國立臺東大學生命科學系碩士班 碩士論文
指導教授:彭仁君 博士 張文炳 博士
棘頰海葵魚初期個體發生學研究 Early ontogeny development of Spine-cheek anemonefish, Premnas
biaculeatus
研 究 生: 江玉瑛 撰
中華民國ㄧ百年十二月
國立臺東大學生命科學系碩士班 碩士論文
棘頰海葵魚初期個體發生學研究 Early ontogeny development of Spine-cheek anemonefish, Premnas
biaculeatus
研 究 生:江玉瑛 撰
指導教授:彭仁君 博士
張文炳 博士
中 華 民 國 ㄧ 百 年 十 二 月
誌 謝
本論文得以順利完成,感謝指導教授彭仁君老師與國立海洋生物博物館副研 究員張文炳老師給予論文指導與審查寶貴意見、東部海洋生物研究中心研究員兼 主任陳文義博士及副研究員何源興先生給予本論文審查諸多建議,於修業期間提 供良好的研究資源並給予鼓勵,在此致上由衷的謝意。感謝海生館張老師研究室 奕彰、依婷、合陞及國立屏東科技大學水產養殖系浩榮與阿甘,於實驗期間給予 諸多協助以利實驗順利進行。感謝水試所東部中心鄭明忠先生、李任棋先生、范 氏碧小姐、施勝中先生、董家宏先生、李建何先生、劉宗誠先生、李沛珊小姐、
陳富美小姐於平日工作上的協助;江偉全博士、許紅虹小姐、許儷玉小姐、蔡碧 玲小姐、周曉慧小姐、陳彥伶小姐、陳宗文先生、洪慈英小姐、邱燕華小姐、林 忠仙先生、蘇東山先生、陳正發先生、吳瑞賢博士、黃梓倫先生、蔡龍泉先生、
桂玉阿姨、田阿姨及秀玲給予平日的關心。於求學期間,感謝系辦惠嵐小姐在行 政上的協助,以及碩班同學怡辰、佳燕、軒佑、雅鈞、尚燁、耀文、弦育、俊成、
美秀及澤宇在課業上聯繫與提醒,在此一併致上最誠摯的謝意。最後,感謝我的 家人一直以來的支持與鼓勵,謝謝你們!!
棘頰海葵魚初期個體發生學研究
江玉瑛
國立台東大學 生命科學系碩士班
摘 要
棘頰海葵魚 (Spine-cheek anemonefish, Premnas biaculeatus Bloch, 1790) 成熟種魚配對後經 3 ~ 6 個月培育會陸續產卵,受精卵呈長橢 圓形橘紅色的附著性卵,平均長徑為 1.81 ± 0.09 mm;平均短徑為 0.79
± 0.03 mm;平均卵黃徑為 1.25 ± 0.09 mm,內有 0.16 ± 0.03 mm 大小 不一之油球數個。平均水溫 27.3 ± 1 ℃下約需 144.5 小時孵化,孵化 時間會在日沒後開始進行。初孵化的仔魚全長為 3.64 ± 0.31 mm,具趨 光性,開口口徑為 470 ± 60 μm 可攝食輪蟲 (Brachionus plicatilis),此 時頭部所有骨骼元素均以軟骨形式存在,上頜骨、齒骨與咽齒骨會隨
著口部發育於第 2 日開始骨化;第 3 日神經棘、血管棘及椎體橫突等
骨骼已達定數;第 5 日起口徑為 0.68 ± 0.26 mm 可開始兼投大型橈足
類,此時準備進入脊索末端上屈中期;第 7 ~ 8 日,體表橫帶開始形
成且下尾骨開始骨化並可發現牙齒形成;第 10 日脊索末端上屈後期
且鰭條開始發育,仔魚游泳能力明顯增強;第 15 日前後體表色素開
始擴散全身,下尾骨及主要鰭條完全骨化;孵化後第 20 日,仔魚全
長已達 8.60 ± 1.04 mm,可開始接受人工粒狀飼料;第 31 日後全長為
9.88 ± 1.48 mm 開始進入稚魚階段。由骨骼與形質結果可知,仔魚發育
初期主要將能量運用攝食相關形質發育上,顯示仔魚之攝食機能發育
必需優先於游泳機能,如此可確保仔稚魚在初期階段不會因飢餓及營
養不良而遭到淘汰。
關鍵詞:棘頰海葵魚、魚苗、形態、骨骼發育
Early ontogeny development of Spine-cheek anemonefish, Premnas biaculeatus
Yu-Ying Jiang
Abstract
After pairing 3 to 6 months, the brooders of spine-cheek anemonefish (Spine-cheek anemonefish, Premnas biaculeatus Bloch, 1790) spawned.
The adhesive demersal eggs were ellipsoidal and orange-red. The diameter of eggs were 1.81 ± 0.09 mm in length and 0.79 ± 0.03 mm in width; the yolks were about 1.25 ± 0.09 mm in diameter and contained several oil droplets (0.16 ± 0.03 mm in diameter). Hatching occurred after sunset and accumulated 144.5 hours (water temperature 27.3 ± 1 ℃). Newly hatched larvae were 3.64 ± 0.31 mm in total length (TL), with phototaxis, the gape height were 470 ± 60 μm can feed rotifers (Brachionus plicatilis), all head bones were cartilage. The maxillary, dentary and pharyngeal dentary of mouth began to ossify on 2nd days post hatch (DPH); Ossification of the neural spines, haemal spines and parapophyses were completed on 3 DPH;
The notochord flexion occurring, gape height were 0.68 ± 0.26 mm and it
can begin to fed larger copepods on 5 DPH; Horizontal band occurred on
body surface, hypurals occurred and ossified, dentary teeth occurred on 7 ~
8 DPH; The development of notochord postflexion was completed, caudal
rays began to develop and it increased larval swimming ability significantly
on 10 DPH; Pigment spread to around the body surface, the hypurals and
principal caudal rays had completely ossified on 15 DPH. The larval can
begin to accept artificial pellet feed (TL, 8.60 ± 1.04 mm) on 20 DPH; The
larvae can development into juvenile stage (TL, 9.88 ± 1.48 mm) on 31 DPH. The results showed that the early stages of larvae may be used the initially energy priority to develop the morphometric characters of feeding than swimming function, to ensure the early stages fish larvae not due to hunger and malnutrition was eliminated.
Key words: Premnas biaculeatus、larva、morphology、osteological development
目錄
中文摘要…...………..……….……..…..
i
英文摘要…...……….………….………...
iii
目錄...……….………...
v
表目錄...………..…………...
vii
圖目錄...
viii
附圖目錄...
x
壹、前言... 1
ㄧ、緒言... 1
二、棘頰海葵魚... 2
三、形態發育... 3
四、骨骼發育... 4
五、研究動機與目的... 5
貳、材料與方法... 6
ㄧ、種魚飼育... 6
二、胚胎發育... 6
三、仔魚培育... 7
四、形態發育... 7
五、透明骨骼製作及標本觀察... 7
六、餌料生物培養... 8
(一) 擬球藻培養... 8
(二) 輪蟲之篩選及蓄養... 9
(三) 橈腳類之篩選及蓄養... 9
(四) 豐年蝦之孵化... 9
參、結果…... 11
ㄧ、種魚培育... 11
二、胚胎發育... 12
三、仔魚培育………... 13
四、形態發育………... 14
五、骨骼發育... 15
肆、討論…... 19
一、種魚培育... 19
二、胚胎發育... 19
三、仔魚培育………... 21
四、形態發育... 22
五、骨骼發育... 23
伍、結論... 28
陸、參考文獻... 29
表目錄
Table 1. Summary data for sampling Premnas biaculeatus………... 39 Table 2. Embryonic development of Premnas biaculeatus……….. 40 Table 3. Morphometric of Premnas biaculeatus larvae (mean±sd ). ... 41 Table 4. A comparison of egg diameter, incubation water temperature, hatching
time and fry length among different species of anemonefish. ... 42
圖目錄
Fig. 1 Relationship between body length (cm) and body weight (g) indices female of Premnas biaculeatus. ………... 43 Fig. 2 Relationship between body length (cm) and body weight (g) indices
male of Premnas biaculeatus. ………... 43 Fig. 3 Relationship between body length (cm) and body weight (g) indices
immature of Premnas biaculeatus. ………... 43 Fig. 4 Spawning behavior of Premnas biaculeatus. ………... 44 Fig. 5 Embryo development of Premnas biaculeatus. ………... 45 Fig. 6 Egg color changes during embryonic development of Premnas
biaculeatus. …………...………...…………... 46 Fig. 7 The morphological changes of the Premnas biaculeatus at larval and fry
stage. ………... 47 Fig. 8 Feed supply and water management scheme in larval rearing of
Premnas biaculeatus. ………...………. 48 Fig. 9 Spine-cheek development of Premnas biaculeatus. ………. 49 Fig. 10 Growth of Premnas biaculeatus larvae from hatching until day 40
(mean±sd). ………... 50 Fig. 11 Some body proportions of Premnas biaculeatus development, shown as
percentages of Total length. ……….. 51 Fig. 12 The growth of gape height in Premnas biaculeatus.………... 52 Fig. 13 Osteological developmental sequence of head, vertebral column, caudal,
dorsal and anal fins in Premnas biaculeatus. ………... 53 Fig. 14 Representative cleared and stained specimens of P. biaculeatus at (A) 1
DPH and (B) 8 DPH. ………... 54 Fig. 15 Representative developmental sequence of the feeding apparatus in
Premnas biaculeatus through ontogeny. ………... 55 Fig. 16 Development of the vertebral column, caudal, dorsal and anal fins in
Premnas biaculeatus. ………... 56
Fig. 17 Development of the caudal fins in Premnas biaculeatus. ……… 58 Fig. 18 Comparison of ostelogical development between A. ocellaris, A.
