single-stranded viral RNA can serve as substrates of DCL enzymes (Blevins et al., 2006; Ding, 2010;
Wang et al., 2012). VsRNA pools with near equal abundance of both sense- and antisense-polarities have been profiled in TuMV- and PVX-infected N. benthamiana, WMV- and MNSV-infected melons, and TYLCV-infected tomato plants (Donaire et al., 2009). Furthermore, other evidence suggests hairpin structures formed by self-annealing within single-stranded viral RNA can also serve as DCL processing templates (Dunoyer and Voinnet, 2005; Molnár et al., 2005). In fact, biased strand polarity and asymmetrical distribution of vsRNAs along virus genomes are often observed. Profiles of TMV, TRV and CMV in Arabidopsis, GRSPaV in grapevine, also CymRSV and PMMoV in N.
benthamiana presented more (+)-stranded vsRNAs (Donaire et al., 2009; Pantaleo et al., 2010; Qi et al., 2009), while GFkV, GRGV and GAMaV vsRNAs were mostly (-)-stranded in co-infected grapevine (Pantaleo et al., 2010).
Furthermore, differential distribution of vsRNAs has been observed with the same virus in different hosts. Negative-strand predominance of BaMV vsRNAs has been found in N. benthamiana, whereas slightly higher sense-polarity presented in A.
thaliana. In addition, while most vsRNAs mapped to the CP and 3’-UTR regions in infected N. benthamiana, vsRNAs profiled in A. thaliana were largely generated from the RdRp region of the 5’ half of the BaMV genome (Lin et al., 2010). Although RSV vsRNAs generally possessed (+)-polarity in both Oryza sativa and N. benthamiana plants, most vsRNAs identified in rice were generated from RSV RNA 3 and 4, whereas RNA 4 is the major source of vsRNAs in tobacco (Xu et al., 2012).
18
2.3. Interactions between virus-modulated RNA silencing and the pathogenesis of viruses in plants
2.3.1. Roles of viral suppressors of RNA silencing (VSRs) in symptom induction As a counter-defense strategy, many viruses evolved VSRs during evolution. A number of studies have shown multiple steps of disrupting RNA silencing pathways of certain VSRs. For example, tombusvirus P19 sequesters double-stranded siRNAs, potyvirus Hc-Pro and P38 interfere with RISC assembly, and cucumovirus 2b inhibits the slicing activity of AGO1 (reviewd in Burgyán and Havelda, 2011). Silencing suppressors can contribute to viral symptoms via indirectly helping virus accumulation or directly modifying endogenous small RNA-regulated pathways (Silhavy and Burgyán 2004). Diminished virus accumulation has been found in the dark-green zones of the mosaic patterns compared to the light-green zones of infected leaves of Tamarillo mosaic virus (TaMV) and Tomato mosaic virus (ToMV) (Moore et al. 2001; Hirai et al.
2008). The mottling seed coat patterns in CMV-infected Glycine max ‘Jack’ seeds has been shown to result from suppression of chalcone synthase (CHS) gene silencing by the suppressor protein 2b (Senda et al., 2004). CMV infection dysregulated the star-type color pattern of Petunia hybrida 'Red Star' flowers mediated by natural RNA silencing of CHS in the white sectors, significantly increasing the CHS mRNA and turning all the white patterns red (Koseki et al., 2005). Viral symptom-like phenotypes are often observed in transgenic plants that express VSR. For instance, transgenic Arabidopsis expressing Tomato bushy stunt virus (TBSV) P19, TuMV P1/Hc-Pro or Turnip crinckle virus (TCV) CP exhibited serrated rosette leaves (Chapmen et al., 2004).
Dysmorphogenic and sterile male inflorescences with narrow sepals and protruding
19
internal whorls also displayed on P19 and HcPro expressing plants (Dunoyer et al., 2004; Mallory et al., 2002).