frenatus and P. biaculeatus in day. ………... 60
附圖目錄
附圖1. 棘頰海葵魚 (Premnas biaculeatus)。………... 61 附圖2. 棘頰海葵魚屬與海葵魚屬的分類特徵。………... 61 附圖3. 棘頰海葵魚的分佈。………... 61
壹、前言
一、緒言
根據世界觀賞魚市場分析中淡水魚類的觀賞魚佔全貿易市場的80 % ~ 90 %。
大部份淡水觀賞魚中約有 90 % 經由人工繁養殖獲得,而海水觀賞魚則有 90 % 需來自野外的捕捉 (Tlusty, 2002)。海水觀賞魚分布的棲息環境,多為熱帶珊瑚礁 海域,由於體色彩繽紛,種類繁多,不僅成為大型水族館中的嬌客,亦是眾多愛 好海水生態水族箱之消費者趨之若騖的選擇對象。近年來因家庭式生態水族箱的 盛行,使得活珊瑚及海水觀賞魚需求量持續增加,加上航空貨運的便捷,讓不少 歐美水族愛好者可選擇品質佳且多樣、多種類的活珊瑚及海水觀賞魚;另外,海 水水族箱設備的不斷提升,更讓水族愛好者有能力自行設置家庭式水族箱,使淡 水水族箱玩家已逐漸趨向多樣化的海水水族箱。目前超過 1000 種珊瑚礁魚類在 海水魚貿易市場中流通,但卻只有 84 種來自於人工繁殖 (Arvedlund et al., 2000)。許多海水觀賞魚供應國,為方便採捕會使用具有毒性的化學藥劑如氰化物 等毒殺珊瑚礁魚類,使受毒害魚隻外表看不出損傷,但經飼養一段時日後便開始 漸漸死亡,而擴散的氰化物不僅污染珊瑚礁也破壞了海洋生態系 (Olivotto et al., 2006; Rubec et al., 2001 )。
由於海水觀賞魚的人工繁殖技術至今仍非常困難,主要在於魚苗孵化後幼魚口 徑小,適合的生物餌料無法持續供應及人工微粒飼料尚未開發完成,導致必須持 續依賴野外族群 (蔡,2009)。水產養殖為台灣重要傳統產業之一,縱觀台灣地理、
天候環境很適宜進行水產養殖工作,水產種苗產業及相關周邊產業已相當發達,
極有機會開創另一海水觀賞魚類繁養殖之新型態產業。目前台灣已能成功繁殖多 種海洋觀賞性魚類如雀鯛 (damselfish)、海葵魚 (anemone fish, genus Amphiprion and genus Premnas)、海馬 (Hippocampus spp.) 與蓋刺魚類等,其中以具明顯市場 偏好、健康穩定且完全不倚賴棲地環境資源的人工培育海葵魚最受消費者所青 睞,透過人工繁養殖培育過程不但明顯提升個體於運輸後及飼養過程之活存率,
同時也在符合動物福利與生態環保議題下,成為外銷北美及歐盟市場主要之繁殖
培育海水觀賞魚種 (黃,2010)。
二、棘頰海葵魚
海葵魚 (Clown anemonefish) 俗名小丑魚,屬雀鯛科 (Pomacentridae) 海葵魚 亞科 (Amphiprioninae) 之魚類,本亞科可分成海葵魚屬 (Amphiprion) 及棘頰海葵 魚屬 (Premna) 二種,在野外環境中色彩及圖案為辨識小丑魚的主要特徵,其它有 用的特徵包括齒形、頭部形態及身體比例作為分類上的標準 (Fautin and Allen, 1992),而棘頰海葵魚於鰓蓋骨下方有兩隻骨質蓋刺,故將棘頰海葵魚獨立為棘頰 海葵魚屬,因此海葵魚亞科分成 2 屬 28 種 (Fautin and Allen, 1997)。台灣目前之 記錄僅有 Amphiprion 一屬共五種;包括眼斑海葵魚 (Amphiprion ocellaris, 公子 小丑)、鞍斑海葵魚 (A. polymnus, 鞍背小丑)、粉紅海葵魚 (A. perideraion, 咖啡小 丑)、白條海葵魚 (A. frenatus, 紅小丑) 及克氏海葵魚 (A. clarkii, 雙帶小丑) 等,
主要分佈於岩礁海域及離島等處 (沈,1993)。
棘頰海葵魚 (Spine-cheek anemonefish,Premnas biaculeatus Bloch, 1790) 俗名 透 紅 小 丑 ( 附 圖 1) 。 分 類 上 屬 於 輻 鰭 魚 綱 (Actinopterygii) 、 新 鰭 魚 亞 綱 (Neopterygii) 、 鱸 形 目 (Perciformes) 、 隆 頭 魚 亞 目 (Labroidei) 、 雀 鯛 科 (Pomacentridae)、海葵魚亞科 (Amphiprioninae)、棘頰海葵魚屬 (Premnas) 之魚 種。在外部型態上,其背鰭硬棘 X,軟條 17 ~ 18;臀鰭硬棘 II,軟條 13 ~ 15;
體呈橢圓,側扁。眼睛下有一對長刺 (附圖 2);三道白細條紋分別於眼後方、軀 幹中段及尾柄處垂直穿過魚體;體色為濃豔的深紫褐色至深紅色,是一種受歡迎 的海水觀賞魚種 (邵,2011)。另,蘇門達臘有一種分類地位與棘頰海葵魚相同的 海葵魚,其原本的三道白細紋為三道黃色寬斑紋所取代 (Froese and Pauly, 2011),
因此水族業界又將此種稱為金透紅小丑。
棘頰海葵魚與其他海葵魚一樣具有性轉變的能力,雌魚體型普遍大於雄魚,
性成熟的雌魚最大體型可達 16 cm (Fautin and Allen, 1992),在熱帶海域中全年皆 有產卵行為,而在亞熱帶及溫帶海域產卵行為僅發生在水溫較高的春天及夏天 (Fautin and Allen, 1997;Richardson et al., 1997)。喜歡與拳頭海葵 (Entacmaea quadricolor) 共生,棲息水深一般不超過 50 公尺,溫度範圍 25 ~ 28 ℃ 間的熱帶
溫暖水域 (Fautin and Allen, 1992)。主要分佈在印度-澳洲島群,包括印度、緬甸、
泰國、馬來西亞、印尼、菲律賓群島、新幾內亞、新不列顛島、所羅門群島、萬 那杜及澳洲等地 (邵,2011) (附圖 3) 。台灣並不在主要分佈範圍內,但在 98 年 12 月農委會野生動物活體輸出入審核要點公告中,棘頰海葵魚與雙帶海葵魚 (Amphiprion clarkii)、白條海葵魚、眼斑海葵魚、粉紅海葵魚、鞍斑海葵魚及黑雙 帶海葵魚 (A. sebae) 皆列為法定同意輸入觀賞水產動物名錄之魚種 (農委會,
2009)。
三、形態發育
魚類發育的初期是一段快速且複雜的發生和分化過程,由於器官與系統的快 速變化會影響發育期間的生理和行為能力。發育過程中除了體型增加,身體部位 的比例為適應環境在形態上會有很大的改變,進而提高攝食、呼吸、運動等重要 器官系統的運作,以增加在早期發育階段存活和發育 (Osse and Boogart, 1995)。透 過形態學研究,比較不同來源或不同批次的受精卵,觀察發育和體結構來判斷魚 隻健康程度與缺陷,進而評估仔魚品質與建立適合的環境條件和飼育方法,降低 由於基因不良或形態畸形造成飼養成本上的損失,以提高養殖收益 (Gisbert et al., 2002)。
珊瑚礁魚類仔稚魚培育中,初期發育階段死亡率最高的二個階段為受精卵孵 化及隨之而來的初期攝餌階段,也就是當魚苗營養來源由內源性的卵黃囊轉變為 外源性攝食時期,因仔魚伴隨著卵黃耗盡後,仔稚魚的組織及器官初步形成,構 造發育不足以應付環境的挑戰。故魚苗必須盡可能在卵黃及油球耗盡或可能面臨 飢餓前達成有能力捕捉和消化食物 (Avila and Juario, 1987; Fukuhara, 1988)。對於 初次攝食的仔魚,可攝食餌料生物的體型受限於口徑大小、器官及攝食構造的發 育狀況,而影響魚苗攝食成功的因素,除了魚苗本身的視覺、感覺器官、游泳能 力、攝食骨骼發育、消化系統及搜索行為等的發育程度之外,餌料生物本身的大 小、顏色、密度及游泳方式也會影響海水魚苗的攝食能力及選擇性 (Green and McCormick, 2001; Job and Bellwood, 1996; Turingan et al., 2005; Wittenrich et al., 2007)。因此魚類自孵化後,經由一連串個體發生學上的形態變化,其外部身體形
態、內部生理代謝與行為習性上因而有所改變,以適應周圍生活環境 (Kendall et al., 1984; Youson, 1988)。
另外硬骨魚類所產的卵,不論是漂浮性、沈著性或粘著性,自仔魚孵化後大 都有一段浮游幼生時期,即所謂的仔稚魚時期。在此期間仔魚不具游泳能力而隨 波逐流,其不具完整的主動游泳能力,即不能游泳達到水平方向位移的目的,而 行暫時性的浮游生活,當其各種游泳的鰭如胸鰭、腹鰭、尾鰭等發育完成後,過 著完全不同的主動游泳生活。將仔稚魚與成魚區分出來在生態及形態上都具有特 別的意義,因仔稚魚所擁有的特殊形態使其在海洋或淡水水體中,佔據與成魚截 然不同的生態地位 (丘,1999)。
四、骨骼發育
初期仔魚在成長過程中由於各部位的器官尚未發育完成,對於主動攝食及避 敵的能力有限,而對於可攝食餌料生物的體型受限於其口徑大小與攝食構造的發 育程度。在魚類的攝食機制中,涉及一套複雜的頭部骨骼、肌肉、韌帶及肌腱的 連動,初次攝食的仔魚具有一個簡單的攝食機制,主要由舌骨與下頜骨的聯繫,
到接近變態時期由舌骨、下頜骨及鰓蓋骨的聯繫以完成更複雜的捕食功能來有效 捕捉到獵物 (Hunt von Herbing, 2001; Turingan et al., 2005)。在ㄧ些商業價值海水魚 類仔魚的研究中,探討攝食偏好和形態功能之間的關係,證明攝食骨骼構造的發 育程度,限制仔魚期攝食成功的能力 (Wittenrich et al., 2007; 2009)。Wittenrich et al.
(2007) 記錄夜棲鋸鱗蝦虎魚 (Priolepis nocturna) 仔稚魚初期發育及攝食能力,因 受 限 於 發 育 未 完 全 的 攝 食 構 造 影 響 , 仔 魚 只 能 攝 食 游 動 力 弱 的 甲 藻 (Dinoflagellates) 和 Tintinnids,因此在開始初次攝食的階段,攝食構造的發育程度 可能是成功捕獲獵物的一個重要因素 (Wittenrich et al., 2007),對於仔稚魚在成長 過程中攝食及游泳機能發育的掌握仍有其重要性。
對於魚類發育的研究,隨著飼育技術的進步,使仔稚魚各發育階段的樣本容 易取得,也克服野生樣本收集困難的問題,骨骼發育除了提供個體發育的基本知 識之外,亦可用於系統分類 (Liu, 2001; Hilton and Johnson, 2007) 與骨骼異常發生
(Gavaia et al., 2002; Fraser et al., 2004) 的研究,為了解魚苗發育過程中骨骼異常的 發生,建立正常骨骼發育的基礎資料仍有其重要性 (Koumoundouros et al., 1997;
Faustino and Power, 1999)。在漁業和水產養殖的學科領域,骨骼個體發育有助於理 解一個物種在不同發育階段的形態功能和環境偏好 (Fukuhara, 1992),對於培育過 程中的改善飼育條件有正面價值 (Koumoundouros et al., 1999, 2001a; Sfakianakis et al., 2005)。
五、研究動機與目的
魚類的生活史過程由各個生長階段所組成,每個生長階段通常都有各自的生 長特徵,而魚類的初期發育階段,形態發生及分化是快速且複雜的過程,了解養 殖物種初期生活史的生長形式,掌握不同階段的生長特徵和轉換時間,降低飼育 環境的限制因素,才能以最適合的飼育方式來進行繁養殖工作以提高魚苗的育成 率。目前關於棘頰海葵魚的研究除了生殖及初期發育 (Kim et al., 2007) 之外,尚 有對仔魚的視覺發育 (Job and Bellwood, 1996) 與餌料生物滋養對仔稚魚成長表現 (Kim et al., 2009) 等相關研究。而本研究以棘頰海葵魚在人工條件下進行親魚培育 及繁養殖飼育,觀察胚胎與仔魚外部形態變化、攝食及游泳等相關骨骼發育過程 的生長特徵,透過了解棘頰海葵魚初期發育階段的轉變,可作為未來仔稚魚培育 管理過程中的參考。
貳、材料與方法
ㄧ、種魚培育
本研究之棘頰海葵魚購入自坊間水族館12 尾及自行繁殖 260 尾,合計為 272 尾。試驗種魚以 1.8 噸 FRP 水槽 (尺寸為 200 × 80 × 60 cm3 );置入 4 ~ 5 個巨 大異幅海葵,供飼驗魚配對及共生,水槽中加設控溫設備,天然採光照明,水溫 維持在 27 ± 1.5 ℃,鹽度為 32 ~ 33 psu,每日餵食二次並交替新鮮蝦肉、魚肉及 乾性粒狀配合飼料餵飼。培育過程中若發現死亡情形,立即進行解剖及鏡檢,以 了解魚致死原因,並記錄種魚性別、生殖腺發育情形及測量體重、全長等資料。
了解雄魚、雌魚及未成熟魚體重與全長之關係,套用指數關係 (power relationship) 以 Excel 統計軟體估算指數迴歸關係式,進而了解棘頰海葵魚雌雄全長與體重之 關係。
試驗種魚經一段時間蓄養後,若有同一朵海葵中有兩尾魚同時躲藏,並追趕 其他魚隻,則可視為初步配對成功,以手抄網將初步配對之 2 尾種魚移入種魚培 育水槽中進一步蓄養。種魚培育缸大小為 50 cm × 45 cm × 40 cm,養殖用水採循 環過濾方式處理,設備包括不織布粗過濾、蛋白除沫機、生物濾床及紫外線殺菌 器等,每日循環次數為10 ~ 12 次,內置產卵床如空心磚、鵝卵石、陶製小花盆或 珊瑚石一個,還有巨大異輻海葵一顆以提供躲藏,平日以自製軟性飼料及市售粒 狀飼料交替餵食,每天於上午及下午各投餵一次至每尾種魚飽食為止。若發現配 對種魚間互有攻擊行為,則馬上將種魚移回蓄養缸中重新配對。另觀察種魚產卵 之行為模式,包括雌雄種魚清理產卵床、產卵行為、排精行為、受精及護卵行為 等,並以數位照相器材拍攝記錄。
二、胚胎發育
吸取受精卵在 40 倍光學顯微鏡下,測量 30 粒受精卵長徑、短徑、油球徑、
卵黃徑,同時每日取受精卵數粒,利用吸管將受精卵吸至凹槽載玻片上,水量剛 好蓋過受精卵,並以數位照相器材拍攝胚胎發育過程,同時記錄時間、水溫與胚 胎發育之關係,直至受精卵孵化為止。
三、仔魚培育
初孵化仔魚具趨光之習性,使用聚光燈及虹吸管收集仔魚進行培育,育苗水 溫為 24 ~ 28 ℃,鹽度為 33 ~ 35 psu,培育槽為 1.8 噸之 FRP 桶。仔魚孵化後 即提供輪蟲 (Brachionus plicatilis) 作為開口餌料生物,輪蟲投餵期間 (1 ~ 6 天) 仔 魚培育採止水式,以滴流方式添加擬球藻 (Nannochloropsis oculata) 穩定水質,水 中透明度維持在50 ~ 80 cm,第 5 日以後開始兼投橈足類 (Copepod) 並與輪蟲混 合投放,第 8 日開始投餵滋養豐年蝦 (Artemia spp.) 無節幼蟲作為仔魚餌料,每 日更換海水 200 ~ 250 L,第 20 日開始以微粒浮性飼料馴餌,每日下午抽底一次,
並開始流水養殖。
四、形態發育
試驗期間每日採樣 6 尾仔魚,以立體顯微鏡拍攝仔稚魚之鰭部、體態與體 色等成長過程之外形變化,並利用生物解剖顯微鏡搭配數位影像測量分析軟體 (NIS -Elements D 2.30, Nikon),記錄全長 (totel length, TL)、標準體長 (standard length, SL)、頭長 (head length, HL)、眼徑 (eye diameter, ED)、吻長 (snout length, SnL)、下顎長 (lower jaw length, LJL)、臀鰭前長 (preanal length, PAL)、體高 (body depth at pectaral-fin, BDP) 及口徑 (gape height, GH) 等各部位形質,形態測量的定 義及縮寫參考 Leis and Carson-Ewart (2000);口徑計算公式為:GH=√(UJL2+LJL2),
UJL (上顎長),LJL (下顎長) (Wittenrich et al., 2007);仔稚魚發育階段參考 Kendall et al. (1984),發育階段區分為 (1) 脊索末端上屈前期 (Preflexion)、(2) 脊索末端 上 屈 中 期 (Flexion) 、 (3) 脊 索 末 端 上 屈 後 期 (Postflexion) 及 (4) 稚 魚 期 (Juvenile)。測量結束後將標本固定於 10 % 中性福馬林,以利後續研究進行。相 對成長是各形質長度與全長之比率關係,相對成長 = 形質長度 ÷ 全長 × 100 %。
五、透明骨骼製作及標本觀察
於孵化後前 20 天每日取樣 6 隻,於第 20 至 32 天每隔三日取樣一次,以 10 % 福馬林固定後,參考 Song and Parenti (1995) 與 Potthoff (1984) 提出的染色 方法製作透明骨骼,經透明與染色的步驟後可觀察到硬骨呈紅色、軟骨呈藍色的
骨骼標本。製作流程修改如下:
(一) 固定:樣本以 10 % 福馬林固定約 24 ~ 48 小時。
(二) 水洗:以蒸餾水浸泡樣本約 1 ~ 2 天。
(三) 漂白:為去除魚體表色素,可浸泡於 3 % H2O2 漂白溶液中約 1 天。
(四) 依序浸泡於 30 % → 50 % → 70 % → 95 % 酒精溶液進行脫水,間隔 30 分鐘。
(五) 軟骨染色:配製軟骨染液 (80ml 的 95 %酒精 + 20 ml 的冰醋酸 + 10 mg Alcian blue 染劑),將樣本浸泡於染液中約 30 ~ 60 分鐘,染色時間視染 色程度而異。
(六) 依序浸泡於 95 % → 70 % → 50 % → 30 % 酒精溶液 → 清水,間隔約 30 分鐘。
(七) 透明:配製酵素溶液 (30 ml 飽和硼酸鈉溶液 + 70 ml 蒸餾水 + 1 g Trypsin ),浸泡樣本至開始呈現透明度。
(八) 置於 0.5 % KOH 溶液中約 30 分鐘。
(九) 硬骨染色:於 0.5 % KOH 溶液中加入少許 Alizarin red S 染劑使溶液呈 紫紅色即可,將樣本浸泡在染液中約 30 ~ 60 分鐘,時間視染色程度而 異。
(十) 脫色:置於 0.5 % KOH 溶液中,約 1 天內更換二次 KOH。
(十一) 保存:將標本浸泡於 0.5 % KOH 與甘油的混合溶液中,視魚體變軟或 呈透明的狀況,按不同比例進行更換 0.5 % KOH:甘油 = 7:3、1:1、
3:7,至魚體透明後置入純甘油中保存。
標本製作完成後,以解剖顯微鏡觀察骨骼發育並拍照紀錄,並以標準體長為 紀錄,各部位骨骼名稱參考 Sfakianakis et al. (2005) 及 Koumoundouros et al. (1999) 所描述。
六、餌料生物培養
(一) 擬球藻培養 (蘇,1999)
擬球藻 (Nannochloris oculata) 取自東港生技研究中心餌料生物實驗 室,培育於 0.5 噸 FRP 桶,引入經逆滲透過濾後鹽度為 32 psu 的海 水,曝氣 3 日後,以加入硫酸氨 (Ammonium sulphate) 30 g、尿素 (Urea) 15 g 及過磷酸鈣 (Single superphosphate) 15 g。藻水與新注入海水比例為 1:5 ~ 1:10,以氣泡石打氣至水色不再變濃時 (約 5 ~ 7 日) 即收穫使用,
可重複進行至藻細胞增殖不良時全數收穫。
(二) 輪蟲之篩選及蓄養
1. 本實驗使用東部海洋生物研究中心自行培育之輪蟲,以 300 網目之浮 游生物網至輪蟲池撈捕收穫後,以 150 網目浮游生物網過濾出殼長約 120 ~ 150 μm 之輪蟲。