2.3.2. Symptom induction through vsRNA-mediated host gene regulation
In contrast to the viral RNA clearance activity of plant RNA silencing pathways, studies have also demonstrated that mRNA of host genes are conversely targeted by virus- or subviral agent-derived siRNA-guided RISC complexes, if high sequence complementarity exists between pathogen and host mRNA (Llave, 2004; Wang et al., 2004). Several studies have proven that some viroids and satellites direct RNA silencing against physiologically important host genes, thus causing disease symptoms. For example, viroid symptoms were observed in transgenic tomato plants expressing Potato spindle tuber viroid sequence hairpin RNA (Wang et al., 2004). The symptom determinant sequence of CMV Y satellite RNA (Y-sat) was complementary to a 22-nt region of a chlorophyll biosynthetic gene ChlI in tobacco, thus inducing bright yellowing symptoms via indirect repression of chlorophyll accumulation (Shimura et al.
2011; Smith et al. 2011). A severe chlorotic symptom called peach-calico resulted from retarded expression of a chloroplastic heat-shock protein 90 via Peach latent mosaic viroid (PLMVd)-derived siRNA-mediated RNA silencing (Navarro et al., 2012).
Moreover, several CaMV vsRNAs have near-perfect sequence complementarity to Arabidopsis mRNAs and therefore, may direct silencing of these genes (Moissiard and Voinnet 2006). Qi et al. have predicted potential target genes of TMV crucifer strain (TMV-Cg) and the cleavage of two host genes, the cleavage and polyadenylation specificity factor (CPSF30) and the translocon-associated protein α (TRAPα), have been verified by 5’ rapid amplification of cDNA ends (5’-RACE) assays (Qi et al. 2009).
20
2.3.3. Infection induced dysregulation of miRNA expression and its association in plant-virus interactions
Plant endogenous miRNAs are broadly involved in plant development, signal transduction, protein degradation, and biotic or abiotic stress responses. Since miRNAs and siRNAs share many common features, it has been proposed that the miRNA-mediated gene regulation pathways may also be perturbed by virus infection (Lu et al., 2008; Silhavy and Burgyán, 2004). Bazzini and colleagues used beta-glucuronidase (GUS) reporter vectors to reveal elevated activity of miR164a promoter upon Oilseed rape mosaic virus (ORMV) and TMV infection, and up regulation of miR164 was verified by RT-PCR in virus infected Arabidopsis (Bazzini et al., 2009). Bazzini likewise observed disease-like phenotypes in TMV CP and MP co-expressing transgenic N. tabacum along with up regulated expression of miR156, miR164, miR165 and miR167, resembling TMV-infected plants and leading to the hypothesis that a complex formed between MP and CP may dysregulate miRNA expression and result in disease symptoms (Bazzini et al., 2007). An array of miRNAs, including miR319 and miR172 in tomato ‘Pusa Ruby’, were responsive to Tomato leaf curl new delhi virus (ToLCNDV) infection and may be involved in the formation of leaf curl symptoms (Naqvi et al., 2010). Reduced levels of miR159, miR165/166 and miR171 were found in cassava and N. benthamiana infected with geminivirus African cassava mosaic virus Cameroon Strain (ACMV) with severe symptoms, whereas the reduction was absent in plants infected with another geminivirus, East African cassava mosaic Cameroon virus (EACMCV), which induced only mild symptoms (Chellapan et al., 2005). In Brassica rapa, miR1885 was identified as a novel miRNA and was
21
specifically induced upon TuMV infection, mediating cleavage of TIR–NBS–LRR class R gene transcripts thus breaking the host’s disease resistance (He et al., 2008). It has also been shown that the phytocyanin transcript targeting miR408 was specifically down regulated at late stage of TMV infection in tobacco, and the repressed expression may impact several metabolites involved in respiratory pathways (Bazzini et al., 2011).