2. 重覆過濾數次後直到鏡檢為輪蟲後再培養於 200 L 錐形桶中,培養桶 中加入擬球藻作為輪蟲之餌料。培養過程中待藻水清澈後再予以添加 綠藻水。
(三) 橈足類之篩選及蓄養
1. 本實驗使用東部海洋生物研究中心自行培育之橈腳類,主要為模糊許 水蚤 (Pseudodiaptomus annandalei) 及短角異劍水蚤 (Apocyclops royi) 二種,成蟲體長為 250 ~ 1050 μm 之間,以 200 網目之浮游生物網至 橈腳類培育池撈捕收穫後,以 150 網目浮游生物網過濾出殼長約 420
± 90 μm 及殼寬約 186.3 ± 12.5 μm 之橈腳類培育仔魚。
2. 重覆過濾數次後直到鏡檢為橈腳類後再培養於 200 L 錐形桶中,並中 加入擬球藻作為橈腳類之餌料。培養過程中待藻水轉清後再予以添加 藻水。
(四) 豐年蝦之孵化
購入市售豐年蝦卵 (Belgium, INVE High-5) 自行孵化,孵化條件為 : 水溫 27 ~ 30 ℃ ,鹽度 25 ~ 30 psu,pH ≧ 8,蝦卵密度 2 ~ 3 g / l,初孵
化豐年蝦無節幼蟲 (Artemia sp.) 體長約 400 ~ 500 μm,以直接投餵方式 餵食仔魚。
參、結果
ㄧ、種魚培育
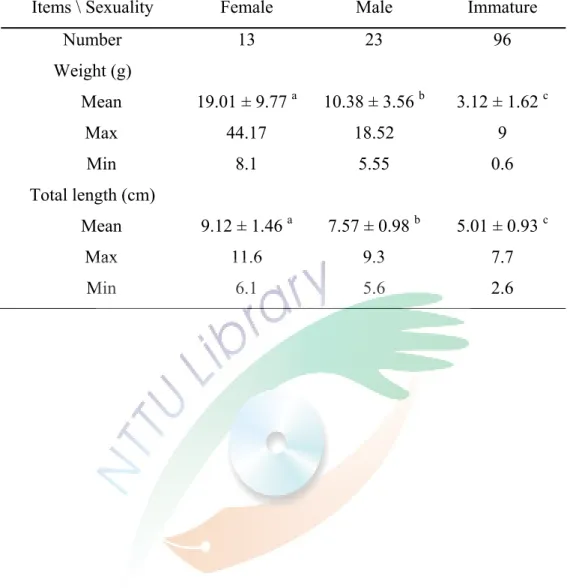
試驗期間獲得的樣本數為 132 尾,解剖樣本中得到 13 尾雌魚,23 尾雄魚,
未成熟有 96 尾。測量數據得知雌魚平均全長為 9.12 ± 1.46 cm (6.10 ~ 11.6 cm),
而平均體重為 19.01 ± 9.77 g (8.10 ~ 44.17 g);雄魚平均全長為 7.57 ± 0.98 cm (5.6
~ 9.3 cm),而平均體重為 10.38 ± 3.56 g (5.55 ~ 18.52 g);未成熟魚之平均全長為 5.01 ± 0.93 cm (2.6 ~ 7.7 cm),而平均體重為 3.12 ± 1.62 g (0.6 ~ 9.0 g),經統計分 析樣本得知雌雄之體重、體長上具顯著差異 (p<0.05) (Table 1)。棘頰海葵魚雌魚、
雄魚及未成熟魚之體長與體重關係式如下:
Fig. 1 為雌性棘頰海葵魚其體長與體重之關係式:
W = 1.36889L0.9738,R2 = 0.8944,n=13 Fig. 2 為雄性棘頰海葵魚其體長與體重之關係式為:
W = 1.36506L0.9317,R2 = 0.8239,n = 23。
Fig. 3 為未成熟棘頰海葵魚其體長與體重之關係式為:
W = 1.72271L0.1769,R2 = 0.8022,n = 96
種魚配對後經過 3 ~ 6 個月培育,陸續開始出現產卵行為,棘頰海葵魚在產 卵床挑選上並沒有明顯的差別,本研究於產卵缸中置入空心磚、花盆及珊瑚礁石,
結果發現種魚並無明顯挑選材質上的偏好,皆會在上面產卵,甚至於有時海葵位 移到玻璃上,種魚也會將卵產在玻璃缸面上,讓卵得到海葵的保護。
棘頰海葵魚之生殖行為如 Fig. 4 所示,於產卵前 2 ~ 3 小時,觀察到雌、雄 魚生殖孔明顯突起,且種魚會選擇海葵旁邊之穩固物作為產卵基床,產卵前雌雄 種魚會積極以其口啄除產卵床上之藻類及附著物,此時可觀察到雌魚腹部膨脹且 生殖突起呈圓鈍狀,而雄魚生殖突起則為細尖狀則可判定種魚即將產卵。開始產 卵後 30 ~ 50 分鐘內,種魚會連續不間斷的產卵及排精,雌魚腹部會緊貼著產卵
床將卵產於卵床上,產出數 10 顆卵粒後雄魚隨後進行受精,有時雌雄亦會同時 產卵排精。觀察發現每回產卵時間在 09:00 ~ 16:00,大部份集中在 9:30 ~ 14:
00 左右,依雌魚體型大小區別計算其卵粒數,得知全長 6 ~ 10 cm 的雌魚產卵量 約在 800 ~ 2500 粒左右;全長 10 ~ 12 cm 的雌魚產卵量約 3000 粒以上,推算 卵粒分佈面積約 5.5 ~ 11.5 cm2。
產卵結束後種魚會有護卵行為,一般以胸鰭搧動水流為主,其主要目的為增 加溶氧及加速胚體代謝物之擴散,同時種魚也會以口啄除死卵,雖然雌、雄種魚 都會護卵,但主要的護卵工作是由雄魚擔任,越接近孵化日以胸鰭扇動水流之頻 率增加,即使是在零光照的環境下,種魚仍持續以胸鰭搧動水流直至仔魚全部孵 化為止。
二、胚胎發育
棘頰海葵魚受精卵之平均長徑為 1.81 ± 0.09 mm;平均短徑為 0.79 ± 0.03 mm;平均卵黃徑為 1.25 ± 0.09 mm,內有 0.16 ± 0.03 mm 大小不一之油球數個,
受精卵為橢圓且呈橘紅色,偏動物極之頂端具有棉絮狀之附著絲,其功用在使卵 粒黏附於卵床上。
受精卵之胚胎發育過程如 Table 2 及 Fig. 5 所示,在水溫 27.3 ± 1 ℃及鹽度 介於 32 ~ 33 psu 下,受精卵之外觀如 Fig. 5 A 所示,受精 35 分後胚胎發育為 2 細胞期 (Fig. 5 B); 1 小時 05 分為 4 細胞期 (Fig. 5 C); 1 小時 50 分後為 8 細胞期 (Fig. 5 D); 2 小時 25 分後為 16 細胞期 (Fig. 5 E); 3 小時 05 分後為 32 細胞期 (Fig. 5 F); 3 小時 40 分後為 64 細胞期 (Fig. 5 G); 5 小時後為桑 椹期 (Morula atage) (Fig. 5 H); 11 小時 20 分後為囊胚期 (Blastula atage) (Fig. 5 I); 20 小時 00 分後胚囊覆蓋卵黃且可見胚體及眼胞形成 (Fig. 5 J); 23 小時後 出現 8 體節(Fig. 5 K); 26 小時 40 分後耳胞形成 (Fig. 5 L); 33 小時 10 分後眼 胞內晶體形成且尾部與卵黃分離 (Fig. 5 M); 37 小時 50 分後心臟開始博動且心跳 速率:96 ~ 114 time / min (Fig. 5 N); 41 小時 20 分後胚體頭部旋轉至卵膜前方,
後卵黃及胚體上已出現色素細胞 (Fig. 5 O); 50 小時後胚體眼上已見色素沈澱 (Fig. 5 P); 69 小時後胚體眼上已積聚鳥糞素;100 小時後胚體眼上已積聚鳥糞素
(Fig. 5 Q); 140 小時尾部末端已延長至眼部,胚體發育完全 (Fig. 5 R); 144 小 時 30 分破卵而出的仔魚及卵膜 (Fig. 5 S),初孵化仔魚測量腹部卵黃長徑約為 1.43 mm,油球徑約為 0.23 mm。
Fig. 6 為棘頰海葵魚胚胎發育過程受精卵顏色變化,第 1 日受精卵顏色呈橘 紅色,與卵黃之顏色有關;第 2 日受精卵顏色為橘色,鏡檢受精卵可發現胚體覆 蓋全卵黃且脊索形成;第 3 日受精卵顏色變為黃褐色,此時胚體已經形成,卵黃 及胚體上有色素細胞,導致受精卵顏色變暗;第 4 日受精卵顏色變為黑褐色,可 發現胚體色素沉著眼部黑化;第 5 日受精卵顏色變為亮黑褐色,可觀察到胚體眼 部閃亮的銀白色光澤;第 6 日受精卵顏色變為亮銀色,此時卵黃囊縮小且眼部積 聚鳥糞素 (Guanine) 呈現銀白色金屬光澤,心臟已可見色素沉著,尾部末端已延 長至眼部,胚體已經發育完全,此時為孵化離床的徵兆,仔魚會在日落後 30 分 鐘左右開始孵化並破殼而出。
三、仔魚培育
棘頰海葵魚仔魚形態變化過程及餌料生物種類如 Fig. 7 及 Fig. 8 所示,在水 溫 27.3 ± 1 ℃下,剛孵化仔魚之平均體長為 3.64 ± 0.3 mm (Fig. 7 A),根據觀察發 現仔魚具驅光性,利用此特性使用小型聚光燈收集仔魚,將仔魚移入培育槽中育
苗,仔魚會浮游於中上層。初孵化仔魚卵黃囊尚未消化殆盡,唯仔魚會在孵化後6
~ 8 小時內開始攝食,因此在孵化當日培育槽中即要添加藻水,並以 150 目的浮 游生物網,篩選出大小為 120 ~ 240 µm 之輪蟲及橈足類幼生,以作為仔魚之初期 餌料生物,輪蟲投餵密度為 5 ~ 10 隻/ ml,未順利攝食之仔魚會在孵化後 65 小時 後全數死亡。
孵化後第 1 日 (Fig. 7 B) 仔魚全長為 3.94 ± 0.14 mm,卵黃消失殆盡,軀幹 有些許黑色素細胞分佈。孵化後第 2 日 (Fig. 7 C) 仔魚全長為 4.04 ± 0.19 mm,
軀幹的黑色素細胞開始增加;仔魚主要分佈於培育槽下層。孵化後第 3 日 (Fig. 7 D) 仔魚全長 4.38 ± 0.31 mm,各鰭仍呈原鰭狀。孵化後第 4 日 (Fig. 7 E) 仔魚全 長 5.14 ± 0.19 mm,體表黑色素持續擴散並分佈至頭部;孵化後第 5 日 (Fig. 7 F) 仔魚全長 5.40 ± 0.21 mm,鰭膜內縮各鰭部開始分化,口徑為 680 ± 25.6 µm 可兼
投大型橈足類;孵化後第 6 日 (Fig. 7 G) 仔魚全長 5.72 ± 0.35 mm;孵化後第 7 日 (Fig. 7 H) 仔魚全長 5.98 ± 0.51 mm,體色開始出現淡橘紅色色素細胞,鰓蓋及 軀幹隱約出現白色橫帶,仔魚背鰭及臀鰭之鰭式已與成魚相似;孵化後第 8 日 (Fig. 7 I) 仔魚全長 6.14 ± 0.40 mm,軀幹白色條紋明顯,口徑達 1.06 ± 0.39 mm 可 兼投豐年蝦無節幼蟲,此時每日換水 1 / 10 以穩定水質;孵化後第 9 日 (Fig. 7 J) 及第 10 日 (Fig. 7 K) 仔魚全長為 6.71 ± 0.43 mm,頭部吻端、背鰭基部與腹部邊 緣開始出現淡橘紅色色素細胞,體側出現兩條白色橫帶;孵化後第 13 日 (Fig. 7 L) 仔魚全長 7.20 ± 0.50 mm,體表橘紅色色素持續擴散,頭部與軀幹白色橫帶形成;
孵化後第 15 日 (Fig. 7 M) 仔魚全長 8.24 ± 0.76 mm,可以兼餵投人工飼料;各 鰭的橘紅色色素持續累積,尾柄第三條白色橫帶出現。孵化後第 20 日 (Fig. 7 N) 仔魚全長已達 8.60 ± 1.04 mm,可以完全投予人工飼料;孵化後第 25 日仔魚 (Fig.
7 O) 全長 8.88 ± 0.64 mm;孵化後第 30 日 (Fig. 7 P) 全長 9.88 ± 0.97 mm,至此 體色斑紋,已完全與成魚一致。孵化後第 70 日全長 23.45 ± 2.53 mm 眶下骨硬棘 尚未出現 (Fig. 9 A),孵化後第 80 日仔魚全長 25.71 ± 2.58 mm 可觀察到頭部眶 下骨硬棘開始出現 (Fig. 9 B)。孵化後第 120 日仔魚全長達 32.95 ± 2.42 mm,眶 下骨硬棘更為明顯 (Fig. 9 C)。成熟種魚的眶下骨與眶前骨之硬棘延長並形成一對 長刺 (Fig. 9 D),此特徵為棘頰海葵魚分類上之主要依據。
四、形態發育
依據棘頰海葵魚在形態發育的特徵將初期生活史分為四個階段 (Table 3 及 Fig.
10) , 仔 稚 魚 日 齡 與 各 部 位 測 量 形 質 之 變 化 分 別 為 (1) 脊 索 末 端 上 屈 前 期 (Preflexion)、(2) 脊索末端上屈中期 (Flexion)、(3) 脊索末端上屈後期 (Postflexion) 及 (4) 稚魚期 (Juvenile)。孵化 0 ~ 6 日之前為脊索末端上屈前期,仔魚脊索尚未 上屈仍行浮游生活,初孵化仔魚測得各部位形質資料為全長為3.64 ± 0.31 mm,頭 長 0.94 ± 0.09 mm,眼徑 0.39 ± 0.04 mm,吻長 0.20 ± 0.03 mm,臀鰭前長 1.72 ± 0.12 mm 及體高為 0.39 ± 0.04 mm。至第 7 日全長為 5.98 ± 0.51 mm 進入脊索末 端上屈期,仔魚脊索明顯上屈,至第10 日全長為 6.71 ± 0.35 mm 後開始進入脊索 末端上屈後期,隨著脊索上屈且各鰭條陸續發育完成,仔魚游泳能力逐漸增強;
第 15 日全長為 8.24 ± 0.49 mm,體高與頭長的比例相等;第 31 日全長為 9.88 ± 1.48 mm 是初孵化全長的 2.71 倍,開始進入稚魚期。
仔魚的相對成長發育是以魚體各部位長度對全長求出相對關係比例算出,測 量標準體長、下頜長、體高、頭長、吻長、眼徑及臀鰭前長等七個部位形質長度 與全長比較如 Fig. 11 所示。棘頰海葵魚一般形質出現正相關成長有頭長、吻長及 臀鰭前長等三個部位,在全長約5 ~ 6 mm 時達到最大形質比例之後,約至 10 mm 時達到最小,之後形質變化趨於平緩。體高則於全長約10 mm 時達到最大比例之 後形質變化趨於緩慢增加。負相關增加的形質有標準體長、下頜長及眼徑等 3 個 部位,普遍在全長 3.64 ± 0.31 mm 時開始出現,至全長約 10 mm 時形質變化趨於 平緩。
Fig. 12 為棘頰海葵魚之仔魚開口口徑,在初孵化仔魚口徑為 470 ± 6.0 µm,
已可攝食 120 ~ 240 µm 的輪蟲及橈足類幼生等游動性較慢的初期餌料,以提供脊 索末端上屈前期的魚苗捕食,孵化第 5 日仔魚口徑為 0.68 ± 0.26 mm 可開始兼投 橈足類幼生,此時準備進入脊索末端上屈期,鰭條正在發育可提供成長較快仔魚 提早攝食;脊索末端上屈後期於第 8 日完成,仔魚口徑也達 1.06 ± 0.17 mm 可輕 易捕食豐年蝦無節幼蟲,因此第 10 日後不再給予橈足類,以豐年蝦無節幼蟲作 為主要餌料,第 20 日仔魚口徑為 2.93 ± 0.33 mm,可投餵微粒人工飼料開始馴餌 (粒徑 ≦ 1 mm)。
五、骨骼發育
棘頰海葵魚頭部、脊椎骨、背鰭、臀鰭及尾鰭的發育順序如 Fig. 13 所示,與 不同階段的骨骼發育圖如 Fig. 14 ~ 17 所示。
(一) 頭部骨骼
頭 部 骨 骼 主 要 紀 錄 上 頜 骨 (maxilla) 、 前 上 頜 骨 (premaxilla) 、 牙 齒 (teeth)、密克爾氏軟骨 (Meckel’s cartilage)、齒骨 (dentary)、方骨 (quadrate)、
角 舌 骨 (ceratohyal) 、 下 舌 骨 (hypohyal) 、 間 舌 骨 (interhyal) 、 鰓 蓋 條 (branchiostegals)、舌頷骨 (hyomandibular)、續接骨 (symplectic)、鰓蓋系列
(opercular series)、咽齒 (pharyngeal teeth)、梁軟骨 (trabecula)、篩骨板 (ethmoid plate)、主匙骨 (cleithrum) (Fig. 14、15)。其中密克爾氏軟骨為軟骨,方骨、
角舌骨、下舌骨、間舌骨、舌頷骨、續接骨、咽齒、小梁骨、篩骨板為軟骨 化骨 (cartilage bone) ,其他上頜骨、前上頜骨、齒、齒骨、鰓蓋條、鰓蓋系 列、主匙骨為膜骨 (membrane bone) (Fig. 13)。孵化後第 1 日的仔魚 (3.72 mm SL),下頜已有密克爾氏軟骨形成,上頜部分尚無骨骼形成但其後方已形成篩 骨板來支撐上頜,其餘可見構造均以軟骨形式存在 (Fig. 15 A)。孵化後第 2 與第3 日的仔魚 (3.74 mm SL 與 3.95 mm SL),可見前頜骨、頜骨、齒骨、
鰓蓋條和主匙骨開始骨化,方骨、角舌骨、下舌骨、間舌骨、舌頷骨、續接 骨、咽齒、小梁骨、篩骨板仍呈軟骨狀態 (Fig. 15 B 與 15 C);下頜的齒骨開 始骨化後會漸漸地包覆密克爾氏軟骨。孵化後第4 日的仔魚 (4.18 mm SL),
咽齒完全骨化 (Fig. 15 D)。孵化後第 5 ~ 6 日的仔魚 (4.29 mm SL),方骨、舌 頷骨和續接骨開始骨化 (Fig. 15 E 與 15 F)。孵化後第 7 ~ 8 日的仔魚 (4.65 mm SL 與 4.72 mm SL),上頜與下頜的牙齒以膜骨形式出現並骨化 (最初 2~3 個隨 成長增加) (Fig. 15 G 與 15 H)。孵化後第 9 日的仔魚 (5.11 mm SL),舌頷骨與 續接骨完全骨化 (Fig. 15 I)。孵化後第 10 日的仔魚 (5.14 mm SL),間舌骨與 篩骨板開始骨化 (Fig. 15 J)。孵化後第 13 日的仔魚 (5.76 mm SL),方骨、角 舌骨與下舌骨完全骨化 (Fig. 15 K)。孵化後第 15 日的仔魚 (5.91 mm SL),間 舌骨與鰓蓋系列完全骨化 (Fig. 15 L)。
(二) 脊椎 (Vertebral column)
紀錄脊椎骨中的椎體 (centra)、神經棘(弓) (neural processes)、血管棘(弓) (haemal processes)、椎體橫突 (parapophyses)、腹肋 (pleural rib) 的發育情形 (Fig. 16)。椎體與腹肋為膜骨,其餘為軟骨化骨 (Fig. 13)。初孵化仔魚椎體尚 未發育,體長 3.95 mm 時由前端開始發育出椎體,於體長 5.14 mm 時達定 數 (Fig. 16 F);脊索上屈 (notochord flexion) 開始於體長 4.36 mm。神經棘與 血管棘於體長 3.72 mm 開始形成 (Fig. 16 A),於體長 3.74 mm 達定數,同 時出現椎體橫突並於體長 4.18 mm 達定數 (Fig. 16 B)。
神經棘與血管棘約同時由前半段開始骨化 (3.95 與 4.18 mm SL) ,觀察至 第 32 日,體長 7.80 mm (Fig. 16 J) 進入稚魚期發現接近尾部的神經棘與血 管棘末端仍未完全骨化,但椎體橫突於體長4.18 mm 時開始骨化 (Fig. 16 B),
體長 4.36 mm 時完全骨化;腹肋於體長 5.14 mm 時開始出現,體長 5.65 mm 時開始骨化 (Fig. 16 G)。
(三) 尾鰭 (Caudal fin)
尾 鰭 部 分 紀 錄 下 尾 骨 (hypurals) 、 上 尾 骨 (epurals) 、 準 下 尾 骨 (parhypural)、尾神經骨 (uroneural)、主要鰭條 (principal caudal ray)、次要鰭 條 (secondary caudal ray) 及附屬軟骨 (accessory cartilage) (Fig. 17)。其中尾神 經骨、主要鰭條、次要鰭條為膜骨,其餘為軟骨化骨 (Fig. 13)。孵化後第 1 日仔魚已形成第一、第二下尾骨與準下尾骨 (3.72 mm SL) (Fig. 17 A),接著形 成第三及第四下尾骨 (3.95 mm SL) (Fig. 17 B、17 C),隨後形成第一至第三上 尾骨 (4.18 mm SL、4.29 mm SL) (Fig. 17 D、17 E),於體長 4.36 mm 時脊索 開始上屈且準下尾骨、第一及第二下尾骨靠近脊索端融合 (4.65 mm SL、5.11 mm SL) (Fig. 17 F、17 G),第三及第四下尾骨之後也融合 (5.14 mm SL、5.65 mm SL) (Fig. 17 H、17 I),於尾鰭下端開始出現附屬軟骨 (5.14 mm SL) (Fig. 17 H),接著尾鰭上端的附屬軟骨 (5.77 mm SL) (Fig. 17 J) 與尾鰭下端的附屬軟 骨 (6.38 mm SL) (Fig. 17 K) 也接著出現。
尾骨的骨化首先發生在第一、第二下尾骨及準下尾骨的融合體 (4.65 mm SL) (Fig. 17 F),尾神經骨於體長 5.77 mm 時出現並骨化,上尾骨是最後出現 骨化現象 (5.77 mm SL),觀察至第 32 日樣本 (7.80 mm SL) (Fig. 17 L),準下 尾骨、下尾骨及上尾骨皆仍未完全骨化。
初孵化仔魚具有僅具有原始的鰭褶 (fin fold),於體長 4.18 mm 時先由下 尾骨末端形成主要尾鰭條 (Fig. 17 D),體長 5.14 mm 時主要尾鰭條達定數共 16 條,上半段由第三及第四下尾骨末端形成的鰭條數為 8 條,下半段由準 下尾骨、第一及第二下尾骨末端形成的鰭條數為 8 條。次要尾鰭條較晚出現 (5.14 mm SL) (Fig. 17 H),於體長 5.91 mm 時達定數,上半段次要鰭條數為 8
條,下半段次要鰭條為 6 ~ 7 條。
(四) 背鰭 (Dorsal fin)
背鰭部分由前背骨 (predorsals)、近擔鰭骨 (proximal radials)、遠擔鰭骨 (distal radials)、硬棘 (hard spines)、鱗質鰭條 (lepidotrichia) 以及擔鰭骨突 (stay) 所組成 (Fig. 16)。其中硬棘及鱗質鰭條為膜骨,其餘為軟骨化骨 (Fig.