Induction of miR168 expression has been recognized as a common response to virus infection (Lang et al., 2011; Várallyay et al., 2010). CymRSV viral RNA and increased miR168 accumulation displayed an overlapping distribution, and this up-regulation of miR168 was absent in suppressor P19-deficient CymRSV-infected Arabidopsis. In contrast, Várallyay and colleagues observed increased accumulation of AGO1 mRNA in plants infected with either wild-type or P19-mutated CymRSV, suggesting AGO1 gene expression was elevated as an antiviral mechanism. In contrast, virus-induced up-regulation of miR168 was due to VSR activity (Várallyay et al., 2010).
In conclusion, these studies indicate that virus infection affects expressions of host genes through vsRNA-mediating RNA silencing or dysregulated miRNA pathways, and is thus involved in pathogenesis and development of plant diseases.
22
Materials and Methods 1. Maintenance of CymMV- and ORSV-free plants 1.1. Plant materials and growing conditions
Mature mericlone P. amabilis plants in 7.5-cm pots were purchased from I-hsin Biotechnology Inc. (Chia-yi, Taiwan). CymMV and ORSV-free plants were examined by reverse transcription-polymerase chain reaction (RT-PCR) and grown in natural sunlight phytotron at the Genome Research Center, Academia Sinica. Growth conditions were day/night temperature of 25/20o C with shade cloths used to maintain maximum light intensity of 300 μmol·m-2·s-1 photosynthetic photon flux density (PPFD). Plants were fertigated with 15N-2.2P-12.5K (Peters 15-5-15 Ca Mg, Scotts Co., Marysville, Ohio, USA) and leached using distilled water as needed.
1.2. Detecting virus infection in newly purchased plants
For detecting CymMV and ORSV infection by RT-PCR, total RNA was purified from 0.1 g lower-positioned leaves of each plant. Disposable razor blades and gloves were used to cut off the sampled tissues to prevent contamination if some plants were virus-infected.
1.2.1. Extracting total RNA from leaf tissues
A CTAB method modified from the pine tree method (Chang et al., 1993) was used to purify total RNA from Phalaenopsis leaves. About 0.1 g fresh or frozen leaf tissues were gounded into fine powder with liquid nitrogen, mixed vigorously with 1 mL grinding buffer [0.55 mM CTAB, 0.1 M Tris (pH7.5), 0.2 M EDTA, 20 mM NaCl, 0.03
23
mM PVP40, and 2% β-mercaptoethanol] and incubated it at 65o C for 15 min. The mixture was centrifuged at 3000 xg for 10 min at room temperature, the supernatant was then mixed with an equal volume of CI (chloroform : isoamylalcohol = 24 : 1), then centrifuged at room temperature again at 12000 xg for 10 min. Total RNA were precipitated by adding one third volume of 8M LiCl into the supernatant and chilling it at -20 oC overnight. The RNA pellet was obtained by centrifuging it at 12000 xg, 4o C for 30 min, then washing it with pre-chilled 70% ethanol and dissolving it in RNase-free water. The concentration of total RNA was determined by NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA).
1.2.2. Reverse transcription-polymerase chain reaction (RT-PCR)
RT-PCR was performed using primer pairs specific for CymMV/ORSV coat protein (CP) genes or RNA-depedent RNA polymerase (RdRp) genes. Details of the primers are listed in Table S1. For the RT reaction, 500 ng total RNA was mixed with 1 μL of 10 μM reverse primer and 2 μL of 10 mM dNTP, and RNase-free water was added to bring up the volume to 12 μL. The mixture was incubated at 70o C for 5 mins then quenched at least 5 mins on ice. 4 μL 5X first strand buffer, 2 μL 0.1M DTT, 0.5 μL RNasin® (40 U·μL-1, Promega, Madison, WI, USA), 0.5 μL Super-ScriptTM III Reverse Transciptase (200 U·μL-1, Invitrogen, Carlsbad, CA, USA), and 1μL RNase-free water were added to the solution to produce a final volume of 20 μL. After incubating at 42o C for 1 hour, the cDNA can be used as template for amplification in PCR.