13)。背鰭最早出現的骨骼為近擔鰭骨 (4.29 mm SL) (Fig. 16 C),由中央往頭 部與尾部兩個方向形成,體長 5.11 mm 時達定數,同時出現遠擔鰭骨 (5.11 mm SL) (Fig. 16E),體長 5.41 mm 時達定數,觀察至第 32 日樣本 (7.80 mm SL) 仍未骨化完全 (Fig. 16 J)。擔鰭骨突於體長 5.76 mm 出現,之後與最後 一根近擔鰭骨融合 (5.91 mm SL)。前背骨於體長 5.14 mm 形成,並達定數。
近擔鰭骨 (5.76 mm SL) 與前背骨 (6.38 mm SL) 開始出現局部骨化現 像,但持續至第32 日後 (7.80 mm SL) 仍未骨化完全。鱗質鰭條與硬棘 (5.14 mm SL) 於發育過程中同時出現且兩者皆於體長 5.41 mm 時達定數。
(五) 臀鰭 (Anal fin)
臀鰭由近擔鰭骨 (Proximal radials)、遠擔鰭骨 (Distal radials)、硬棘 (Hard spines)、鱗質鰭條 (lepidotrichia)以及擔鰭骨突 (stay) 所組成 (Fig. 16)。除硬 棘及鱗質鰭條為膜骨外,其餘為軟骨化骨 (Fig. 13)。臀鰭最早出現的骨骼為 近擔鰭骨 (4.29 mm SL) (Fig. 16 C),由中央往頭部與尾部兩個方向形成 (Fig.
16 D),並於體長 5.11 mm 達定數 (Fig. 16 E)。遠擔鰭骨於體長 5.11 mm 形 成,並於體長 5.41 mm 達定數。擔鰭骨突於體長 5.41 mm 出現,於 5.65 mm 時與最末端近擔鰭骨融合 (Fig. 16 G)。
近擔鰭骨於體長 5.41 mm 開始骨化,與遠擔鰭骨觀察至第 32 日樣本 (7.80 mm SL) (Fig. 16 J) 仍未完全骨化。鱗質鰭條與硬棘於體長 5.14 mm 同 時出現並達定數 (Fig. 16 F)。
肆、討論
ㄧ、種魚培育
在解剖樣本中得知雌魚平均全長為 9.12 ± 1.46 cm (6.10 ~ 11.6 cm),平均體重 為 19.01 ± 9.77 g (8.10 ~ 44.17 g);雄魚平均全長為 7.57 ± 0.98 cm (5.6 ~ 9.3 cm),
平均體重為 10.38 ± 3.56 g (5.55 ~ 18.52 g),雌魚的體型明顯大於雄魚。雌性為海 葵魚群聚中第一優勢魚,體型最大,第二 (或加上第三) 為雄魚,其餘無性別分化,
雌魚失去時有依順序遞補變性的現象 (邵與陳,1990)。在眼斑海葵魚研究中卵巢 的產卵類型是屬於分批非同步型 (施,1994),卵巢中含有各種不同發育時期的卵 母細胞,這種類型魚類在一個延續較長的產卵時期中多次分批產卵,根據本研究 資料顯示棘頰海葵魚在水溫 25 ~ 30 ℃ 可終年不間斷產卵,何 (2007) 在眼斑海 葵魚也發現相同特性,故推測棘頰海葵魚也屬多次分批產卵的生殖型態。
棘頰海葵魚不具有性別兩色變異 (Sexually dichromatic),因此在配對繁殖時,
無法由體色來區分性別,強迫配對發現較強勢的魚 (可能是雌魚) 會驅趕或攻擊較 弱勢的魚 (可能是雄或雌魚),棘頰海葵魚生性好鬥,在仔稚魚群體中會有互相攻 擊而造成死亡情形,具地域優勢的大體型魚會攻擊體型較小的弱勢魚。本研究進 行人工配對時常發生互相攻擊致死情形,故配對過程需注意養殖密度及空間。何 (2007) 眼斑海葵魚研究中發現最佳的配對方式是大體型魚和體型較小之未成熟魚 來配對,配對缸中以放入一顆巨大異輻海葵,可減少攻擊行為發生及提高配對的 成功率。而應用於棘頰海葵魚種魚配對時依體型差異及配合海葵的置入也確實減 少了棘頰海葵魚的攻擊行為,提升了配對成功率。
二、胚胎發育
棘頰海葵魚受精卵偏動物極具有附著絲得固著於卵床上,形狀多呈長橢圓 形,剛產出之受精卵產卵數依雌魚體型大小可達 800 ~ 3000 粒不等,比較鞍斑海 葵魚受精卵數 1,400 ~ 2,000 粒 (陳等,2003)、粉紅海葵魚 300 ~ 700 粒 (何等,
2006)、白條海葵魚 800 ~ 2,500 粒 (錢,2006) 及眼斑海葵魚 300 ~ 1,000 粒 (何,
2007) 之最大產卵量來的多。造成產卵數差異的原因,除了不同魚種之間的差異
外,同種魚類之間體型較大、獲取較多養份以及相對成熟的個體而言,可預期其 產卵量也較多 (Hoff, 1996)。
魚的卵巢 (Ovaries) 及卵由於含有類胡蘿蔔素,因而有不同之色澤表現,不同 魚類其卵巢中類胡蘿蔔素含量在生殖細胞或其他細胞之變化相當大,視魚類所攝 食的食物種類而異 (王,1999)。棘頰海葵魚剛產出的受精卵顏色為橘色到橘紅色 之間,餵食配合飼料的種魚肉眼觀察產下的卵顏色多徧淡橘色,但投餵動物性餌 料卵色較呈橘紅色也對仔魚破卵後的活存率有正面的提升。Turano et al. (2000) 也 表示以新鮮魚肉、魷魚和蝦肉交替投餵種魚有助於提升卵質;但將生鮮餌料應用 於種魚培育,對棘頰海葵魚雖有較明顯提升仔稚魚破卵能力,但同時也發現容易 讓水質惡化,因此本研究將生鮮餌練製成軟性飼料後投餵,可以減少水質劣化之 結果。
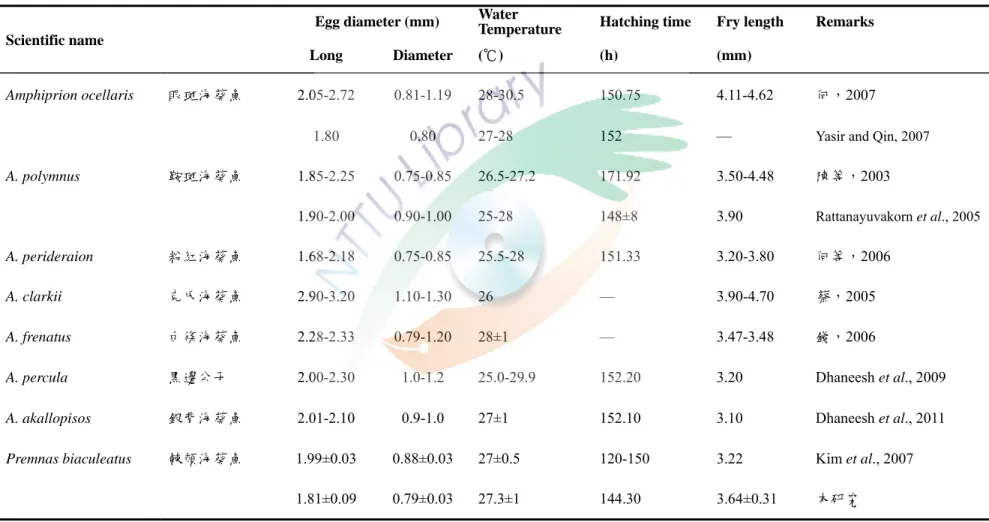
有些魚類雖屬同一種,但因產卵期或產卵場環境之異,卵徑也會發生差異 (朱,1997),本研究受精卵平均長、短徑為 1.81 ± 0.09 mm 及 0.79 ± 0.03 與韓國 棘頰海葵魚培育研究之受精卵的平均長、短徑為 1.99 ± 0.03 mm 及 0.88 ± 0.03 mm (Kim et al., 2007) 稍小。另與其他品種海葵魚比較,如鞍斑海葵魚平均長、短 徑 1.85 ~ 2.25 mm 及 0.75 ~ 0.85 mm (陳等,2003)、泰國鞍斑海葵魚平均長、短 徑 1.90 ~ 2.00 mm 及 0.90 ~ 1.00 mm (Rattanayuvakorn et al., 2005)、眼斑海葵魚平 均長、短徑為 2.05 ~ 2.72 mm 及 0.81 ~ 1.19 mm (何,2007)、克氏海葵魚平均長、
短徑為 2.90 ~ 3.20 mm及 1.10 ~ 1.30 mm (蔡,2005) 及白條海葵魚 2.28 ~ 2.33 mm及 0.79 ~ 1.20 mm (錢,2006)、黑邊公子 (A. percula) 平均長、短徑 2.00 ~ 2.30 mm 及 1.0 ~ 1.2 mm (Dhaneesh et al., 2009) 及銀背海葵魚 (A. akallopisos) 平均 長、短徑 2.01 ~ 2.10 mm 及 0.9 ~ 1.0 mm (Dhaneesh et al., 2011) 之受精卵卵徑來 的小。棘頰海葵魚的卵徑僅大於粉紅海葵魚平均長、短徑為 1.68 ~ 2.18 mm 及 0.75 ~ 0.85 mm (何等,2006),較近似於澳洲眼斑海葵魚平均長、短徑為 1.80 mm 及 0.80 mm (Yasir and Qin, 2007) (Table 4)。
另根據試驗結果發現,棘頰海葵魚產卵最適水溫在 25 ~ 30 ℃,受精卵在水溫 27.3 ± 1 ℃以上約需 144.5 小時孵化,比較 Kim et al. (2007) 在韓國培育的棘頰海