PCR amplification was performed in a 25 μL reaction mix containing 2 μL RT product, 0.5 μL of each 10 μM forward and reverse primers, 12.5 μL Taq 2X Master Mix (GeneMark, Taichung, Taiwan) and 9.5 μL RNase-free water. The Tpersonal
24
Thermocycler (Biometra, Göttingen, Germany) was set to the following program: 1) 42o C, 45 min, 2) 95o C, 5 min, 3) 95o C, 30 sec, 4) 55o C, 30 sec, 5) 72o C, 30 sec, 6) repeat step 3 to 5 for 34 cycles (total 35 cycles), 7) 72o C, 7 min, 8) 4o C, pause. The PCR products was then analyzed by 1% agarose gel electrophoresis with 1 Kb Plus DNA Ladder (Invitrogen, Carlsbad, CA, USA) as size markers.
2. Detection of CymMV and ORSV spread in P. amabilis 2.1. Experimental locations
When conducting inoculation assays, plants were moved from the phytotron to a growth room in the basement of the Institute of Plant and Microbial Biology Building, Academia Sinica. Growth conditions were set to a constant temperature of 28o C, 70%
relative humidity, and the light period from 0600AM to 0800PM with a light intensity of 100 μmol·m-2·s-1 PPFD supplied by fluorescent lights.
2.2. Virus source
The purified virion and viral RNA were used in inoculation assays. CymMV and ORSV isolates were obtained through three successive single lesion passages on C.
quinoa (Lin et al., unpublished). After propagating in N. benthamiana, virus particles were purified using a method described previously (Lin and Chen, 1991; Lin et al., 1992). Infected leaves were ground in 4 volume of borate buffer (0.5 M boric acid, 1 mM EDTA, and 0.5% β-mercaptoethanol, pH 9.0) and filtrated through micracloth (Calbiochem, Billerica, MA, USA). The filtrate was centrifuged at 8,800 rpm for 22 min in a JA-10 rotor (Beckman Coulter, Brea, CA, USA). K2HPO4 and CaCl2 were then added to the supernatant to reach a final concentration of 40 mM. The mixture was
25
stirred well for 30 min at 4° C and was then centrifuged at 8,800 rpm for 22 min. PEG 6000 was added to the supernatant to a final concentration of 6% and stirred for 60 min at 4° C. The pellet was obtained after centrifuging at 8,800 rpm for 18 min, and resuspended in 0.05 M borate buffer (0.05 M boric acid and 1 mM EDTA, pH 8.0).
Triton X-100 was added to a final concentration of 2% and the mixture was stirred for 20 min at 4° C. After centrifuging at 6,000 rpm for 5 min, the supernatant was layered under a 5-ml cushion of 20% sucrose. Ultrahigh speed centrifugation of 43,000 rpm for 1 hr 40 min was carried out by a RP70T rotor (Hitachi, Tokyo, Japan). The pellet was then dissolved in 5 ml 0.05 M borate buffer at 4° C overnight and yield of purified virions was determined by ultraviolet absorption, assuming the extinction coefficient (ε)
= 3 for CymMV and ORSV at 260 nm (Choi et al., 2002; Frowd and Tremarine, 1977).
Viral RNA was isolated from purified virions as previously described (Lin and Chen, 1991; Lin et al., 1992). Virions were disrupted by adding one-fourth volume of 4X disruption buffer [100 mM Na2HPO4, 50 mM NaH2PO4, 5 mM EDTA, 500 mg/ml bentonite (Sigma), 5% SDS, 1 μL RNasin® (40 U·μL-1, Promega, Madison, WI, USA) and 5% β-mercaptoethanol] and incubated at 60° C for 5 min. Proteins were removed by extracting twice with PCI (phenol:chloroform:isoamyl alcohol = 25:24:1, pH 8.0). The RNA was precipitated by NaOAc and ethanol and finally dissolved in RNase-free water.
The concentration of viral RNA was measured by the NanoDrop 1000 Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Virions and viral RNA were stored at -70° C for further use.
2.3. Virus inoculation and sequential sampling
To observe cell-to-cell and systemic movements of CymMV and ORSV in P.
26
amabilis inoculated leaves, preliminary tests were conducted from 18 Mar 2011 to 29 Mar 2011 and 5 July 2011 to 4 Aug 2011. The fully expanded second leaf from the top was used as the inoculated leaf. The inoculums were as follows:
1) 1.5 μg ORSV viral RNA dissolved in 20 μL inoculation buffer [5mM Tris-HCl (pH 8.0), 1mM EDTA (pH 8.0), 5mM phosphate buffer (pH 8.0), and 1 g·L-1 bentonite.]
2) 1.5 μg CymMV viral RNA dissolved in 20 μL inoculation buffer
3) 0.75 μg of each ORSV and CymMV viral RNA mixed in 20 μL inoculation buffer 4) Mock-inoculation with buffer only
Three plants were subjected to each treatment. Assuming the virus would spread along parallel veins, plants were mechanically inoculated with carborundum and cotton swabs at the tip side of about 30% of the area of the leaf. Four areas were marked parallel to veins for sequential sampling at 2, 4, 7, and 11 days post inoculation (dpi) or 4, 11, 16, and 22 dpi to determine the timing of virus movement (Fig. 1). And a longer test period with ORSV inoculation treatment only was conducted starting from 16 Aug 2011 and sampling at 5, 10, 20, 30 dpi. Virus infection and distribution were detected by tissue blotting.
2.4. Tissue blotting and hybridization assay for detecting virus 2.4.1. Tissue blotting
The study employed tissue blotting to reveal virus infection and distribution in inoculated leaves (Lin et al., 1990). Sampling areas were cut off with disposable razor blades and sliced perpendicular to the veins from base to tip sides sequentially with about 3 mm spacing. Each single plane cut surface was pressed on Hybond™-N+ (GE Healthcare Life Sciences, Little Chalfont, UK) nylon membranes for about two seconds.
27
Pressed leaf strips were collected in 15 mL Falcon tubes and frozen in liquid nitrogen, stored at -70o C for further experiments. Positive and negative controls and serial diluted (1 ng to 0.1 pg) viral RNA samples were dotted on the membranes to measure probe specificity and sensitivity. The blots were air-dried, UV corss-linked with 3-minute exposure to 1,200 μW·cm-2 UV light using the HL-2000 Hybrilinker UV crosslinker (UVP, Cambridge, UK) and kept in a dry box for following hybridization assays.
2.4.2. Preparation of digoxigenin (DIG)-labeled probes
The plasmid pGOCP2 and pGCCP (Fig. S1) were constructed for synthesis of ORSV CP and CymMV CP probes, respectively. ORSV and CymMV CP gene fragments were amplified from virus infected Phalaenopsis using RT-PCR with ORSV-3, ORSV-6 and CymMV CP-F1, CymMV CP-R1 primer pairs. Detailed information about the primers is listed in Table S1 and the RT-PCR reaction was performed as described in Section 1.2. Desired products were cleaned up using Gel/PCR DNA fragments Extraction Kit (Geneaid Biotech Ltd., New Taipei City, Taiwan), cloned to pGEM®-T Easy vector (Promega, Madison, WI, USA) and transformed into Fast-TransTM Escherichia coli DH5α competent cells (Protech Technology Enterprise Co., Ltd., Taipei, Taiwan) following the manufacturer’s protocol. After selection, the plasmids were purified from cultures of positive clones by NucleoBond® PC 500 Maxi-Prep kit (MACHEREY-NAGEL GmbH & Co. KG, Düren, Deutschland) and stored them at -20o C for further assays.