葵魚胚胎發育過程中發現,水溫在 27.0 ± 0.5 ℃孵化時間需 150 小時,於相近水 溫條件下比本研究孵化時間提早約 6 小時。另,眼斑海葵魚在水溫 28 ~ 30.5 ℃ 下約 150.75 小時孵化 (何,2007)、鞍斑海葵魚在水溫 26.5 ~ 27.2℃ 下約 171.92 小時孵化、黑邊公子在水溫 25.09 ~ 29.9 ℃下約 152.20 小時孵化 (Dhaneesh et al., 2009) 及銀背海葵魚在水溫 27 ± 1 ℃下約 52.10 孵化 (Dhaneesh et al., 2011) 皆 較本研究之孵化時間長,如 Table 4 所示。Falk-Petersen (2005) 指出胚胎發育的時 間受魚種與孵化水溫不同而有所差異,水溫愈高會加速胚胎發育速度,即胚胎發 育所需的時間短 (何,2007),棘頰海葵魚也發現相似情形,於冬季水溫降至 25 ± 1 ℃時,孵化時間會延長至 7 日孵化。
三、仔魚培育
魚類在生殖策略上主要可分為以量取勝 (r-selection) 及以質取勝 (k-selection) 兩種 (李,2006),多數產浮性卵的魚類,通常其卵徑小而卵量多,孵化期也較短,
由於缺少親魚的保護所以存活率較彽,屬於以量取勝的生殖策略,例如翻車魚可 產三億粒卵,是採重量不重質的卵海戰術來確保後代的存活機會;而產沉底性、
附著性卵或口孵的魚類,通常其卵徑較大而量少,孵化期也較長,常有各種護卵 或護幼的行為,屬於以質取勝的生殖策略,如雀鯛、慈鯛、鰕虎、鳚科魚類等 (邵 與陳,2004;朱,1997)。受精卵孵化天數愈長,可預期仔魚體內器官及消化道發 育愈完整,孵化後仔魚可以很快進行攝食,研究發現棘頰海葵魚仔魚孵化後 7 小 時鏡檢胃內容物,即可發現橈足類幼生等餌料生物;在眼斑海葵魚仔魚孵化後 8 小時也發現胃內有輪蟲存在 (何,2007);鞍斑海葵魚在孵化後 14 小時開始投餵 輪蟲,並在 4 小時後發現胃內也有輪蟲存在 (陳等,2003)。
有學者提出對於初次攝食的仔魚,應提供與自身卵黃囊成份相近的營養或餌 料生物,有利於仔魚的營養需求和代謝能力,以提高魚苗活存率 (Tocher and Sargent, 1984; Heming and Buddington, 1988),因此,考慮餌料對仔魚的適口性及營 養在初期餌料的選擇是相當重要的因素。棘頰海葵魚仔魚口徑為 470 ± 6.0 μm 之 間,比較眼斑海葵魚仔魚的口徑及口幅分別為 450 ~ 750 μm 及 450 ~ 500 μm (何,2007);鞍斑海葵魚仔魚的口徑及口幅為 459.6 ~ 777.8 μm 與 450 ~ 500 μm
(陳等,2003);粉紅海葵魚仔魚的口徑及口幅為 466 ~ 594 μm 與 375 ~ 500 μm (何 等,2006) 皆以提供殼長為 120 ~ 240 μm 的海水輪蟲或橈足類幼生作為初期餌 料。
研究中發現棘頰海葵魚仔魚同時投餵橈足類幼生或輪蟲後觀察胃內容物,捕 食橈足類幼生與輪蟲的比例約為 9:1,可見橈足類幼生具有韻律型的跳躍比輪蟲 緩慢移動的方式更能夠吸引棘頰海葵魚初孵化仔魚的注意進而捕食。棘頰海葵魚 育苗過程中發現,仔魚成長至第 5 日後可投放大型橈足類,而輪蟲及橈足類幼生 亦必需重疊投放,如此可提高仔魚的育成率,第 8 日可以兼投餵豐年蝦無節幼 蟲。在石斑魚及雀鯛的研究指出,育苗過程中餌料重疊的交替餵食可提高魚苗的 成長和存活 (Doi et al., 1997; Gopakumar and Santhosh, 2009)。
四、形態發育
仔稚魚培育最重要階段,就是初期攝食時期也就是魚苗由內因性營養 (卵黃囊) 轉變為外因性捕食的時期,仔魚的口徑大小與各攝食器官的發育狀況,直接影響 了可以攝取的餌料生物種類 (Olivotto et al., 2005),而初期階段捕食成功率及飢餓 是導致仔魚死亡的主要原因 (Bailey and Houde, 1989),因此攝食與運動系統的協調 必需同步發展 (Osse et al., 1997)。
初孵化的棘頰海葵魚各鰭部為圓鰭狀,仔魚鰭條因尚未分化游泳速度緩慢,
皆在水表層行浮游生活,開口口徑為 470 μm 因此初次攝食可捕食 120 ~ 240 μm 的輪蟲及小型橈足類幼生,符合張與謝 (1997) 研究海水魚仔魚的餌料粒徑大小,
最好為其開口口徑的三分之一至一半左右為佳的攝食口徑。孵化後第 5 日仔魚口 徑為 680 µm 可開始兼投大型投橈足類 110 ~ 500 µm,第 20 日齡仔魚口徑為 2.09 mm,可投餵微粒人工飼料 (粒徑≦1mm)。開口大小決定魚苗可以攝食的餌枓 生物,而棘頰海葵魚的口徑大小與餌料生物投餵序列與眼斑海葵魚 450 μm (何,
2007)、鞍斑海葵魚 459.6 μm (陳等,2003) 及粉紅海葵魚口徑 466 μm (何等,2006) 相近。
黃 (2009) 在產浮性卵的赤鰭笛鯛初期發育研究發現,赤鰭笛鯛在孵化後第 20
日前主要將能量用在攝餌相關形質發育上,至孵化後第30 日,則用在游泳相關形 質發育上,而產附著卵的棘頰海葵魚,由其各部位形質對全長成長比例的變化發 現,在孵化後到進入稚魚期之前,各部位形質發育變化明顯,配合骨骼發育的結 果可知,初期仔魚主要將能量運用在攝食相關的發育上,兩者結果皆顯示仔魚之 攝食機能發育必需優先於游泳機能,如此可確保仔稚魚在初期階段不會因飢餓及 營養不良而遭到淘汰。
Job and Bellwood (1996) 在棘頰海葵魚視覺發育的觀察發現,於水溫 21 ~ 24
℃下,沉降發生在 12 ~ 14 日,仔魚對於攝食輪蟲成功的機率,由孵化後 3 日的 96 % 提高到孵化後 10 日(沉降前期) 的 100 %,顯示仔魚在接近沈降期時對於 獵物的捕捉能力提高。仔魚的游泳能力會先以前進運動為主,再陸續完成轉向與 瞬間急速行進的操控運動,游泳機能的完備不僅可增加攝食成功率,也可逃避敵 害 (黃,2009),隨著攝食、視覺與游泳能力發育完備的程度,再配合適當的餌料 提供,有助於降低初期仔魚的耗損。
在赤鰭笛鯛初期發育之攝餌與游泳相關形質得知,在建立模式化的種苗生產 流程中,種苗生產最重要的關鍵在於充足的餌料,而仔魚初期餌料更直接影響仔 魚存活率,因此,生物餌料的選擇可針對仔魚不同成長階段與攝餌及游泳能力進 行考量與規劃,才能使仔稚魚攝餌成功率增加 以獲得最佳的飼育效果 (黃,2009)。
五、骨骼發育
由目前的研究已知,產浮性卵魚類的卵徑較小且孵化期較短,初孵化仔魚的 發育較不完全,例如鯛科的金頭鯛 Sparus aurata (Koumoundouros et al., 1997)、細 點牙鯛 Dentex dentex (Koumoundouros et al., 1999) 以及尖吻重牙鯛 Diplodus puntazzo (Sfakianakis et al., 2005) 的仔魚孵化後骨骼才開始發育。相較之下,產附 著性卵魚類的卵徑較大且孵化期長 (Green & McCormick, 2001),例如海葵魚亞科 的棘頰海葵魚 (本研究)、眼斑海葵魚 Amphiprion ocellaris 與白條海葵魚 A.
frenatus ( 蘇 , 2010) 、 雀 鯛 科 的 多 刺 棘 光 鰓 雀 鯛 Acanthochromis polyacanthus (Kavanagh and Alford, 2003) 及 蝦 虎 科 的 夜 棲 鋸 鱗 蝦 虎 魚 Priolepis nocturna (Wittenrich et al., 2007) 的初孵化仔魚發育較完整。
魚 類 攝 食 構 造 的 功 能 性 , 隨 著 仔 魚 從 孵 化 到 變 態 階 段 而 增 加 (Green &
McCormick, 2001; Turingan et al., 2005),而頭部骨骼是最先發育的部份,其負責攝 食及呼吸功能等主要要素 (Koumoundouros et al., 2000)。棘頰海葵魚於孵化後第 1 日即開始攝食,此時可觀察到篩骨板、密克爾氏軟骨及舌骨,密克爾氏軟骨則為 下頷的初始構造,篩骨板在初期攝食骨骼發育中扮演支撐上頷的角色,而舌骨則 有利於下頜活動 (Wittenrich et al., 2007),此時攝食骨皆以軟骨形式存在,因此僅 投以輪蟲作為初期餌料。孵化後第 2 日除了頜骨與齒骨之外,主匙骨也開始骨 化,其作用為支撐胸骨舌骨肌 (sternohyoideus),參與開口攝食的功能 (Matsuoka, 1987),另外咽齒骨的出現有助於仔魚的攝食。棘頰海葵魚孵化後第 2 與第 3 日 的仔魚,可見前頜骨、頜骨、齒骨、鰓蓋條和主匙骨陸續骨化,而方骨、角舌骨、
下舌骨、間舌骨、舌頷骨、續接骨、咽齒、小梁骨、篩骨板仍呈軟骨狀態;下頷 的齒骨開始骨化後會漸漸地包覆密克爾氏軟骨。第 4 日仔魚的咽齒骨化完全,研 究指出咽齒骨的骨化可用於順利捕捉獵物,其作用是為了輾碎和破壞獵物結構(甲 殼),以增加表面積來提升消化食物及養分吸收 (Hunt von Herbing et al., 1996;
Wainwright and Bellwood, 2002),在克氏海葵魚 Amphiprion clarkii 的研究也發現 在孵化後第 5 日咽齒骨骨化可順利捕食豐年蝦無節幼蟲 (Anto et al., 2009),隨著 棘頰海葵魚仔魚成長,口徑也會有所改變,孵化第 5 日後也開始兼投橈足類以供 捕食。孵化後第 7 ~ 8 日的仔魚,上頜與下頜的牙齒出現,使魚苗在攝食過程中 有助於捕捉獵物以防止逃脫,此時以提供較大型橈足類為主。孵化後第 13 ~ 15 日 的仔魚,鰓蓋骨陸續完成骨化,鰓蓋條與鰓蓋骨的骨化組成的鰓蓋骨系,主要功 能是保護鰓及形成鰓腔,使鰓的呼吸效率大為提高 (黃,1997)。
Kohno et al. (1996) 於金目鱸 Lates calcarifer 初期仔魚的骨骼發育研究提出,
仔魚於孵化後 40 小時開口時已具有初始的樑軟骨、鰓弓、舌骨、方骨、續接骨
-舌頜軟骨、上頜骨與密克爾氏軟骨等構造形成口腔,此時以「吸 (sucking feeding)」方式攝食,接著前頜骨與牙齒的出現,與現有構造的骨化與長度增加,
使原有的攝食機制增加 「抓 (grasping feeding)」 方式攝食。Aristazabal (2005) 在 赤鯛 Pagrus pagrus 的研究提出,初期仔魚於孵化後第 3 日開口,頭部初始骨骼構 造的長度增加與骨化,提高「吸」的攝食方式增加所攝食的輪蟲量,咽齒與牙齒
的出現有利於抓住食物,結合「吸」與「抓」的攝食方式,伴隨攝食與游泳相關 骨骼的骨化,有利於游泳能力的改善以提高餌料生物的捕食。
鰭的發育在魚類早期生活史是重要的特徵,且鰭的發育程度會影響仔魚的游 泳模式、速度、攝食能力與表現 (Kendall et al., 1984 ; Fukuhara, 1992)。隨著仔魚 成長,不同階段的骨骼發育與各部位鰭的形成會造成不同的運動能力,尾鰭由分 節的鰭條所構成,隨著發育會提升仔魚的推進、轉向能力 (Koumoundouros et al., 2001a; 邵與陳,2004)。尾鰭是棘頰海葵魚最早出現鰭條的部位,此與眼斑海葵魚 及白條海葵魚 (蘇,2010)、安邦雀鯛 Pomacentrus amboinensis (Murphy, 2007) 及 鯛科魚類 (Koumoundouros et al., 2001a, b; Sfakianakis et al., 2005) 的觀察相同。而 不同魚種為適應環境可能會有不同的鰭條發育順序,例如深棕連鰭鳚 Enchelyurus brunneolus (Watson, 1987) 以及雲紋裸皮鮋 Gymnapistes marmoratus (Neira, 1989) 則是胸鰭最先發育出鰭條,鰭除了有平衡和協助運動的作用外,為了適應棲地和 不同的生活方式,鰭會特化成不同的形狀或構造,以協助魚類進行攝食、呼吸、
生殖、爬行、飛翔、跳躍、吸附、發聲和防禦等作用 (Koumoundouros et al., 2001a;
邵與陳,2004)。
大多數魚類的骨骼是硬骨,其產生方式可分為兩種:ㄧ是由軟骨直接骨化而 來,這類骨骼稱為軟骨化骨 (cartilage bone) 或替代骨 (replacing bone);另一種方 式是在膜質 (結締組織) 上直接骨化而成,中間越過軟骨階段,所形成的硬骨稱為 膜骨 (membrane bone) (黃,1997)。在不同魚種之間,骨骼形成的方式與方向也會 有所差異,例如在大西洋笛鯛 (Potthoff et al., 1988)、鯛科魚類 (Koumoundouros et al., 2001a, b; Sfakianakis et al., 2005)、雀鯛科的眼斑海葵魚 (蘇,2010) 以及本研究 棘頰海葵魚的椎體皆以膜骨方式出現,由前端循著尾部方向骨化形成;而 Fraser et al. (2004) 在尖吻鱸的研究則觀察到椎體是以軟骨化骨的方式,從第七節附近開始 出現後再分別朝吻端及尾部方向陸續形成椎體。另外,棘頰海葵魚、眼斑海葵魚 (蘇,2010)、沙重牙鯛 Diplodus sargus (Koumoundouros et al., 2001b) 及尖吻重牙 鯛 Diplodus puntazzo (Sfakianakis et al., 2005) 的背鰭近擔鰭骨是先由靠近脊柱中 央的位置出現,再分別朝向吻端及尾部陸續形成;而Coban et al. (2009) 觀察到赤 鯛 Pagrus pagrus 的背鰭近擔鰭骨是由前端開始循著尾部方向陸續形成。
尾部骨骼的發育通常包含下尾骨之間的融合,下尾骨、準下尾骨與尾桿骨的 融合,以及尾神經骨的消失或融合 (Johnson, 1984)。棘頰海葵魚與眼斑海葵魚 (蘇,2010) 尾部同樣具有三根上尾骨,上尾骨達定數後不會像白條海葵魚 (蘇,
2010) 長出第四根上尾骨再消失的現象;但同樣觀察到第一、第二下尾骨以及準下 尾骨先出現融合,而後第三及第四下尾骨也融合的現象。其它如大西洋笛鯛 Lutjanus campechanus (Potthoff et al., 1988)、鯛科魚類 (Koumoundouros et al., 2001a, b) 以及紅甘鰺 Seriola dumerili (Liu, 2001) 同樣也觀察到下尾骨融合的現象。
Johnson (1981) 提出下尾骨的融合通常發生在快速游泳的魚類,其目的可能是為了 有更有效的游泳能力;在個體發育的轉換過程,骨骼的減少和融合,對於不同骨 骼的起源和可能的同源性提供了關鍵的價值 (Fritzsche and Johnson, 1980)。
幼魚尋找未來的棲息地的行為在魚類行為學上稱為「著苗行為」(Settlement behavior),此階段對海洋生物而言十分的重要,著苗時幼魚不僅要考慮該棲地的水 質、週遭地形還有天敵威脅等變因 (李,2006)。Green and McCormick (2001) 在三 色小丑 Amphiprion melanopus 的觀察發現,水溫 28 ± 1℃下,於孵化後第 3 日,
卵黃消耗的同時消化道也開始發育,接著下頜開始骨化,重要的構造正在形成與 發育,此急速結構性的變化可能是其發育的關鍵時期。於孵化後第 8 日,仔魚的 各部位的鰭完全骨化且轉變至成魚的體色外形,此時進入變態期正值沈降期 (settlement) 階段;鰭有助於機動性的在海葵與珊瑚礁附近活動,而顏色有助於偽 裝和辨識同種個體。蘇 (2010) 在眼斑海葵魚的觀察發現,水溫 27 ± 1 ℃下,於 孵化後第 10 日觀察到明顯硬骨化的前上頜骨、上頜骨與齒骨,此時正好是其生 活史由浮游期轉變為沉降期的階段,因此可推測攝食骨骼在這個時間點有明顯的 變化是為了因應生活史的轉變,沉降後食物種類更多元化,除了浮游性生物外還 增加了底棲性浮游動物以及藻類,所以需要更強而有力的攝食骨骼。而本研究的 棘頰海葵魚於水溫 24 ~ 28 ℃下,孵化後第 13 ~ 15 日發現仔魚各部位的鰭條大多 骨化完成且體色開始轉變,開始會聚集於水槽底部及角落,此時期開時進入沈降 期階段。可見棘頰海葵魚的沉降時機相較於眼斑海葵魚 (蘇,2010) 及白條海葵魚 (Green and McCormick, 2001) 來得晚。這些發育過程中的轉變有助於存活、成長和 分 化 , 除 了 解 不 同 物 種 的 生 活 模 式 , 也 突 顯 仔 魚 發 育 過 程 的 重 要 性
(Koumoundouros et al., 2001a)。
飼育環境與魚種差異等因素會間接影響發育速度,蘇 (2010) 在 27 ± 1 ℃下 觀察眼斑海葵魚及白條海葵魚骨骼發育結果與本研究在 27 ± 1 ℃觀察棘頰海葵魚 的各部位骨骼發育順序有相似之處。在頭骨部分,白條海葵魚比眼斑海葵魚發育 速度快,初孵化仔魚皆可觀察到篩骨板與密克爾氏軟骨;眼斑海葵魚孵化後第 7 日上下頜皆可看見牙齒,第 10 日明顯觀察到前上頜骨、上頜骨與齒骨的硬骨化,
白條海葵魚孵化後第 5 ~ 6 日於上下頜可看見牙齒,第 7 日明顯觀察到前上頜
骨、上頜骨、齒骨與主齒骨的硬骨化。而棘頰海葵魚觀察上發現孵化後第 1 日即
可發現篩骨板、密克爾氏軟骨與舌骨;而第 3 日可發現前上頜骨、上頜骨與齒骨 的硬骨化,明顯較眼斑及白條海葵魚來的早;而牙齒發育則於 7 ~ 8 日後可明顯 發現與眼斑海葵魚發育時機相當。其它各部位骨骼發育順序詳細比較如 Fig. 18 所 示:在脊椎骨部分,棘頰海葵魚除椎體形成的速度比眼斑海葵魚及白條海葵魚快;
神經棘與血管棘於孵化第 1 日皆已形成,但棘頰海葵魚骨化速度較兩者慢;而棘 頰海葵魚椎體橫突較兩者形成慢,但骨化速度較兩者快。在尾鰭部分,棘頰海葵 魚、眼斑海葵魚及白條海葵魚的準下尾、下尾骨於孵化第 1 日皆已形成,但棘頰 海葵魚骨化速度明顯較兩者慢;上尾骨、尾神經骨、主要鰭條與次要鰭條的形成 較兩者慢。在背鰭部分,白條海葵魚的前背骨形成較眼斑海葵魚及棘頰海葵魚要 快,但棘頰海葵魚的骨化速度較兩者慢;白條海葵魚及眼斑海葵魚在近擔鰭骨、
遠擔鰭骨、硬棘、鱗質鰭條與擔鰭骨突的形成速度皆較棘頰海葵魚要快。在臀鰭 部分,白條海葵魚及眼斑海葵魚在近擔鰭骨、遠擔鰭骨、硬棘、鱗質鰭條與擔鰭 骨突的形成速度比棘頰海葵魚皆要來的快。