Non-radioactive DIG-labeling RNA probes were generated by in vitro transcription from pGOCP2 and pGCCP plasmids. The RNA probes were complementary to (+)-strand viral RNAs and recognized the CP region of CymMV and ORSV,
28
respectively. Plasmids were first linearized with restriction enzyme NdeI (New England Biolabs Inc., Beverly, MA, USA). The 1 μg linearized plasmid in 2 μL DIG-labeling Mix (Roche Applied Science, Penzberg, Germany) was mixed with 0.5 μL RNasin® (40 U·μL-1, Promega, Madison, WI, USA), 2 μL 10X polymerase reaction buffer and 2 μL T7 polymerase (50 U·μL-1, New England Biolabs Inc., Beverly, MA, USA) and brought the final volume to 20 μL with RNase-free water. After incubating 2 hrs at 37o C, 2 μL RQ1 RNase-Free DNase (1 U·μL-1, Promega, Madison, WI, USA) was added and incubated at 37o C for another 30 mins to digest the DNA template. Enzyme reaction was terminated by adding 2 μL 0.2 M EDTA. RNA transcripts were precipitated with 3 M NaOAc and 100% ethanol after refrigeration at -70o C for 1 hr. Centrifuged and dried RNA pellet was resuspended with RNase-free water to a final concentration about 400 ng·μL-1and stored at -70 oC for further use.
2.4.3. Hybridization and chemiluminescent detection
Tissue blots were hybridized with ~1 μg DIG-labeling probes for about 200 cm2 membranes. The following detection was performed using DIG detection kit (Roche Applied Science, Penzberg, Germany) according to the manufacturer’s protocol.
After adding probes and overnight-incubation at 65o C with a hybridization buffer [50%
formamide, 50 mM PO4, 0.8M NaCl, 1mM EDTA, 0.5% SDS, 10X Denhardt’s reagent, 250 μg·L-1 D-1626 Sodium salt Type 3 salmon sperm DNA (Sigma-Aldrich Corporation, St. Louis, MO, USA), and 500 μg·L-1 s R-6750 Type 6 baker’s yeast ribosomal RNA (Sigma-Aldrich Corporation, St. Louis, MO, USA)], unhybridized probes were washed off with high stringency buffer (0.1X SSC containing 0.1% SDS) and low stringency buffer (2X SSC containing 0.1% SDS) at 65o C and 25o C, respectively. Membranes
29
were incubated with blocking solution to reduce non-specific binding, then hybridized with 2 μL Anti-digoxigenin-AP Fab fragments (1.5 U·μL-1, Roche Applied Science, Penzberg, Germany). CDP-Star (Roche Applied Science, Penzberg, Germany) was used as chemiluminscent substrate and light emission was recorded on X-ray films.
3. Genome wide analysis of small RNAs from ORSV-infected P. amabilis by deep sequencing
3.1. ORSV inoculation assay and RNA sample preparation
To construct small RNA libraries from ORSV-infected P. amabilis, 1 μg of purified ORSV virion per leaf was inoculated on the first and second fully expanded leaf of each plant at the tip side as performed in preliminary tests. Plants mock-inoculated with 20 μL inoculation buffer were used as controls. The inoculated area (i) and non-inoculated area (c) of ORSV-inoculated (O) leaves were designated as Oi and Oc, respectively.
Corresponding non-infected tissues from the mock (M) plants were designated as Mi and Mc (Fig. 6). At 10 dpi, Oi, Oc, Mi, and Mc tissues were collected separately and frozen-stored at -70o C. After confirming virus infection by tissue blotting, tissues from three leaves were pooled for total RNA purification using the CTAB method. In addition, we performed RT-PCR to detect virus infection at a higher level of sensitivity. Protocol for tissue blotting, RNA extraction and RT-PCR reaction were the same as previously described.
3.2. Analyzing viral RNA and siRNA accumulation
3.2.1. Analyzing viral RNA accumulation by Northern blot
Northern blot assays were conducted to analyze viral genomic and subgenomic
30
RNA accumulation. Two μg of total RNA was denatured with glyoxal and separated on 1% agarose gel as previously described (Lin et al., 1996). RNA was transferred to Hybond™-N+ (GE Healthcare Life Sciences, Little Chalfont, UK) nylon membranes by the capillary method with 3 M NaCl and 0.01 N NaOH and immobilized with UV cross-link non-radioactive probes. Hybridization methods used for northern blot were the same as those used for tissue blotting as described in Section 2.4.
3.2.2. Analyzing viral siRNA accumulation by small RNA Northern blot
To detect viral siRNA accumulation, twenty-five μg total RNA was mixed with equal volume of 50% deionized formamide with bromophenol blue, boiled for 5 mins then placed on ice until loaded into the gel. MicroRNA Marker (New England Biolabs Inc., Beverly, MA, USA) containing a set of 17, 21 and 25-nt synthetic single-stranded RNA oligonucleotides was used as size markers. After separation in the 15%
acrylamide/7M urea gel, RNA were transferred to Hybond™-N+ (GE Healthcare Life Sciences, Little Chalfont, UK) nylon membranes by Trans-Blot® SD Semi-Dry Transfer Cell (BioRad Laboratories, Hercules, CA, USA) and immobilized by UV cross-link.
The following hybridization assay was performed with radioactive 32P-labeled probes. To detect a pool of siRNAs generated from ORSV RdRp or ORSV CP to 3’-UTR regions, viral gene fragments were cloned with ORSV RdRp-F/R and ORCP3UTR-F/R primer pairs (Table S1) from infected Phalaenopsis using RT-PCR, and ligated into pGEM®-T Easy (Promega, Madison, WI, USA) vectors according to the same method as described for the DIG-labeled probes. The consequent plasmids pGORd and pGOC3U (Fig. S2) were first linearized with restriction enzyme NcoI, and then used for SP6 polymerase-driven preparation of in vitro transcribed [α-32P]CTP
31
labeled probes as described previously (Lin et al., 1993; Lin et al., 1996). To increase the efficiency of small RNA detection, probes were chemically hydrolyzed to reduce their size with alkaline carbonate buffer (containing 8.5 mM Na2CO3 and 1 mM NaHCO3, pH10.2) at 60o C for 1 hr 30 min before hybridization. Alternatively, when detecting a specific viral siRNA, corresponding anti-sense synthetic single-stranded DNA oligonucleotide was used as probe and labeled with [γ-32P]ATP by T4 Polynucleotide Kinase (New England Biolabs Inc., Beverly, MA, USA) at the 5’-end. A 21-nt DNA oligonucleotide complementary to the microRNA Marker was also labeled by 5’-end labeling and used to hybridize with the size marker.
The membranes for small RNA northern blot were incubated with ULTRAhyb®-Oligo hybridization solution (Ambion, Carlsbad, California, USA) at 42o C for 1 hr for pre-hybridization. Radioactive probes were added into roller bottles and incubated overnight at 42o C. After two post-hybridization washes with wash solution (2X SSC and 0.1% SDS), radioactive signals were recorded by reusable phosphorimager screens and scanned by Typhoon® FLA7000 Biomolecular Imager (GE Healthcare Life Sciences, Little Chalfont, UK).
3.3. Small RNA deep-sequencing and annotation 3.3.1. cDNA library construction and sequencing
Small RNAs from inoculated (Oi and Mi) and non-inoculated (Oc and Mc) tissues of ORSV- or mock-inoculated plants were subjected to deep sequencing on 28 Nov 2011. The integrity of the RNA was checked by Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). For each library, 10 μg of total RNA was size fractionated on 15% tris-borate-EDTA urea polyacrylamide gel. Small RNAs were